The Effect of Silver and Copper Nanoparticles on the Condition of English Oak (Quercus robur L.) Seedlings in a Container Nursery Experiment

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design, Plant Material, Soil Substrate and Sampling Scheme
2.2. Characteristics of Nano-Silver and Nano-Copper Colloids
2.3. Biometric Parameters and Powdery Mildew Assessment
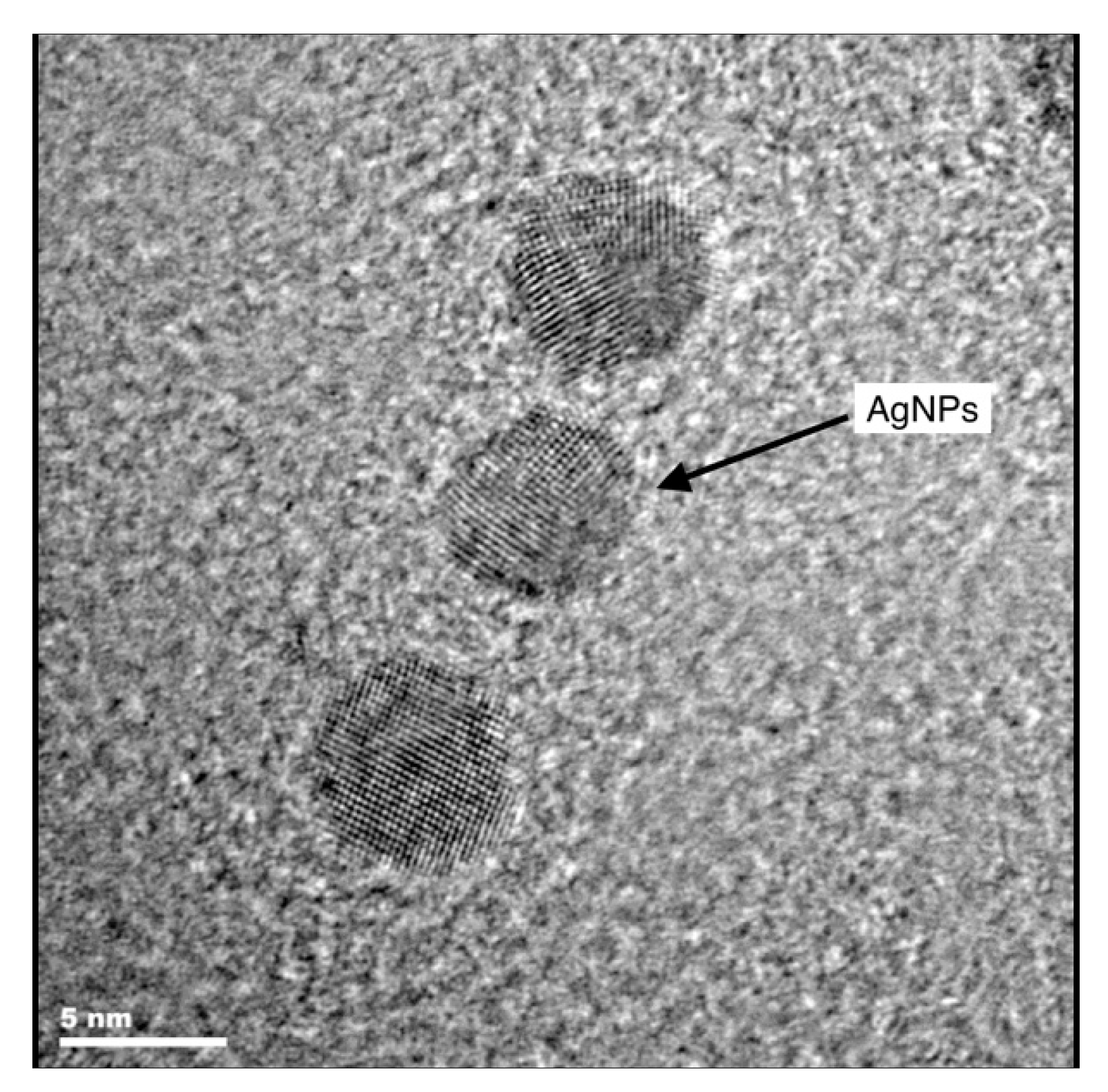
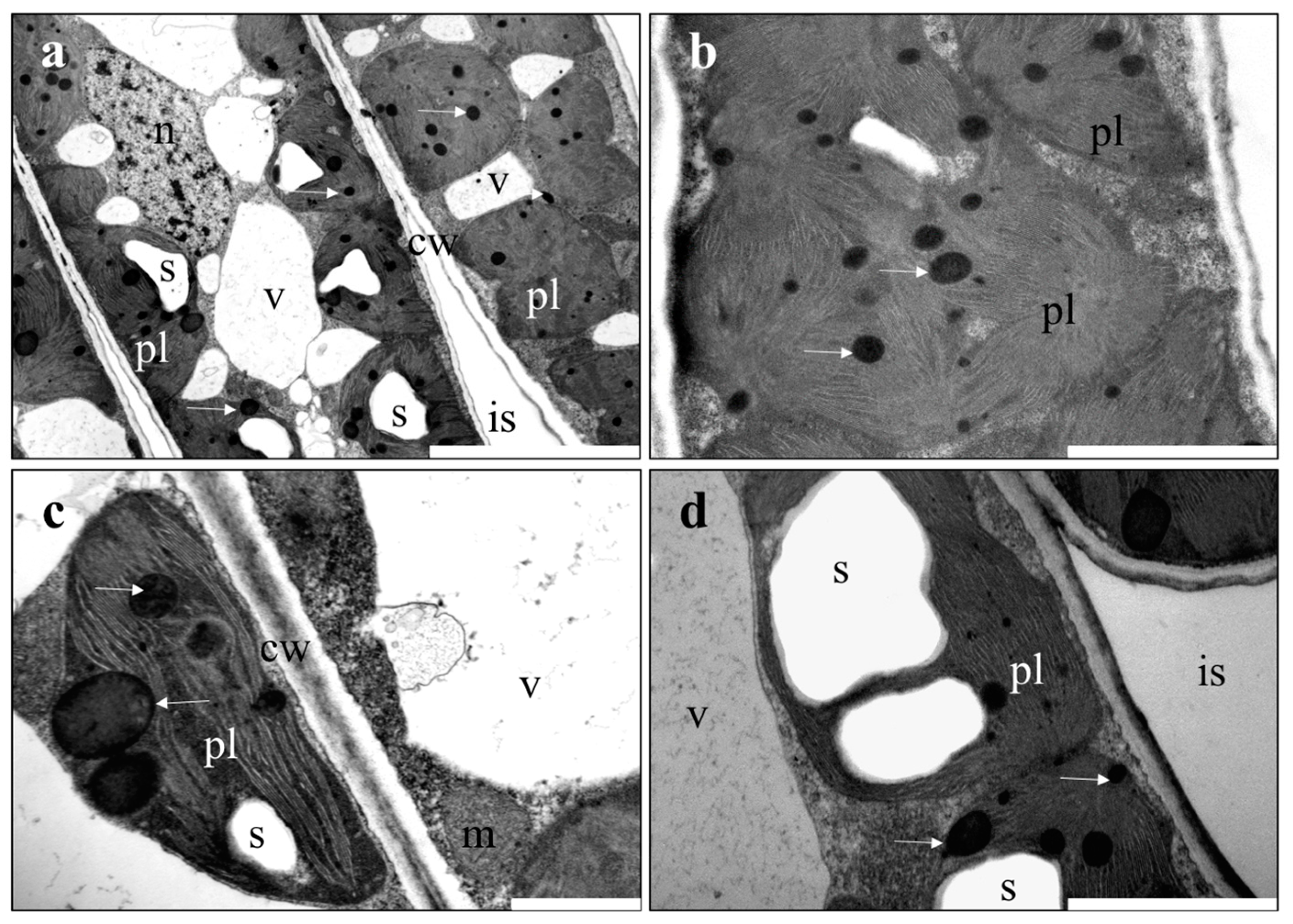
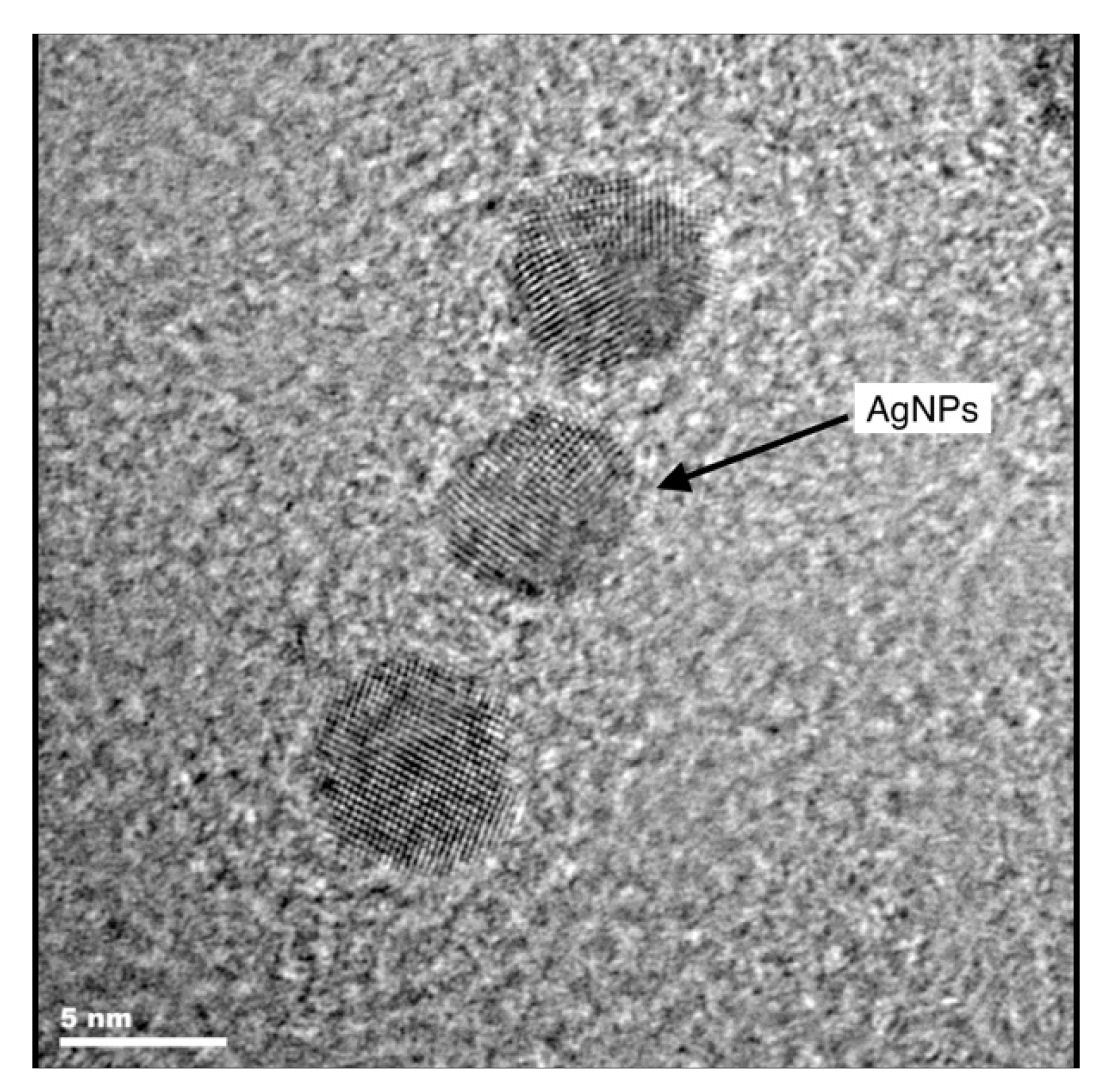
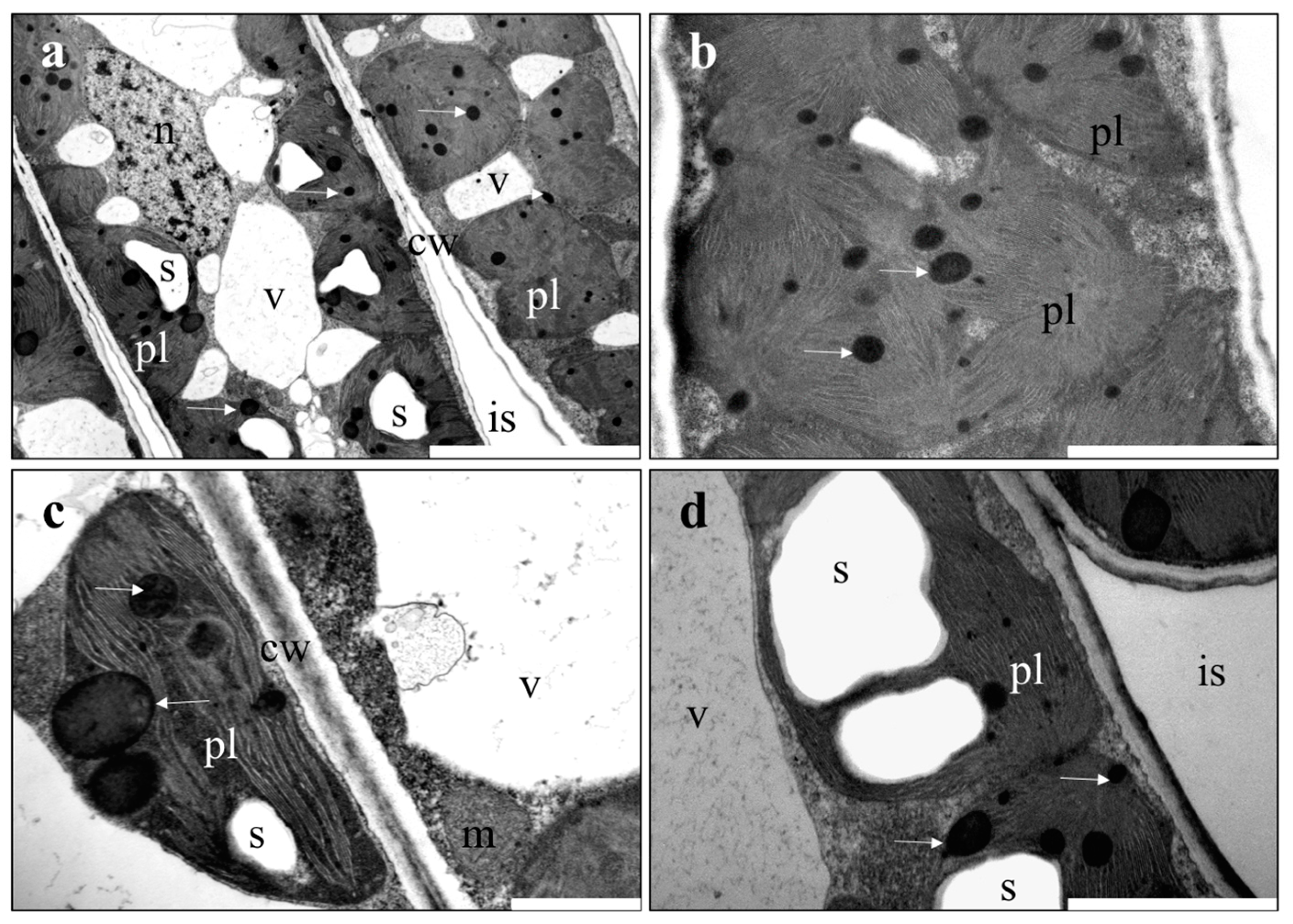
2.4. Microscopic Investigations
2.5. Ectomycorrhizae Assessment
2.6. Data Analysis
3. Results
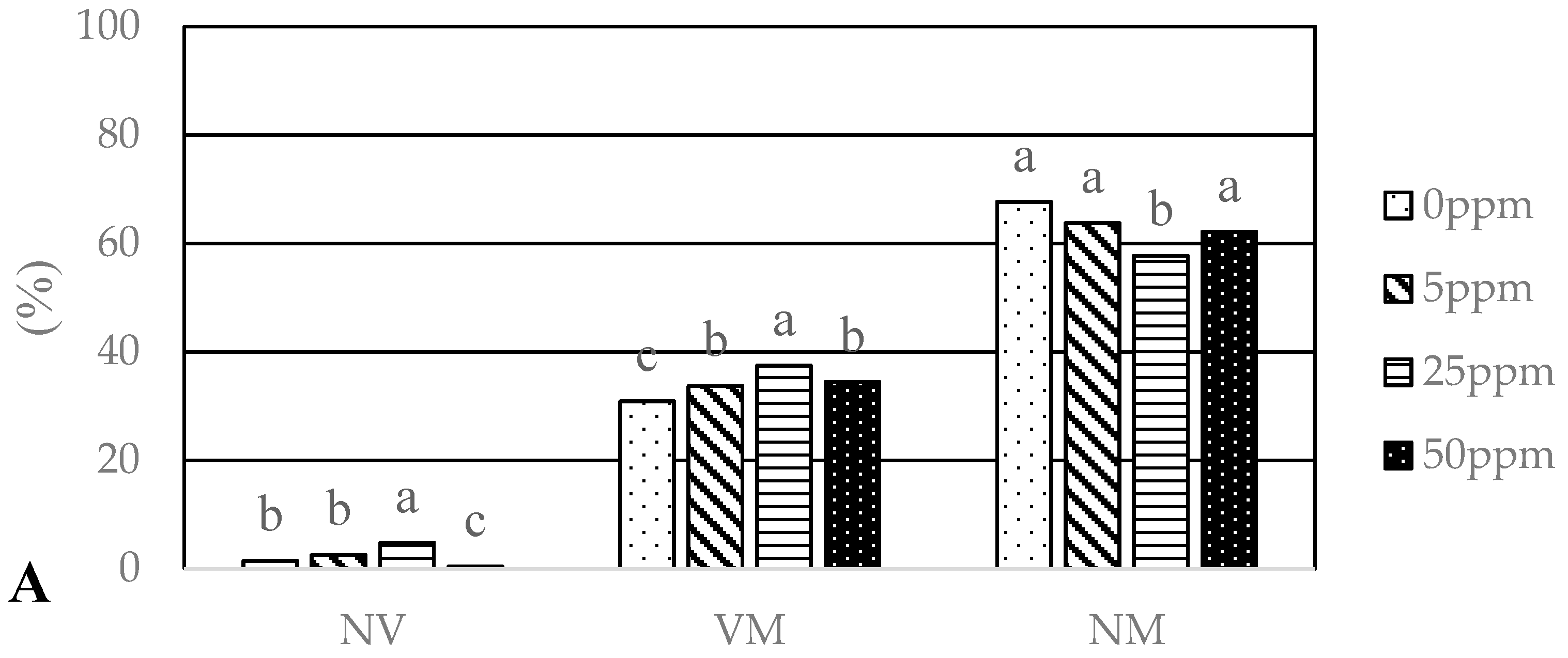
3.1. Growth Parameters and Powdery Mildew
3.2. Microscopic Investigations of Plant Material
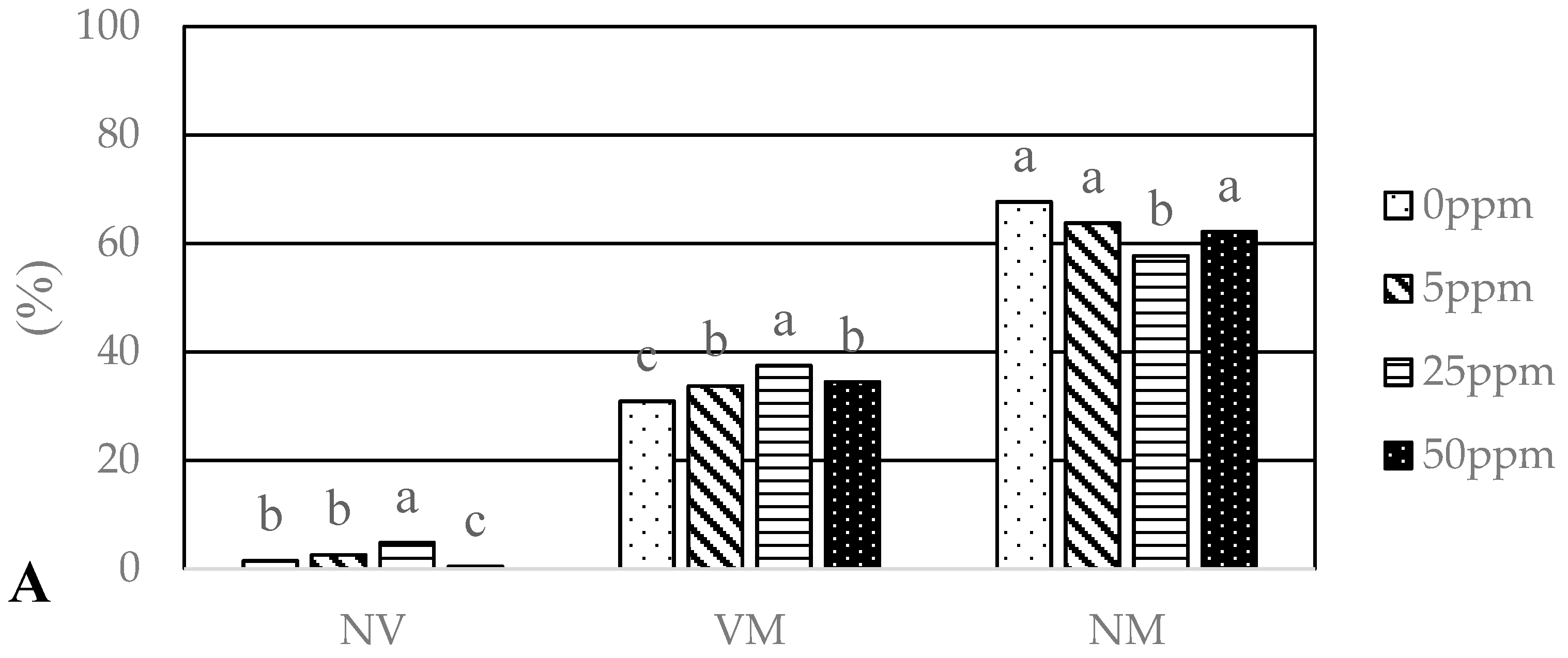
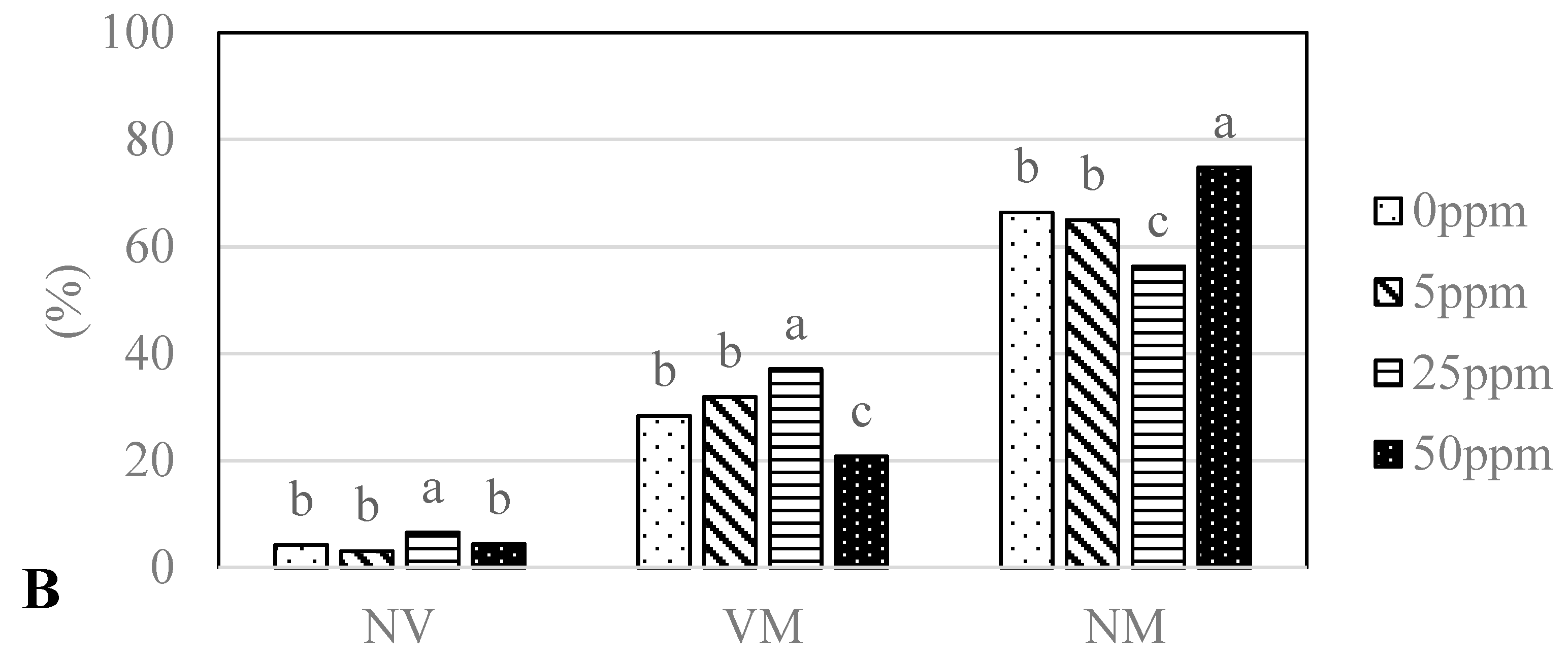
3.3. Mycorrhizal Colonization
3.4. Mycorrhizal Fungal Communities
4. Discussion
4.1. The Effect of Foliar Applications of AgNPs and CuNPs on Biometric Parameters of Oak Seedlings and on Powdery Mildew Disease
4.2. Ultrastructure of Organs of Plants Treated with NPs
4.3. Mycorrhizal Colonization
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ball, P. Natural strategies for the molecular engineer. Nanotechnology 2002, 13, 15–28. [Google Scholar] [CrossRef]
- Roco, M.C. Broader societal issue of nanotechnology. J. Nanopart. Res. 2003, 5, 181–189. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Madler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Ciancio, A.; Mukerji, K.G. General Concepts in Integrated Pest and Disease Management; Springer: Dordrecht, The Netherlands, 2007; pp. 27–43. [Google Scholar]
- Smita, S.; Gupta, S.K.; Bartonova, A.; Dusinska, M.; Gutleb, A.C.; Rahman, Q. Nanoparticles in the environment: Assessment using the causal diagram approach. Environ. Health 2012, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Han, J.; Xiao, J.Q.; Jin, Y. Uptake, translocation and accumulation of manufactured iron oxide nanoparticles by pumpkin plants. J. Environ. Monit. 2008, 10, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Husen, A.; Siddiqi, K.S. Phytosynthesis of nanoparticles: Concept, controversy and application. Nanoscale Res. Lett. 2014. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.S.; Diao, S.Q.; Li, C.H.; Fang, L.J.; Qiao, S.C.; Yu, M. Effect of TMS (nanostructured silicon dioxide) on growth of Changbai larch seedlings. J. For. Res. 2004, 15, 138–140. [Google Scholar]
- Lin, D.; Xing, B. Phytotoxicity of nanoparticles: Inhibition of seed germination and root growth. Environ. Pollut. 2007, 150, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Geiser-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between engineered nanoparticles (ENPs) and plants: Phytotoxicity, uptake and accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Lian, K.; Wu, Q.; Li, Y.; Danzy, M.; Menard, R.; Chin, K.L.; Collins, D.; Oliveria, F.; Klepzig, K. Nanotechnology application in forest protection. In Proceedings of the TAPPI International Conference on Nanotechnology for Renewable Materials, Washington, DC, USA, 6–8 June 2011; pp. 271–301. [Google Scholar]
- Lee, W.; An, Y.; Yoon, H.; Kweon, H. Toxicity and bioavailability of copper nanoparticles to the terrestrial plants mung bean (Phaseolus radiatus) and wheat (Triticum aestivum): Plant uptake for water insoluble nanoparticles. Environ. Toxicol. Chem. 2008, 27, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.J.; Singleton, I. Soil contamination with silver nanoparticles reduces Bishop pine growth and ectomycorrhizal diversity on pine roots. J. Nanopart. Res. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostami, A.A.; Shahsavar, A. Nano-silver particles eliminate the in vitro contaminations of olive ‘mission’ explants. Asian J. Plant Sci. 2009, 8, 505–509. [Google Scholar] [CrossRef]
- Seeger, E.M.; Baun, A.; Kästner, M.; Trapp, S. Insignificant acute toxicity of TiO2 nanoparticles to willow trees. J. Soil. Sediment 2009, 9, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Cui, X.; He, S.; Dong, G.; Chen, M.; Wang, J. The role of metal nanoparticles in influencing arbuscular mycorrhizal fungi effects on plant growth. Environ. Sci. Technol. 2013, 47, 9496–9504. [Google Scholar] [CrossRef] [PubMed]
- El-Temsah, Y.S.; Joner, E.J. Impact of Fe and Ag nanoparticles on seed germination and differences in bioavailability during exposure in aqueous suspension and soil. Environ. Toxicol. 2012, 27, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Savithramma, N.; Ankanna, S.; Bhumi, G. Effect of nanoparticles on seed germination and seedling growth of Boswellia ovalifoliolata an endemic and endangered medicinal tree taxon. Nano Vis. 2012, 2, 61–68. [Google Scholar]
- Stampoulis, D.; Sinha, S.K.; White, J.C. Assay-dependent phytotoxicity of nanoparticles to plants. Environ. Sci. Technol. 2009, 43, 9473–9479. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.O.; McLean, J.E.; Latta, D.E.; Manangón, E.; Britt, D.W.; Johnson, W.P.; Boyanov, M.I.; Anderson, A.J. CuO and ZnO nanoparticles: Phytotoxicity, metal speciation, and induction of oxidative stress in sand-grown wheat. J. Nano Res. 2012, 14, 1–15. [Google Scholar] [CrossRef]
- Lilja, A.; Poteri, M. Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CBA International: Wallingford, UK, 2013; pp. 567–592. [Google Scholar]
- Ouda, S.M. Antifungal Activity of Silver and Copper Nanoparticles on Two Plant Pathogens, Alternaria alternata and Botrytis cinerea. Res. J. Microbiol. 2014, 9, 34–42. [Google Scholar] [CrossRef]
- Gorczyca, A.; Pociecha, E.; Kasprowicz, M.; Niemiec, M. Effect of nanosilver in wheat seedlings and Fusarium culmorum culture systems. Eur. J. Plant Pathol. 2015, 142, 251–261. [Google Scholar] [CrossRef]
- Ashrafi, S.J.; Rastegar, M.F.; Nekouei, M.K. Use of silver Nano particle for controlling the powdery mildew diseases at Celastrales plants. Am. Eurasian J. Agric. Environ. Sci. 2010, 7, 70–74. [Google Scholar]
- Lamsal, K.; Kim, S.W.; Jung, J.H.; Kim, Y.S.; Kim, K.S.; Lee, Y.S. Inhibition Effects of Silver Nanoparticles against Powdery Mildews on Cucumberand Pumpkin. Mycobiology 2011, 39, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, UK, 2008. [Google Scholar]
- Marin, M. Effects of pesticides on the growth of ectomycorrhizal fungi and ectomycorrhiza formation. In Diversity and Biotechnology of Ectomycorrhizae; Rai, M., Varma, A., Eds.; Springer: Berlin, Germany, 2010; pp. 323–346. [Google Scholar]
- Scholl, L.; Hoffland, E.; van Breemen, N. Organic anion exudation by ectomycorrhizal fungi and Pinus sylvestris in response to nutrient deficiencies. New Phytol. 2006, 170, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Scholl, L.; Kuyper, T.W.; Smits, M.M.; Landeweert, R.; Hoffland, E.; van Breemen, N. Rock-eating mycorrhizas: Their role in plant nutrition and biogeochemical cycles. Plant Soil 2008, 303, 35–47. [Google Scholar] [CrossRef]
- Finlay, R.D. Ecological aspects of mycorrhizal symbiosis: With special emphasis on the functional diversity of interactions involving the extraradical mycelium. J. Exp. Bot. 2008, 59, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Finlay, R.D.; Söderström, B. Mycorrhiza and carbon flow to soil. In Mycorrhizal Functioning; Allen, M.F., Ed.; Chapman and Hall: London, UK, 1992; pp. 134–160. [Google Scholar]
- Colpaert, J.V. Heavy metal pollution and genetic adaptations in ectomycorrhizal fungi. In Stress in Yeasts and Filamentous Fungi; Avery, S., Stratford, M., van West, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 157–173. [Google Scholar]
- Finlay, R.D.; Lindahl, B.D.; Taylor, A.F.S. Responses of mycorrhizal fungi to stress. In Stress in Yeasts and Filamentous Fungi; Avery, S., Stratford, M., van West, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 201–220. [Google Scholar]
- Izumi, H.; Anderson, I.C.; Alexander, I.J.; Killam, K.; Moore, E.R.B. Diversity and expression of nitrogenase genes (nifH) from ectomycorrhizas of Corsican pine (Pinus nigra). Environ. Microbiol. 2006, 8, 2224–2230. [Google Scholar] [CrossRef] [PubMed]
- Power, S.A.; Ashmore, M.R. Nutrient relations and root mycorrhizal status of healthy and declining beech (Fagus sylvatica L) in southern Britain. Water Air Soil Pollut. 1996, 86, 317–333. [Google Scholar] [CrossRef]
- Kovacs, G.; Pausch, M.; Urban, A. Diversity of ectomycorrhizal morphotypes and oak decline. Phyton Ann. Rei Bot. A 2000, 40, 109–116. [Google Scholar]
- Scattolin, L.; Dal Maso, E.; Mutto Accord, S.; Sella, L.; Montecchio, L. Detecting asymptomatic ink-diseased chestnut trees by the composition of the ectomycorrhizal community. For. Pathol. 2012, 42, 501–509. [Google Scholar] [CrossRef]
- Manceau, A.; Nagy, K.L.; Marcus, M.A.; Lanson, M.; Geoffroy, N.; Jacquet, T.; Kirpichtchikova, T. Formation of metallic copper nanoparticles at the soil-root interface. Environ. Sci. Technol. 2008, 42, 1766–1772. [Google Scholar] [CrossRef] [PubMed]
- Dubchak, S.; Ogar, A.; Mietelski, J.W.; Turnau, K. Influence of silver and titanium nanoparticles on arbuscular mycorrhiza colonization and accumulation of radiocaesium in Helianthus annuus. Span. J. Agric. Res. 2010, 8, 103–108. [Google Scholar] [CrossRef]
- Judy, J.D.; Kirby, J.K.; Creamer, C.; McLaughlin, M.J.; Fiebiger, C.; Wright, C.; Cavagnaro, T.R.; Bertsch, P.M. Effects of silver sulfide nanomaterials on mycorrhizal colonization of tomato plants and soil microbial communities in biosolid-amended soil. Environ. Pollut. 2015, 206, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, S. Zasady postępowania technologicznego przy zastosowaniu polskiego biopreparatu z grzybem Hebeloma crustuliniforme w zabiegu sterowanej mikoryzacji sadzonek drzew leśnych. In Ektomikoryzy. Nowe Biotechnologie w Polskim Szkółkarstwie Leśnym; Kowalski, S., Ed.; CILP: Warszawa, Poland, 2007; pp. 101–103. [Google Scholar]
- Nanogrp.com. Available online: http://www.nanogrp.com/pl/ (accessed on 15 April 2017).
- Karnovsky, M.J. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell. Biol. 1965, 27, 1A–149A. [Google Scholar]
- Agerer, R. Colour Atlas of Ectomycorrhizae, 1st–15th ed.; Einhorn: Munich, Germany, 1987–2008. [Google Scholar]
- Agerer, R. Exploration types of ectomycorrhizae. A proposal to classify ectomycorrhizal mycelial systems according to their patterns of differentiation and putative ecological importance. Mycorrhiza 2001, 11, 107–114. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Iotti, M.; Zambonelli, A. A quick and precise technique for identifying ectomycorrhizas by PCR. Mycol. Res. 2006, 110, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B. Vegan: Community Ecology Package. R Package Version 2.4-2. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 24 August 2017).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Khot, L.R.; Sankaran, S.; Maja, J.M.; Ehsani, R.; Schuster, E.W. Applications of nanomaterials in agricultural production and crop protection. Crop. Prot. 2012, 35, 64–70. [Google Scholar] [CrossRef]
- Zhang, Z.; Kong, F.; Vardhanabhuti, B.; Mustapha, A.; Lin, M. Detection of engineered silver nanoparticle contamination in pears. J. Agric. Food Chem. 2012, 60, 10762–10767. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Mukherjee, A.; Chandrasekaran, N. Genotoxicity of silver nanoparticles in Allium cepa L. Sci. Total Environ. 2010, 407, 5243–5246. [Google Scholar] [CrossRef] [PubMed]
- Battke, F.; Leopold, K.; Maier, M.; Schmidhalter, U.; Schuster, M. Palladium exposure of barley: Uptake and effects. Plant. Biol. 2009, 10, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.T.; Bali, R. On the formation and extent of uptake of silver nanoparticles by live plants. J. Nanopart. Res. 2008, 10, 691–695. [Google Scholar] [CrossRef]
- Hong, J.; Peralta-Videa, J.R.; Rico, C.; Sahi, S.; Viveros, M.N.; Bartonjo, J.; Zhao, L.; Gardea-Torresdey, J.L. Evidence of translocation and physiological impacts of foliar applied CeO2 nanoparticles on cucumber (Cucumis sativus) plants. Environ. Sci. Technol. 2014, 48, 4376–4385. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Wagner, B.; Odzak, N.; Sigg, L.; Behra, R. Effects of Differently Coated Silver Nanoparticles on the Photosynthesis of Chlamydomonas reinhardtii. Environ. Sci. Technol. 2015, 49, 8041–8047. [Google Scholar] [CrossRef] [PubMed]
- Lalau, C.M.; de Almeida Mohedano, R.; Schmidt, E.C.; Bouzon, Z.; Ouroques, L.C.; dos Santos, R.; da Costa, C.H.; Vicentini, D.S.; Matia, W.G. Toxicological effects of copper oxide nanoparticles on the growth rate, photosynthetic pigment content, and cell morphology of the duckweed Landoltia punctata. Protoplasma 2014, 252, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Selga, T.; Selga, M. Response of Pinus silvestris L. needles to electromagnetic fields. Cytological and ultrastructural aspects. Sci. Total Environ. 1996, 180, 65–73. [Google Scholar] [CrossRef]
- Piller, L.E.; Abraham, M.; Dormann, P.; Kessler, F.; Besagni, C. Plastid lipid droplets at the crossroads of prenylquinone metabolism. J. Exp. Bot. 2012, 63, 1609–1618. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Tarafdar, J.C.; Biswa, S.P. Nanoparticle synthesis and delivery by an aerosol route for water melon plant foliar uptake. J. Nanopart. Res. 2013. [Google Scholar] [CrossRef]
- Birbaum, K.; Brogioli, R.; Schellenberg, M.; Martinoia, E.; Stark, W.J.; Gunter, D.; Limbach, L.K. No evidence for cerium dioxide NP translocation in maize plants. Environ. Sci. Technol. 2010, 44, 8718–8723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Qiu, Y.L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Myzorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Corcobado, T.; Vivas, M.; Moreno, G.; Solla, A. Ectomycorrhizal symbiosis in declining and non-declining Quercus ilex trees infected with or free of Phytophthora Cinnamomi. Forest Ecol. Manag. 2014, 324, 72–80. [Google Scholar] [CrossRef]
- Kuikka, K.; Härmä, E.; Markkola, A.; Rautio, P.; Roitto, M.; Saikkonen, K.; Ahonen-Jonnarth, U.; Finlay, R.; Tuomi, J. Severe defoliation of Scots pine reduces reproductive investment by ectomycorrhizal symbionts. Ecology 2003, 84, 2051–2061. [Google Scholar] [CrossRef]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.; Stenlid, J.; Finlay, R. Fungal communities in mycorrhizal roots of conifer seedlings in forest nurseries under different cultivation systems, assessed by morphotyping, direct sequencing and mycelial isolation. Mycorrhiza 2005, 16, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Abbasian, M.; Kashani, A.; Ardakani, M.R.; Rejali, F.; Timajchi, M.; Seifi, S.M.; Mafakheri, S. The effects of chemical, biological and nano fungicides on mycorrhizal colonization and quality of sunflower. Ann. Biol. Res. 2012, 3, 4239–4245. [Google Scholar]
- Urban, A. Metal elements and the diversity and function of ectomycorrhizal communities. In Diversity and Biotechnology of Ectomycorrhiza; Rai, M., Varma, A., Eds.; Springer: Berlin, Germany, 2011; Volume 25, pp. 231–254. [Google Scholar]
- Yamasaki, H.; Pilon, M.; Shikanai, T. How do plants respond to copper deficiency? Plant Signal Behav. 2008, 3, 231–232. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.H.; Bradshaw, A.D. A comparison of the toxicity of heavy metals, using root elongation of rye grass, Lolium perenne. New Phytol. 1982, 91, 255–261. [Google Scholar] [CrossRef]
- Arduini, I.; Godbold, D.L.; Onnis, A. Influence of copper on root growth and morphology of Pinus pinea L. and Pinus pinaster Ait. seedlings. Tree Physiol. 1995, 15, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Trappe, J.M.; Molina, R.; Castellano, M. Reactions of mycorrhizal fungi and mycorrhiza formation to pesticides. Annu. Rev. Phytopathol. 1984, 22, 331–359. [Google Scholar] [CrossRef]
- Gherghel, F.; Krause, K. Role of mycorrhiza in re-forestation at heavy metal-contaminated sites. In Bio-Geo Interactions in Metal-Contaminated Soils; Kothe, E., Varma, A., Eds.; Springer: Berlin, Germany, 2012; Volume 31, pp. 183–199. [Google Scholar]
- Sánchez, S.; Gómez, E.; Martín, M.; De Miguel, A.M.; Urban, A.; Barriuso, J. Experiments on the life cycle and factors affecting reproduction of Sphaerosporella brunnea provide evidence for rapid asexual propagation by conidiospores and for homothallism in an ectomycorrhizal competitor of cultivated truffle species. Fungal Ecol. 2014, 8, 59–65. [Google Scholar] [CrossRef]
- Baum, C.; Schmid, K.; Makeschin, F. Interactive effects of substrates and ectomycorrhizal colonization on growth of a poplar clone. J. Plant Nutr. Soil Sci. 2000, 163, 221–226. [Google Scholar] [CrossRef]
- Baum, C.; Hrynkiewicz, K.; Leinweber, P.; Meißner, R. Heavy-metal mobilization and uptake by mycorrhizal and nonmycorrhizal willows (Salix × dasyclados). J. Plant Nutr. Soil Sci. 2006, 169, 516–522. [Google Scholar] [CrossRef]
- Urban, A.; Puschenreiter, M.; Strauss, J.; Gorfer, M. Diversity and structure of ectomycorrhizal and co-associated fungal communities in a serpentine soil. Mycorrhiza 2008, 18, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Marx, D.H.; Cordell, C.E.; France, R.C. Effects of triadimefon on growth and ectomycorrhizal development of loblolly and slash pines in nurseries. Phytopathology 1986, 76, 824–831. [Google Scholar] [CrossRef]
- Deng, Y.; White, J.C.; Xing, B. Interactions between Engineered Nanomaterials and Agricultural Crops: Implications for Food Safety. J. Zhejiang Univ. Sci. A 2014, 15, 552–572. [Google Scholar] [CrossRef]
- Pacheco, I.; Bueza, C. Nanoparticle Interaction with Plants. In Nanoscience and Plant–Soil Systems; Ghorbanpour, M., Manika, K., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; Volume 48, pp. 323–355. [Google Scholar]
- Schreck, E.; Foucault, Y.; Sarret, G.; Sobanska, S.; C´ecillon, L.; Castrec-Rouelle, M.; Uzu, G.; Dumat, C. Metal and metalloid foliar uptake by various plant species exposed to atmospheric industrial fallout: Mechanisms involved for lead. Sci. Total Environ. 2012, 427, 253–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NPs and Woody Plants | ||||
| NPs | Concentration | Plant Species | Effect | References |
| Ag | 0, 350 and 790 mg kg−1 | Bishop pine | Inhibition of root growth of root and shoot biomass | [13] |
| 100–400 mg L−1 submerged | Mission olive explant | Severe injuries and browning of the explants | [14] | |
| SiO2 | 500 µL. L−1 | Changbai larch | Increased shoot height, root collar diameter, root length | [8] |
| TiO2 | 1–100 mg L−1 | Willow tree | No significant effects on transpiration rate, growth, and water use efficiency | [15] |
| CCCS | 4 g/tree | Bald cypress | No significant effects on the growth parameters of the seedling | [11] |
| NPs and non-woody plants | ||||
| Ag | 0.01 mg kg−1 | Clover | Reduced aboveground biomass | [16] |
| 10 mg L−1 | Barley | Reduced germination and shoot length | [17] | |
| 10–30 g mL−1 | Boswellia ovalifoliolata | Increased germination and seedling growth | [18] | |
| 500 and 1000 mg L−1 | Zucchini | Reduced biomass | [19] | |
| Cu | <200 mg L−1 | Mung bean, Wheat | Reduced root and seedling growth | [12] |
| 1000 mg L−1 | Zucchini | Reduced biomass and root growth | [19] | |
| CuO | 500 mg kg−1 | Common Wheat | Increased biomass | [20] |
| Treatment (n = 40) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Parameters | Ag0 | Ag5 | Ag25 | Ag50 | Cu0 | Cu5 | Cu25 | Cu50 |
| Length of shoot (cm) | 29.10 | 26.63 | 28.23 | 28.60 | 28.25 | 29.31 | 29.48 | 27.13 |
| ±9.69 | ±8.67 | ±7.91 | ±9.06 | ±8.79 | ±7.49 | ±7.98 | ±7.33 | |
| Root collar diameter (mm) | 5.78 | 5.43 | 5.43 | 5.72 | 5.42 | 5.77 | 5.62 | 5.76 |
| ±0.78 | ±0.91 | ±0.83 | ±0.87 | ±0.99 | ±0.87 | ±0.90 | ±0.83 | |
| Dry mass of shoot (g) | 1.62 | 1.44 | 1.41 | 1.56 | 1.50 | 1.56 | 1.61 | 1.51 |
| ±0.67 | ±0.63 | ±0.58 | ±0.72 | ±0.65 | ±0.56 | ±0.59 | ±0.52 | |
| Dry mass of root (g) | 3.27 | 3.40 | 3.21 | 3.41 | 3.13 | 2.93 | 3.26 | 3.13 |
| ±1.08 | ±1.23 | ±1.30 | ±1.06 | ±1.41 | ±1.08 | ±1.26 | ±1.05 | |
| Total root length (cm) | 397.07 | 384.69 | 403.94 | 386.00 | 375.00 | 406.83 | 432.98 | 421.56 |
| ±131.52 | ±76.28 | ±145.89 | ±140.23 | ±144.02 | ±142.24 | ±138.05 | ±126.30 | |
| Number of tips (n) | 917.85 | 1006.75 | 1027.15 | 893.45 | 856.90 | 973.48 | 1084.10 | 1001.30 |
| ±262.16 | ±235.19 | ±331.61 | ±309.25 | ±302.36 | ±304.43 | ±378.44 | ±298.07 | |
| Infestation index of powdery mildew | 2.69 | 2.60 | 2.71 | 2.59 | 2.78 | 2.74 | 2.64 | 2.80 |
| Treatment (n = 40) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Identification | Closest Match | ID with Accession No. | Identity (%) | Ag0 | Ag5 | Ag25 | Ag50 | Cu0 | Cu5 | Cu25 | Cu50 |
| Ascomycota | |||||||||||
| Sphaerosporella brunnea | Sphaerosporella brunnea | KC008076.1 | 98 | 45.0/54.0 | 57.5/7.0 | 67.5/65.6 | 62.5/68.9 | 45.0/79.1 | 47.5/68.9 | 60.0/59.2 | 52.5/74.5 |
| Basidiomycota | |||||||||||
| Thelephora terrestris | Thelephora terrestris | FJ809998.1 | 98 | 55.0/17.6 | 47.5/16.8 | 70.0/28.1 | 35.0/28.6 | 25.0/16.8 | 55.0/38.3 | 70.0/28.5 | 25.0/20.1 |
| Paxillus involutus | Paxillus involutus | EU078725.1 | 98 | -/- | 10.0/11.5 | 10.0/6.4 | 10.0/2.5 | -/- | 2.5/0.9 | 17.5/12.3 | 7.5/5.3 |
| Laccaria proxima | Laccaria proxima | GQ267477.1 | 99 | -/- | 10.0/3.5 | -/- | -/- | 10.0/4.7 | -/- | -/- | -/- |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olchowik, J.; Bzdyk, R.M.; Studnicki, M.; Bederska-Błaszczyk, M.; Urban, A.; Aleksandrowicz-Trzcińska, M. The Effect of Silver and Copper Nanoparticles on the Condition of English Oak (Quercus robur L.) Seedlings in a Container Nursery Experiment. Forests 2017, 8, 310. https://doi.org/10.3390/f8090310
Olchowik J, Bzdyk RM, Studnicki M, Bederska-Błaszczyk M, Urban A, Aleksandrowicz-Trzcińska M. The Effect of Silver and Copper Nanoparticles on the Condition of English Oak (Quercus robur L.) Seedlings in a Container Nursery Experiment. Forests. 2017; 8(9):310. https://doi.org/10.3390/f8090310
Chicago/Turabian StyleOlchowik, Jacek, Roman Mariusz Bzdyk, Marcin Studnicki, Magdalena Bederska-Błaszczyk, Alexander Urban, and Marta Aleksandrowicz-Trzcińska. 2017. "The Effect of Silver and Copper Nanoparticles on the Condition of English Oak (Quercus robur L.) Seedlings in a Container Nursery Experiment" Forests 8, no. 9: 310. https://doi.org/10.3390/f8090310