Fine Scale Determinants of Soil Litter Fauna on a Mediterranean Mixed Oak Forest Invaded by the Exotic Soil-Borne Pathogen Phytophthora cinnamomi

 , ,
, ,
Abstract
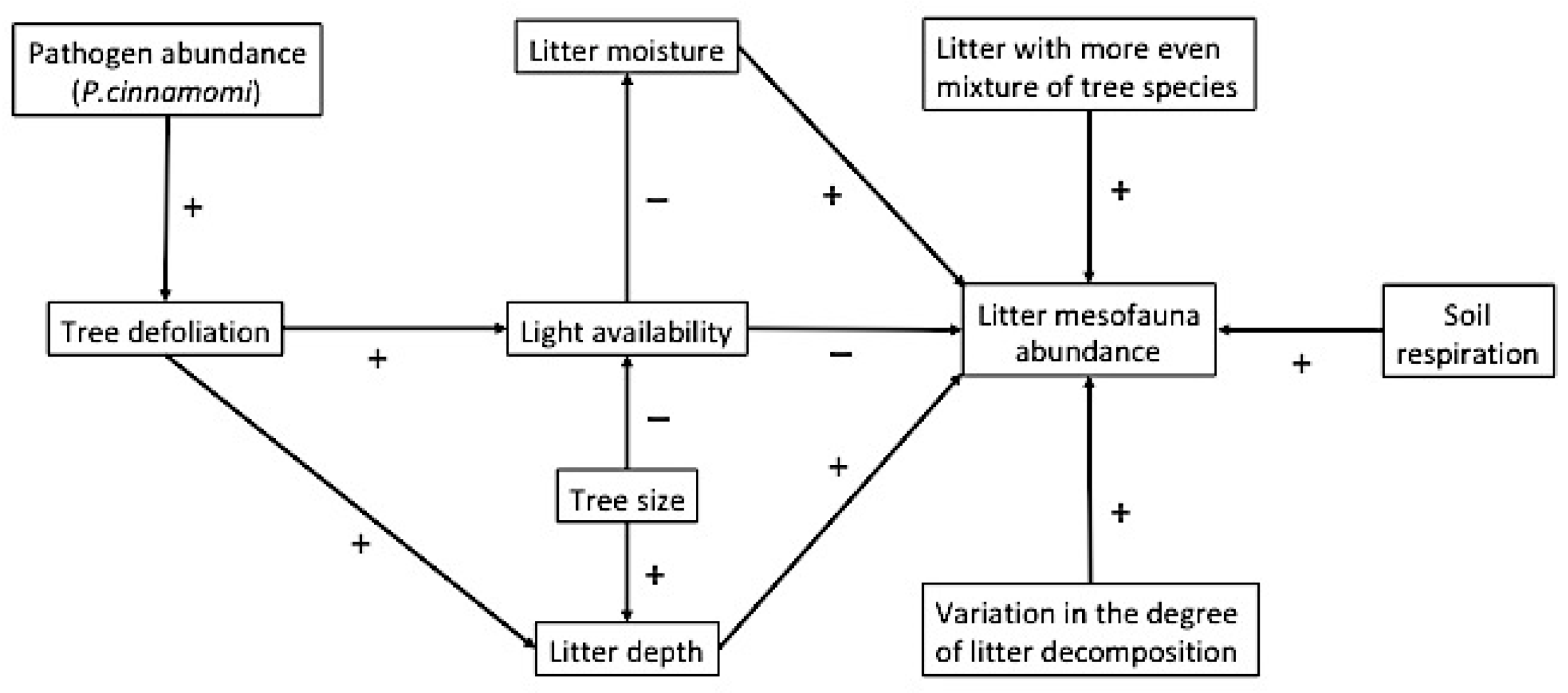
:1. Introduction
2. Material and Methods
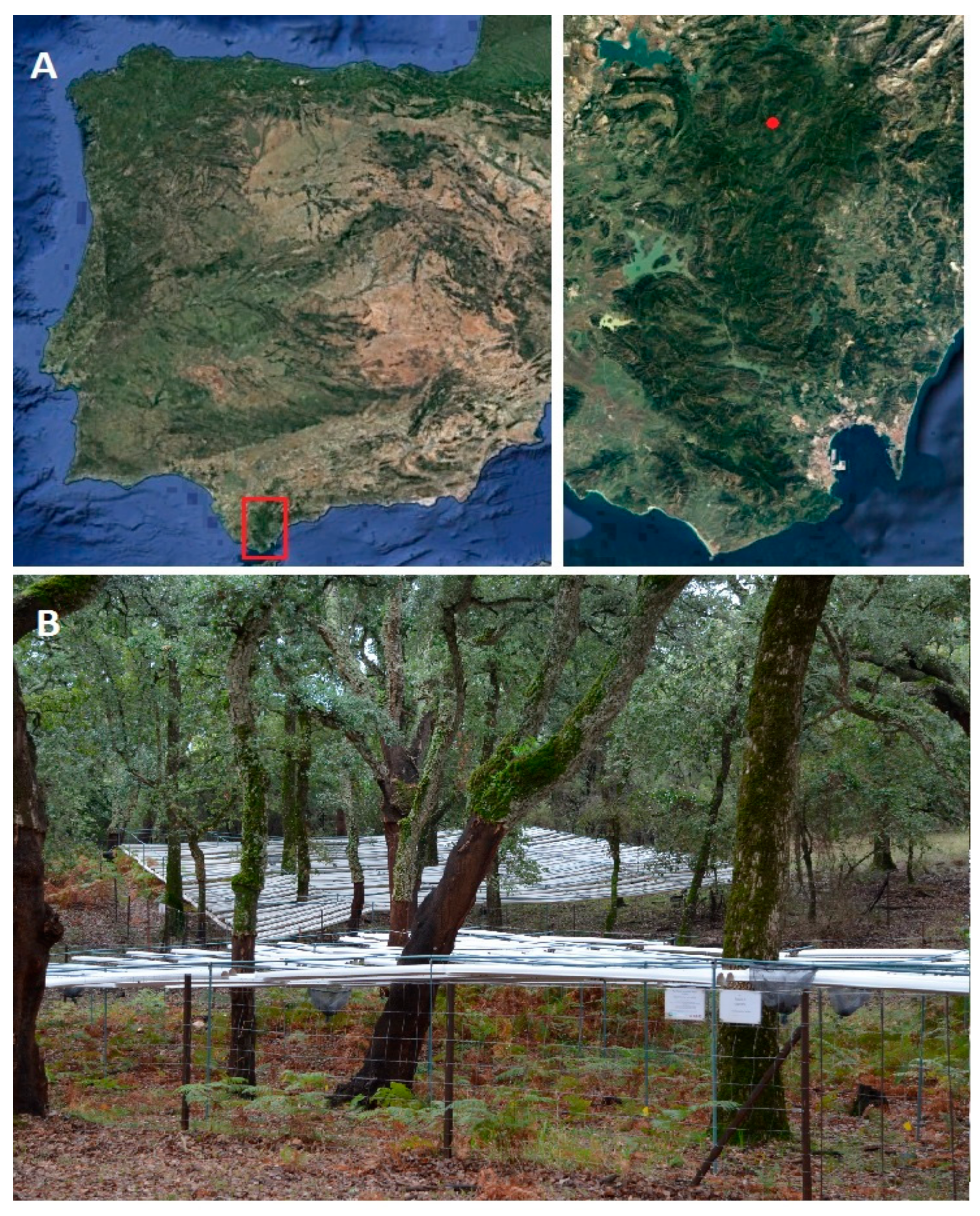
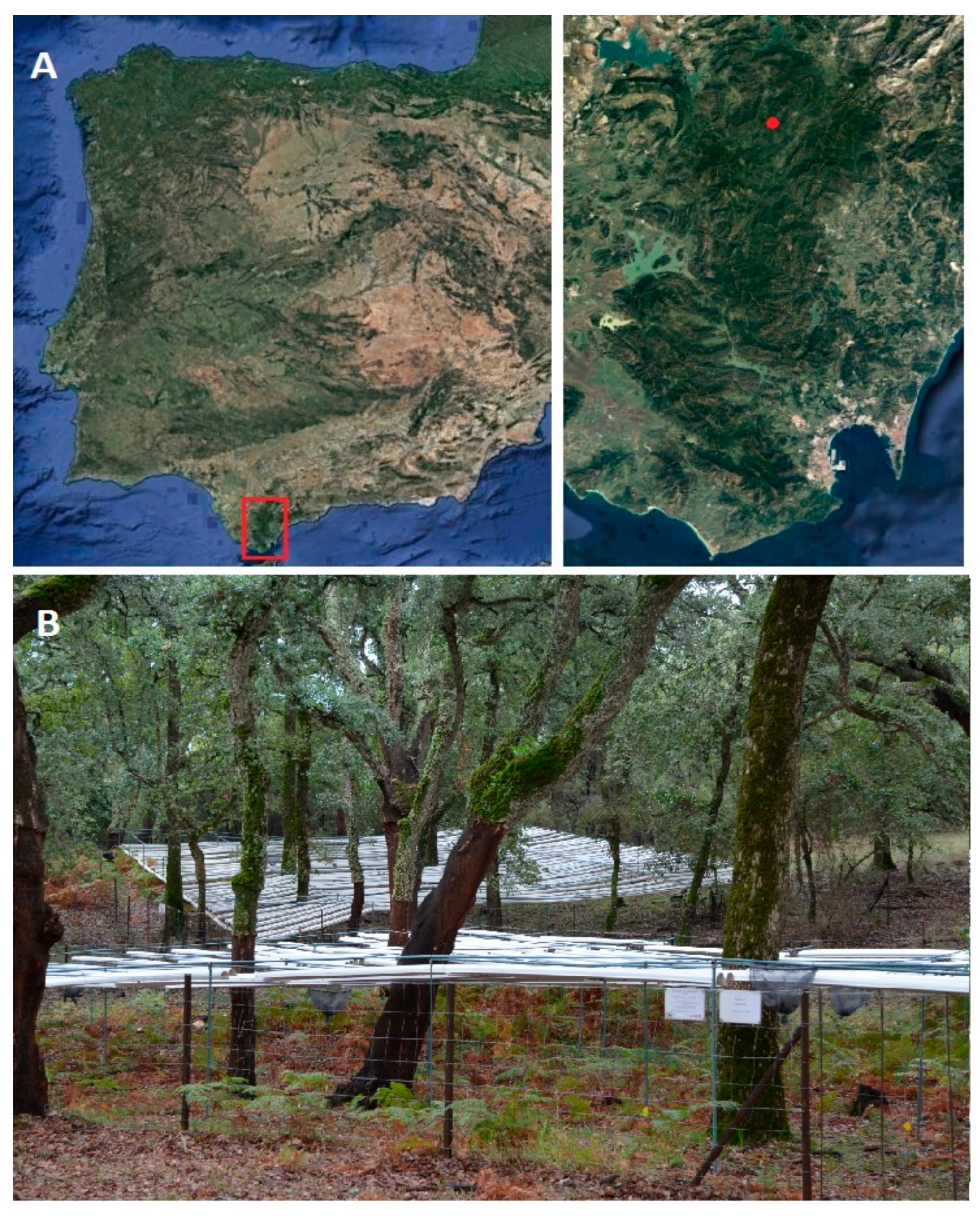
2.1. Study Site
2.2. Experimental Setup
2.3. Collection of Soil Fauna Invertebrates and Characterization of Litter Properties
2.4. Characterization of Fine-Scale Variation in Abiotic and Biotic Determinants of Soil Fauna under Field Conditions
2.5. Data Analysis
3. Results
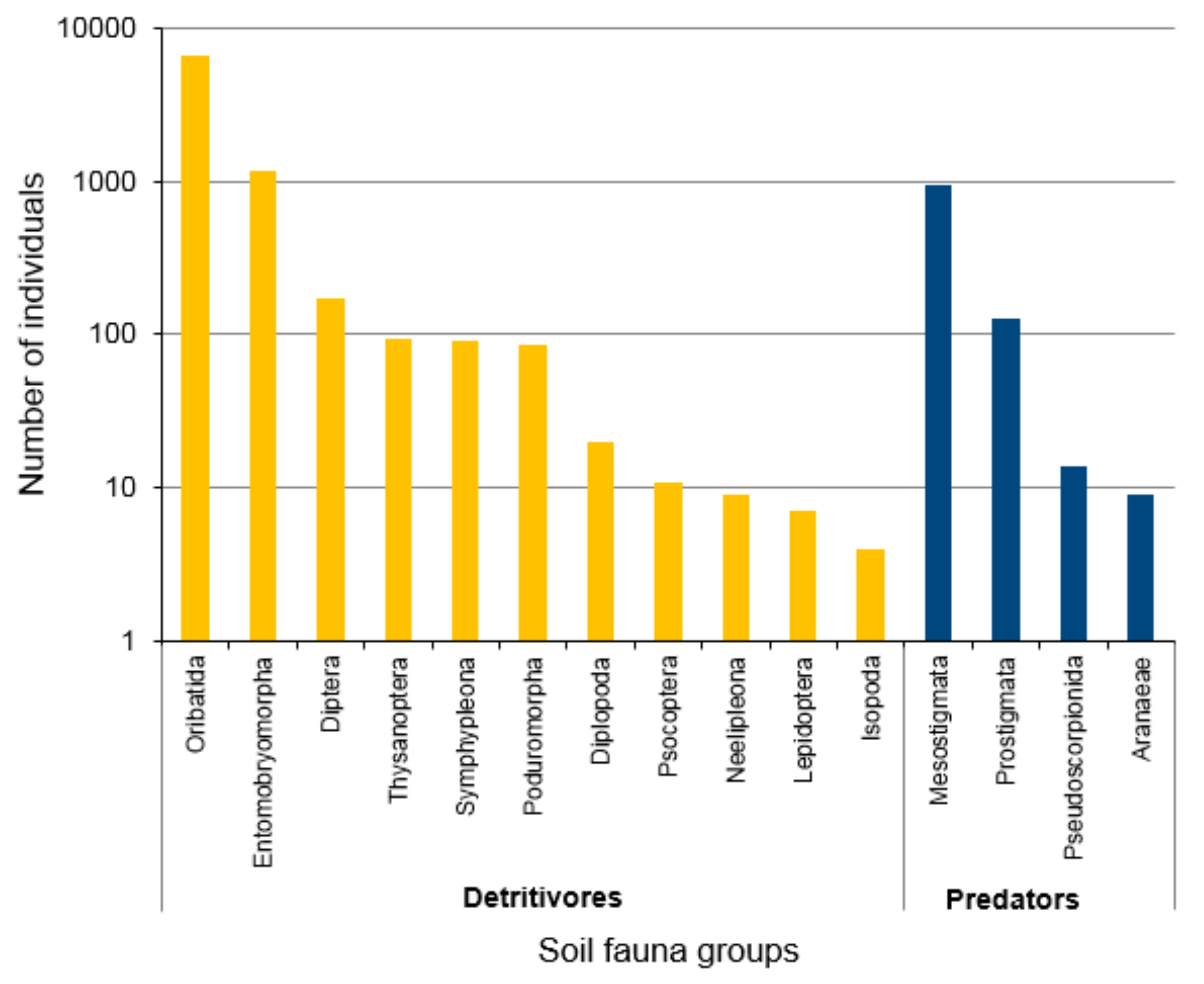
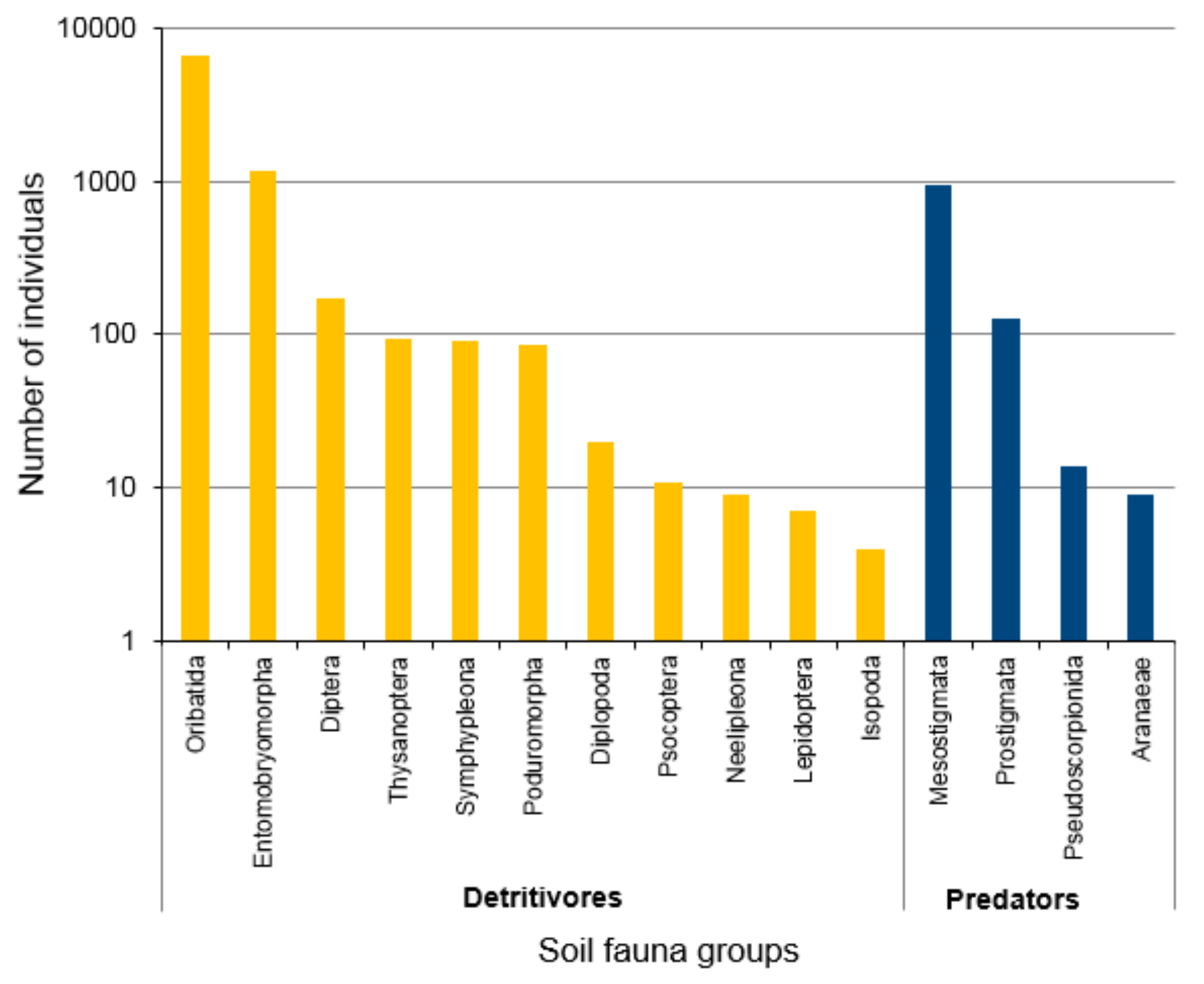
3.1. Abundance of Litter Mesofauna
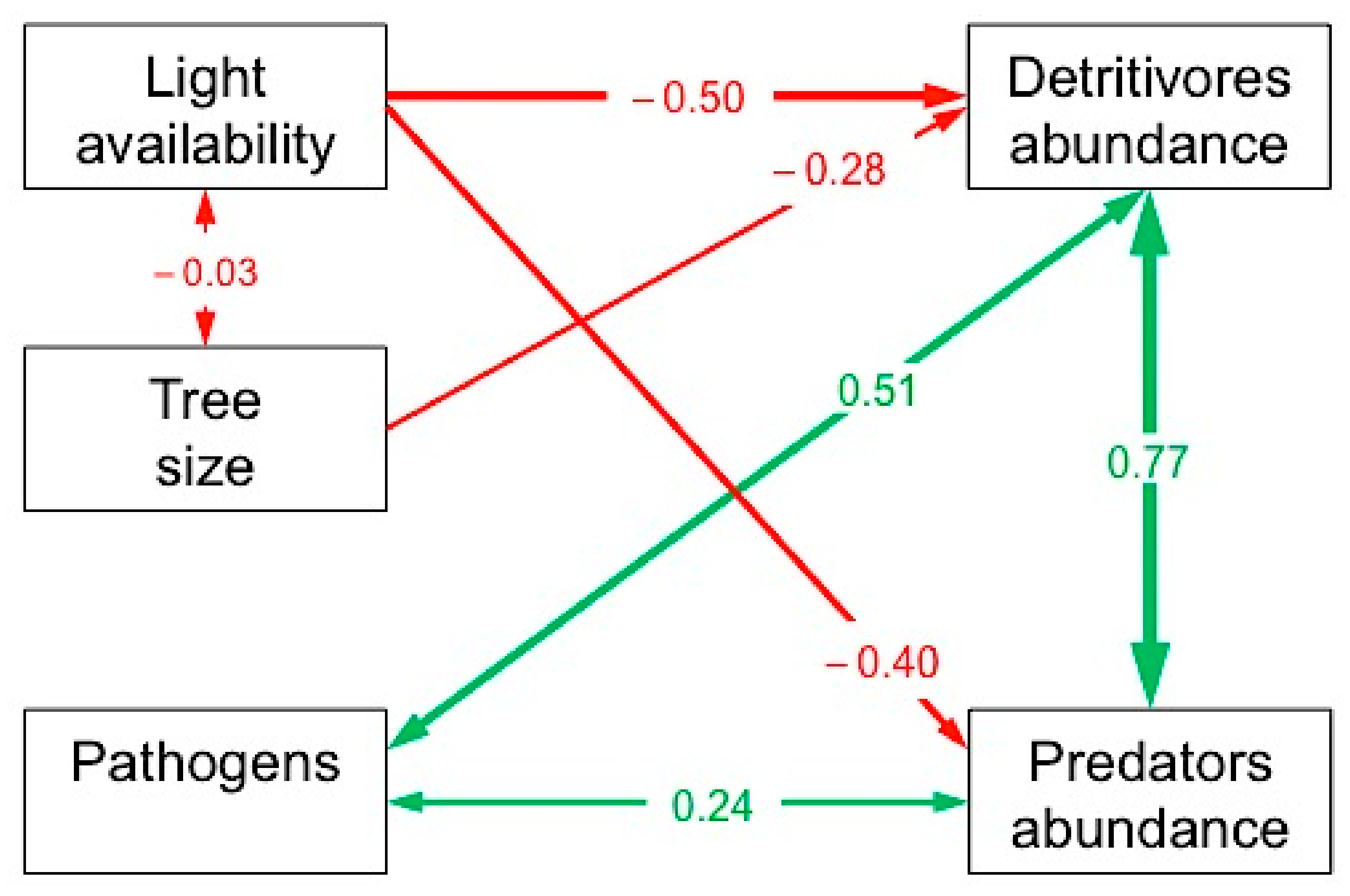
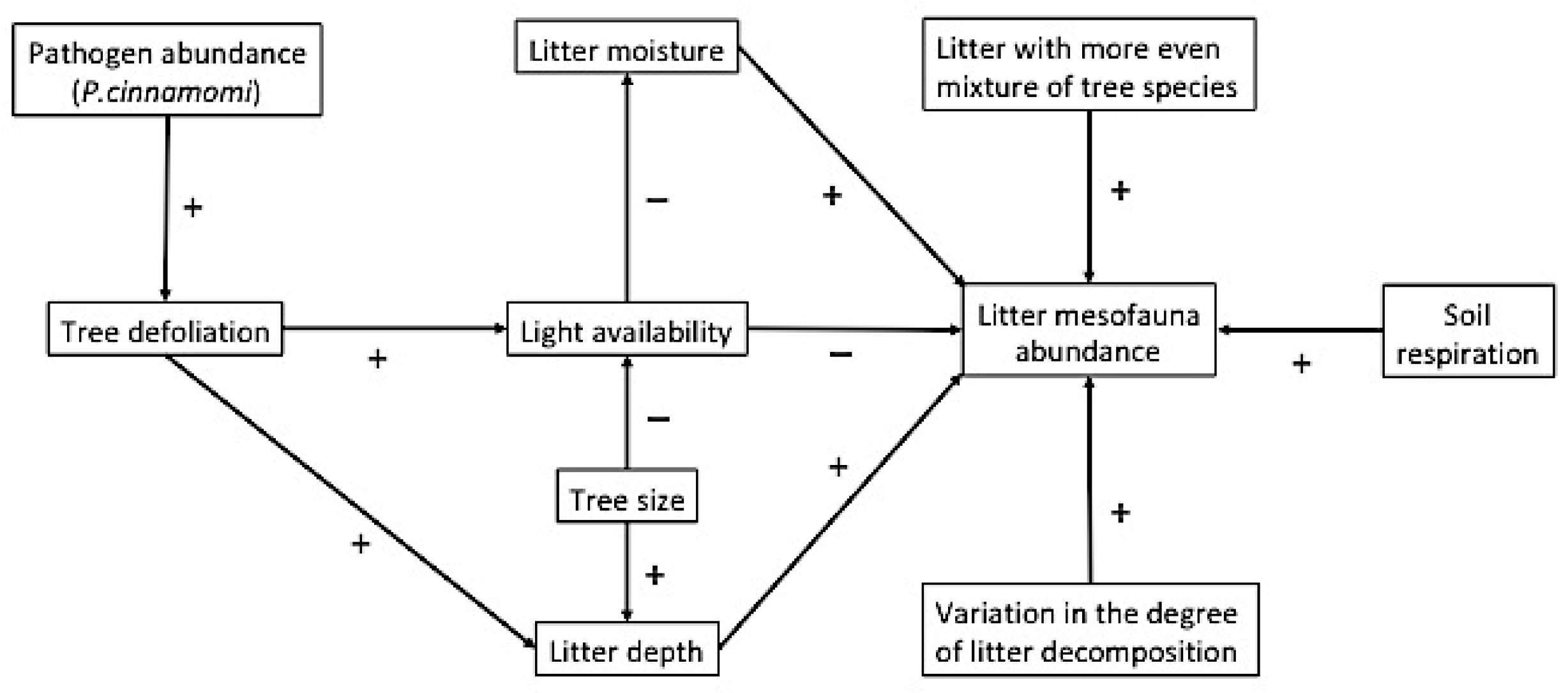
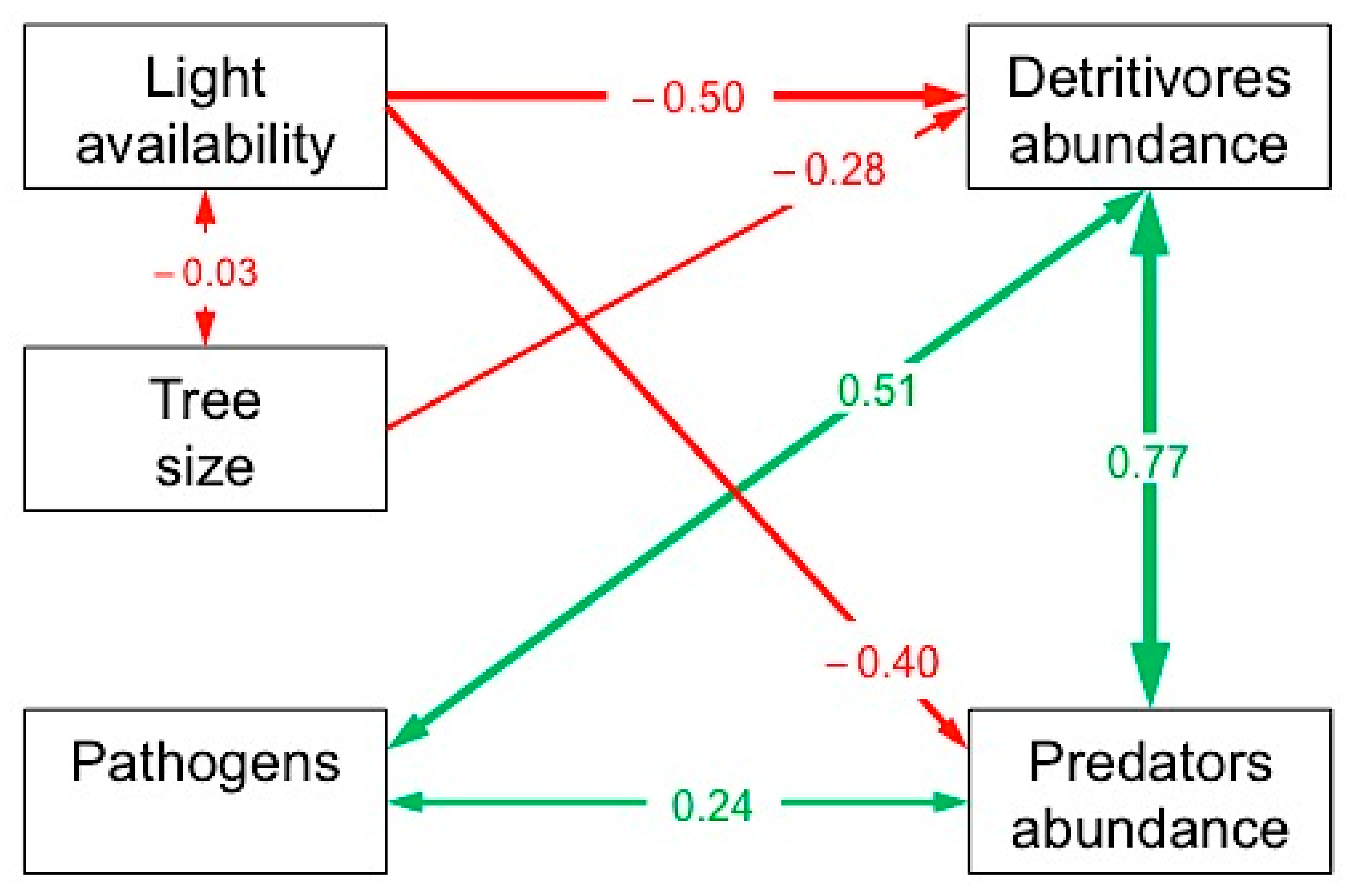
3.2. Direct and Indirect Determinants of the Abundance of Detritivore and Predatory Mesofauna
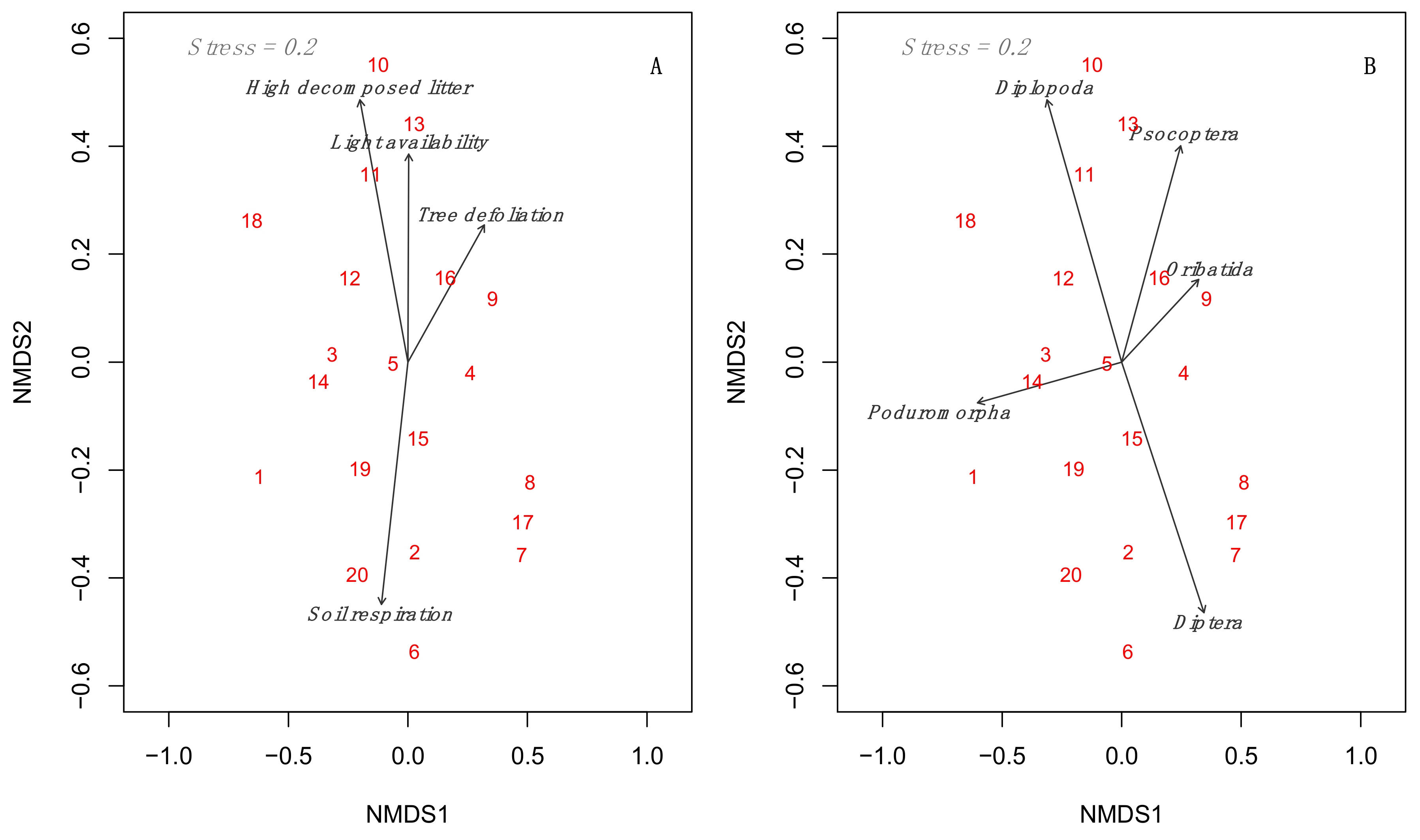
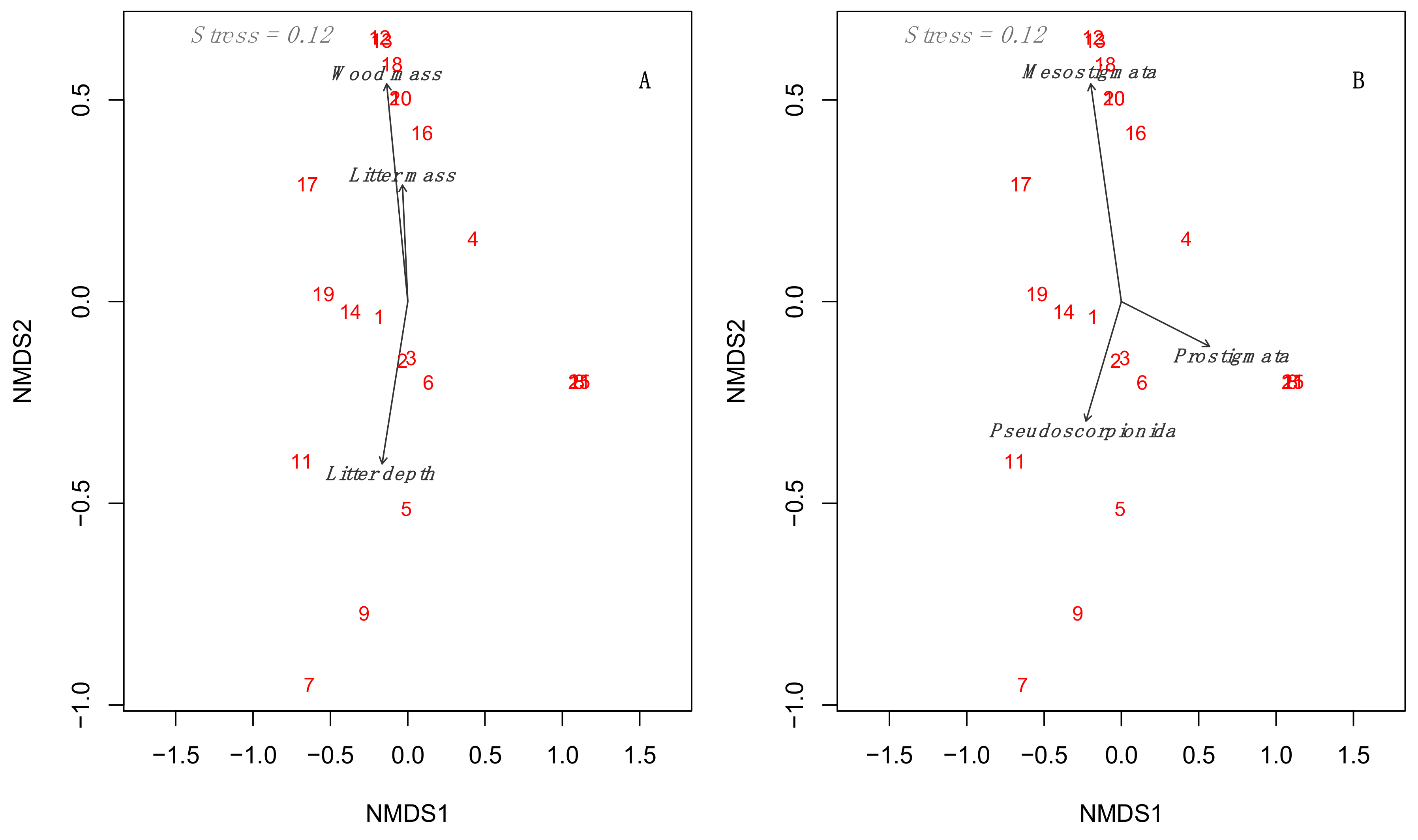
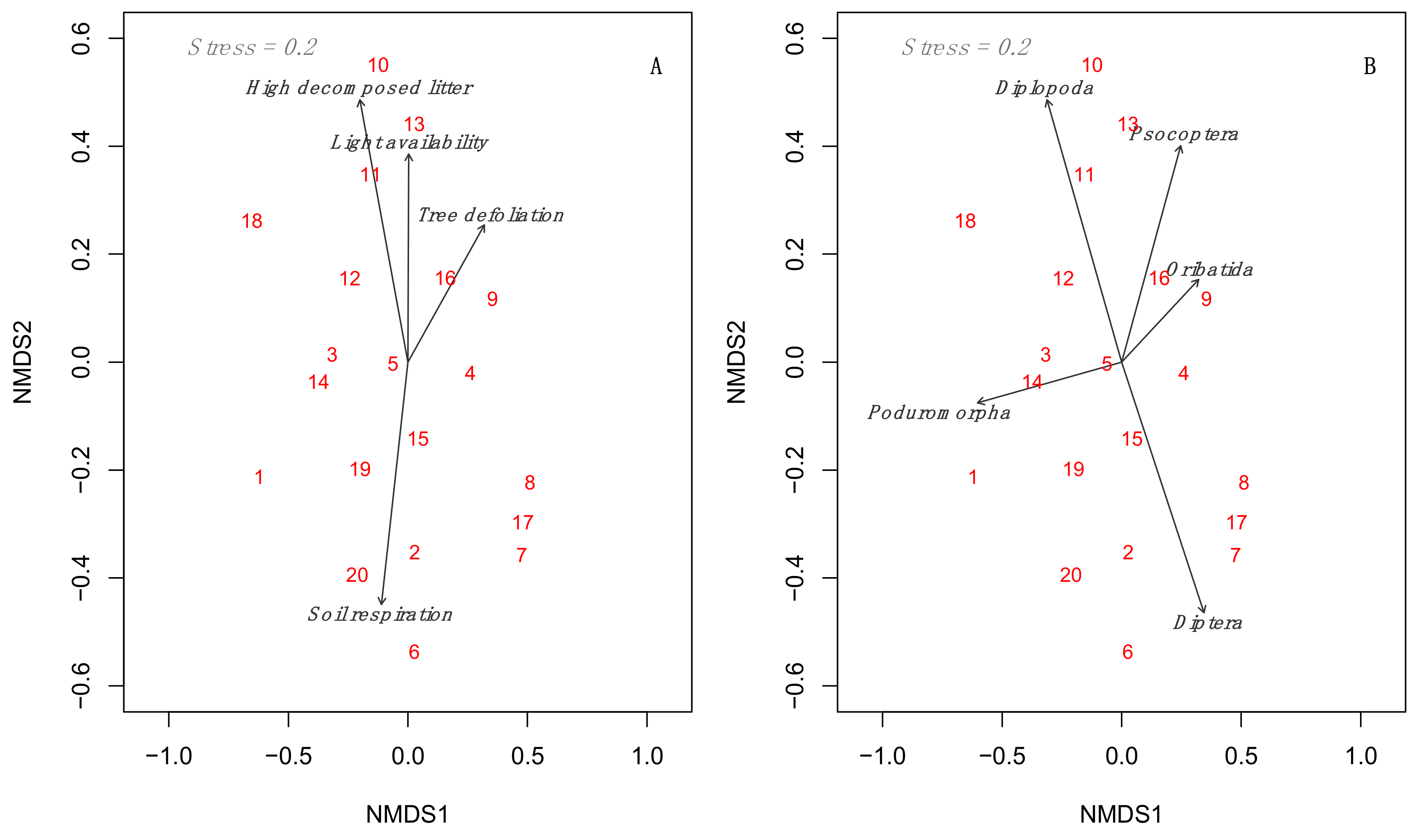
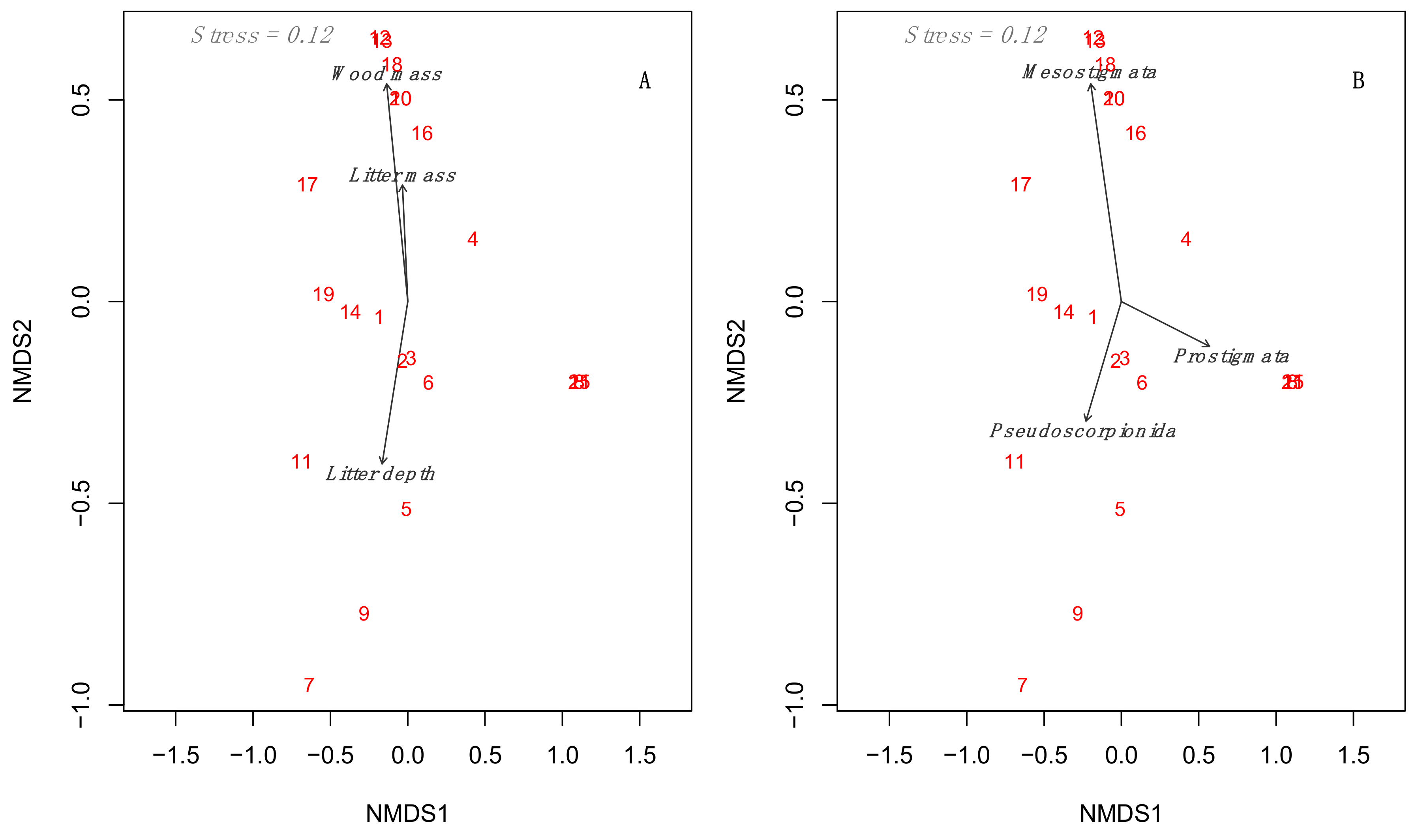
3.3. Multidimensional Analyses Relating to Environmental Predictors to Mesofauna Composition
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

References
- García-Palacios, P.; Maestre, F.T.; Kattge, J.; Wall, D.H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol. Lett. 2013, 16, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Soong, J.L.; Nielsen, U.N. The role of microarthropods in emerging models of soil organic matter. Soil Biol. Biochem. 2016, 102, 37–39. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkely, CA, USA, 1979; Volume 5. [Google Scholar]
- Aerts, R. The freezer defrosting: Global warming and litter decomposition rates in cold biomes. J. Ecol. 2006, 94, 713–724. [Google Scholar] [CrossRef]
- Cadisch, G.G. Driven by Natureplant Litter Quality and Decomposition; CAB International: Wallingford, UK, 1997. [Google Scholar]
- Eisenbeis, G. Biology of soil invertebrates. In Intestinal Microorganisms of Termites and Other Invertebrates; Springer: Berlin, Germany, 2006; pp. 3–53. [Google Scholar]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Wu, F.; Yang, W.; Xu, Z.; Zhang, L.; Liu, Y. Soil fauna significantly contributes to litter decomposition at low temperatures in the alpine/subalpine forests. Pol. J. Ecol. 2015, 63, 377–386. [Google Scholar] [CrossRef]
- Brussaard, L.; Pulleman, M.M.; Ouédraogo, É.; Mando, A.; Six, J. Soil fauna and soil function in the fabric of the food web. Pedobiologia 2007, 50, 447–462. [Google Scholar] [CrossRef]
- Pauli, N.; Abbott, L.; Negrete-Yankelevich, S.; Andrés, P. Farmers’ knowledge and use of soil fauna in agriculture: A worldwide review. Ecol. Soc. 2016, 21, 1–21. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, M.A. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Mueller, K.E.; Eisenhauer, N.; Reich, P.B.; Hobbie, S.E.; Chadwick, O.A.; Chorover, J.; Dobies, T.; Hale, C.M.; Jagodziński, A.M.; Kałucka, I. Light, earthworms, and soil resources as predictors of diversity of 10 soil invertebrate groups across monocultures of 14 tree species. Soil Biol. Biochem. 2016, 92, 184–198. [Google Scholar] [CrossRef]
- Santonja, M.; Fernandez, C.; Proffit, M.; Gers, C.; Gauquelin, T.; Reiter, I.M.; Cramer, W.; Baldy, V. Plant litter mixture partly mitigates the negative effects of extended drought on soil biota and litter decomposition in a mediterranean oak forest. J. Ecol. 2017, 105, 801–815. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Ibánez, B.; Serrano, M.S.; De Vita, P.; Avila, J.M.; Pérez-Ramos, I.M.; García, L.V.; Esperanza Sánchez, M.; Maranón, T. Spatial patterns of soil pathogens in declining mediterranean forests: Implications for tree species regeneration. New Phytol. 2012, 194, 1014–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valladares, F. Light heterogeneity and plants: From ecophysiology to species coexistence and biodiversity. In Progress in Botany; Springer: Berlin, Germany, 2003; pp. 439–471. [Google Scholar]
- Ma, Z.; Yang, W.; Wu, F.; Tan, B. Effects of light intensity on litter decomposition in a subtropical region. Ecosphere 2017, 8, e01770. [Google Scholar] [CrossRef]
- Pande, Y.; Berthet, P. Observations on the vertical distribution of soil oribatei in a woodland soil. Ecol. Entomol. 1975, 127, 259–275. [Google Scholar] [CrossRef]
- Boyd, I.; Freer-Smith, P.; Gilligan, C.; Godfray, H. The consequence of tree pests and diseases for ecosystem services. Science 2013, 342, 1235773. [Google Scholar] [CrossRef] [PubMed]
- Lovett, G.M.; Weiss, M.; Liebhold, A.M.; Holmes, T.P.; Leung, B.; Lambert, K.F.; Orwig, D.A.; Campbell, F.T.; Rosenthal, J.; McCullough, D.G.; et al. Nonnative forest insects and pathogens in the united states: Impacts and policy options. Ecol. Appl. 2016, 26, 1437–1455. [Google Scholar] [CrossRef] [PubMed]
- Preston, D.L.; Mischler, J.A.; Townsend, A.R.; Johnson, P.T. Disease ecology meets ecosystem science. Ecosystems 2016, 19, 737–748. [Google Scholar] [CrossRef]
- Desprez-Loustau, M.-L.; Robin, C.; Buée, M.; Courtecuisse, R.; Garbaye, J.; Suffert, F.; Sache, I.; Rizzo, D.M. The fungal dimension of biological invasions. Trends Ecol. Evol. 2007, 22, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Hansen, E.M. Phytophthora species emerging as pathogens of forest trees. Curr. For. Rep. 2015, 1, 16–24. [Google Scholar] [CrossRef]
- Cobb, R.C.; Rizzo, D.M. Litter chemistry, community shift, and non-additive effects drive litter decomposition changes following invasion by a generalist pathogen. Ecosystems 2016, 19, 1478–1490. [Google Scholar] [CrossRef]
- Ruess, R.W.; McFarland, J.M.; Trummer, L.M.; Rohrs-Richey, J.K. Disease-mediated declines in N-fixation inputs by alnus tenuifolia to early-successional floodplains in interior and south-central alaska. Ecosystems 2009, 12, 489–502. [Google Scholar] [CrossRef]
- Lovett, G.M.; Arthur, M.A.; Weathers, K.C.; Griffin, J.M. Long-term changes in forest carbon and nitrogen cycling caused by an introduced pest/pathogen complex. Ecosystems 2010, 13, 1188–1200. [Google Scholar] [CrossRef]
- Cobb, R.C.; Eviner, V.T.; Rizzo, D.M. Mortality and community changes drive sudden oak death impacts on litterfall and soil nitrogen cycling. New Phytol. 2013, 200, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M. Phytophthora cinnamomi and oak decline in southern europe. Environmental constraints including climate change. In Annales des Sciences Forestieres; EDP Sciences: Ray Ulysse, France, 1996; pp. 347–358. [Google Scholar]
- Sánchez, M.; Caetano, P.; Ferraz, J.; Trapero, A. Phytophthora disease of quercus ilex in south-western Spain. For. Pathol. 2002, 32, 5–18. [Google Scholar] [CrossRef]
- Sánchez, M.; Caetano, P.; Romero, M.; Navarro, R.; Trapero, A. Phytophthora root rot as the main factor of oak decline in southern Spain. In Progress in Research on Phytophthora Diseases of Forest Trees; Forest Research: Farnham, UK, 2006; pp. 149–154. [Google Scholar]
- Romero, M.; Sánchez, J.; Jiménez, J.; Belbahri, L.; Trapero, A.; Lefort, F.; Sánchez, M. New pythium taxa causing root rot on mediterranean quercus species in South-west Spain and Portugal. J. Phytopathol. 2007, 155, 289–295. [Google Scholar] [CrossRef]
- Zentmyer, G.A. Phytophthora Cinnamomi and the Diseases It Causes; American Phytopathological Society: Saint Paul, MN, USA, 1980. [Google Scholar]
- Zentmyer, G. Origin and distribution of four species of Phytophthora. Trans. Br. Mycol. Soc. 1988, 91, 367–378. [Google Scholar] [CrossRef]
- Da Clara, M.I.; de Almeida Ribeiro, N.M. Decline of Mediterranean oak trees and its association with Phytophthora cinnamomi: A review. Eur. J. For. Res. 2013, 132, 411–432. [Google Scholar]
- Weste, G.; Marks, G.C. The biology of Phytophthora cinnamomi in Australasian forests. Annu. Rev. Phytopathol. 1987, 25, 207–229. [Google Scholar] [CrossRef]
- Shearer, B.; Crane, C.; Barrett, S.; Cochrane, A. Phytophthora cinnamomi invasion, a major threatening process to conservation of flora diversity in the South-west botanical province of western Australia. Aust. J. Bot. 2007, 55, 225–238. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Biodiversity hotspots in the Mediterranean basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Stocker, T. Climate change 2013: The Physical Science Basis: Working Group i Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Limousin, J.-M.; Rambal, S.; Ourcival, J.-M.; Joffre, R. Modelling rainfall interception in a Mediterranean quercus ilex ecosystem: Lesson from a throughfall exclusion experiment. J. Hydrol. 2008, 357, 57–66. [Google Scholar] [CrossRef]
- Berlese, A. Apparecchio per Raccogliere Presto ed in Gran Numero piccoli Arthropodi. J. Hydrol. 1905, 2, 85–89. [Google Scholar]
- Barrientos, J.A. Curso Práctico de Entomología; Universitat Autònoma de Barcelona: Bellaterra, Spain, 2004; Volume 41. [Google Scholar]
- Gullan, P.J.; Cranston, P.S. The Insects: An Outline of Entomology; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Scoble, M.J. The Lepidoptera. Form, Function and Diversity; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Alatalo, J.M.; Jägerbrand, A.K.; Juhanson, J.; Michelsen, A.; Ľuptáčik, P. Impacts of twenty years of experimental warming on soil carbon, nitrogen, moisture and soil mites across alpine/subarctic tundra communities. Sci. Rep. 2017, 7, 44489. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Guzmán, B. Canopy structure and spatial heterogeneity of understory light in an abandoned holm oak woodland. Ann. For. Sci. 2006, 63, 749–761. [Google Scholar] [CrossRef]
- Bekku, Y.; Koizumi, H.; Nakadai, T.; Iwaki, H. Measurement of soil respiration using closed chamber method: An IRGA technique. Ecol. Res. 1995, 10, 369–373. [Google Scholar] [CrossRef]
- Rosseel, Y. Lavaan: An R Package for Structural Equation Modeling and More Version 0.4-9 (Beta); Ghent University: Ghent, Belgium, 2011. [Google Scholar]
- Kline, R.B. Principles and Practice of Structural Equation Modeling; Guilford Publications: New York, NY, USA, 2015. [Google Scholar]
- Grace, J.B. Structural Equation Modeling and Natural Systems; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package “vegan”. Community Ecology Package, Version 2.5-1; 2013. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 February 2018).
- Clarke, K.; Ainsworth, M. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- George, P.B.; Keith, A.M.; Creer, S.; Barrett, G.L.; Lebron, I.; Emmett, B.A.; Robinson, D.A.; Jones, D.L. Evaluation of mesofauna communities as soil quality indicators in a national-level monitoring programme. Soil Biol. Biochem. 2017, 115, 537–546. [Google Scholar] [CrossRef]
- Lindberg, N.; Engtsson, J.B.; Persson, T. Effects of experimental irrigation and drought on the composition and diversity of soil fauna in a coniferous stand. J. Appl. Ecol. 2002, 39, 924–936. [Google Scholar] [CrossRef]
- O’Neill, K.P.; Godwin, H.W.; Jiménez-Esquilín, A.E.; Battigelli, J.P. Reducing the dimensionality of soil microinvertebrate community datasets using indicator species analysis: Implications for ecosystem monitoring and soil management. Soil Biol. Biochem. 2010, 42, 145–154. [Google Scholar] [CrossRef]
- Gerlach, J.; Samways, M.; Pryke, J. Terrestrial invertebrates as bioindicators: An overview of available taxonomic groups. J. Insect Conserv. 2013, 17, 831–850. [Google Scholar] [CrossRef]
- Ritter, E.; Dalsgaard, L.; Einhorn, K.S. Light, temperature and soil moisture regimes following gap formation in a semi-natural beech-dominated forest in Denmark. For. Ecol. Manag. 2005, 206, 15–33. [Google Scholar] [CrossRef]
- Prévost, M.; Raymond, P. Effect of gap size, aspect and slope on available light and soil temperature after patch-selection cutting in yellow birch–conifer stands, Quebec, Canada. For. Ecol. Manag. 2012, 274, 210–221. [Google Scholar] [CrossRef]
- Dhooria, M.S. Fundamentals of Applied Acarology; Springer: Berlin, Germany, 2016. [Google Scholar]
- Hansen, R.A.; Coleman, D.C. Litter complexity and composition are determinants of the diversity and species composition of oribatid mites (Acari: Oribatida) in litterbags. Appl. Soil Ecol. 1998, 9, 17–23. [Google Scholar] [CrossRef]
- Anderson, J.; Ineson, P.; Huish, S. Nitrogen and cation mobilization by soil fauna feeding on leaf litter and soil organic matter from deciduous woodlands. Soil Biol. Biochem. 1983, 15, 463–467. [Google Scholar] [CrossRef]
- Shearer, B. Time course studies of temperature and soil depth mediated sporangium production by phytophthora cinnamomi. Australas. Plant Pathol. 2014, 43, 235–244. [Google Scholar] [CrossRef]
- Birkhofer, K.; Schöning, I.; Alt, F.; Herold, N.; Klarner, B.; Maraun, M.; Marhan, S.; Oelmann, Y.; Wubet, T.; Yurkov, A. General relationships between abiotic soil properties and soil biota across spatial scales and different land-use types. PLoS ONE 2012, 7, e43292. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Units | Mean ± Standard Error | |
|---|---|---|---|
| Abiotic | Light availability | Glare Spread Function values (GSF) | 0.336 ± 0.016 |
| Soil humidity | % volumetric water content | 20.618 ± 1.022 | |
| Wood | Grams (g) | 13.997 ± 1.682 | |
| Litter depth | Centimeters (cm) | 4.268 ± 0.235 | |
| Litter moisture | % water in mass | 36.586 ± 1.956 | |
| Biotic | Soil respiration | µmol CO2 m−2 s−1 | 5.685 ± 0.358 |
| Tree size | Diameter at breast height (DBH) in cm | 37.455 ± 3.624 | |
| Tree defoliation | From 0 (no defoliation) to 3 (full defoliation) | 0.636 ± 0.151 | |
| Litter Q. canariensis | Grams (g) | 9.724 ± 0.930 | |
| Litter Q. suber | Grams (g) | 7.941 ± 0.683 | |
| Litter other species | Grams (g) | 0.088 ± 0.086 | |
| Litter nonidentifiable | Grams (g) | 41.355 ± 5.299 | |
| Decomposition level 1 | Grams (g), litter low decomposed | 11.028 ± 1.084 | |
| Decomposition level 2 | Grams (g), litter medium decomposed | 6.725 ± 0.601 | |
| Decomposition level 3 | Grams (g), litter highly decomposed | 41.355 ± 10.270 | |
| Pathogen (Phytophthora cinnamomi) | Colony Forming Units/Grams (CFU g−1) | 105.636 ± 23.707 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Chacón, A.; Homet, P.; Matías, L.; Gómez-Aparicio, L.; Godoy, O. Fine Scale Determinants of Soil Litter Fauna on a Mediterranean Mixed Oak Forest Invaded by the Exotic Soil-Borne Pathogen Phytophthora cinnamomi. Forests 2018, 9, 218. https://doi.org/10.3390/f9040218
Jiménez-Chacón A, Homet P, Matías L, Gómez-Aparicio L, Godoy O. Fine Scale Determinants of Soil Litter Fauna on a Mediterranean Mixed Oak Forest Invaded by the Exotic Soil-Borne Pathogen Phytophthora cinnamomi. Forests. 2018; 9(4):218. https://doi.org/10.3390/f9040218
Chicago/Turabian StyleJiménez-Chacón, Alejandro, Pablo Homet, Luis Matías, Lorena Gómez-Aparicio, and Oscar Godoy. 2018. "Fine Scale Determinants of Soil Litter Fauna on a Mediterranean Mixed Oak Forest Invaded by the Exotic Soil-Borne Pathogen Phytophthora cinnamomi" Forests 9, no. 4: 218. https://doi.org/10.3390/f9040218