The Role of Environmental Filtering in Structuring Appalachian Tree Communities: Topographic Influences on Functional Diversity Are Mediated through Soil Characteristics

Abstract
:1. Introduction
2. Materials and Methods
3. Results
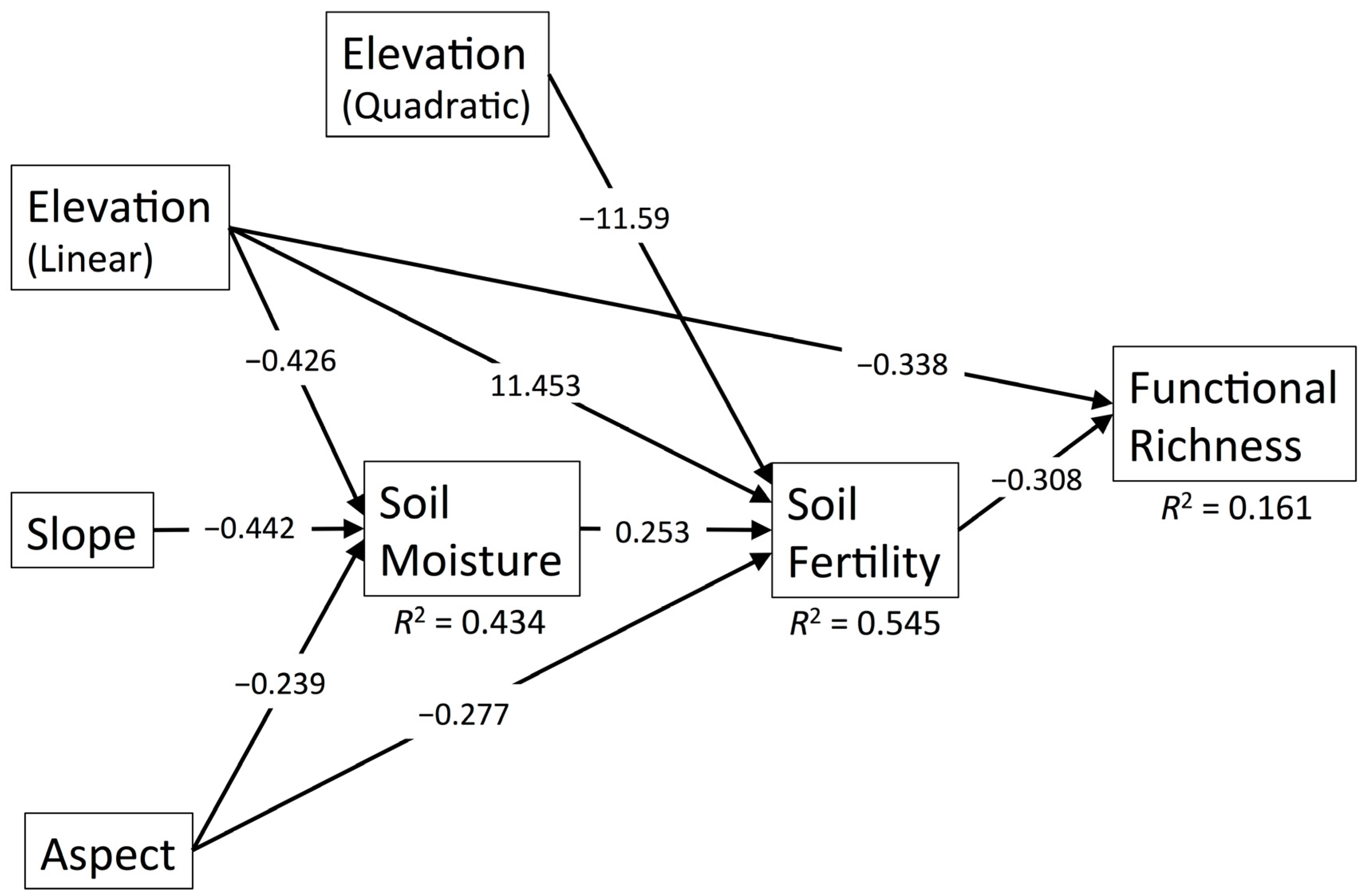
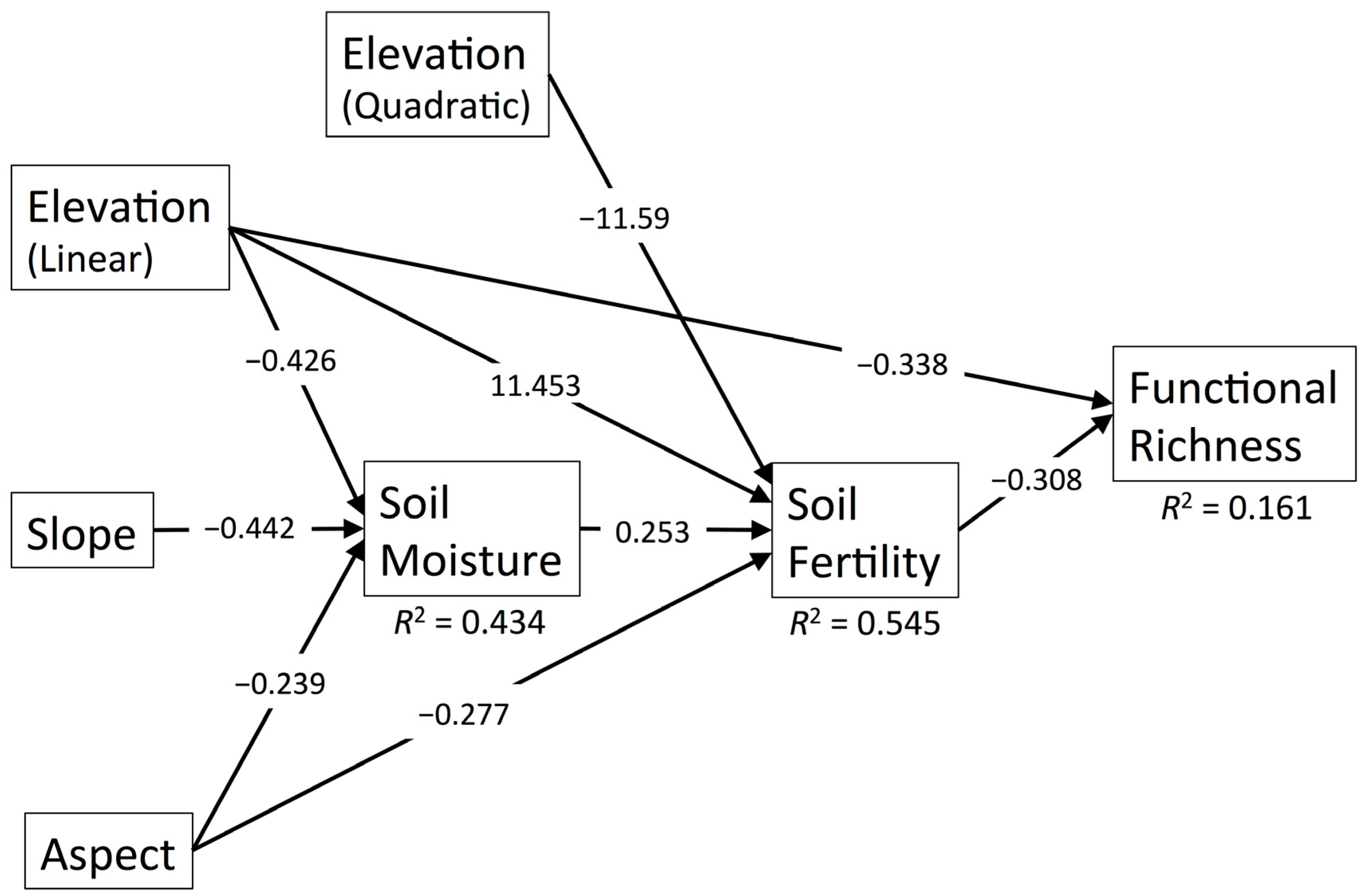
3.1. Functional Richness
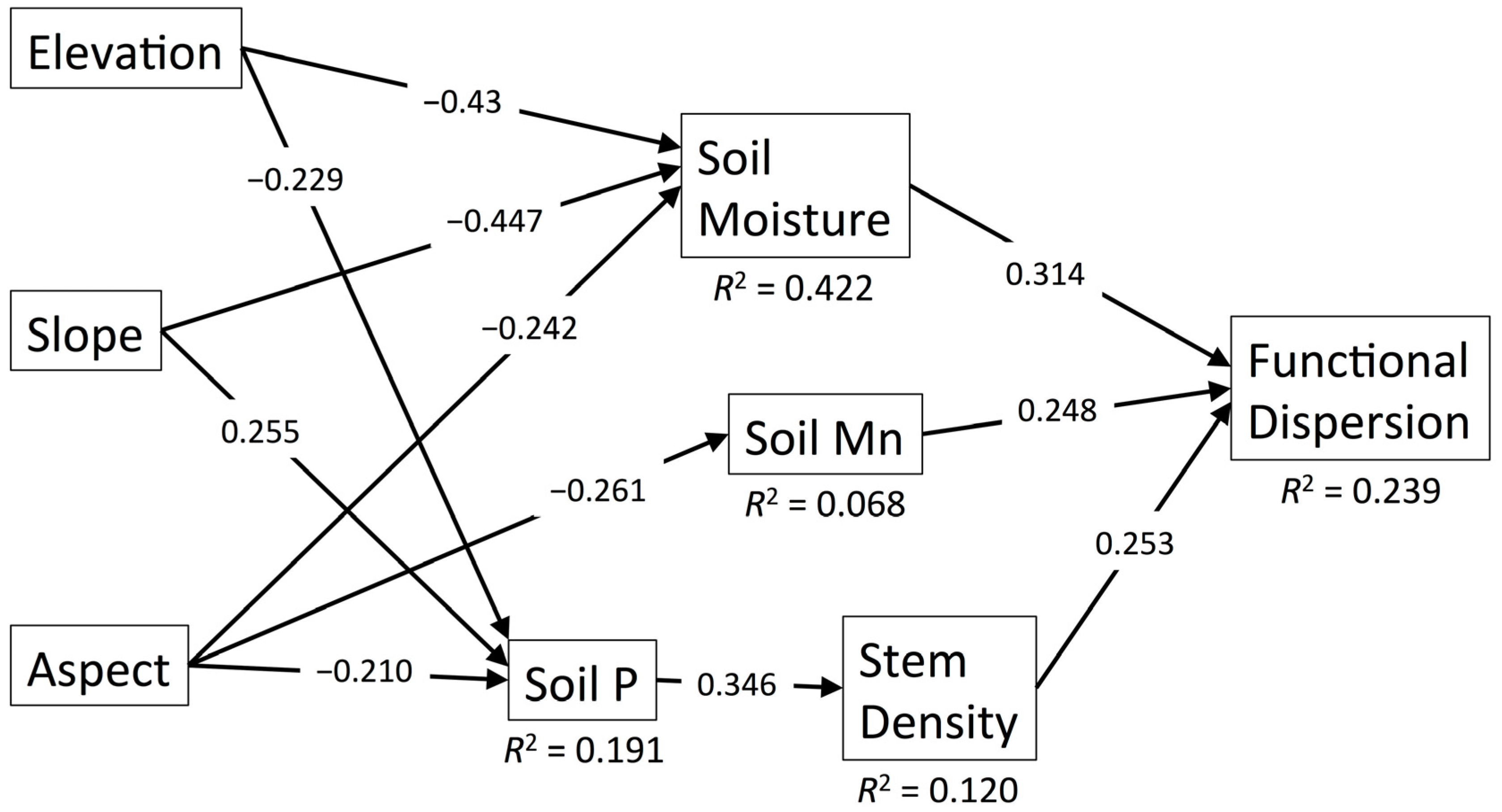
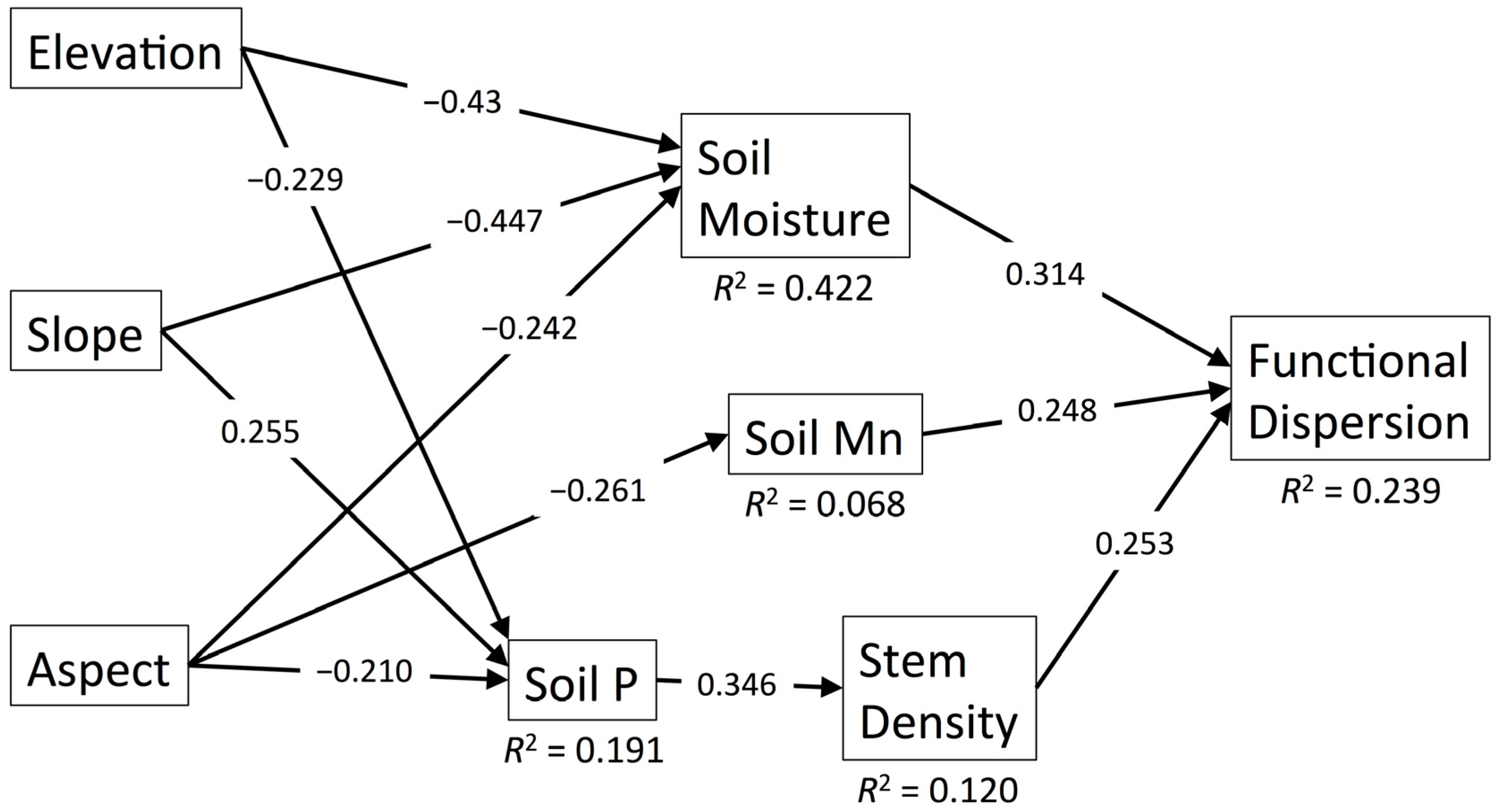
3.2. Functional Dispersion
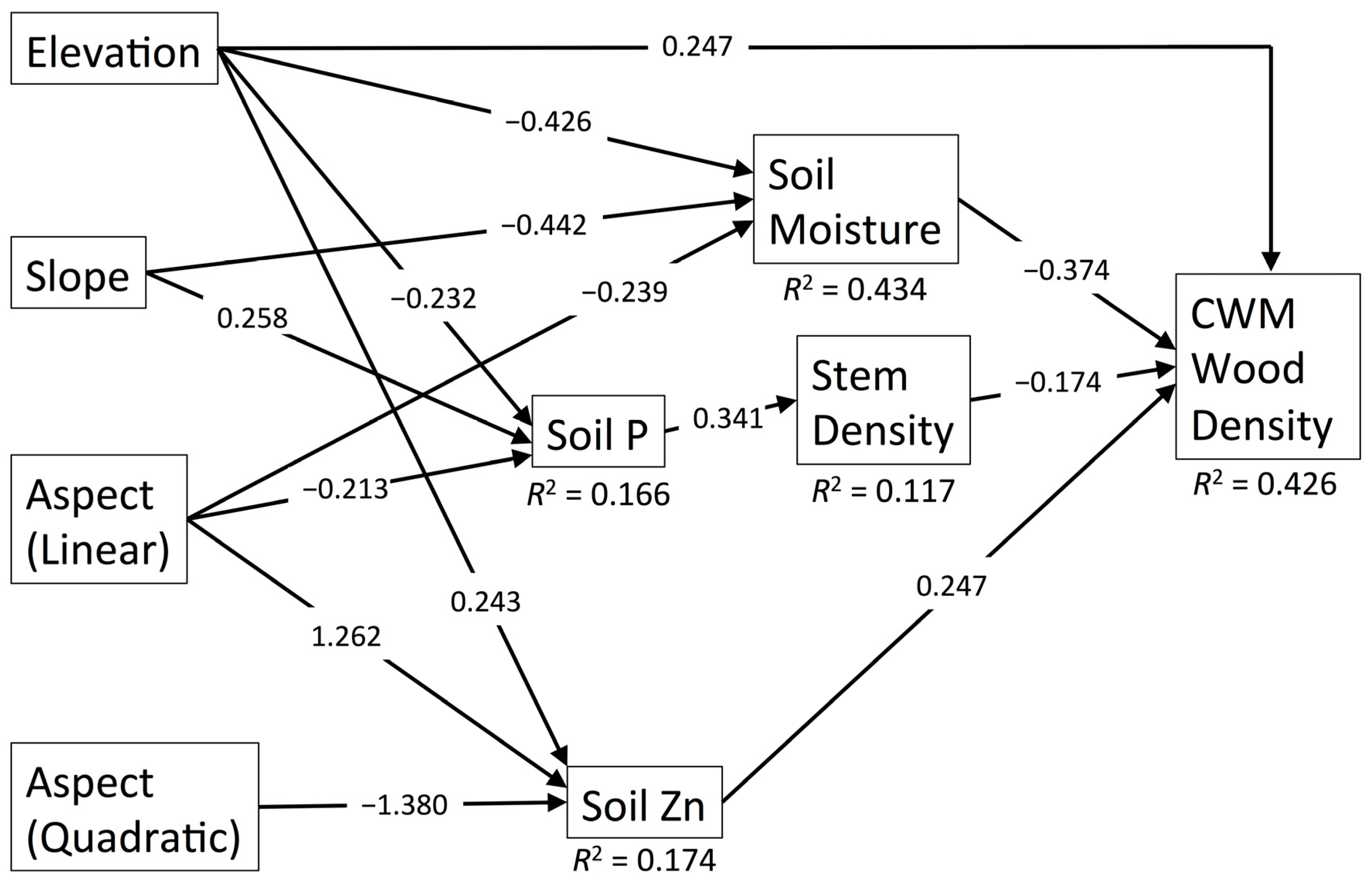
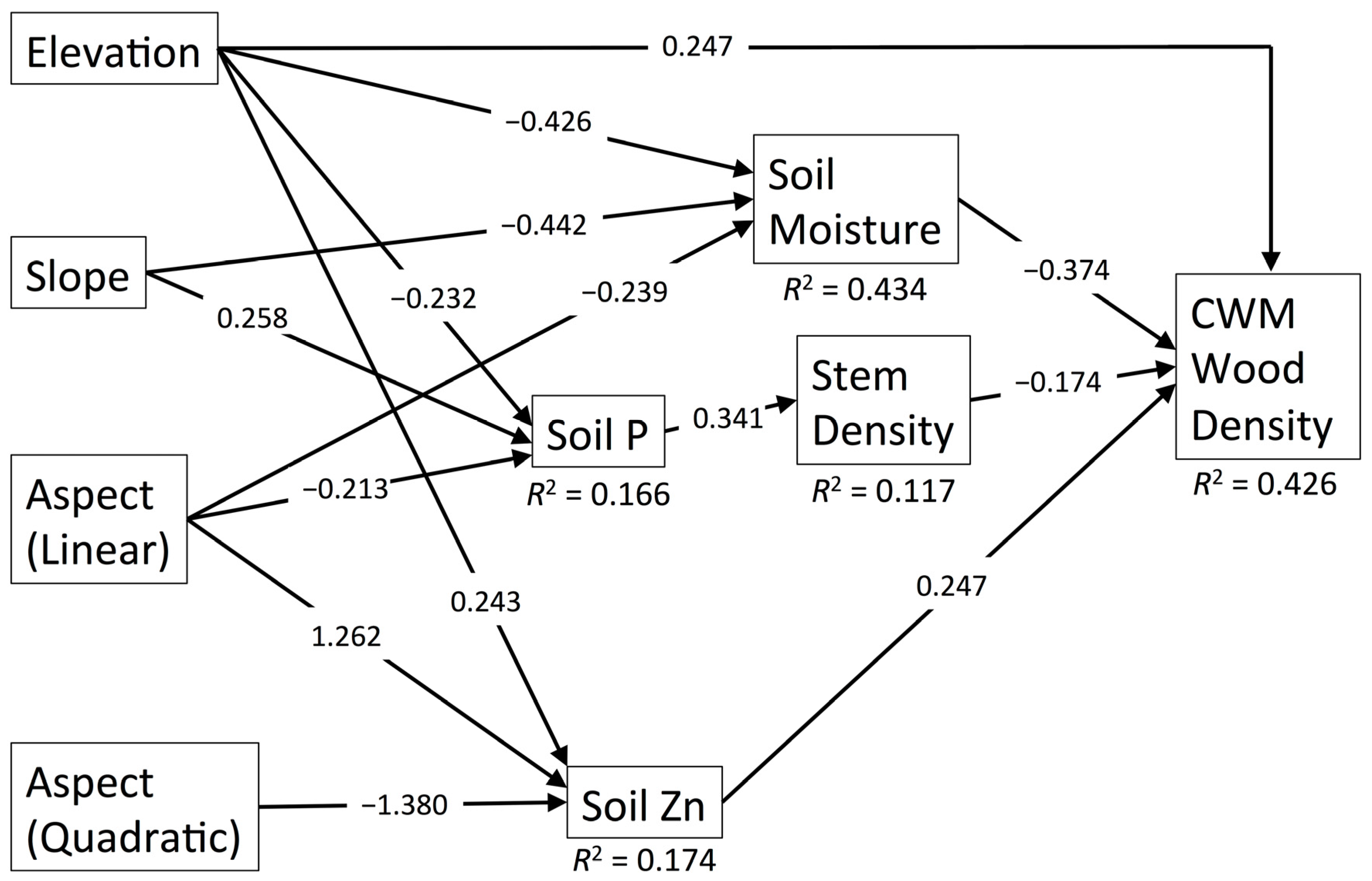
3.3. CWM Wood Density
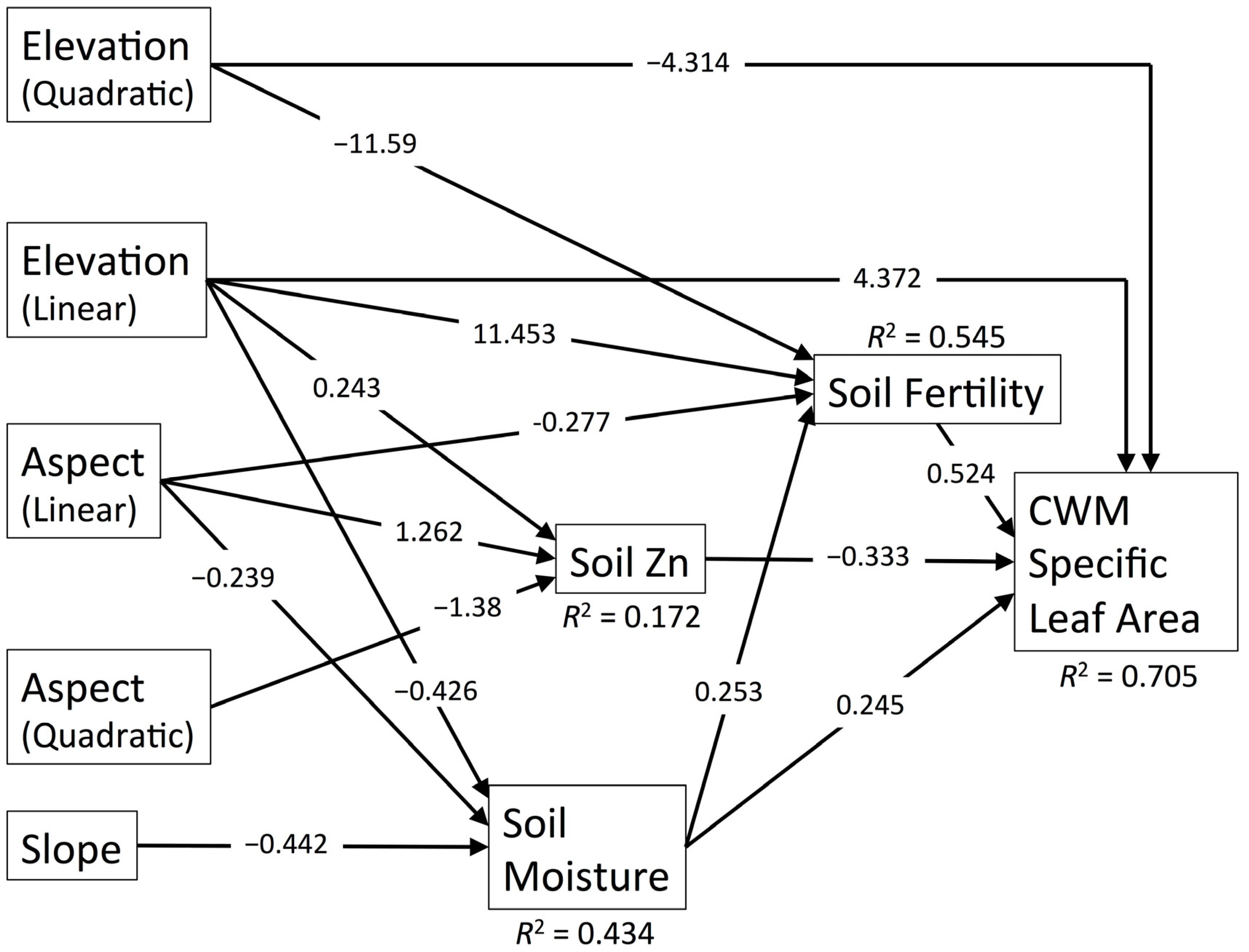
3.4. CWM Specific Leaf Area
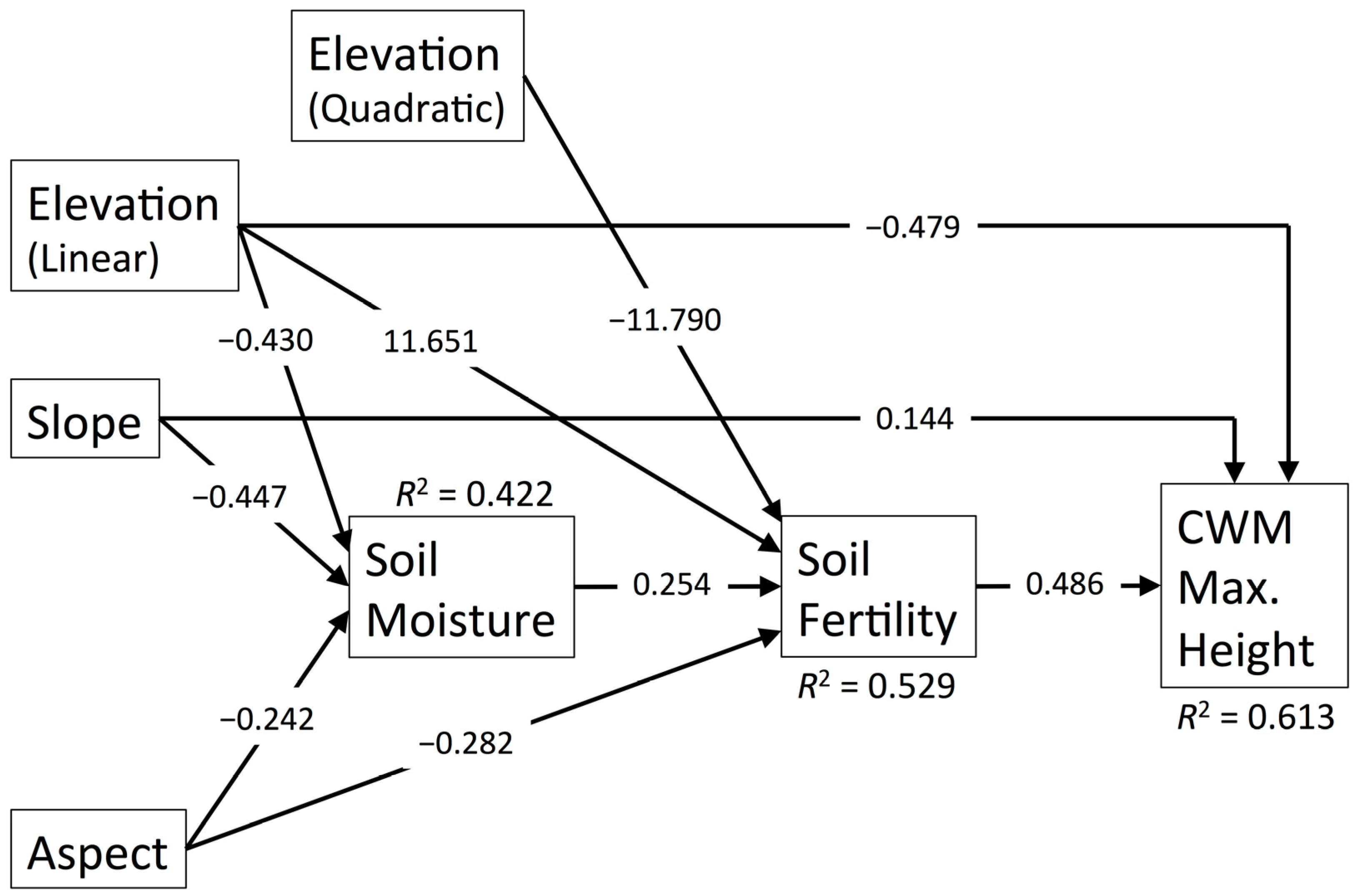
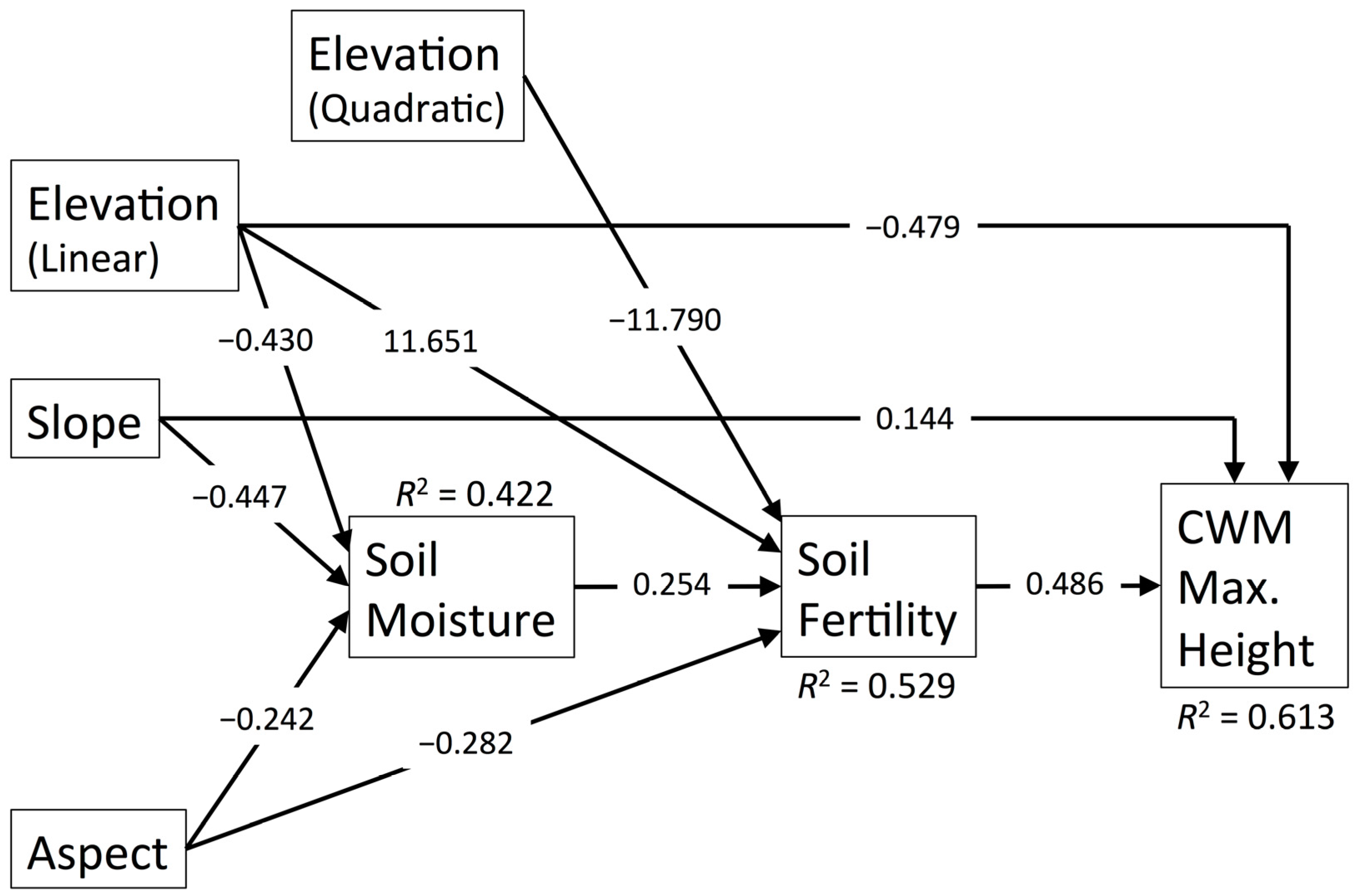
3.5. CWM Maximum Height
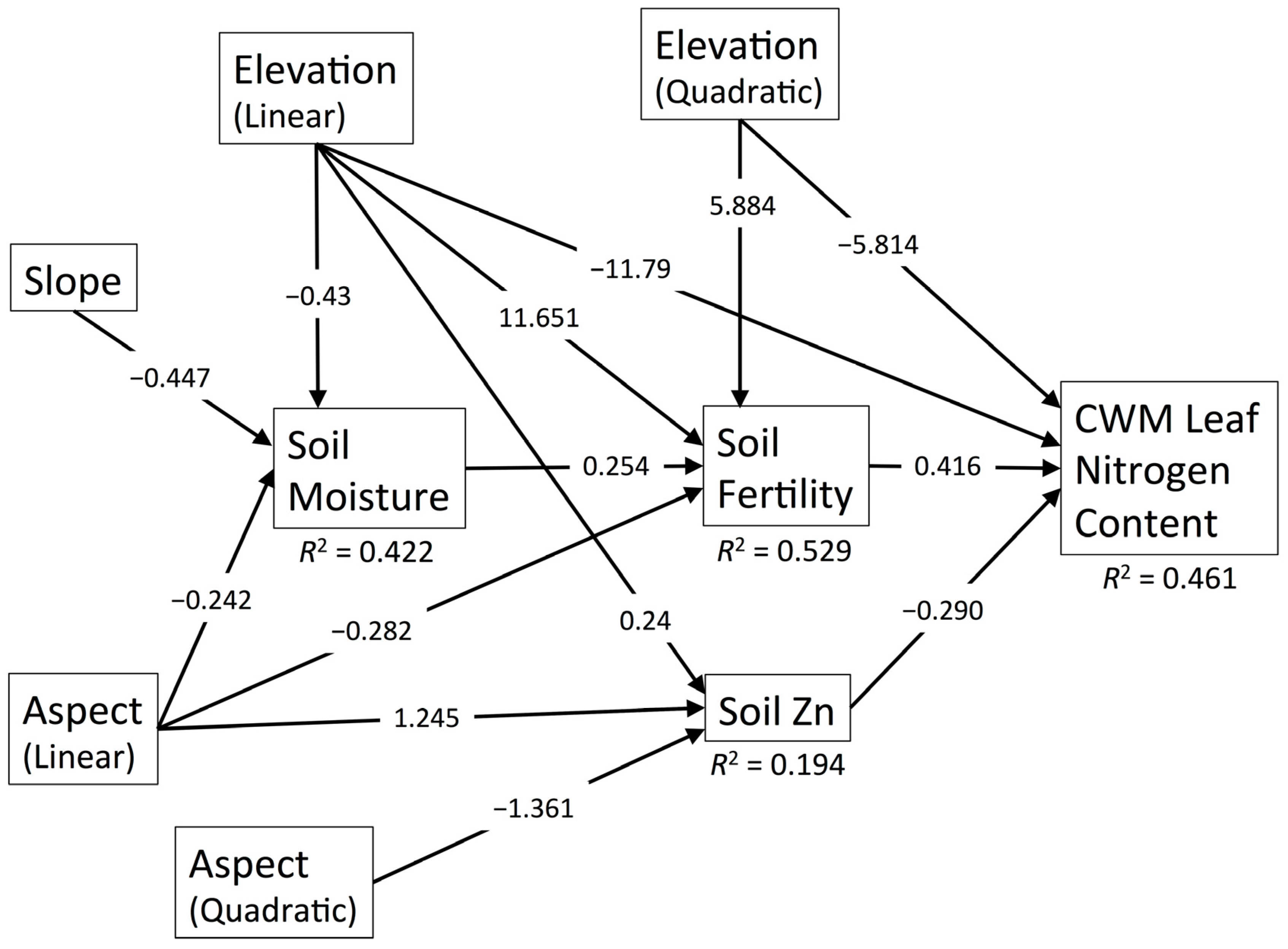
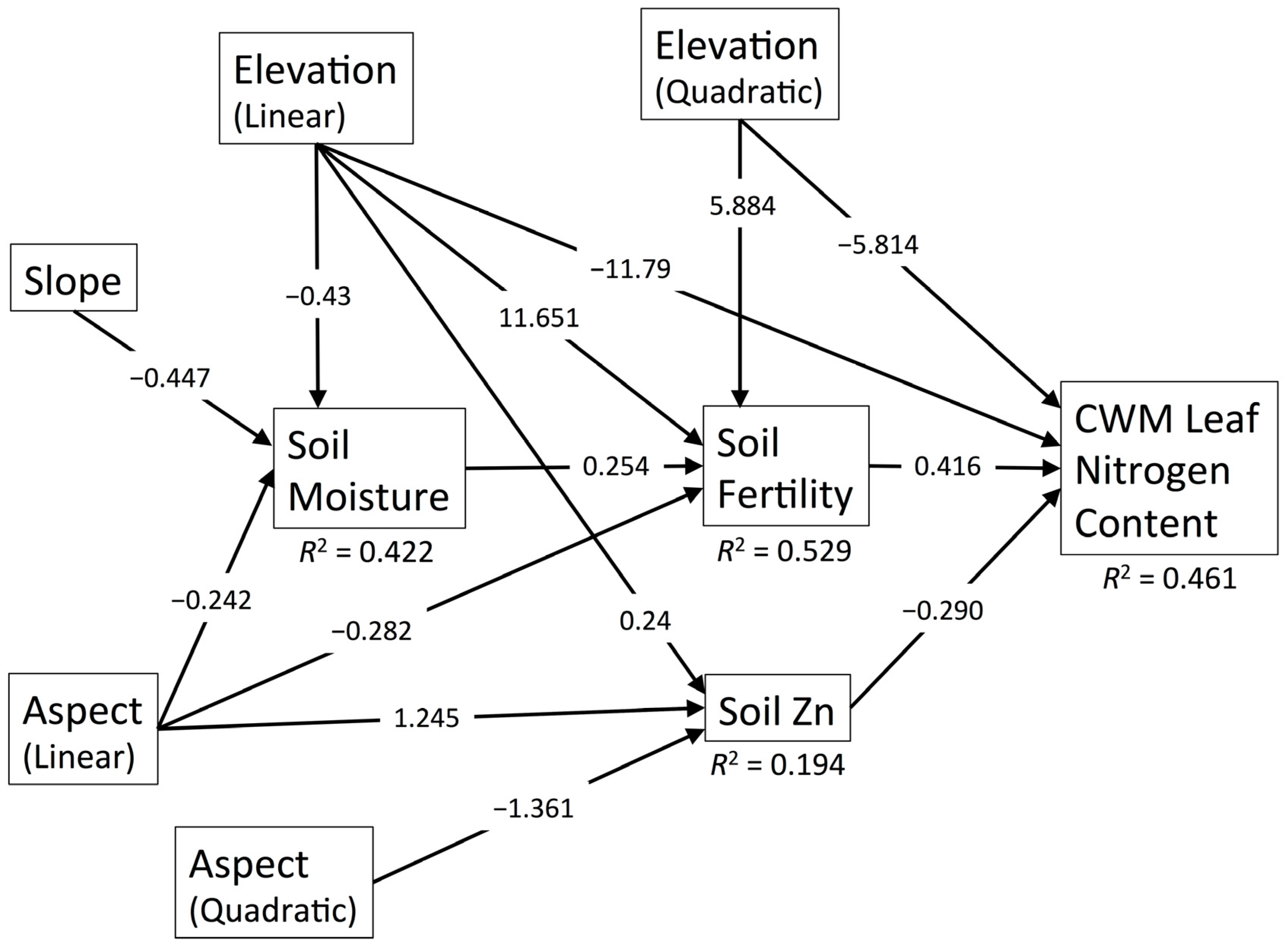
3.6. CWM Leaf Nitrogen Content
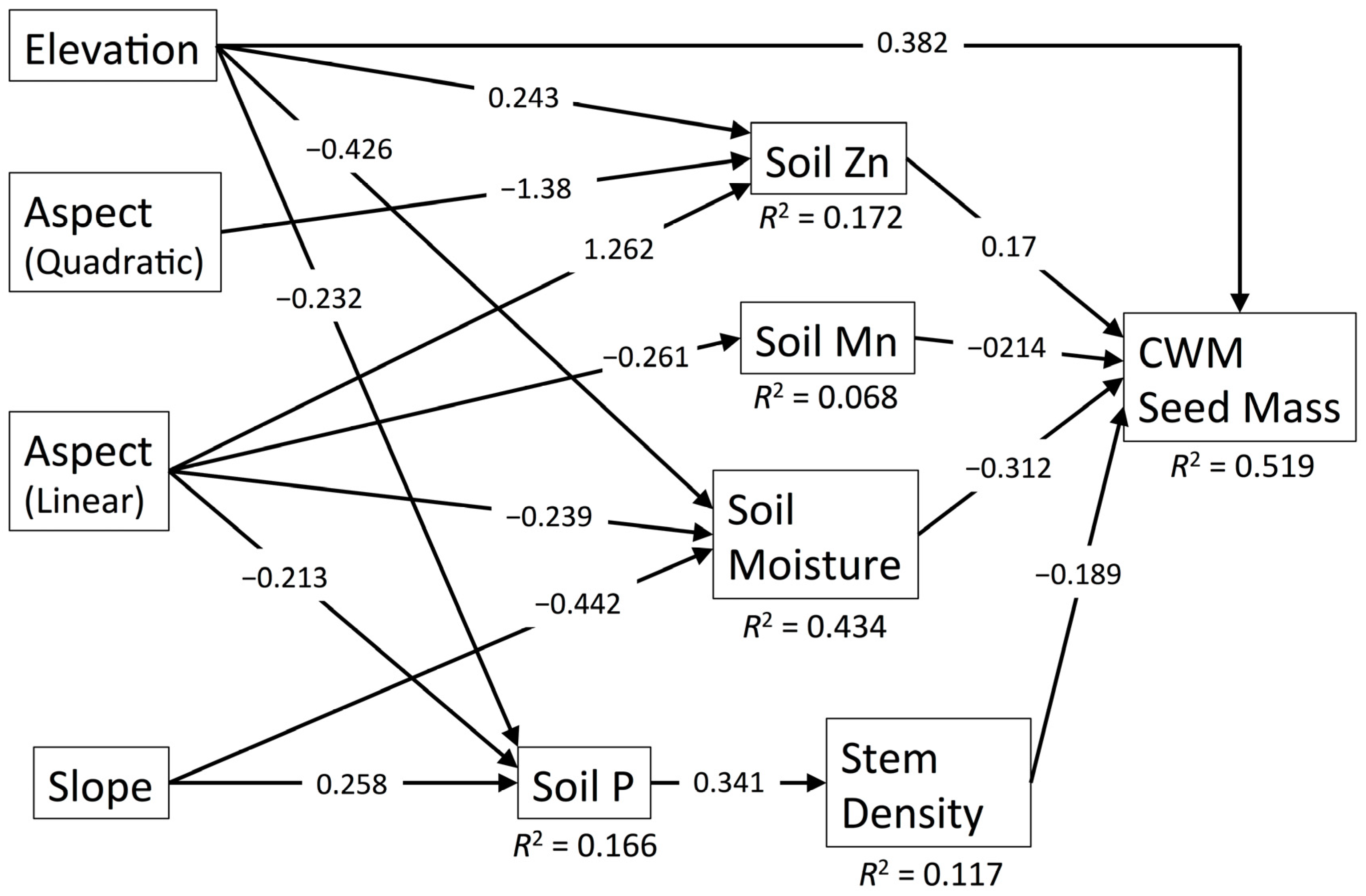
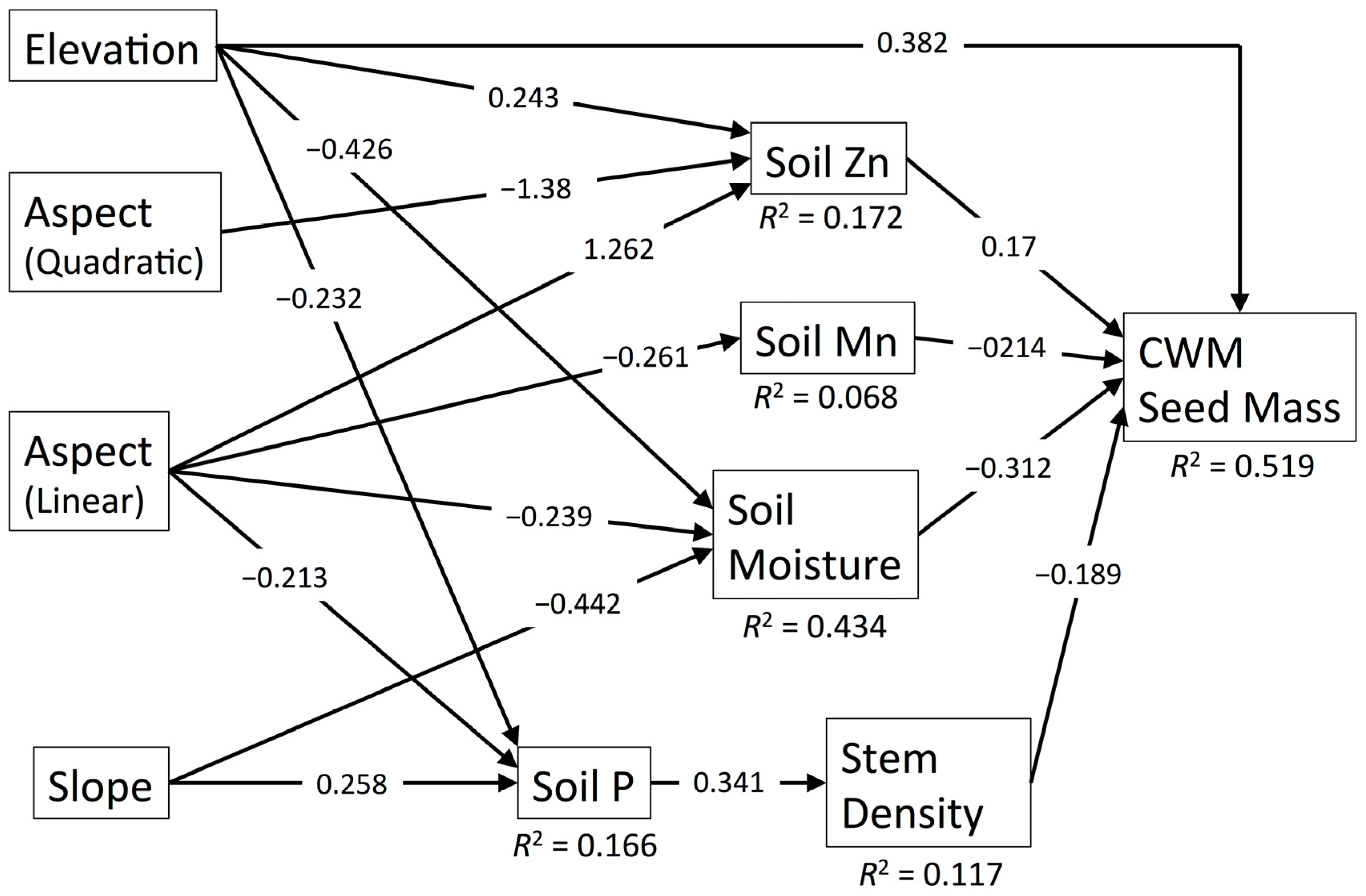
3.7. CWM Seed Mass
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Braun, E.L. The vegetation of Pine Mountain, Kentucky. Am. Midl. Nat. 1935, 16, 517–565. [Google Scholar] [CrossRef]
- Whittaker, R.H. Vegetation of the Great Smoky Mountains. Ecol. Monogr. 1956, 26, 1–80. [Google Scholar] [CrossRef]
- Braun, E.L. Forests of the Cumberland Mountains. Ecol. Monogr. 1942, 12, 413–447. [Google Scholar] [CrossRef]
- Boerner, R.E.J. Unraveling the Gordian Knot: Interactions among vegetation, topography, and soil properties in the central and southern Appalachians. J. Torrey Bot. Soc. 2006, 133, 321–361. [Google Scholar] [CrossRef]
- Hutchins, R.L.; Hill, J.D.; White, E.H. The influence of soil and microclimate on vegetation of forested slopes in eastern Kentucky. Soil Sci. 1976, 121, 234–241. [Google Scholar] [CrossRef]
- Schmalzer, P.; Hinkle, C.; DeSelm, H. Descriminant Analysis of Cove Forests of the Cumberland Plateau of Tennessee. In Proceedings of the Central Hardwoods Forest Conference II; Pope, P., Ed.; Purdue University: West Lafayette, IN, USA, 1978; pp. 62–86. [Google Scholar]
- Weiher, E.; Keddy, P.A. Assembly Rules, Null Models, and Trait Dispersion, New Questions from Old Patterns. Oikos 1995, 74, 159–164. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. A Trait-Based Test for Habitat Filtering: Convex Hull Volume. Ecology 2006, 87, 1314–1324. [Google Scholar] [CrossRef]
- Currie, D.J.; Mittelbach, G.G.; Cornell, H.V.; Field, R.; Guégan, J.F.; Hawkins, B.A.; Kaufman, D.M.; Kerr, J.T.; Oberdorff, T.; O’Brien, E.; et al. Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 2004, 7, 1121–1134. [Google Scholar] [CrossRef]
- Coyle, J.R.; Halliday, F.W.; Lopez, B.E.; Palmquist, K.A.; Wilfahrt, P.A.; Hurlbert, A.H. Using trait and phylogenetic diversity to evaluate the generality of the stress-dominance hypothesis in eastern North American tree communities. Ecography (Cop.) 2014, 37, 814–826. [Google Scholar] [CrossRef]
- Sundqvist, M.K.; Sanders, N.J.; Wardle, D.A. Community and Ecosystem Responses to Elevational Gradients: Processes, Mechanisms, and Insights for Global Change. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 261–280. [Google Scholar] [CrossRef]
- Culmsee, H.; Leuschner, C. Consistent patterns of elevational change in tree taxonomic and phylogenetic diversity across Malesian mountain forests. J. Biogeogr. 2013, 40, 1997–2010. [Google Scholar] [CrossRef]
- Fortunel, C.; Paine, C.E.T.; Fine, P.V.A.; Kraft, N.J.B.; Baraloto, C. Environmental factors predict community functional composition in Amazonian forests. J. Ecol. 2014, 102, 145–155. [Google Scholar] [CrossRef]
- Liu, J.; Yunhong, T.; Slik, J.W.F. Topography related habitat associations of tree species traits, composition and diversity in a Chinese tropical forest. For. Ecol. Manag. 2014, 330, 75–81. [Google Scholar] [CrossRef]
- Satdichanh, M.; Millet, J.; Heinimann, A. Using Plant Functional Traits and Phylogenies to Understand Patterns of Plant Community Assembly in a Seasonal Tropical Forest in Lao PDR. PLoS ONE 2015, 26, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Yu, S.-X.; Lian, J.-Y.; Shen, H.; Cao, H.-L.; Lu, H.-P.; Ye, W.-H. Inferring community assembly processes from trait diversity across environmental gradients. J. Trop. Ecol. 2016, 32, 290–299. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Valencia, R.; Ackerly, D.D. Functional Traits and Niche-based Tree Community Assembly in an Amazonian Forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Lasky, J.R.; Sun, I.F.; Su, S.H.; Chen, Z.S.; Keitt, T.H. Trait-mediated effects of environmental filtering on tree community dynamics. J. Ecol. 2013, 101, 722–733. [Google Scholar] [CrossRef]
- Kluge, J.; Kessler, M. Phylogenetic diversity, trait diversity and niches: Species assembly of ferns along a tropical elevational gradient. J. Biogeogr. 2011, 38, 394–405. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, D.; Wang, R.; Liu, J.; Du, N. Altitudinal patterns of species diversity and phylogenetic diversity across temperate mountain forests of northern China. PLoS ONE 2016, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, F.M.; Burton, J.I.; Scheller, R.M.; Amatangelo, K.L.; Mladenoff, D.J. Functional diversity of ground-layer plant communities in old-growth and managed northern hardwood forests. Appl. Veg. Sci. 2014, 17, 398–407. [Google Scholar] [CrossRef]
- Spasojevic, M.J.; Grace, J.B.; Harrison, S.; Damschen, E.I. Functional diversity supports the physiological tolerance hypothesis for plant species richness along climatic gradients. J. Ecol. 2014, 102, 447–455. [Google Scholar] [CrossRef]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, R.; Mimura, M.; Mori, A.S.; Sakai, A. Topographic patterns in the phylogenetic structure of temperate forests on steep mountainous terrain. AoB Plants 2015, 7, plv134. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, G.E.; Wolfe, J.N. Soil Moisture Investigations at Neotoma, a Forest Bioclimatic Laboratory in Central Ohio. Ohio J. Sci. 1959, 59, 38–46. [Google Scholar]
- Franzmeier, D.; Pederson, E.; Longwell, T.; Byrne, J.; Losche, C. Properties of Some Soils in the Cumberland Plateau as Related to Slope Aspect and Position. Soil Sci. Soc. Am. Proc. 1969, 33, 755–761. [Google Scholar] [CrossRef]
- Losche, C.; McCracken, R.; Davey, C. Soils of Steeply Sloping Landscapes in Southern Appalachian Mountains. Soil Sci. Soc. Am. Proc. 1970, 4, 473–478. [Google Scholar] [CrossRef]
- Hairston, A.; Grigal, D. Topographic influences on soils and trees within single mapping units on a sandy outwash landscape. For. Ecol. Manag. 1991, 43, 35–45. [Google Scholar] [CrossRef]
- Chapman, J.I.; McEwan, R.W. Thirty Years of Compositional Change in an Old-Growth Temperate Forest: The Role of Topographic Gradients in Oak-Maple Dynamics. PLoS ONE 2016, 11, e0160238. [Google Scholar] [CrossRef] [PubMed]
- McEwan, R.W.; Muller, R.N. Spatial and temporal dynamics in canopy dominance of an old-growth central Appalachian forest. Can. J. For. Res. 2006, 36, 1536–1550. [Google Scholar] [CrossRef]
- McEwan, R.W.; Muller, R.N.; McCarthy, B.C. Vegetation-Environment Relationships Among Woody Species in Four Canopy-Layers in an Old-Growth Mixed Mesophytic Forest. Castanea 2005, 70, 32–46. [Google Scholar] [CrossRef]
- Chapman, J.I.; McEwan, R.W. Spatiotemporal dynamics of a and β diversity across topographic gradients in the herbaceous layer of an old-growth deciduous forest. Oikos 2013, 122, 1679–1686. [Google Scholar] [CrossRef]
- Martin, W.H. The Lilley Cornett Woods: A Stable Mixed Mesophytic Forest in Kentucky. Bot. Gaz. 1975, 136, 171–183. [Google Scholar] [CrossRef]
- Trewartha, G. An Introduction to Climate, 4th ed.; McGraw and Hill: New York, NY, USA, 1968. [Google Scholar]
- Hill, J.D. Climate of Kentucky, Report No 221; University of Kentucky College of Agriculture: Lexington, KY, USA, 1976. [Google Scholar]
- Muller, R.N. Vegetation Patterns in the Mixed Mesophytic Forest of Eastern Kentucky. Ecology 1982, 63, 1901–1917. [Google Scholar] [CrossRef]
- Braun, E.L. Deciduous Forests of Eastern North America; The Blakiston Company: Philadelphia, PA, USA, 1950. [Google Scholar]
- Carter, M. Soil Sampling and Methods of Analysis; Lews Publishing: Boca Raton, FL, USA, 1993. [Google Scholar]
- McEwan, R.W.; Chapman, J.I.; Muller, R.N. 2010 Lilley Cornett Woods Overstory Data. Data Files: Big Everidge Hollow Permanent Plots. Available online: http://ecommons.udayton.edu/mcewanlab_1_data/1 (accessed on 5 January 2018). [CrossRef]
- McEwan, R.W.; Chapman, J.I.; Muller, R.N. 2001 Lilley Cornett Woods Soil Data. Data Files: Big Everidge Hollow Permanent Plots. Available online: http://ecommons.udayton.edu/mcewanlab_1_data/6 (accessed on 5 January 2018). [CrossRef]
- McEwan, R.W.; Chapman, J.I.; Muller, R.N. Lilley Cornett Woods Plot Information and Topography Data. Data Files: Big Everidge Hollow Permanent Plots. Available online: http://ecommons.udayton.edu/mcewanlab_1_data/8 (accessed on 5 January 2018). [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Bonisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. A global database of plant traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef] [Green Version]
- Kattge, J.; Knorr, W.; Raddatz, T.; Wirth, C. Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Glob. Chang. Biol. 2009, 15, 976–991. [Google Scholar] [CrossRef]
- Bolster, K.; Martin, M.; Aber, J. Determination of carbon fraction and nitrogen concentration in tree foliage by near infrared reflectance: Comparison of statistical methods. Can. J. For. Res. 1996, 26, 590–600. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Harmon, M.E.; Woodall, C.W.; Fasth, B.; Sexton, J. Woody Detritus Density and Density Reduction Factors for Tree Species in the United States: A Synthesis; USDA Forest Service Northern Research Station: Newtown Square, PA, USA, 2008; p. 84.
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Data from: Towards a worldwide wood economics spectrum. Dryad Digit. Repos. 2009. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Seed size and plant strategy across the whole life cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Kew, R.B.G. Seed Information Database (SID). Version 7.1. Available online: http://data.kew.org/sid/ (accessed on 17 July 2014).
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global patterns in plant height. J. Ecol. 2009, 97, 923–932. [Google Scholar] [CrossRef]
- USDA, NRCS. The PLANTS Database; National Plant Data Team: Greensboro, NC, USA. Available online: http://plants.usda.gov (accessed on 6 August 2014).
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from multiple traits, and other tools for functional ecology. R package version 1.0–1.2. 2014. Available online: https://cran.r-project.org/web/packages/FD/FD.pdf (accessed on 14 October 2014).
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Mason, N.W.H.; De Bello, F.; Mouillot, D.; Pavoine, S.; Dray, S. A guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients. J. Veg. Sci. 2013, 24, 794–806. [Google Scholar] [CrossRef]
- Swenson, N.G. Functional and Phylogenetic Ecology in R; Springer-Verlag: New York, NY, USA, 2014. [Google Scholar]
- Kline, R.B. Principles and Practice of Structural Equation Modeling, 2nd ed.; The Guilford Press: New York, NY, USA, 2005. [Google Scholar]
- Denslow, J.S. Disturbance and Diversity in Tropical Rain Forests: The Density Effect. Ecol. Appl. 1995, 5, 962–968. [Google Scholar] [CrossRef]
- Hu, L.T.; Bentler, P.M. Cutoff criteria for fit indexes in covariance structure analysis: Conventional criteria versus new alternatives. Struct. Equ. Model. 1999, 6, 1–55. [Google Scholar] [CrossRef]
- Rosseel, Y. lavaan: An R Package for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Stahl, U.; Kattge, J.; Reu, B.; Voigt, W.; Ogle, K.; Dickie, J.; Wirth, C. Whole-plant trait spectra of North American woody plant species reflect fundamental ecological strategies. Ecosphere 2013, 4, 128. [Google Scholar] [CrossRef]
- Barbour, M.G.; Burk, J.H.; Pitts, W.D.; Gilliam, F.S.; Schwartz, M.W. Terrestrial Plant Ecology, 3rd ed.; Bejamin Cummings: Menlo Park, CA, USA, 1999. [Google Scholar]
- Finzi, A.C.; Canham, C.D.; van Breemen, N. Canopy Tree-Soil Interactions within Temperate Forests: Species Effects on pH and Cations. Ecol. Appl. 1998, 8, 447–454. [Google Scholar]
- Alban, D.H. Effects of Nutrient Accumulation by Aspen, Spruce, and Pine on Soil Properties. Soil Sci. Soc. Am. J. 1982, 46, 853–861. [Google Scholar] [CrossRef]
- McFee, W.W.; Kelly, J.M.; Beck, R.H. Acid precipitation effects on soil pH and base saturation of exchange sites. Water Air Soil Pollut. 1977, 7, 401–408. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity Metric | χ2 | df | p | RMSEA | SRMR | CFI | R2 |
|---|---|---|---|---|---|---|---|
| FRic | 12.298 | 11 | 0.342 | 0.038 | 0.074 | 0.998 | 0.161 |
| FDis | 11.796 | 14 | 0.623 | 0.000 | 0.054 | 1.000 | 0.239 |
| CWM WD | 39.075 | 21 | 0.010 | 0.104 | 0.087 | 0.950 | 0.426 |
| CWM SLA | 35.909 | 19 | 0.011 | 0.105 | 0.086 | 0.982 | 0.705 |
| CWM Leaf N | 39.843 | 20 | 0.005 | 0.111 | 0.078 | 0.978 | 0.466 |
| CWM Max. Ht. | 10.362 | 10 | 0.409 | 0.021 | 0.073 | 0.999 | 0.603 |
| CWM Seed Mass | 45.491 | 28 | 0.020 | 0.088 | 0.077 | 0.954 | 0.519 |
| Predictor | Pathway | FRic | FDis | ||
|---|---|---|---|---|---|
| Coeff | p | Coeff | p | ||
| Elevation (Linear) | Direct | −0.338 | 0.001 | - | - |
| Indirect through Soil Moisture | - | - | −0.135 | 0.007 | |
| Indirect through Soil Moisture and Soil Fertility | 0.033 | 0.055 | - | - | |
| Indirect through Soil Fertility | −3.53 | 0.006 | - | - | |
| Indirect through Soil P and Stem Density | - | - | −0.020 | 0.130 | |
| Total | −3.835 | 0.003 | −0.155 | 0.003 | |
| Elevation (Quadratic) | Indirect through Soil Fertility | 3.572 | 0.006 | - | - |
| Aspect (Linear) | Indirect through Soil Moisture | – | – | −0.076 | 0.035 |
| Indirect through Soil Moisture and Soil Fertility | 0.019 | 0.094 | - | - | |
| Indirect through Soil P and Stem Density | - | - | −0.018 | 0.148 | |
| Indirect through Soil Fertility | 0.085 | 0.024 | - | - | |
| Indirect through Soil Mn | - | - | −0.065 | 0.080 | |
| Total | 0.104 | 0.015 | −0.159 | 0.002 | |
| Slope | Indirect through Soil Moisture | - | - | −0.140 | 0.006 |
| Indirect through Soil Moisture and Soil Fertility | 0.034 | 0.054 | - | - | |
| Indirect through Soil P and Stem Density | - | - | 0.022 | 0.114 | |
| Total | 0.034 | 0.054 | −0.118 | 0.027 | |
| Stem Density | Direct | - | - | 0.253 | 0.010 |
| Soil Moisture | Direct | - | - | 0.314 | 0.001 |
| Indirect through Soil Fertility | −0.078 | 0.039 | - | - | |
| Soil Fertility | Direct | −0.308 | 0.003 | - | - |
| Indirect through Stem Density | - | - | 0.088 | 0.042 | |
| Soil Mn | Direct | - | - | 0.248 | 0.011 |
| Predictor | Pathway | WD | SLA | Max. Ht. | Leaf N | Seed Mass | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Coeff | p | Coeff | p | Coeff | p | Coeff | p | Coeff | p | ||
| Elevation (Linear) | Direct | 0.247 | 0.011 | 4.372 | 0.005 | −0.485 | 0.000 | 5.856 | 0.004 | 0.382 | 0.000 |
| Indirect through Soil Moisture | 0.159 | 0.002 | −0.104 | 0.005 | - | - | - | - | 0.133 | 0.003 | |
| Indirect through Soil Moisture and Soil Fertility | - | - | −0.056 | 0.019 | −0.054 | 0.018 | −0.045 | 0.032 | - | - | |
| Indirect through Soil Fertility | - | - | 6.003 | 0.000 | 5.738 | 0.000 | 4.819 | 0.000 | - | - | |
| Indirect through Soil Zn | 0.060 | 0.068 | −0.081 | 0.029 | - | - | −0.069 | 0.051 | 0.041 | 0.112 | |
| Indirect through Soil P and Stem Density | 0.014 | 0.168 | – | – | – | – | – | – | 0.015 | 0.139 | |
| Total | 0.480 | 0.000 | 10.13 | 0.000 | 5.199 | 0.000 | 10.56 | 0.000 | 0.572 | 0.000 | |
| Elevation (Quadratic) | Direct | - | - | −4.314 | 0006 | - | - | −5.787 | 0.005 | - | - |
| Indirect through Soil Fertility | - | - | −6.075 | 0.000 | −5.807 | 0.000 | −4.877 | 0.000 | - | - | |
| Total | - | - | −10.39 | 0.000 | −5.807 | 0.000 | −10.66 | 0.000 | - | - | |
| Aspect (Linear) | Indirect through Soil Moisture | 0.090 | 0.020 | −0.059 | 0.030 | - | - | - | - | 0.075 | 0.025 |
| Indirect through Soil Moisture and Soil Fertility | - | - | −0.032 | 0.053 | −0.030 | 0.051 | −0.025 | 0.069 | - | - | |
| Indirect through Soil P and Stem Density | 0.013 | 0.182 | - | - | - | - | - | - | 0.014 | 0.154 | |
| Indirect through Soil Fertility | - | - | −0.145 | 0.002 | −0.139 | 0.002 | −0.117 | 0.008 | - | - | |
| Indirect through Soil Zn | 0.312 | 0.042 | −0.420 | 0.010 | - | - | −0.359 | 0.027 | 0.215 | 0.085 | |
| Indirect through Soil Mn | - | - | - | - | - | - | - | - | 0.056 | 0.069 | |
| Total | 0.414 | 0.009 | −0.656 | 0.000 | −0.169 | 0.000 | −0.501 | 0.003 | 0.359 | 0.007 | |
| Aspect (Quadratic) | Indirect through Soil Zn | −0.341 | 0.034 | 0.460 | 0.006 | - | - | 0.393 | 0.020 | −0.235 | 0.076 |
| Slope | Direct | - | - | - | - | 0.146 | 0.040 | - | - | - | - |
| Indirect through Soil Moisture | 0.165 | 0.001 | −0.108 | 0.005 | - | - | - | - | 0.138 | 0.003 | |
| Indirect through Soil Moisture and Soil Fertility | - | - | −0.059 | 0.018 | −0.056 | 0.017 | −0.047 | 0.031 | - | - | |
| Indirect through Soil P and Stem Density | −0.015 | 0.153 | - | - | - | - | - | - | −0.017 | 0.123 | |
| Total | 0.150 | 0.005 | −0.167 | 0.001 | 0.090 | 0.223 | −0.047 | 0.031 | 0.121 | 0.010 | |
| Stem Density | Direct | −0.174 | 0.041 | - | - | - | - | - | - | −0.189 | 0.015 |
| Soil Moisture | Direct | −0.374 | 0.000 | 0.245 | 0.001 | - | - | - | - | −0.312 | 0.000 |
| Indirect through Soil Fertility | - | - | 0.132 | 0.008 | 0.127 | 0.007 | 0.106 | 0.018 | - | - | |
| Total | −0.374 | 0.000 | 0.377 | 0.000 | 0.127 | 0.007 | 0.106 | 0.018 | - | - | |
| Soil Fertility | Direct | - | - | 0.524 | 0.000 | 0.501 | 0.000 | 0.421 | 0.000 | - | - |
| Soil Zn | Direct | 0.247 | 0.005 | −0.333 | 0.000 | - | - | −0.285 | 0.001 | 0.170 | 0.033 |
| Soil P | Indirect through Stem Density | −0.059 | 0.083 | - | - | - | - | - | - | −0.065 | 0.052 |
| Soil Mn | Direct | - | - | - | - | - | - | - | - | −0.214 | 0.006 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chapman, J.I.; McEwan, R.W. The Role of Environmental Filtering in Structuring Appalachian Tree Communities: Topographic Influences on Functional Diversity Are Mediated through Soil Characteristics. Forests 2018, 9, 19. https://doi.org/10.3390/f9010019
Chapman JI, McEwan RW. The Role of Environmental Filtering in Structuring Appalachian Tree Communities: Topographic Influences on Functional Diversity Are Mediated through Soil Characteristics. Forests. 2018; 9(1):19. https://doi.org/10.3390/f9010019
Chicago/Turabian StyleChapman, Julia I., and Ryan W. McEwan. 2018. "The Role of Environmental Filtering in Structuring Appalachian Tree Communities: Topographic Influences on Functional Diversity Are Mediated through Soil Characteristics" Forests 9, no. 1: 19. https://doi.org/10.3390/f9010019