Day Extension with Far-Red Light Enhances Growth of Subalpine Fir (Abies lasiocarpa (Hooker) Nuttall) Seedlings

 and
and 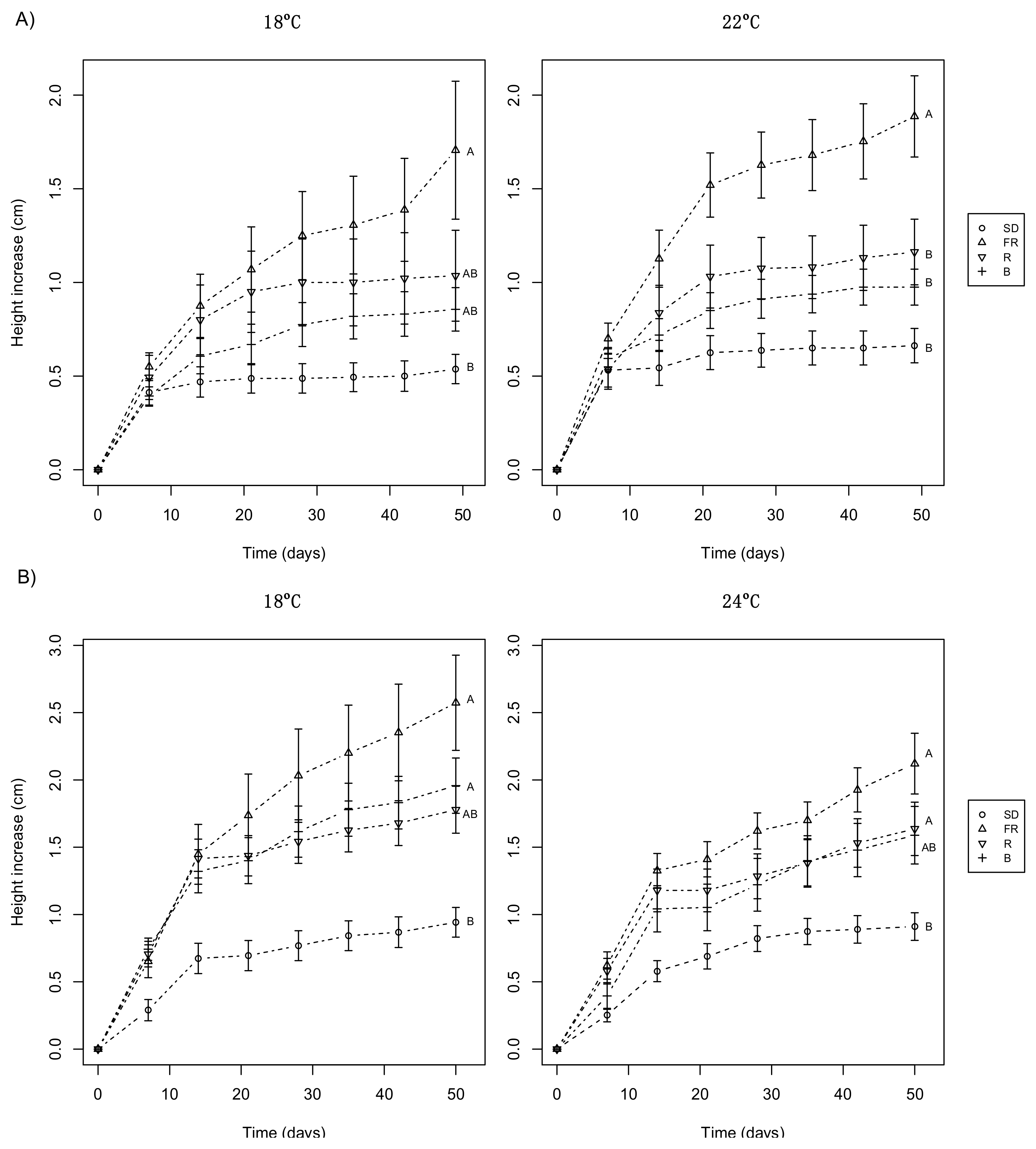{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Pre-Growing Conditions
2.2. Experimental Design and Conditions
2.3. Recording of Growth Parameters
2.4. Statistical Analysis
3. Results
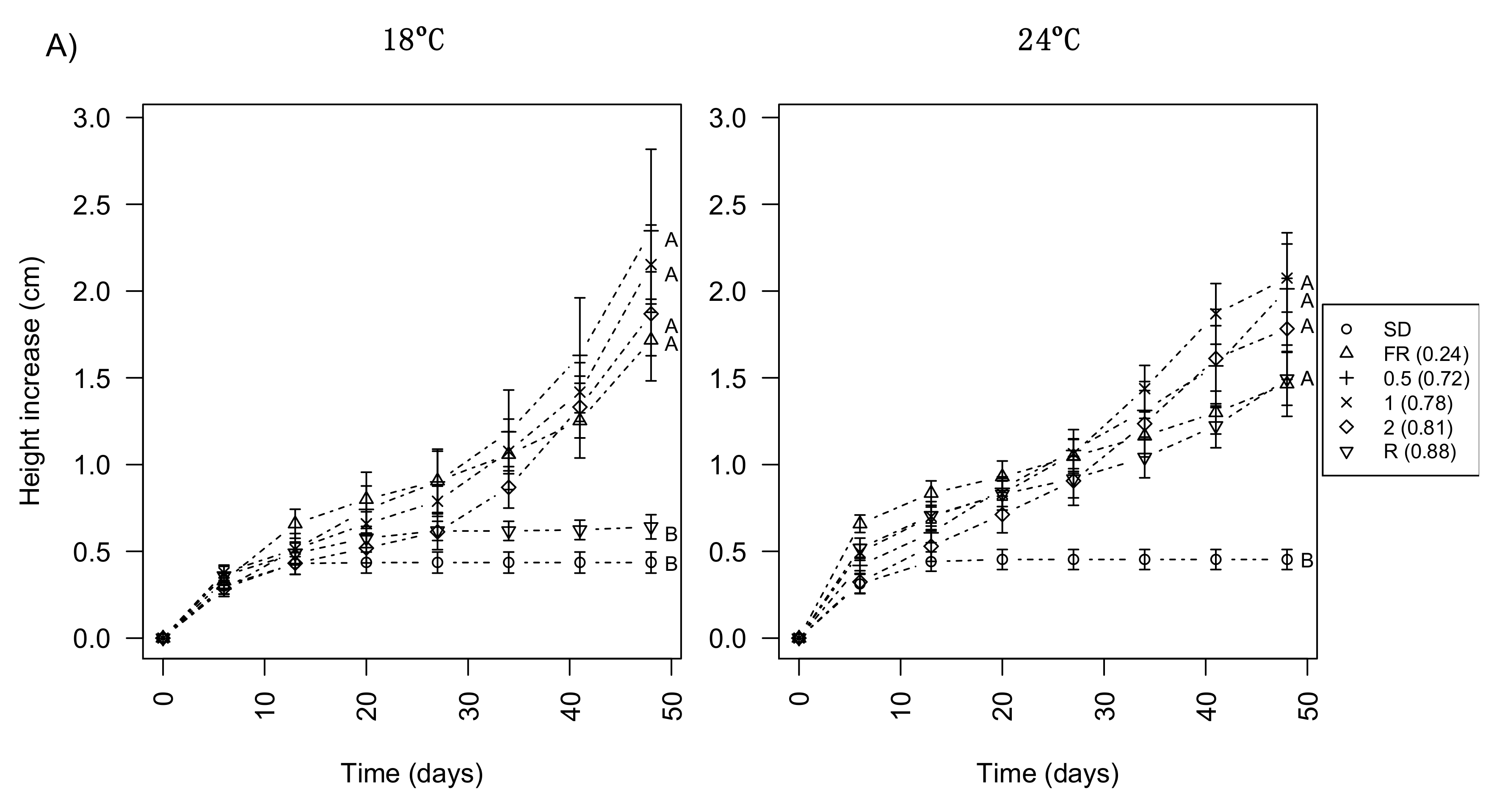
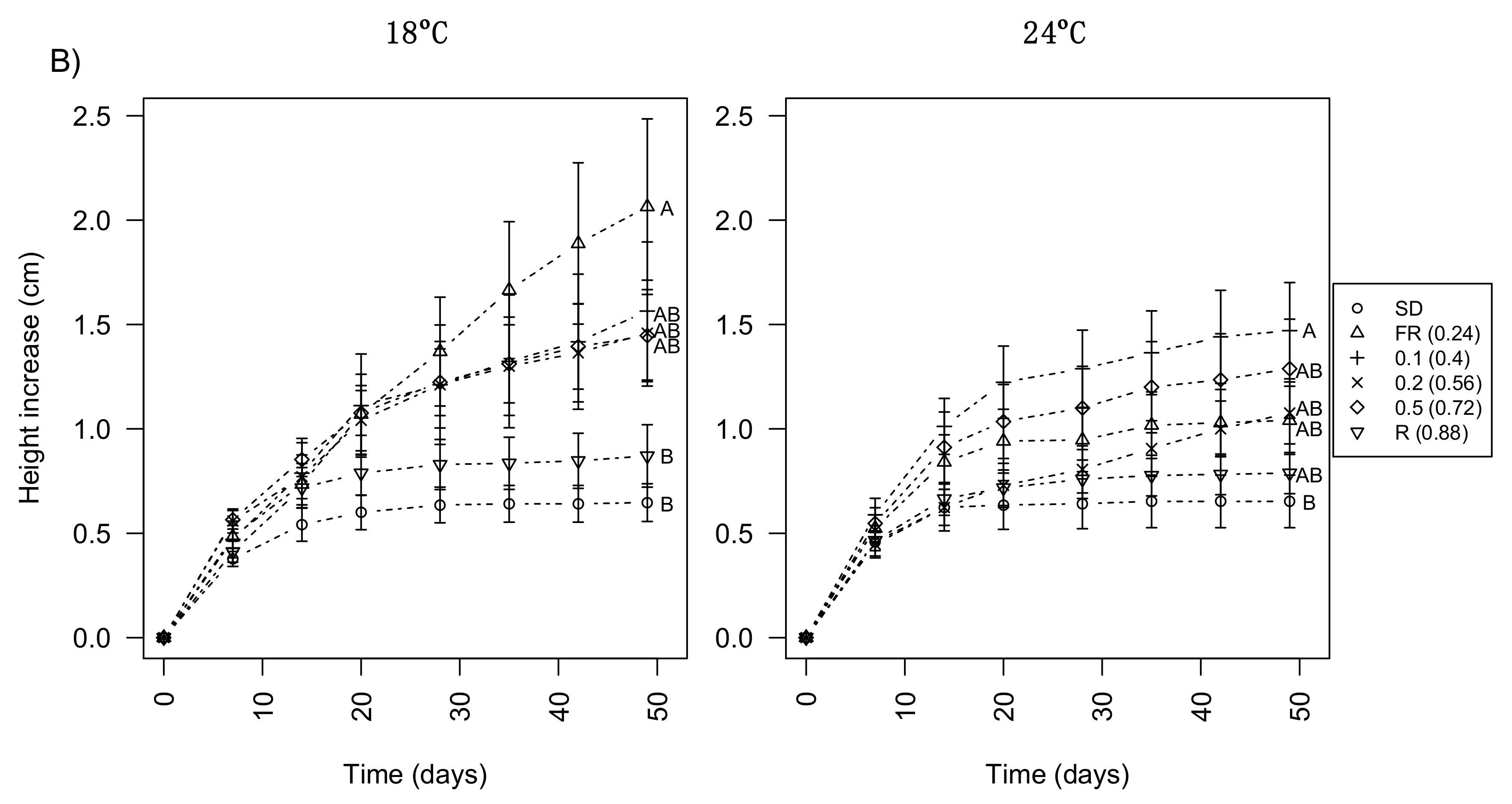
3.1. Day Extension with FR Light Enhances Shoot Elongation in Subalpine Fir Seedlings
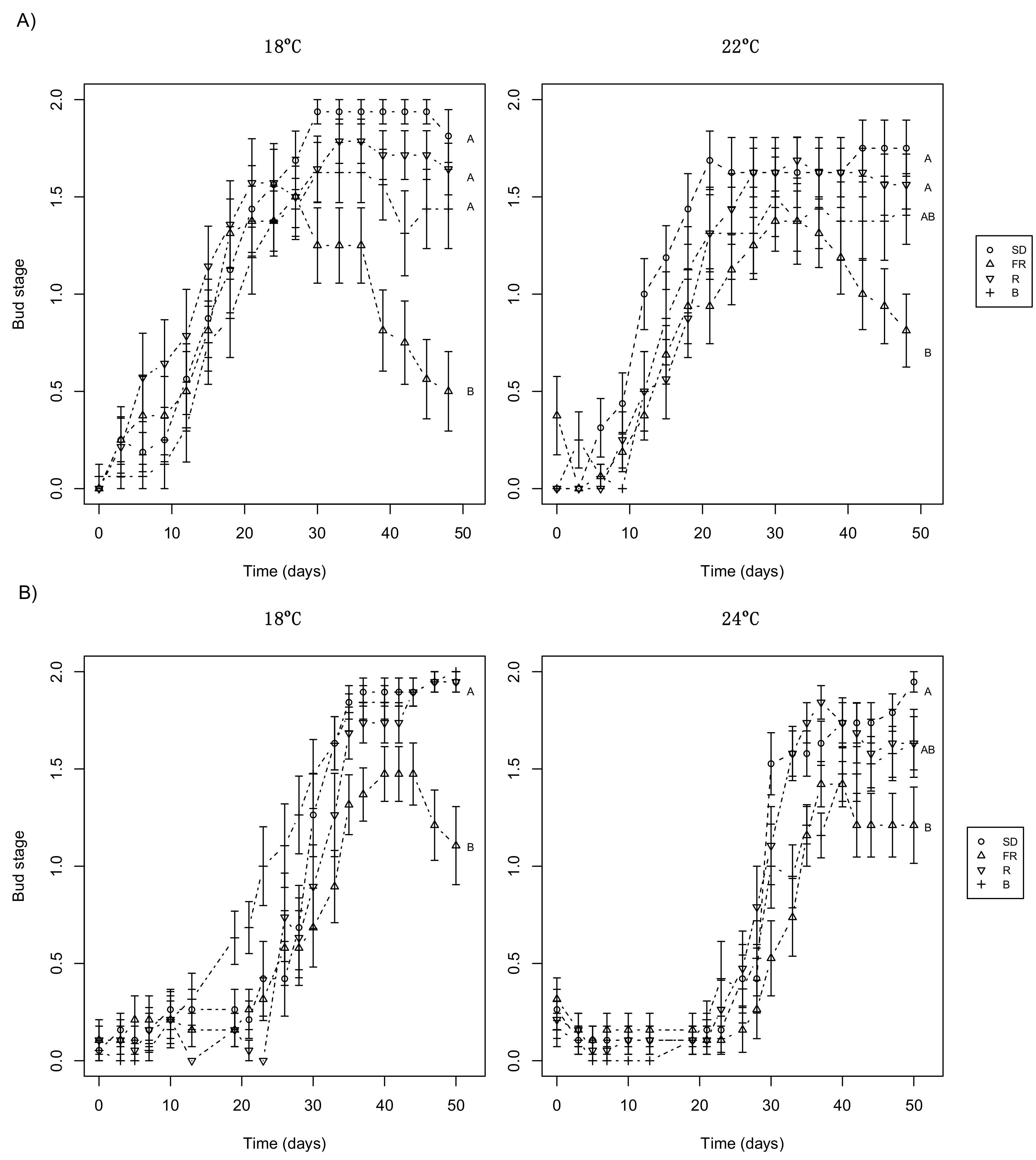
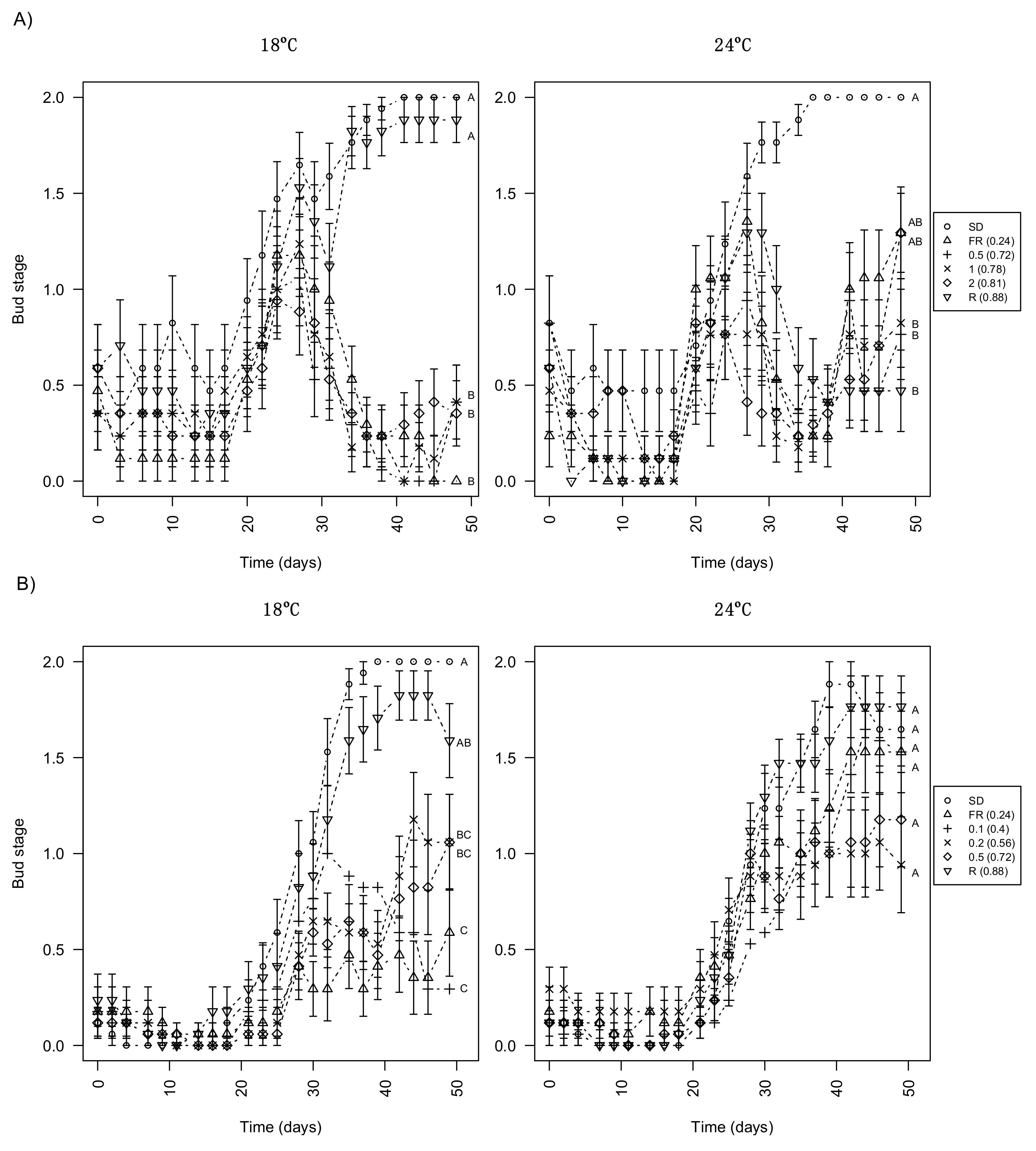
3.2. Day Extension with FR Light Delays Terminal Bud Development
3.3. Day Extension with Different R:FR Ratios Delays Bud Development Similarly to FR Only
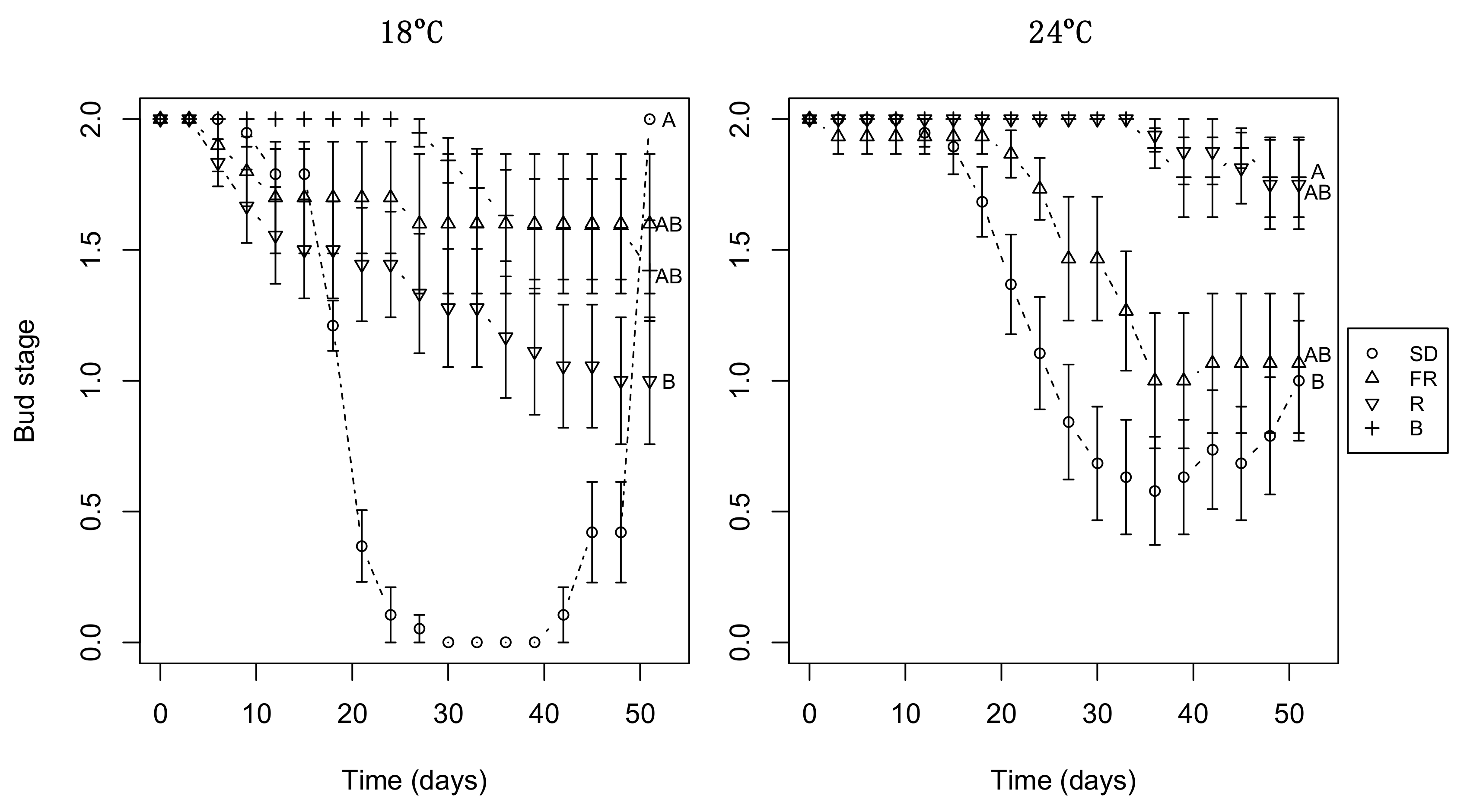
3.4. Day Extension Treatment Affects Subsequent Bud Burst
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Parker, W.H.; Maze, J. Intraspecific variation in Abies lasiocarpa from British Columbia and Washington. Am. J. Bot. 1984, 71, 1051–1059. [Google Scholar] [CrossRef]
- Strande, J.A. Christmas trees in “the land of the midnight sun”. In Proceedings of the 12th International Christmas Tree Research and Extension Conference, Honne, Norway, 6–11 September 2015; NIBIO: Ås, Norway, Talgø, V., Fløistad, I.S., Eds.; Volume 1, p. 16. [Google Scholar]
- Mattsson, A. Predicting field performance using seedling quality assessment. New For. 1997, 13, 227–252. [Google Scholar] [CrossRef]
- Sæbø, A.; Fløistad, I.S.; Netland, J.; Skulason, B.; Edvardsen, Ø. Weed control measures in Christmas tree plantations of Abies nordmanniana and Abies lasiocarpa on agricultural land. New For. 2009, 38, 143–156. [Google Scholar] [CrossRef]
- Strande, J.A. Forest Economical Analysis of Input Factors in Christmas Tree Production. Master’s Thesis, Norwegian University of Life Sciences, Ås, Norway, 2013. [Google Scholar]
- Wareing, P.J. Photoperiodism in woody plants. Annu. Rev. Plant. Physiol. 1956, 7, 191–214. [Google Scholar] [CrossRef]
- Nitsch, J.P. Photoperiodism in woody plants. Proc. Am. Soc. Hort. Sci. 1957, 70, 526–544. [Google Scholar]
- Olsen, J.E. Light and temperature sensing and signaling in induction of bud dormancy in woody plants. Plant Mol. Biol. 2010, 73, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.E.; Lee, Y.K. Trees and boreal forests. In Temperature Adaptation in a Changing Climate: Nature at Risk; Storey, K., Tanino, K., Eds.; CABI Climate Change Series; CABI: Wallingford, UK, 2011; pp. 160–178. ISBN 978-1-84593-822-2. [Google Scholar]
- Håbjørg, A. Effects of photoperiod and temperature on growth and development of three latitudinal and three altitudinal populations of Betula pubescens Ehrh. Meld. Nor. Landbrukshøgsk. 1972, 51, 1–27. [Google Scholar]
- Junttila, O.; Kaurin, Å. Climatic control of apical growth cessation in latitudinal ecotypes of Salix pentandra L. In Plant Production in the North; Kaurin, Å., Junttila, O., Nilsen, J., Eds.; Norwegian University Press: Oslo, Norway, 1985; pp. 83–91. ISBN 82-00-07385-8. [Google Scholar]
- Clapham, D.; Dormling, I.; Ekberg, I.; Eriksson, G.; Qamaruddin, M.; Vince-Prue, D. Latitudinal cline of requirement for far-red light for the photoperiodic control of bud set and extension growth in Picea abies (Norway spruce). Physiol. Plant. 1998, 102, 71–78. [Google Scholar] [CrossRef]
- Clapham, D.; Ekberg, I.; Eriksson, G.; Norell, L.; Vince-Prue, D. Requirement for far-red light to maintain secondary needle extension growth in northern but not southern populations of Pinus sylvestris (Scots pine). Physiol. Plant. 2002, 114, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Tsegay, B.A.; Lund, L.; Nilsen, J.; Olsen, J.E.; Mølmann, J.M.; Ernstsen, A.; Junttila, O. Growth responses of Betula pendula ecotypes to red and far-red light. Electron. J. Biotechnol. 2005, 8, 17–23. [Google Scholar] [CrossRef]
- Mølmann, J.A.; Junttila, O.; Johnsen, Ø.; Olsen, J.E. Effects of red, far-red and blue light in maintaining growth in latitudinal populations of Norway spruce (Picea abies). Plant Cell Environ. 2006, 29, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Opseth, L.; Holefors, A.; Rosnes, A.K.R.; Lee, Y.K.; Olsen, J.E. FTL2 preceding bud set corresponds with timing of bud set in Norway spruce provenances under different light quality treatments. Environ. Exp. Bot. 2016, 121, 121–131. [Google Scholar] [CrossRef]
- Van Ieperen, W. Plant morphological and developmental responses to light quality in a horticultural context. Acta Hort. 2012, 956, 131–140. [Google Scholar] [CrossRef]
- Zhao, X.; Yu, X.; Foo, E.; Synons, G.M.; Lopez, J.; Bendehakkalu, K.T.; Xiang, J.; Weller, J.L.; Liu, X.; Reid, J.B.; et al. A study of gibberellin homeostasis and cryptochrome-mediated blue light inhibition of hypocotyl elongation. Plant. Physiol. 2007, 145, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.; Lee, C.; Chakrabarty, D.; Pae, K. Growth responses of marigold and salvia bedding plants as affected by monochromic or mixture radiation provided by a light-emitting diode (LED). Plant Growth Regul. 2002, 3, 225–230. [Google Scholar] [CrossRef]
- Fukuda, N.; Ishii, Y.; Ezura, H.; Olsen, J.E. Effects of light quality under red and blue light emitting diodes on growth and expression of FBP28 in petunia. Acta Hort. 2011, 907, 361–366. [Google Scholar] [CrossRef]
- Gautham, P.; Terfa, M.T.; Olsen, J.E.; Torre, S. Red and blue effects on morphology and flowering of Petunia x hybrida. Scienta Hort. 2015, 184, 171–178. [Google Scholar] [CrossRef]
- Morgan, D.C.; Smith, H. A systematic relationship between phytochrome-controlled development and species habitat, for plants grown in simulated natural radiation. Planta 1979, 145, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Feller, M.C. Influence of ecological conditions on Engelmann spruce (Picea engelmannii) and subalpine fir (Abies lasiocarpa) germinant survival and initial seedling growth in south-central British Columbia. For. Ecol. Manag. 1998, 107, 55–69. [Google Scholar] [CrossRef]
- Klinka, K.; Wang, Q.; Kayahara, G.J.; Carter, R.E.; Blackwell, B.A. Light-growth response relationships in Pacific silver fir (Abies amabilis) and subalpine fir (Abies lasiocarpa). Can. J. Bot. 1992, 70, 1919–1930. [Google Scholar] [CrossRef]
- Tanino, K.; Kalcsits, L.; Silim, S.; Kendall, E.; Gray, G.R. Temperature-driven plasticity in growth cessation and dormancy development in deciduous woody plants: A working hypothesis suggesting how molecular and cellular function is affected by temperature during dormancy induction. Plant Mol. Biol. 2010, 73, 49–75. [Google Scholar] [CrossRef] [PubMed]
- Rohde, A.; Bastien, C.; Boerjan, W. Temperature signals contribute to the timing of photoperiodic growth cessation and bud set in poplar. Tree Physiol. 2011, 31, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.E.K.; Eriksson, M.; Junttila, O. The dynamic nature of bud dormancy in trees: Environmental control and molecular mechanisms. Plant Cell. Environ. 2012, 35, 1707–1728. [Google Scholar] [CrossRef] [PubMed]
- Junttila, O.; Hänninen, H. The minimum temperature for budburst in Betula depends on the state of dormancy. Tree Physiol. 2012, 32, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Westergaard, L.; Eriksen, E.N. Autumn temperature affects the induction of dormancy in first-year seedlings of Acer platanoides L. Scand. J. For. Res. 1997, 12, 11–16. [Google Scholar] [CrossRef]
- Heide, O.M. High autumn temperature delays spring bud burst in boreal trees, counterbalancing the effect of climatic warming. Tree Physiol. 2003, 23, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Junttila, O.; Nilsen, J.; Igeland, B. Effect of temperature in the induction of bud dormancy in various ecotypes of Betula pubescens and B. pendula. Scand. J. For. Res. 2003, 18, 208–217. [Google Scholar] [CrossRef]
- Søgaard, G.; Johnsen, Ø.; Nilsen, J.; Junttila, O. Climatic control of bud burst in young seedlings of nine provenances of Norway spruce. Tree Physiol. 2008, 28, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Kalcsits, L.; Silim, S.; Tanino, K. Warm temperature accelerates short-photoperiod-induced growth cessation and dormancy induction in hybrid poplar (Populus x spp.). Trees 2009, 23, 971–979. [Google Scholar] [CrossRef]
- Olsen, J.E.; Lee, Y.K.; Junttila, O. Effect of alternating day and night temperature on short day-induced bud set and subsequent bud burst in long days in Norway spruce. Front. Plant Sci. 2014, 5, 691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, J.A.; El Kayal, W.; Hart, A.T.; Runcie, D.E.; Arango-Velez, A.; Cooke, J.E.K. The joint influence of photoperiod and temperature during growth cessation and development of dormancy in white spruce (Picea glauca). Tree Physiol. 2016, 36, 1432–1448. [Google Scholar] [CrossRef] [PubMed]
- Fløistad, I.S.; Patil, G.G. Growth and terminal bud formation in Picea abies seedlings grown with alternating diurnal temperature and different light qualities. Scand. J. For. Res. 2002, 17, 15–27. [Google Scholar] [CrossRef]
- Strømme, C.B.; Julkunen-Tiito, R.; Krishna, U.; Lavola, A.; Olsen, J.E.; Nybakken, L. UV-B and temperature enhancement affect spring and autumn phenology in Populus tremula. Plant Cell Environ. 2015, 38, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Strømme, C.B.; Julkunen-Tiito, R.; Olsen, J.E.; Nybakken, L. High daytime temperature delays autumnal bud formation in Populus tremula under field conditions. Tree Physiol. 2017, 37, 71–81. [Google Scholar] [PubMed]
- Claveau, Y.; Messier, C.; Comeau, P.G.; Coates, K.D. Growth and crown morphological responses of boreal conifer seedlings and saplings with contrasting shade tolerance to a gradient of light and height. Can. J. For. Res. 2002, 32, 458–468. [Google Scholar] [CrossRef] [Green Version]
- Sager, J.C.; Smith, W.O.; Edwards, J.L.; Cyr, K.L. Photosynthetic efficiency and phytochrome photoequilibria determination using spectral data. Trans. ASAE 1988, 31, 1882–1889. [Google Scholar] [CrossRef]
- Bae, G.; Choi, G. Decoding of light signals by plant phytochromes and their interacting proteins. Ann. Rev. Plant. Biol. 2008, 59, 281–311. [Google Scholar] [CrossRef] [PubMed]
- Crawley, M.J. The R Book, 1st ed.; John Wiley and Sons Ltd.: Chichester, UK, 2007; ISBN 13-978-0-470-51024-7. [Google Scholar]
- Brelsford, C.C.; Robson, M. Blue light advances bud burst in branches of three temperate deciduous tree species under short-day conditions. BioRixv 2018. [Google Scholar] [CrossRef]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiang, C.; Aas, O.T.; Jetmundsen, M.R.; Lee, Y.; Torre, S.; Fløistad, I.S.; Olsen, J.E. Day Extension with Far-Red Light Enhances Growth of Subalpine Fir (Abies lasiocarpa (Hooker) Nuttall) Seedlings. Forests 2018, 9, 175. https://doi.org/10.3390/f9040175
Chiang C, Aas OT, Jetmundsen MR, Lee Y, Torre S, Fløistad IS, Olsen JE. Day Extension with Far-Red Light Enhances Growth of Subalpine Fir (Abies lasiocarpa (Hooker) Nuttall) Seedlings. Forests. 2018; 9(4):175. https://doi.org/10.3390/f9040175
Chicago/Turabian StyleChiang, Camilo, Oda Toresdatter Aas, Marianne Rindedal Jetmundsen, YeonKyeong Lee, Sissel Torre, Inger Sundheim Fløistad, and Jorunn E. Olsen. 2018. "Day Extension with Far-Red Light Enhances Growth of Subalpine Fir (Abies lasiocarpa (Hooker) Nuttall) Seedlings" Forests 9, no. 4: 175. https://doi.org/10.3390/f9040175