
Characteristics of Fine Roots of Pinus massoniana in the Three Gorges Reservoir Area, China
by
Yafei Shen
1,†, 
Na Wang
1,†,
Ruimei Cheng
1,2,*,
Wenfa Xiao
1,2,
Shao Yang
1,
Yan Guo
1,
Lei Lei
1,
Lixiong Zeng
1 and
Xiaorong Wang
1 1
Key Laboratory of Forest Ecology and Environment, State Forestry Administration; Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China
2
Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China
*
Author to whom correspondence should be addressed.
†
Y.S. and N.W. contributed equally to this study.
Forests 2017, 8(6), 183; https://doi.org/10.3390/f8060183
Submission received: 1 May 2017
/
Revised: 19 May 2017
/
Accepted: 22 May 2017
/
Published: 25 May 2017
(This article belongs to the Special Issue Forest Fine Roots in Changing Climate)
Abstract
:Several studies have focused on fine roots characteristics because they provide a major pathway for nutrient cycling and energy flow in forest ecosystems. However, few studies have evaluated changes in fine root characteristics according to their diameter. Pinus massoniana forests are the main vegetative component in the Three Gorges Reservoir area and play an important role in providing forest resources and ecological services. Pinus massoniana fine roots were sorted into 0–0.5, 0.5–1, and 1–2 mm diameter classes, and their fine root standing biomass (FRB), necromass, annual production and decomposition rates were determined and correlated with soil characteristics. These fine roots in three diameter classes significantly differed in their initial carbon (C), C/N ratio, FRB, necromass, annual C and N production and decomposition rate. The production and decomposition of these different diameter classes varied significantly with soil variables including soil temperature, moisture, calcium and ammonium concentration but the strength of these interactions varied dependent on diameter class. The very fine roots had a faster decomposition ratio than larger fine roots due to the lower N content, higher C/N ratio and higher sensitivity to soil environmental factors. These results clearly indicate heterogeneity among fine roots of different diameters, and these variations should be taken into account when studying fine root characteristics and their role in the C cycle.
1. Introduction
Forests are an important component of the global terrestrial ecosystem and play a key role in the carbon (C) cycle. They store approximately 80% of all terrestrial aboveground C and over 70% of all soil organic C [1,2,3]. Furthermore, within a root system, the roots of forest trees, especially fine roots (i.e., <2 mm in diameter), are a vital source of soil C [4,5,6] because of their shorter life span and higher metabolic activity than those of coarse roots [7,8,9]. Although fine roots constitute a very small proportion (<5%) of the total standing root biomass in many terrestrial ecosystems [10], it is estimated that fine root turnover represents up to 33% of total net primary production in most ecosystems [11]. Thus, fine root dynamics are widely recognized for essential understanding of their role as C stores and sources of soil litter input in terrestrial ecosystems.
Fine root decomposition provides a major pathway for nutrient cycling and energy flow in forest ecosystems [12,13]. Research shows that, compared to aboveground leaf litter, belowground fine roots are continuously growing, dying, and decomposing, which contributes to increase soil organic matter content, especially in deep soil, and provides a major route for ecosystem C and nutrient input [14,15]. Studies have also shown that 30–80% of forest soil organic C is derived from the turnover and decomposition of fine root biomass [13,16,17], and it is generally accepted that fine roots are components within the overall root system sensitive to rooting environmental variations [18,19]. For example, fine root responses to changes in soil moisture are diameter dependent [20] and the morphological characteristics and biomass of very fine roots (diameter < 0.5 mm) are affected differently by changes in soil warming [21] and soil Ca content [22]. It has been demonstrated that root life span is also affected by soil properties [23,24]. Considerable related research has been published on the biomass and traits of fine roots, especially very fine roots, in response to environmental characteristic and soil nutrient status in forest fields.
Most published literature is on the characteristics of fine roots in diameter class <2 mm in forests across different species, stand ages, and geo-graphic regions [16,25,26]. However, fine roots in diameter class <2 mm alone are not indicative of the functional potential of the fine root system because this system is composed of individual roots with heterogeneous physiological and morphological traits [27,28]. For example, Noguchi et al. [29] considered that very fine roots were crucial in studies of spruce stands growing on a shallow permafrost table because the higher biomass was allocated to very fine roots. Moreover, morphological characteristics and physiological functions differ significantly among fine roots of different sizes, and roots of different diameter classes showed significant differences in their respiration rates as root respiration rate decreased with root diameter increase [30]. In most tree species, mounting evidence shows that fine roots <2 mm diameter can be divided into smaller diameter classes, due to their wide range of variability in root functions, according to root diameter size class or root branching order [9,31,32,33]. Hence, the traditional category of fine roots as a single root diameter class of <2 mm might be inherently arbitrary for predicting variation in root characteristics [9,30,34,35]. In addition, given the difficult nature of long-term studies of root production and decomposition dynamics, it is especially unclear how these patterns will vary among diameter classes. Studying the characteristics of fine roots is challenging, because roots are hidden below the soil surface [36], and studies considering the underground root system are scarcer than those considering aboveground vegetation and its contribution to terrestrial C cycling. However, in the context of global environmental changes, variations in fine root production would affect forest soil nutrients, thereby affecting the patterns and processes of C allocation in the ecosystems [37,38,39]. Therefore, to better understand below-ground C cycling, we must evaluate the dynamics of fine roots in relation to their traits.
Plantations are an important component of global forest resources, and forest management therefore plays a key role in global C dynamics. The Three Gorges Reservoir Area of the Yangtze River is located in subtropical China and has been greatly affected by the Three Gorges Project, in which many plantations of indigenous tree species have been established to provide timber and enhance biodiversity and ecosystem services. Pinus massoniana forests play an important role in providing forest resources and ecological services and are the main vegetative component with the largest distribution area in the Three Gorges Reservoir area [40]. Therefore, studying the dynamics of the fine roots of P. massoniana provides an opportunity to better understand patterns of root ecological processes.
At present, there are numerous studies concerning the leaf litter decomposition rate of P. massoniana forests of different ages [40,41] while there are few studies considering fine roots with different diameters in the Three Gorges Reservoir Area. In this study, we quantified the dynamics of fine roots within three different diameter classes (0–0.5 mm; 0.5–1 mm; and 1–2 mm) [42,43], which are thought to play a key role in accurately predicting underground C storage and nutrient cycling in the P. massoniana forests of the Three Gorges Reservoir Area. Our objectives were to determine: (1) whether standing biomass, annual production, and decomposition rates differed among fine roots with different diameter classes; and (2) whether such differences were similarly or differentially affected by different soil factors including soil temperature and moisture as well as aspects of soil fertility.
2. Materials and Methods
2.1. Study Site
The study area was located at the Jiulingtou Forest Farm (30°59′ N, 110°47′ E) in Zigui County, Hubei Province, China (Figure 1). The values of annual mean temperature and annual rainfall were determined from 2010 to 2011 in Zigui County, Hubei Province, China. Annual mean temperature was approximately 16.9 °C and annual rainfall ranged from 1000 to 1250 mm, primarily from April to September. The monospecific plantations of P. massoniana, which is the most commonly used indigenous species for reforestation in this area, were aerially seeded in the 1970s and those grown on Haplic Luvisol soil were selected for this study [44]. Six plots (each 20 × 20 m) were randomly delineated for sampling at an elevation of 1225 m above sea level, in an area >1 ha. The study area was relatively steep, with a northwest slope of 34°, average diameter at breast height (DBH; tree diameter at 1.30 m) of 20.4 cm, average tree height of 12.4 m and tree density of 1688 hm−2. The average DBH of understory shrubs, such as Lespedeza bicolor Turcz., Pyracantha fortuneana (Maxim.), and Litsea pungens Hemsl., was 5.00 cm and average height was 5.60 m. Herbs in the study area were mainly Woodwardia japonica L.f., Carex tristachya Thunb., Aster ageratoides Turcz., and Parathelypteris nipponica (Franchet and Savatier) Ching.
2.2. Determination of Fine Root Standing Biomass (FRB) and Necromass
The six plots were divided into 24 subplots (10 × 10 m) and the fine roots within each subplot were randomly sampled every two months, from May 2014 to March 2015 (no samples were collected in January as the Jiulingtou Forest Farm was closed due to low temperatures). These soil cores were collected using the soil-coring method (18 cm height × 10 cm diameter steel bucket type soil auger with a T-handle). One hundred and twenty soil cores (24 subplots × 5 replicates per subplot) were collected during each sampling period from 0 to 20 cm depth using soil sampler with 20 cm length sign. The distance between cores within each subplot was more than 2 m. Core samples were transferred to a cool box, transported to the laboratory, and kept at 4 °C until processed (no more than one week).
Soil cores were washed with tap water on a 0.5 mm pore sieve to remove small particles of soil and impurities, and roots were manually separated using tweezers. Dead and living fine roots were assessed based on their colour, lustre, smell, elasticity, toughness, and shape [45]. To determine FRB, all P. massoniana roots in diameter class <2 mm were sorted into three different diameter classes (<0.5 mm, 0.5–1 mm, and 1–2 mm) and oven-dried at 70 °C for 48 h before being weighed. The average values of FRB and necromass were obtained from the five replicates per subplot.
2.3. Fine Root Decomposition Experiment
Fresh roots were collected at 0–10 cm depth in a P. massoniana plantation in March 2014. Living fine roots of P. massoniana were divided into three different diameter classes (0–0.5 mm, 0.5–1 mm, and 1–2 mm) and then air-dried to a constant weight [46]. The litter bag method was used to determine decomposing litter samples [47].
In the in situ fine root decomposition experiment, each litterbag (litterbag size: 100 mm ×100 mm and 0.1 mm-mesh polyethylene bag) was filled with precisely 1 g of air-dried fine roots. A subset of sample material was also retained for analyses to determine initial litter chemistry. In May 2014, the fine roots collected in the plantation were buried in the top soil (0–10 cm depth) of each of the six plots within the study site. Nine of the root litterbags (a total of 270 litter bags) were randomly collected from each plot at 69, 136, 169, 245, and 368 day after incubation, from May 2014 to March 2015. Once in the laboratory, the roots were taken from the bags using tweezers, carefully cleaned to remove soil particles, and oven-dried at 50 °C for 48 h before being weighed [48]. Samples were ground in a mill to pass through a 0.25 mm sieve before chemical analysis. In addition, soil surface temperature and moisture were measured in the six plots, simultaneously and at 1 h intervals, every day during the decomposition period using WatchDog 2000-series weather stations (Spectrum Technologies, Plainfield, IL, USA).
2.4. Chemical Analysis of Litter and Soil Samples
Soil samples were collected from 0 to 20 cm depth when root samples were collected in the same 20 × 20 m plots, taken to the laboratory, washed and sieved (2 mm) to remove impurities, and air-dried before chemical analysis. Subsamples of root litter and soil were analysed for: (1) organic C, using the dichromate oxidation method [49]; (2) soil nitrogen (N), using the Kjeldahl method (NH4+-N and NO3−-N concentrations using a flow injection analyser) [50]; (3) total calcium (Ca) and total phosphorus (P), using inductively coupled plasma mass spectrometry (ICP-MS) analysis (IRIS Intrepid II XSP system; Thermo Electric Co., West Chester, PA, USA); (4) soil pH, using an electrode-equipped pH meter (M229 Lab pH Meter); and (5) total and available potassium (K) and available P, using the standard protocol of the State Forestry Administration [51].
2.5. Date Analyses
A single exponential curve for the constant potential mass lost was fitted using the model:
where X is the mass remaining at time t (i.e., 69, 136, 169, 245, and 368 day after incubation), X0 is the initial mass value, and k is the decomposition coefficient [52]. Fine root production was calculated according to:
where P is the production, B is the average annual biomass value of living fine roots, Bt2−t1 is the change in living FRB between each sampling interval, Mt2−t1 is the change in dead FRB between each sampling interval, D is the annual decomposition mass of fine roots, M is the average annual necromass value of fine roots, and Dr is the annual decomposition rate of fine roots [53,54].
ln(X/X0 = −kt)
P = Bt2−t1 + Mt2−t1 + D, D = M × Dr
Remaining nutrients in fine roots were calculated as:
where W0 is the root initial dry weight, Wt is the root dry weight at time t (i.e., 69, 136, 169, 245, and 368 day after incubation), C0 is the fine root initial nutrient concentration, and Ci is the fine root nutrient concentration at time t (i.e., 69, 136, 169, 245, and 368 day after incubation).
CiWt × 100%/C0W0
All data were statistically analysed using SPSS 22.0 software (SPSS Inc., Chicago, IL, USA). All data were subjected to one-way analysis of variance (ANOVA) and comparisons among means were made using the Duncan multiple analysis test calculated at p < 0.05 and p < 0.01. Pearson’s correlation coefficients were calculated between fine root production or fine root decomposition rates and soil properties (temperature, moisture, total N, NH4+-N, NO3−-N, and total Ca) or roots’ initial properties. The statistical significance levels were set at p < 0.05 and p < 0.01. Significant differences are identified with a, b, c, and d (p < 0.05) or A, B, C, and D (p < 0.01). Significant correlations are indicated by * (p < 0.05) or ** (p < 0.01).
3. Results
3.1. Belowground Environment and Soil Chemical Properties
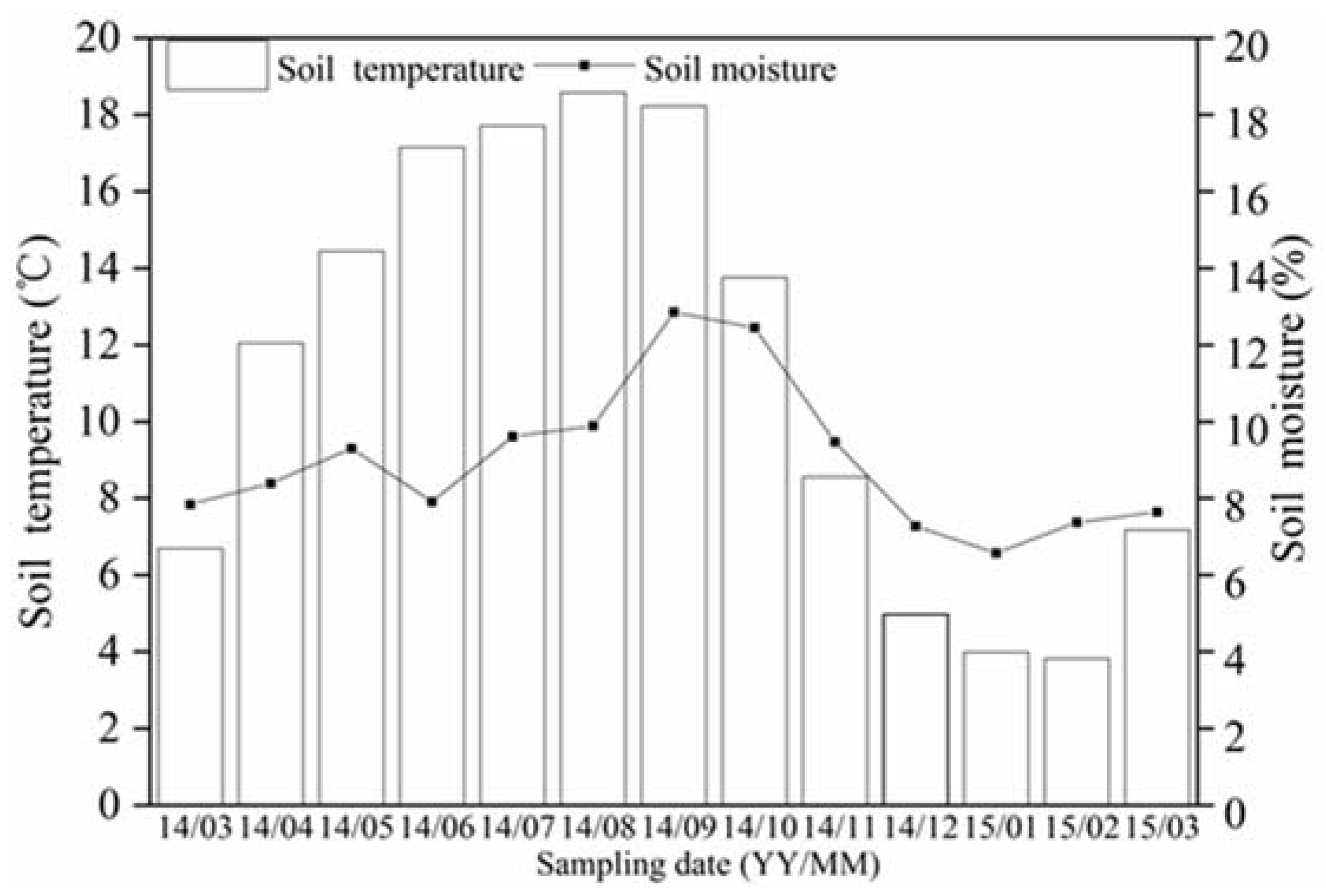
Soil annual average temperature and moisture are shown in Figure 2. The soil was weakly acidic and its total N, P, and K concentrations were 1.05, 0.20, and 0.25 g·kg−1, respectively. During the monitoring period, soil temperature ranged from 3.82 °C to 18.58 °C and moisture from 6.57% to 12.85%. Soil moisture increased from June to September 2014, when the highest value was recorded, decreasing sharply from September 2014 to January 2015. Soil temperature increased from March to August 2014, when the highest temperature was recorded, and then rapidly decreased until January and February 2015.
3.2. Fine Root Biomass, Necromass and Annual Production
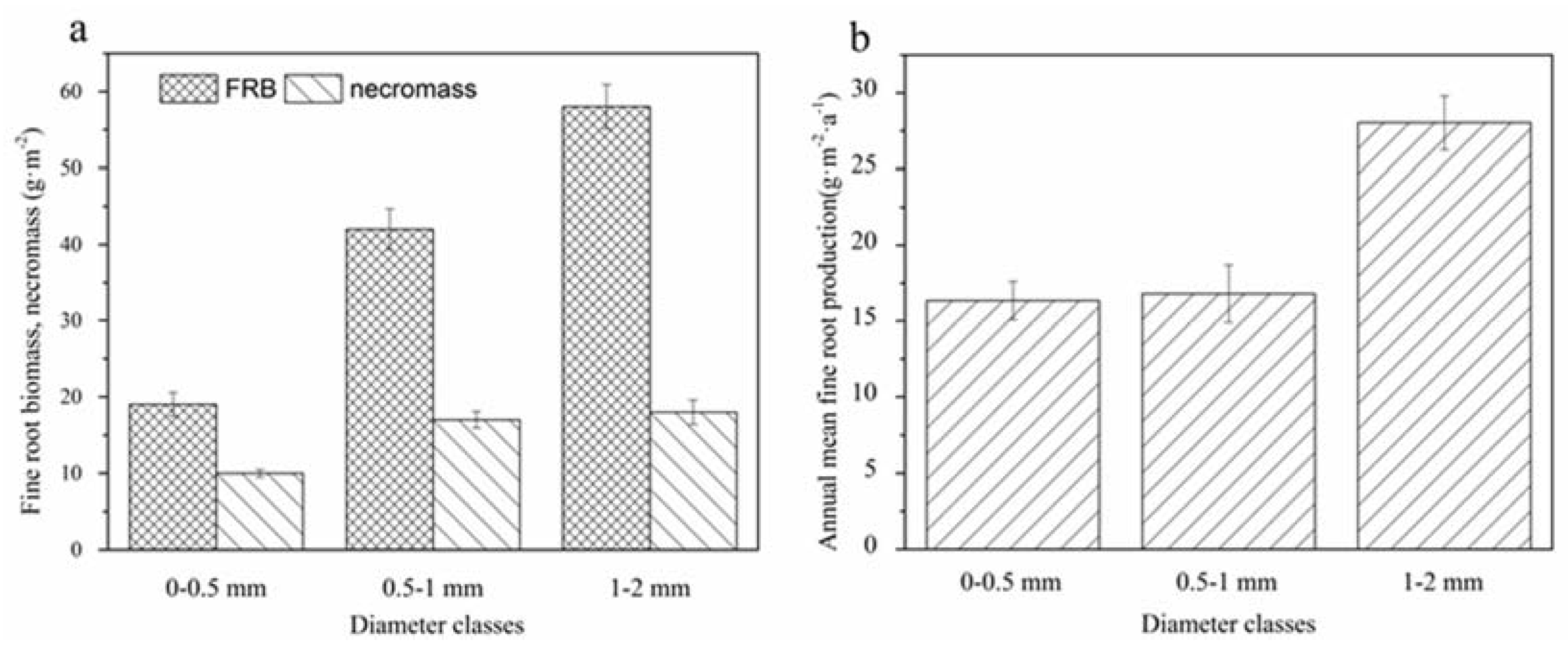
The accumulation of total fine root total biomass (FRB plus necromass) down to 20 cm in mineral soil in the P. massoniana plantation was 164 g·m−2, including 119 g·m−2 from FRB and 45 g·m−2 from necromass (Figure 3). The highest FRB was observed in 1–2 mm roots, which was about twice that of roots in diameter class <0.5 mm. The annual production of the three classes of P. massoniana fine roots down to 20 cm in mineral soil ranged from 16.8 g·m−2·a−1 to 28.03 g·m−2·a−1. The production of roots in diameter class 1–2 mm was the highest (45.81% of total production) whereas that of roots in diameter class <0.5 mm was the lowest (27.46% of total production).
Total FRB, necromass, and annual production increased with root diameter and differed significantly among the three classes of roots (p < 0.05) (Figure 3). In addition, there were significant differences between the total FRB of roots in diameter classes <0.5 mm and 0.5–1 mm (p < 0.05), and the necromass of roots in diameter class <0.5 mm roots was significantly different from that of 0.5 to 1 and 1 to 2 mm (p < 0.05).
3.3. The Initial C and N Content and Annual C and N Production in Fine Roots
Carbon concentration, annual C production, and C/N ratio in roots showed a clear increasing trend with increasing root diameter, while N concentration decreased (Table 1). The total annual C and N production in fine roots down to 20 cm in mineral soil was 27.5 g·m−2·a−1 and 0.61 g·m−2·a−1, respectively. The highest annual production of C and N was obtained in 1–2 mm roots, which represented 46.84% and 40.98% of total fine root C and N production, respectively. The minimum annual C production was observed in roots in diameter class <0.5 mm (25.42%) while the minimum annual N production was observed in roots in diameter class 0.5–1 mm (27.87%).
Fine root N contents were not significantly different among the three classes (p > 0.05), and showed a narrow range (0.9–1.17 g·kg−1). Moreover, the C/N ratio, annual C and N production of fine roots were significantly higher in 1–2 mm roots than those of roots in diameter classes < 0.5 mm and 0.5–1 mm (p < 0.01). Initial C concentration and C/N ratio of roots in diameter class < 0.5 mm were significantly smaller than those of roots in diameter class 0.5–1 mm (p < 0.01).
3.4. Remaining Fine Root Mass after Decomposition Experiment
In the decomposition experiment, all P. massoniana fine roots showed a rapid decomposition rate during the first 136 day of incubation time (Table 2), and there was a significant difference among the different root classes (p < 0.05). All fine root decomposition rates decreased with time of incubation increasing, after 136 day of incubation time, the mass loss of roots in diameter classes <0.5 mm, 0.5–1 mm and 1–2 mm was approximately 26.00%, 21.67%, and 18.67%, respectively. After 368 day of incubation time, the initial mass of roots in diameter classes <0.5 mm, 0.5–1 mm and 1–2 mm decreased by nearly 34.00%, 28.00%, and 25.72%, respectively. Fine root annual decomposition rate decreased with increasing root diameter to values of 0.52 a−1, 0.31 a−1, and 0.31 a−1, for diameter classes <0.5 mm, 0.5–1 mm and 1–2 mm fine root, respectively. However, decomposition rates of roots in diameter classes <0.5 mm and 0.5–1 mm gradually decreased with increasing incubation time (368 day), while decomposition rate of 1–2 mm roots gradually increased and then decreased.
3.5. Changes in C and N Concentrations in Fine Roots during Decomposition
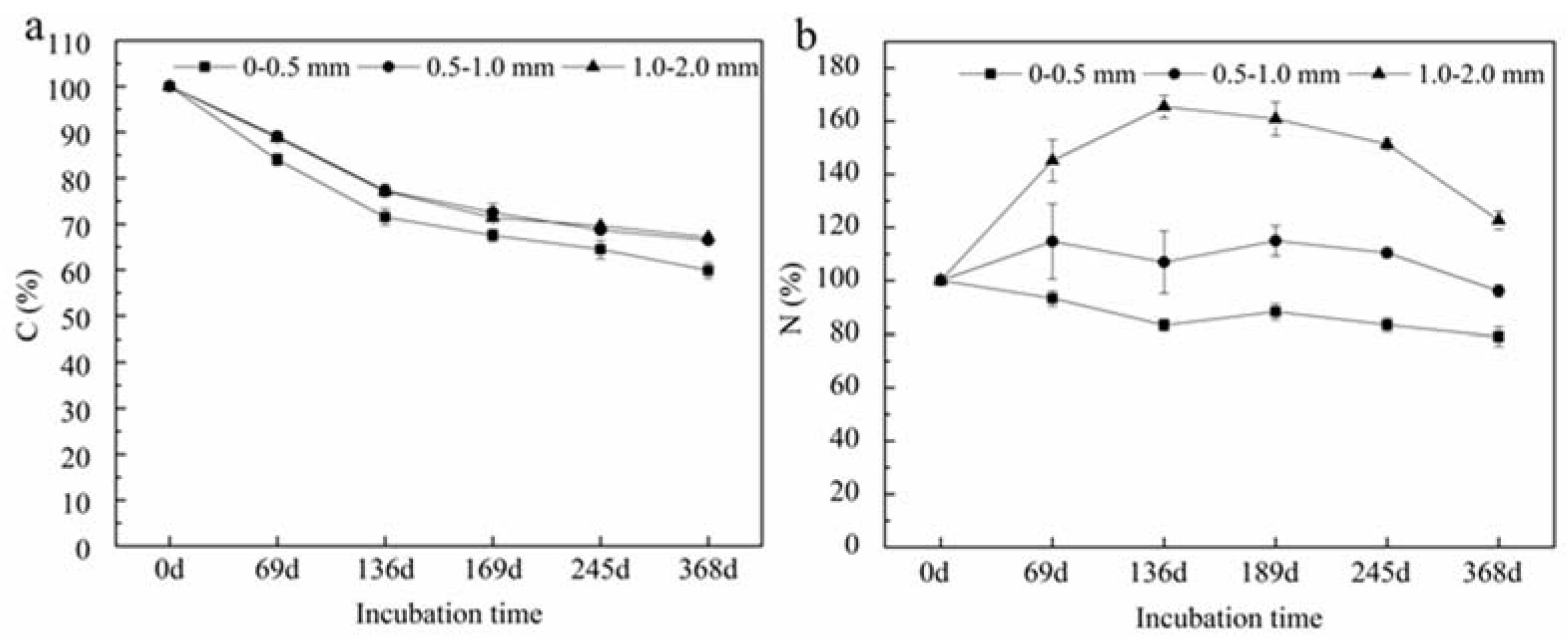
Carbon concentrations in P. massoniana fine roots decreased with incubation time (Figure 4). Approximately 40.12%, 33.52%, and 32.92% of the initial C concentration in roots in diameter classes <0.5 mm, 0.5–1 mm and 1–2 mm, respectively, was lost after 368 day of incubation. On the other hand, N concentration of roots in diameter class <0.5 mm roots decreased by 20.83% and 0.5–1 mm roots decreased by 3.86% during the decomposition process while N concentration in 1–2 mm roots increased.
3.6. Relationships between Fine Root Indicators and Soil Characteristics
Table 3 show that production of roots in diameter classes <0.5 mm and 0.5–1 mm was significantly correlated with soil Ca status (r = 0.810, p < 0.01; r = 0.855, p < 0.01, respectively). Production of roots in diameter class <0.5 mm was also significantly correlated with soil temperature (r = 0.572, p < 0.05). Decomposition rates in all fine root classes were positively correlated with soil temperature (r = 0.617, p < 0.05; r = 0.815, p < 0.01; and r = 0.968, p < 0.01, respectively, decomposition rates vary from warmer periods to colder periods in a year) and initial N (r = 0.717, p < 0.05; r = 0.227, p < 0.05; and r = 0.439, p < 0.05 for roots in diameter classes <0.5 mm, 0.5–1 mm and 1–2 mm, respectively), and significantly and negatively correlated with soil Ca status (r = −0.585, p < 0.05; r = −0.720, p < 0.05; and r = −0.835, p < 0.01 for roots in diameter classes < 0.5 mm, 0.5–1 mm and 1–2 mm,, respectively) and initial C/N ratio (r = −0.901, p < 0.01; r = −0.886, p < 0.01; and r = −0.927, p < 0.01 for roots in diameter classes <0.5 mm, 0.5–1 mm and 1–2 mm, respectively).
4. Discussion
4.1. Fine Roots Biomass and Production in Different Diameter Classes
The FRB and necromass increased with increasing root diameter. Furthermore, production of 1–2 mm roots was significantly higher than that of roots in diameter classes <0.5 mm and 0.5–1 mm, suggesting that the biomass and production of 1–2 mm roots made the largest contribution to total fine root production and biomass. This pattern was similar to that found by Wang et al. [40] and supported the results of Finér et al. [55] and Ostonen et al. [56]. In our P. massoniana stands in the Three Gorges Reservoir area, the mean total live fine root biomass and production were lower than other values reported for Scots pine [42]. Fine root production can vary owing to differences in climate and forest types, as production and turnover rates increase gradually from boreal to tropical forests [18].
Fine root N and C production showed different patterns among the three classes of P. massoniana fine roots. The study results, combined with the findings of Guo et al. [31], suggest that larger dimeter roots are more important in C storage, and small diameter roots have higher N concentrations. The root system is a crucial trait for plants, as it largely determines how efficiently water and nutrients will be extracted from the soil [27]. In our study, fine root N concentrations of very fine roots were higher than those of larger roots and C/N ratios of very fine roots were significantly lower than those of larger roots. However, if only one type of root parameter is applied, as in previous studies of very fine roots, changes in the root plasticity of species-specific traits cannot be explained.
4.2. The Decomposition Rate and C, N Dynamics of Fine Roots in Decomposition Experiment
We assessed whether diameter-related morphological and soil chemical properties affect the decomposition pattern of P. massoniana fine roots (<0.5 mm, 0.5–1 mm and 1–2 mm) in the Three Gorges Reservoir area. The decomposition constant value (k year−1) of roots in diameter classes <0.5 mm, 0.5–1 mm and 1–2 mm after 368 days was 0.52, 0.31 and 0.31, respectively, suggesting that k fell within a range of its global median value. The values in our study were lower than the k value (0.61) of P. massoniana fine roots in diameter classes <2 mm alone [57].
The three classes of P. massoniana fine roots showed two distinct stages during their decomposition, a pattern that has been reported in several studies [40,58,59,60]. Those two stages were rapid decomposition and mass loss very early and then a transition to relatively slower mass loss in later decomposition stages. The rapid decomposition rate of fine roots detected in the beginning of the experiment could be due to fungi and microbes that rapidly degrade labile tissues to access the relatively high carbohydrate content in root organs at the initial stage of decomposition. The later stages of decomposition include the degradation of more recalcitrant compounds in fine roots including lignin, cellulose, and other insoluble residues leading to a decrease in fine root decomposition rates with incubation time [14]. In addition, the results may also be partially due to the fact that the bags were placed in the field in April, and the first 6 months of incubation were very warm. Then, the later months (November to March) were very cold, during which period microbial activity and decomposition would be very slow regardless of the lability of recalcitrance of the root tissue.
During fine root decomposition, nutrients are released, affecting the soil nutrient content [60]. The C concentrations in all diameter classes of fine roots decreased with incubation time. However, although N was released from roots in diameter classes <0.5 mm, 0.5–1 mm and 1–2 mm became N-enriched with incubation time. These patterns were similar to those observed by Goebel et al. [61].
4.3. Soil Factors Influences on Fine Root Traits
Soil temperature had little effect on fine root production, only presenting a significant correlation with roots in diameter <0.5 mm across a year. However, there was a significantly positive correlation between fine root decomposition rate and soil temperature, which was, therefore, the major driver of fine root decomposition, similar to that found in previous studies [57,62,63,64,65,66]. According to Cusack et al. [62], precipitation-related parameters are strong predictors of litter decomposition in neotropical forests, and Hobbie et al. [67] also suggested that moister soils could facilitate rapid mass loss by promoting leaching and microbial activity or a faster colonisation of decomposing material by microbes. Moreover, Zhang et al. [68] and Violita et al. [69] stated that the decomposition rate of roots increased with increasing internal Ca concentration in roots. However, in our study, soil Ca concentration had a negative influence on the decomposition rate of all fine root classes. If decomposition is higher with higher root Ca content, it implies that microbes are decomposing the roots to access the Ca. If soil Ca is high, microbes would have no need for the Ca in roots; therefore, there is less drive to decompose the roots. Thus, the chemical composition of fine roots affects their decomposition rate. Fine roots with a thinner diameter tend to have a higher initial N concentration, which is thought to have result in faster decomposition [70,71,72]. Hishi and Takeda [32] reported that thinner roots with a lower C/N ratio decomposed faster than thicker roots. In addition, it is well known that substrates with higher C/N ratios have low amounts of N available to decomposers, suggesting that the low levels of N released from fine roots might have been influenced by both initial and residual chemistry [73,74]. Therefore, litter composed mainly of thicker roots will generally have a high C/N, or a low N concentration, and could lead to faster immobilization of N than could thinner roots, leading to lower decomposition rates. In our study, we found a relatively high initial C/N in the 0.5–2 mm roots; hence, it is likely that N availability was a limiting factor for decomposers [75,76]. Consequently, litter from thinner roots of P. massoniana should have decomposed faster than thicker roots by the end of the experiment because of the different chemical properties prior to decay.
5. Conclusions
In the Pinus massoniana stand in the Three Gorges Reservoir area, tree fine roots varied depending on different diameter classes, especially very fine roots with a faster decomposition ratio than larger fine roots. The differences found were mainly due to, first, the effect of soil temperature and moisture on the production and decomposition rate of fine roots of different diameters, especially soil moisture. Second, soil Ca concentration was especially important and slowed the decomposition rates of the three classes of fine roots. Third, fine root characteristics were important for both decomposition rate and production, which were also significantly different among classes (e.g., lower C/N ratios resulted in the slower decomposition of fine roots). These results can help explain differences in important mechanisms of root decomposition among roots less than 2 mm in diameter and can improve our understanding of the mechanisms related to soil C fluxes and the quantification of soil C sequestration in forest ecosystems.
Supplementary Materials
Supplementary File 1Acknowledgments
We appreciate M. Luke McCormack at University of Minnesota for his valuable comments and suggestions on the manuscript. This project was financially supported by the National Key Research and Development Program of the Ministry of Science and Technology of China (No. 2016YFD0600204).
Author Contributions
Y.S. analyzed data and drafted the manuscript. N.W. participated in collecting the experiment data. R.C. was involved in planning of study and designing of the work. W.X. was substantial contributions to the conception or design of the work. The remaining authors contributed to refining the ideas, carrying out additional analyses and finalizing this paper. All authors discussed the results and revised the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Six, J.; Callewaert, P.; Lenders, S.; De Gryze, S.; Morris, S.; Gregorich, E.; Paul, E.; Paustian, K. Measuring and understanding carbon storage in afforested soils by physical fractionation. Soil Sci. Soc. Am. J. 2002, 66, 1981–1987. [Google Scholar] [CrossRef]
- Chapin, F.S.; Ruess, R.W. Carbon cycle: The roots of the matter. Nature 2001, 411, 749–752. [Google Scholar] [CrossRef] [PubMed]
- Trumbore, S.E.; Gaudinski, J.B. The secret lives of roots. Science 2003, 302, 1344–1345. [Google Scholar] [CrossRef] [PubMed]
- Upson, M.; Burgess, P. Soil organic carbon and root distribution in a temperate arable agroforestry system. Plant Soil 2013, 373, 43–58. [Google Scholar] [CrossRef]
- Leppälammi-Kujansuu, J.; Aro, L.; Salemaa, M.; Hansson, K.; Dan, B.K.; Helmisaari, H.S. Fine root longevity and carbon input into soil from below- and above-ground litter in climatically contrasting forests. For. Ecol. Manag. 2014, 326, 79–90. [Google Scholar] [CrossRef]
- McCormack, M.; Eissenstat, D.; Prasad, A.; Smithwick, E. Regional scale patterns of fine root lifespan and turnover under current and future climate. Glob. Chang. Biol. 2013, 19, 1697–1708. [Google Scholar] [CrossRef] [PubMed]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine North American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- Mccormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- McClaugherty, C.A.; Aber, J.D.; Melillo, J.M. The role of fine roots in the organic matter and nitrogen budgets of two forested ecosystems. Ecology 1982, 63, 1481–1490. [Google Scholar] [CrossRef]
- Freschet, G.T.; Cornwell, W.K.; Wardle, D.A.; Elumeeva, T.G.; Liu, W.; Jackson, B.G.; Onipchenko, V.G.; Soudzilovskaia, N.A.; Tao, J.; Cornelissen, J.H.C. Linking litter decomposition of above- and below-ground organs to plant-soil feedbacks worldwide. J. Ecol. 2013, 101, 943–952. [Google Scholar] [CrossRef]
- Yuan, Z.; Chen, H.Y. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses. Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Lin, C.F.; Yang, Y.S.; Guo, J.F.; Chen, G.S.; Xie, J.S. Fine root decomposition of evergreen broadleaved and coniferous tree species in mid-subtropical China: Dynamics of dry mass, nutrient and organic fractions. Plant Soil 2011, 338, 311–327. [Google Scholar] [CrossRef]
- Sun, T.; Mao, Z.J.; Dong, L.L.; Hou, L.L.; Song, Y.; Wang, X.W. Further evidence for slow decomposition of very fine roots using two methods: Litterbags and intact cores. Plant Soil 2013, 366, 633–646. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.; Schulze, E.-D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Talhelm, A.F.; Pregitzer, K.S. Fine roots are the dominant source of recalcitrant plant litter in sugar maple-dominated northern hardwood forests. New Phytol. 2015, 208, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Helmisaari, H.S.; Makkonen, K.; Kellomäki, S.; Valtonen, E.; Mälkönen, E. Below- and above-ground biomass, production and nitrogen use in Scots pine stands in eastern Finland. For. Ecol. Manag. 2002, 165, 317–326. [Google Scholar] [CrossRef]
- Montagnoli, A.; Terzaghi, M.; Di Iorio, A.; Scippa, G.S.; Chiatante, D. Fine-root morphological and growth traits in a Turkey oak stand in relation to seasonal changes in soil moisture in the Southern Apennines. Italy. Ecol. Res. 2012, 27, 1015–1025. [Google Scholar] [CrossRef]
- Björk, R.G.; Majdi, M.; Klemedtsson, L.; Jonsson, L.L.; Molau, U. Long-term warming effects on root morphology, root mass distribution, and microbial activity in two dry tundra plan communities in northern Sweden. New Phytol. 2007, 176, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Park, B.B.; Yanai, R.D.; Fahey, T.J.; Bailey, S.W.; Siccama, T.G.; Shanley, J.B.; Cleavitt, N.L. Fine root dynamics and forest production across a calcium gradient in northern hardwood and conifer ecosystems. Ecosystems 2008, 11, 325–341. [Google Scholar] [CrossRef]
- Jackson, R.B.; Schenk, H.J.; Jobbágy, E.G.; Canadell, J.; Colello, G.D.; Dickinson, R.E.; Field, C.B.; Friedlingstein, P.; Heimann, M.; Hibbard, K.; et al. Belowground consequences of vegetation change and their treatment in models. Ecol. Appl. 2000, 10, 470–483. [Google Scholar] [CrossRef]
- Joslin, J.D.; Gaudinski, J.B.; Torn, M.S.; Riley, W.J.; Hanson, P.J. Fine-root turnover patterns and their relationship to root diameter and soil depth in a C14 labeled hardwood forest. New Phytol. 2006, 172, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Finér, L.; Helmisaari, H.S.; Lohmus, K.; Majdi, H.; Brunner, I.; Børja, I.; Eldhuset, T.; Godbold, D.; Grebenc, T.; Konopka, B.; et al. Variation in fine root biomass of three European tree species: Beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.), and Scots pine (Pinus sylvestris L.). Plant Biosyst. 2007, 141, 394–405. [Google Scholar] [CrossRef]
- Noguchi, K.; Konopka, B.; Satomura, T.; Kaneko, S.; Takahashi, M. Biomass and production of fine roots in Japanese forests. J. For. Res. 2007, 12, 83–95. [Google Scholar] [CrossRef]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Hishi, T. Heterogeneity of individual roots within the fine root architecture: Causal links between physiological and ecosystem functions. J. For. Res. 2007, 12, 126–133. [Google Scholar] [CrossRef]
- Noguchi, K.; Matsuura, Y.; Sparrow, S.D.; Hinzman, L.D. Fine root biomass in two black spruce stands in interior Alaska: Effects of different permafrost conditions. Trees 2016, 30, 441–449. [Google Scholar] [CrossRef]
- Makita, N.; Hirano, Y.; Dannoura, M.; Kominami, Y.; Mizoguchi, T.; Ishii, H.; Kanazawa, Y. Fine root morphological traits determine variation in root respiration of Quercus serrata. Tree physiol. 2009, 29, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.L.; Mitchell, R.J.; Hendricks, J.J. Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest. Oecologia 2004, 140, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Hishi, T.; Takeda, H. Dynamics of heterorhizic root systems: Protoxylem groups within the fine-root system of Chamaecyparis obtusa. New Phytol. 2005, 167, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Rewald, B.; Ephrath, J.E.; Rachmilevitch, S. A root is a root is a root? Water uptake rates of Citrus root orders. Plant Cell Environ. 2011, 34, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Mitchell, R.J.; Withington, J.M.; Fan, P.P.; Hendricks, J.J. Endogenous and exogenous controls of root life span, mortality and nitrogen flux in a longleaf pine forest: Root branch order predominates. J. Ecol. 2008, 96, 737–745. [Google Scholar] [CrossRef]
- Guo, D.; Xia, M.; Wei, X.; Chang, W.; Liu, Y.; Wang, Z. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, J.; Vogt, K.A.; Wargo, P.M. Tree root turnover and senescence. In Plant Roots:The Hidden Half, 2nd ed.; Waisel, Y., Eshel, A., Kafkafi, U., Eds.; Dekker: New York, NY, USA, 1996; pp. 363–381. [Google Scholar]
- Rustad, L.E. The response of terrestrial ecosystems to global climate change: Towards an integrated approach. Sci. Total Environ. 2008, 404, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Valverde-Barrantes, O.J.; Smemo, K.A.; Feinstein, L.M.; Kershner, M.W.; Blackwood, C.B. Aggregated and complementary: Symmetric proliferation, overyielding, and mass effects explain fine-root biomass in soil patches in a diverse temperate deciduous forest landscape. New Phytol. 2015, 205, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Vesterdal, L.; Clarke, N.; Sigurdsson, B.D.; Gundersen, P. Do tree species influence soil carbon stocks in temperate and boreal forests? For. Ecol. Manag. 2013, 309, 4–18. [Google Scholar] [CrossRef]
- Wang, R.L.; Cheng, R.M.; Xiao, W.F.; Feng, X.H.; Liu, Z.B.; Ge, X.G.; Wang, X.R.; Zhang, W.Y. Fine root production and turnover in Pinus massoniana plantation in Three Gorges Reservoir area of China. Chin. J. Appl. Ecol. 2012, 23, 2346–2352. [Google Scholar]
- Ge, X.G.; Xiao, W.F.; Zeng, L.X.; Huang, Z.L.; Fu, T.; Feng, X.H. Relationships between litter substrate quality and soil nutrients in different-aged Pinus massoniana stands. Acta Ecol. Sin. 2012, 32, 852–862. [Google Scholar]
- Hansson, K.; Helmisaari, H.S.; Sah, S.P.; Lange, H. Fine root production and turnover of tree and understorey vegetation in Scots pine, silver birch and Norway spruce stands in SW Sweden. For. Ecol. Manag. 2013, 309, 58–65. [Google Scholar] [CrossRef]
- Hirano, Y.; Tanikawa, T.; Makita, N. Biomass and morphology of fine roots in eight Cryptomeria japonica stands in soils with different acid-buffering capacities. For. Ecol. Manag. 2017, 384, 122–131. [Google Scholar] [CrossRef]
- Gong, Z.T. Chinese Soil Taxonomy; China Science Press: Beijing, China, 2003. [Google Scholar]
- Brassard, B.W.; Chen, H.Y.; Cavard, X.; Laganiere, J.o.; Reich, P.B.; Bergeron, Y.; Pare, D.; Yuan, Z. Tree species diversity increases fine root productivity through increased soil volume filling. J. Ecol. 2013, 101, 210–219. [Google Scholar] [CrossRef]
- Mo, J.; Brown, S.; Xue, J.; Fang, Y.; Li, Z. Response of litter decomposition to simulated N deposition in disturbed, rehabilitated and mature forests in subtropical China. Plant Soil 2006, 282, 135–151. [Google Scholar] [CrossRef]
- Crossley, D.; Hoglund, M.P. A litter-bag method for the study of microarthropods inhabiting leaf litter. Ecology 1962, 43, 571–573. [Google Scholar] [CrossRef]
- Ostertag, R.; Marín-Spiotta, E.; Silver, W.L.; Schulten, J. Litterfall and decomposition in relation to soil carbon pools along a secondary forest chronosequence in Puerto Rico. Ecosystems 2008, 11, 701–714. [Google Scholar] [CrossRef]
- Nelson, D.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis; ASA-SSSA: Madison, WI, USA, 1982; pp. 539–579. [Google Scholar]
- Zhu, T.B.; Zhang, J.B.; Meng, T.Z.; Zhang, Y.C.; Yang, J.J.; Müller, C.; Cai, Z.C. Tea plantation destroys soil retention of NO3—And increases N2O emissions in subtropical China. Soil Biol. Biochem. 2014, 73, 106–114. [Google Scholar] [CrossRef]
- Zhang, W.R.; Yang, G.C.; Tu, X.N. Adiministration Forestry Standard of People’s Republic of China—Method of Forest Soil Analysis; Chinese Standard Press: Beijing, China, 1999. [Google Scholar]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Mei, L.; Wang, Z.Q.; Zhang, X.J.; Yu, L.Z.; Du, Y. Effects of nitrogen fertilization on fine root biomass production and turnover of Fraxinus mandshurica plantation. Chin. J. Ecol. 2008, 27, 1663–1668. [Google Scholar]
- Vogt, K.A.; Vogt, D.J.; Bloomfield, J. Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level. Plant Soil 1998, 200, 71–89. [Google Scholar] [CrossRef]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Factors causing variation in fine root biomass in forest ecosystems. For. Ecol. Manag. 2011, 261, 265–277. [Google Scholar] [CrossRef]
- Ostonen, I.; Lõhmus, K.; Pajuste, K. Fine root biomass, production and its proportion of NPP in a fertile middle-aged Norway spruce forest: Comparison of soil core and ingrowth core methods. For. Ecol. Manag. 2005, 212, 264–277. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Mo, J. Correlation between leaf litter and fine root decomposition among subtropical tree species. Plant Soil 2010, 335, 289–298. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant litter. In Decomposition, Humus Formation, Carbon Sequestration, 2nd ed.; Springer: Berlin, Germany, 2008. [Google Scholar]
- Lin, C.F.; Yang, Y.S.; Chen, G.S.; Guo, J.F.; Han, Z.G. Decomposition dynamics of fine roots of Cunninghamia lanceolata in mid-subtropics. J. Subtrop. Resour. Environ. 2008, 3, 15–23. [Google Scholar]
- Wang, C.G.; Chen, Z.X.; Ma, C.E.; Lin, G.G.; Han, S.J. Three potential pathways influencing contrasting decomposition rates of fine roots. J. Beijing For. Univ. 2016, 38, 123–128. [Google Scholar]
- Goebel, M.; Hobbie, S.E.; Bulaj, B.; Zadworny, M.; Archibald, D.D.; Oleksyn, J.; Reich, P.B.; Eissenstat, D.M. Decomposition of the finest root branching orders: Linking belowground dynamics to fine-root function and structure. Ecol. Monogr. 2011, 81, 89–102. [Google Scholar] [CrossRef]
- Cusack, D.F.; Chou, W.W.; Yang, W.H.; Harmon, M.E.; Silver, W.L. Controls on long-term root and leaf litter decomposition in neotropical forests. Glob. Chang. Biol. 2009, 15, 1339–1355. [Google Scholar] [CrossRef]
- Gimbel, K.F.; Felsmann, K.; Baudis, M.; Puhlmann, H.; Gessler, A.; Bruelheide, H.; Kayler, Z.; Ellerbrock, R.H.; Ulrich, A.; Welk, E. Drought in forest understory ecosystems—A novel rainfall reduction experiment. Biogeosciences 2015, 12, 14319–14358. [Google Scholar] [CrossRef]
- Steinberger, Y.; Degani, R.; Barness, G. Decomposition of root litter and related microbial population dynamics of a Negev Desert shrub, Zygophyllum dumosum. J. Arid Environ. 1995, 31, 383–399. [Google Scholar] [CrossRef]
- Wieder, R.K.; Wright, S.J. Tropical forest litter dynamics and dry season irrigation on Barro Colorado Island, Panama. Ecology 1995, 76, 1971–1979. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Wang, W. The decomposition of fine and coarse roots: Their global patterns and controlling factors. Sci. Rep. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, S.E.; Oleksyn, J.; Eissenstat, D.M.; Reich, P.B. Fine root decomposition rates do not mirror those of leaf litter among temperate tree species. Oecologia 2010, 162, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Q.; Hui, D.F.; Luo, Y.Q.; Zhou, G.Y. Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors. J. Plant Ecol. 2008, 1, 85–93. [Google Scholar] [CrossRef]
- Violita, T.; Triadiati, T.; Anas, I.; Miftahudin, M. Fine root production and decomposition in lowland rainforest and oil palm plantations in Sumatra, Indonesia. Hayati J. Biosci. 2016, 23, 7–12. [Google Scholar] [CrossRef]
- Berg, B.; Berg, M.P.; Bottner, P.; Box, E.; Breymeyer, A.; Ca de Anta, R.; Couteaux, M.; Escudero, A.; Gallardo, A.; Kratz, W.; et al. Litter mass loss rates in pine forests of Europe and Eastern United States: Some relationships with climate and litter quality. Biogeochemistry 1993, 20, 127–159. [Google Scholar] [CrossRef]
- Ruess, R.W.; Hendrick, R.L.; Burton, A.J.; Pregitzter, K.S.; Sveinbjornsson, B.; Allen, M.F.; Maurer, G.E. Coupling fine root dynamics with ecosystem carbon cycling in black spruce forests of interior Alaska. Ecol. Monogr. 2003, 73, 643–662. [Google Scholar] [CrossRef]
- Yang, Y.S.; Chen, G.S.; Guo, J.F.; Lin, P. Decomposition dynamic of fine roots in a mixed forest of Cunninghamia lanceolata and Tsoongiodendron odorum in midsubtropics. Ann. For. Sci. 2004, 61, 65–72. [Google Scholar] [CrossRef]
- Parton, W.; Silver, W.L.; Burke, I.C.; Grassens, L.; Harmon, M.E.; Currie, W.S.; King, J.Y.; Carol Adair, E.; Brandt, L.A.; Hart, S.C.; et al. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 2007, 315, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, S.; Jackson, R.B.; Trofymow, J.A.; Porporato, A. The global stoichiometry of litter nitrogen mineralization. Science 2008, 321, 684–686. [Google Scholar] [CrossRef] [PubMed]
- Fahey, T.J.; Hughes, J.W.; Pu, M.; Arthur, M.A. Root decomposition and nutrient flux following whole-tree harvest of northern hardwood forest. For. Sci. 1988, 34, 744–768. [Google Scholar]
- Fan, P.; Jiang, Y. Nitrogen dynamics differed among the first six root branch orders of Fraxinus mandshurica and Larix gmelinii during short-term decomposition. J. Plant Res. 2010, 123, 433–438. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Location of the study area. The study area, located at the Jiulingtou Forest Farm (30°59′ N, 110°47′ E), is indicated by a triangle and Zigui City is marked in green. The blue line represents the Yangtze River flowing through the Three Gorges Reservoir Area, which is the region surrounded by the red line.
Figure 1.
Location of the study area. The study area, located at the Jiulingtou Forest Farm (30°59′ N, 110°47′ E), is indicated by a triangle and Zigui City is marked in green. The blue line represents the Yangtze River flowing through the Three Gorges Reservoir Area, which is the region surrounded by the red line.
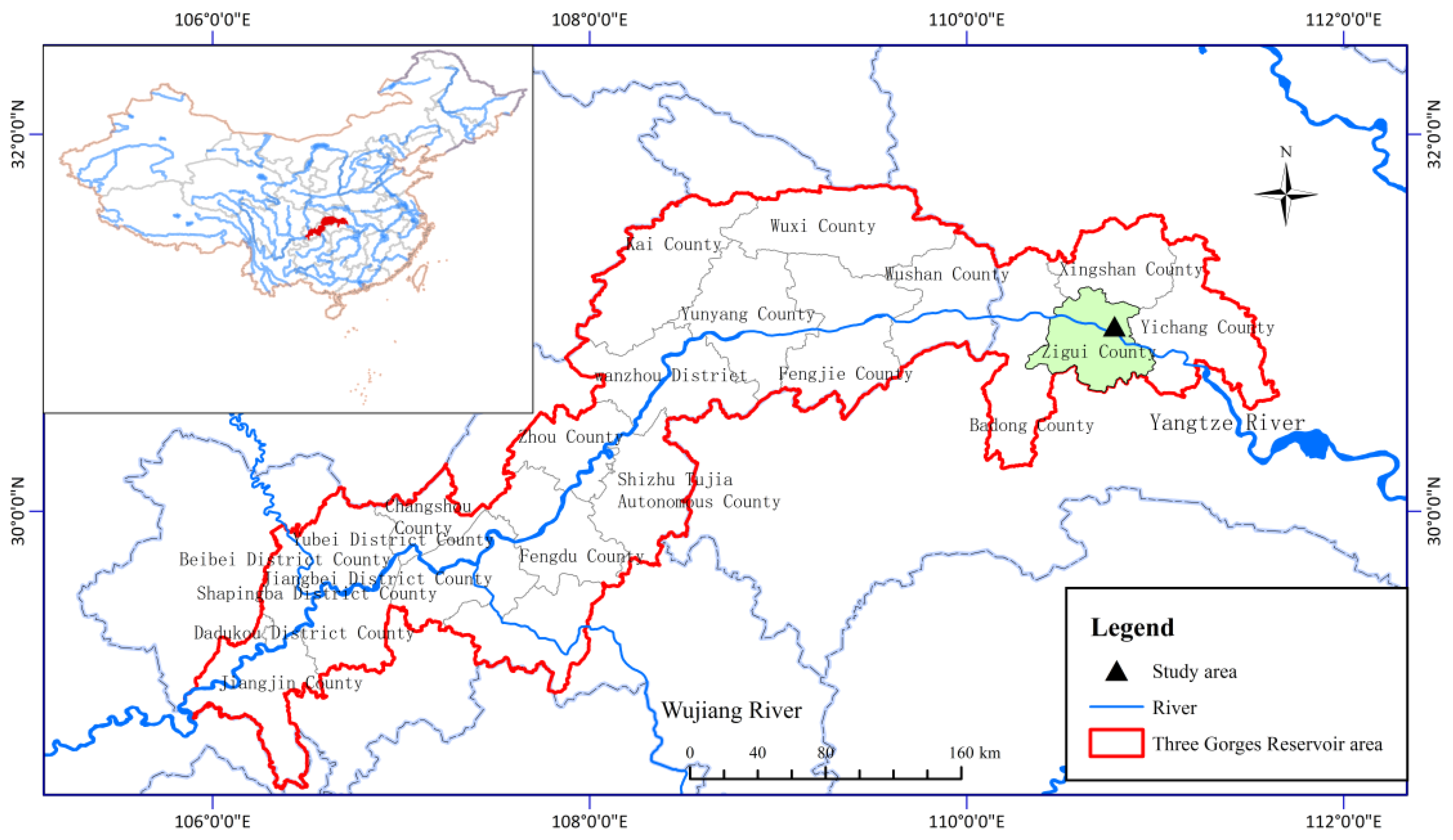
Figure 2.
Changes in soil temperature and moisture at 10 cm depth. Soil temperature (°C) and moisture (%) at 0–10 cm depth, from March 2014 to March 2015, are shown. The bars represent soil temperature and the lines soil moisture.
Figure 2.
Changes in soil temperature and moisture at 10 cm depth. Soil temperature (°C) and moisture (%) at 0–10 cm depth, from March 2014 to March 2015, are shown. The bars represent soil temperature and the lines soil moisture.

Figure 3.
Root biomass and necromass (a); and annual production (b) of the three classes of Pinus massoniana fine roots. Bars with solid and dashed lines in (a) represent the necromass and fine root biomass (FRB), respectively, of roots within the 0–0.5, 0.5–1, and 1–2 mm diameter classes. The annual production of fine roots from the three diameter classes is indicated in (b). Values are means ± standard error (n = 5).
Figure 3.
Root biomass and necromass (a); and annual production (b) of the three classes of Pinus massoniana fine roots. Bars with solid and dashed lines in (a) represent the necromass and fine root biomass (FRB), respectively, of roots within the 0–0.5, 0.5–1, and 1–2 mm diameter classes. The annual production of fine roots from the three diameter classes is indicated in (b). Values are means ± standard error (n = 5).

Figure 4.
Changes in carbon (C) (a) and nitrogen (N) (b) concentrations in fine roots during decomposition. The three lines with circles, triangles, and diamonds represent the changes in C and N concentrations in fine roots with 0–0.5, 0.5–1, and 1–2 mm diameter, respectively, during the decomposition experiment. Values are means ± standard error (n = 9).
Figure 4.
Changes in carbon (C) (a) and nitrogen (N) (b) concentrations in fine roots during decomposition. The three lines with circles, triangles, and diamonds represent the changes in C and N concentrations in fine roots with 0–0.5, 0.5–1, and 1–2 mm diameter, respectively, during the decomposition experiment. Values are means ± standard error (n = 9).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Initial carbon (C) and (N) concentration and production (mean value ± standard error) in fine roots (n = 5).
Table 1.
Initial carbon (C) and (N) concentration and production (mean value ± standard error) in fine roots (n = 5).
| Size Classes | C Concentration (g·kg−1) | N Concentration (g·kg−1) | C/N Ratio | Annual C Production (g·m−2·a−1) | Annual N Production (g·m−2·a−1) |
|---|---|---|---|---|---|
| 0–0.5 mm | 42.72 ± 0.39 A | 1.17 ± 0.02 a | 36.66 ± 0.39 A | 6.99 ± 0.38 A | 0.19 ± 0.02 A |
| 0.5–1 mm | 45.04 ± 0.12 B | 1.00 ± 0.01 b | 45.04 ± 0.57 B | 7.63 ± 0.07 A | 0.17 ± 0.01 A |
| 1–2 mm | 45.95 ± 0.16 B | 0.90 ± 0.03 c | 51.06 ± 1.54 C | 12.88 ± 0.16 B | 0.25 ± 0.04 B |
C Carbon. Significant differences are identified with a, b, c, and d (p < 0.05) or A, B, C, and D (p < 0.01) based on the analysis of variance.
Table 2.
Fine root mass of Pinus massoniana remaining after the decomposition experiments (mean value ± standard error; n = 9).
Table 2.
Fine root mass of Pinus massoniana remaining after the decomposition experiments (mean value ± standard error; n = 9).
| Incubation Time (Days) | 0–0.5 mm | 0.5–1 mm | 1–2 mm |
|---|---|---|---|
| Mass Remaining (%) | Mass Remaining (%) | Mass Remaining (%) | |
| 69 day | 85.5 ± 0.01 a | 89.8 ± 0.01 b | 91.1 ± 0.02 c |
| 136 day | 74.3 ± 0.02 a | 78.4 ± 0.01 b | 81.1 ± 0.02 c |
| 169 day | 71.5 ± 0.02 a | 75.5 ± 0.01 a | 75.6 ± 0.02 a |
| 245 day | 69.1 ± 0.02 a | 73.2 ± 0.01 a | 76.2 ± 0.01 a |
| 368 day | 66.2 ± 0.01 a | 72.8 ± 0.01 a | 74.4 ± 0.03 a |
| k (a−1) | 0.52 ± 0.02 a | 0.31 ± 0.01 b | 0.31 ± 0.01 b |
Significant differences are identified with a, b, c, and d (p < 0.05) or A, B, C, and D (p < 0.01) based on the analysis of variance.
Table 3.
Correlation analysis between production or decomposition coefficient (k) of fine roots and soil characteristics (n = 9).
Table 3.
Correlation analysis between production or decomposition coefficient (k) of fine roots and soil characteristics (n = 9).
| Size Classes | Fine Root Characteristic | Fine Root Initial Indicators | Soil Indicators | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C | N | C/N | Moisture | Temp. | Total N | NO3−-N | NH4+-N | Total Ca | ||
| 0–0.5 mm | Production | 0.213 | 0.416 | 0.119 | 0.168 | 0.572 * | −0.051 | −0.004 | 0.810 ** | 0.810 ** |
| k | −0.687 * | 0.717 * | −0.701 * | 0.206 | 0.617 * | 0.241 | 0.438 | −0.585 * | −0.585 * | |
| 0.5–1 mm | Production | 0.337 | −0.290 | 0.321 | 0.227 | 0.258 | −0.267 | 0.207 | 0.855 ** | 0.855 ** |
| k | −0.669 * | 0.227 | −0.786 * | 0.356 | 0.815 ** | 0.032 | 0.497 | −0.720 * | −0.720 * | |
| 1–2 mm | Production | 0.326 | 0.417 | 0.232 | 0.339 | 0.186 | 0.111 | −0.353 | 0.054 | 0.054 |
| k | −0.671 * | 0.439 * | −0.727 * | 0.738 ** | 0.968 ** | −0.015 | 0.556 * | −0.835 ** | −0.835 ** | |
NO3−N: nitrate nitrogen, NH4+-N: ammonium nitrogen, Ca: calcium, k: decomposition coefficient. Significant correlations are indicated by * (p < 0.05) or ** (p < 0.01), based on Pearson’s correlation analysis.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shen, Y.; Wang, N.; Cheng, R.; Xiao, W.; Yang, S.; Guo, Y.; Lei, L.; Zeng, L.; Wang, X. Characteristics of Fine Roots of Pinus massoniana in the Three Gorges Reservoir Area, China. Forests 2017, 8, 183. https://doi.org/10.3390/f8060183
AMA Style
Shen Y, Wang N, Cheng R, Xiao W, Yang S, Guo Y, Lei L, Zeng L, Wang X. Characteristics of Fine Roots of Pinus massoniana in the Three Gorges Reservoir Area, China. Forests. 2017; 8(6):183. https://doi.org/10.3390/f8060183
Chicago/Turabian StyleShen, Yafei, Na Wang, Ruimei Cheng, Wenfa Xiao, Shao Yang, Yan Guo, Lei Lei, Lixiong Zeng, and Xiaorong Wang. 2017. "Characteristics of Fine Roots of Pinus massoniana in the Three Gorges Reservoir Area, China" Forests 8, no. 6: 183. https://doi.org/10.3390/f8060183
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.