Identification of Floral Relicts Based on Spatial Distance of Isolation

1
Department of Ecology, University of Debrecen, H-4032 Debrecen, Egyetem tér 1, Hungary
2
Department of Conservation Zoology, Hortobágy National Park Directorate—University of Debrecen, H-4024 Debrecen, Sumen u. 2, Hungary
3
Hungarian Academy of Sciences—University of Debrecen Biodiversity and Ecosystem Services Research Group, H-4032 Debrecen, Egyetem tér 1, Hungary
*
Author to whom correspondence should be addressed.
Forests 2017, 8(11), 459; https://doi.org/10.3390/f8110459
Submission received: 12 September 2017
/
Revised: 5 November 2017
/
Accepted: 15 November 2017
/
Published: 21 November 2017
(This article belongs to the Special Issue How Topography Impacts Forests under Global Change?)
Abstract
:The identification of climatic relicts is seldom straightforward. These species are threatened owing to current climatic trends, which underlines the importance of carrying out ecological and biogeographic investigations of them. Here we introduce a novel approach to improve the identification of climatic relicts. We are focusing on thermophilic relict plants of the Pannonian biogeographic region from the Holocene Thermal Maximum (HTM). We argue that a minimal mean annual temperature difference (MATD) of the HTM compared to the recent climate allowed a continuous northward expansion for the taxa investigated. We measured latitudinal distances between the recent occurrences of relicts and those of the main distribution found further south. Regarding estimates for MATD (1.0–2.5 °C), we only consider species with a distribution which has a 150–350 km North-South gap, since a latitudinally directed distance can be translated into temperature, showing a poleward cooling trend. Of the 15 selected species, 12 were recorded with values of 1.0–1.7 °C MATD, and three with values of 1.8–2.5 °C, some of which are presumably interglacial species. Woody species are over-represented among them (four species), using the Hungarian flora as a reference. The proposed method allows the prediction of potential climate-related changes in the future distribution of species, constrained by the topographic features of their habitats.
1. Introduction
Climatic relicts [1] are at risk, as a result of current climatic trends; some of their populations may disappear, while some may expand their ranges, underscoring the importance of ecological and biogeographical investigations of these taxa. Relicts can be either certain populations or all populations of species that are restricted in geographic range. Climatic relicts are typical biogeographic (and not taxonomic) relicts, which are descendants of once widespread taxa (or populations) that now have a narrow geographic distribution in refugia as a result of unfavourable climatic conditions acting on a significant part of their distribution. As regards species, the restriction holds true for all living populations. When there is an endangered population of a relict species (local relict), this does not necessarily mean that other populations of the species also have relict character [2]. Henceforth, we will consider climatic relict populations in this paper.
The use of the concept of climatic relicts has declined in biogeography over the past few decades [3], although the importance of the concept is evident in classical works [4]. This conceptual change is probably governed by the confusing classification of species as biogeographic relicts [5,6]: the older the supposed origin of a relict, the less clear its classification [5]. Identifying the character of a climatic relict in Central Europe [7], deciding whether it originated in an interglacial, glacial or postglacial period [2,8], or even whether it is a relict at all, is frequently problematic because of a lack of evidence. Widely distributed species in the middle of their range are not uncommonly treated as relicts, resulting in the overestimation of the proportion of relicts in the flora or fauna of a given region. Zahariev summarized the current knowledge of the Bulgarian vascular plant relicts [9], mentioning 183 taxa with a tertiary origin (before the Pleistocene), a number which seems to be far too high (approximately (ca.) 5% of the whole flora) in a country located in the temperate climatic zone, as compared to the 143 younger relicts on the list. Most of the mentioned species classified as Tertiary relicts are widespread in Europe, such as Cypripedium calceolus L., Fraxinus excelsior L., or widespread in Central and South Europe, such as Acer tataricum L., which makes their Bulgarian Tertiary relict status questionable.
Assuming an edge population of an investigated taxon to be a relict when it is in fact widespread and frequently occurring in the close neighbourhood casts doubt on its relict status. Kaplan and his colleagues [10,11] provided distribution maps of various species considered postglacial relicts in Bohemia, such as Achillea collina J. Becker ex Reichenb., Linum flavum L., Viola ambigua Waldst. & Kit. These species are sporadic in South-Bohemia and frequent in Northwest Hungary [12], only ca. 100 km to the South-East. Most of these species—such as Linum flavum [13] and Viola ambigua [14]—also occur in the region between Bohemia and Hungary, i.e., in Slovakia. Achillea collina is one of the most frequent generalist herbaceous species on dry grassland habitats in Hungary. These taxa can be relicts only if the Bohemian occurrences are in a proximal position or located in ecotonal microrefugia [15] in the proximity of their main distribution. Without topographic barriers, and considering their preferred habitats, this is not very likely [16].
Hájková and her colleagues considered a number of species with disjunct distributions on the hay-meadows of the Bílé Karpaty Mts. to be ancient relicts [17]. However, the authors do not provide references and specifying species or measurements of disjunctions. In contrast, Kaplan [10] claims that in the Bohemian flora, all relicts present in the area have a Quaternary origin—more precisely a Late Quaternary origin—suggesting the absence of ancient relicts in this region.
Over the past two decades, studies in molecular genetics have attempted to clarify unclear details of historical biogeography [18,19], in the process establishing the field of phylogeography [20], although the results are often no less confusing than those obtained by biogeographic approaches [21]. The revitalisation of the concept of relicts [15,22] has also begun with the application of other approaches such as those applied by Hájková and her colleagues [6], who used paleontological methods. To find species (vascular plants and snails), which are restricted to old, post-glacial fens, thus indicating post-glacial relict status, they aged fens and then compared plant and snail species lists of these different-aged fens. However, these types of paleontological methods are not always applicable [16], because the preferred habitats of the investigated species are dry and warm, which makes them unsuitable for fossilisation processes.
Our assumptions for considering populations as climatic relicts are:
- An inner position in the continuous distribution of a species does not imply relict occurrence.
- An edge position on a continuous distribution can imply relict occurrence, although with a very low probability.
- An isolated occurrence from a large, continuous distribution but close to it can be regarded as a relict with a slightly higher probability.
- An isolated occurrence far from a large, continuous distribution can be regarded as a relict occurrence with much a higher probability than in the former cases.
We outline here a historical biogeographic method which provides a panbiogeographic [23] approach, and we (i) identify relict status and (ii) estimate the age of the origin of plant species in the Pannonian region, focusing on thermophilic relict populations supposedly from the Holocene Thermal Maximum (HTM).
Besides thermal relicts, several other types of relicts are present in the Carpatho-Pannon region e.g., so-called steppe-relicts, such as Gagea szowitsii (A. F. Láng) Besser, whose range is located further east following a 600 km gap, in Dobrogea around the Black Sea, or Nepeta parviflora Bieb. with an 800 km wide disjunction in this direction. Similarly, Artemisia laciniata Willd., or Krascheninnikovia ceratoides (L.) Gueldenst. are probably cold mammoth-steppe relicts, showing hundreds of kilometres of east-west disjunction in their distribution. Regarding their status, some ancient endemic species of the region are between biogeographic and taxonomic relicts such as Syringa josikaea Jacq. fil. ex Reichenb. or Leucanthemella serotina (L.) Tzvelev.
We considered south-north orientated disjunctions to be the focus of our research, because it is widely accepted that the majority of the holothermic or thermal relicts of Europe derive mainly from Mediterranean refugial populations which later experienced range expansion northwards in the early post-glacial periods [24,25]. Our investigated species are not remnants of any drier or more humid climatic periods, and their habitats are neither extremely dry or nor extremely wet.
We suppose that the HTM with its temperature differences from recent values left characteristic fingerprints on the vegetation patterns as shown by distal or remote microrefugial occurrences [15]. We investigate horizontal distances between isolated occurrences and recent continuous distributions, emerging from the retreat of the HTM-distribution with poleward extensions in the past. The baseline is provided by the existence of area disjunctions which often indicate the relict status of species [15,26]. The latitudinally directed, North-South distance (N-SD) component of the disjunction can be translated into temperature differences, based on a poleward latitudinal cooling trend at global scales, calculated as 6.9 °C/1000 km [27,28]. The HTM was followed by a cooling period which resulted in the retreat of the area of the investigated thermophilic species. Established estimates for MATD (Mean Annual Temperature Difference) between the HTM and recent times range between 1 and 2.5 °C [29,30], thus a gap of 150–300 km satisfies the requirement of a distant, isolated occurrence with a potential HTM origin. If the regression left relict and patchy occurrences, the latitudinal component of the distance between the relict patch and the continuous regressed area indicates the minimal northern extent of the original distribution during the HTM. Thus, we suppose that isolated occurrences at great distances are attributable to the simple environmental factor of temperature. This might imply that the distribution ranged even further poleward, but that recently the examined isolated occurrences are the northernmost ones (Figure 1).
Our study questions are the following:
- (1)
- Which species, as candidates for the status of an HTM (or postglacial) relict, satisfy the criteria of N-SD of 150–350 km in the Pannonian flora? We expect a limited number of such species.
- (2)
- How is MATD distributed among these taxa? The distribution of N-SD values, if they are not evenly distributed, can be informative with respect to assessing the exact value of MATD within the wider range (of 1.0–2.5 °C) or with regard to judging whether some relicts originated from earlier interglacial periods.
2. Materials and Methods
The choice of MAT as an effective temperature parameter is justifiable on the basis of recent results of studies using species distribution models (SDM) which show that averages play an important role in these phenomena, often more important than the extremes themselves [31,32,33]. Furthermore, as the distributions of most vascular plant species are nested in certain vegetation types (e.g., Mediterranean woodland, temperate deciduous broad-leaved forest, etc.), and the climatic background of these vegetation types is described globally by climatic diagrams which use average and not extreme data [34], we also omit temperature-extremes. Temperature extremes themselves can be important in the limitation of certain species which are usually widely distributed. Some articles emphasize the importance of these extreme parameters, as in Woodward [35], but others such as [36,37,38] found averages of years or certain months to have greater or equal importance. On the other hand, climatic extremes are rarely independent from average values: within a range of the same continentality the relationship is usually linear: the warmer the MAT, the higher the yearly absolute and average maximum and minimum temperature values.
At middle latitudes, the northward decline of the MAT is measured as ca. 6.9 °C/1000 km [27,28]. For instance, when we calculate for a 500 km N-SD of a disjunction, this value indicates a minimum 0.69 × 5 = 3.45 °C MATD.
Our approach is macroecological [39], hence once a population has survived in a microrefugium, the fine-scale dynamics and habitat-heterogeneity [1] of the microrefugia is not the focus of our attention; only large scale habitat patterns of the recent distribution-gap are important. The estimations are based on the following assumptions:
- The HTM-expansion was not a rapid process (anemochory and zoochory are excluded);
- The area of the gap is flat or hilly, where micro- and mesoclimatic effects are weak, or, if the investigated species depends on habitats influenced by the micro- and mesoclimate, then their effect is evenly distributed;
- The disjunction is not the result of human impacts and re-occupation was not limited by anthropogenic influences.
We identified Pannonian species (2200–2300 native species in Hungary and ca. 3000 altogether in the basin) likely to be early Holocene relicts according to our approach. In doing so, we selected taxa which have disjunctions of more than 150 km latitudinal gap-distance (ca. 1.4° latitude) to indicate a minimal MATD of the HTM (ca. 1 °C), and where the size of the gap is less than 350 km (ca. 3.0–3.5° latitude), indicating cooler conditions than a maximum of 2.5 °C MATD. As another important criterion we specified that the isolated occurrence should be the northernmost one at least on a regional scale (in Central Europe), as the supposed northernmost HTM-edge of the area is fundamentally important in our approach. Interglacial relicts (indicating a MATD greater than 2.5 °C) are not dealt with in our research, as an MATD value of 2.5 °C is the highest reliable paleontological estimation in most paleontological reconstructions [29,30,40,41] of the HTM. A higher MATD probably indicates an origin from a warmer, interglacial period.
We divided the interval of 1–2.5 °C into two equal subintervals: 1–1.7 °C, corresponding to the HTM MATD estimates of Feurdean et al. [40] and Molnár and Végvári [29], and 1.8–2.5 °C, corresponding to the HTM MATD estimates of Magyari et al. [41] and Tóth et al. [30], and compared the number of observations from the two intervals. The lack of observations from the second subinterval does not imply that the HTM MATD estimates with larger maximum values are necessarily wrong, because the MATD values in the current analysis are minimum values. If some observations fall within the second subinterval, but less so than in the first one, they may be either relicts from an earlier period (supposing that the lower HTM MATD estimates are correct), or HTM relicts (preserving the wider HTM MATD interval). If there are several observations in the second subinterval, i.e., as many as in the first one, those observations with MATD values between 1.8 and 2.5 °C are most probably HTM relicts, because the number of earlier relicts must be lower than those originating in the HTM, given that their survival must be accidental in the Pannonian region. In this case, the HTM MATD estimates with smaller maximum values can be questioned. In our study, we also considered those known occurrences of the investigated species which have disappeared over the past years or decades as a result of human impacts.
The Pannonian region is the largest basin in Europe and has global significance, covering ca. 250,000 km2, comparable in size with the Great Basin of North-America, and extending ca. 10° longitudinally and 4° latitudinally. Furthermore, most of the region is lowland or hills not higher than 400–500 m, hence the mesoclimatic influences are less diverse.
The latitudinal changing trends of the MAT are unambiguous in Eastern Europe but questionable further west. However, the extent and location of our region allows us to use East-West parallel isotherm lines as a starting point. This basin is located far from the Atlantic coast where the January isotherm-line runs entirely North-South around the European oceanic coast. The size of the Pannonian region, the weak climatic effect of the Atlantic Ocean and the moderately pronounced topography allow an almost parallel East-West isotherm structure for the basin on a large scale. Supplementary Table S1 illustrates differences in the MAT on a north-south gradient in the Pannonian region schematically. The approximately calculated northward decline of the MAT is estimated at 0.76 °C/100 km, while with the altitudinal correction of the MAT using the latitude-lapse rate value of 6.5 °C/1000 m [42] the decline amounts to 0.57 °C/100 km, which is similar to the cited value of 0.69 °C/100 km [27,28]. In our subsequent calculations we use the statistically established 0.69 °C/100 km value.
Firstly, we assessed the flora of Hungary, which occupies almost 50% of the area of the Pannonian biogeographic region. To search for a continuous area of the species to be selected we devoted special attention to the flora of Croatia, because most of the identified taxa are found in W-Hungary, and their continuous distribution can be located with a high probability further south. The orographic features of central and N-Croatia as well as W-Hungary are very similar: an even mixture of hills, lower mountains and smaller basins. When the continuous areas of some species occur further to the southeast in Bosnia and Herzegovina, in Serbia or in Romania, these floras were also considered.
The selection of species was based on the Hungarian flora [43], supplemented with the recently completed floral map of Hungary [12] and the open access floral map of Croatia (http://hirc.botanic.hr/fcd/). In the case of the eastern part of the Pannonian region, (the small basins and hills of Transylvania), we used the flora-monography of Romania [44,45,46,47,48] and the red data book of the Romanian flora [49]. Volumes of the Atlas Florae Europaeae [50,51,52,53,54] were also used. The specific literature items for the most important continuous occurrences in Bosnia and Herzegovina, in Serbia or in Romania are cited in the species list.
We selected species which have the expected N-SD value by examining the grid maps of the distribution of each species. Although some species show gaps inside Hungary and Romania, most of them show this gap only if we use the pooled distribution maps of the neighbouring countries.
3. Results
The following 15 species met our criteria (woody species are marked with an asterisk):
As regards their Raunkiaer life-form, four of the species are woody (two phanerophytes and two chamaephytes). In the Hungarian flora, the ratio of woody species is 9.3% (5.9% phanerophytes and 3.4% chamaephytes) [43]. This constitutes 65–70% of the Pannonian flora. Thus, woody species are overrepresented among the assumed HTM-relicts.
Most of the 15 species listed above occupy south facing rocky or grass-covered slopes and thermophylous Quercus pubescens Willd. woods. The lowlands of the Pannonian region are dominated by floodplains and cannot provide suitable habitats for them. Consequently, these species probably dispersed across landscapes dominated by the warm slopes of hills or low mountains, and not across lowlands. Members of our set are widespread further south in the Balkan Peninsula but absent or rare on the Mediterranean coast of the nearby Adriatic Sea.
As far as we know, Vincetoxicum pannonicum is strictly endemic, restricted to the Pannonian region, and to Hungary. Its closest living relative is V. fuscatum (Hornem.) Reichenb. fil., which is widespread in the Balkan Peninsula. Our distance data in this case indicate the N-SD between the only two, very local occurrences and no difference between a poleward patch and a larger continuous occurrence, as recently this species only has microrefugial occurrences [16].
Recent investigations show that the taxonomic status of the former Onosma tornensis Jáv. (treated as local endemic taxa) is correctly O. viridis [61]; therefore this species falls into the category of the Balkan floral elements.
We excluded maritime species (such as Schoenoplectus litoralis (Schrader) Palla) which occur on alkaline soils of the Pannonian lowlands as potential HTM relicts. Most of these species can disperse by intensive zoochory in jumps (dispersed by shorebirds, ducks and other waterbirds; see the first assumption in the Methods section). Although these taxa show 200–500 km gaps in their distribution, ancient colonisation processes can be excluded because these gaps are characterized by the absence of suitable habitats for them in the Balkan Mountains, such as shallow sodic lakes, or smaller lakes supplied by geothermal water and evaporites on the surface.
When quantifying N-SD accurately, we need to outline probable paths of dispersion (or tracks in a panbiogeographic approach). Ten species have one possible track. Five of these ten taxa (Carpinus orientalis, Cuscuta approximata, Physospermum cornubiense, Ranunculus psilostachys, Trigonella gladiata,) occur in West Hungary, west of the River Danube. For these plants, and for the NE-Hungarian Micromeria thymifolia, theroretical tracks from Croatia or from the Fruška Gora Mts. in Serbia are highly probable as suitable landscapes are continuous in this direction. Regarding the other four species of these ten, we suppose Vincetoxicum pannonicum has a similar simple path between the two occurrences in a south-to-north direction. Hypericum umbellatum, Ilex aquifolium and Saponaria bellidifolia are distributed in W-Transylvania and their tracks are directed towards the eastern part of the Balkan Peninsula.
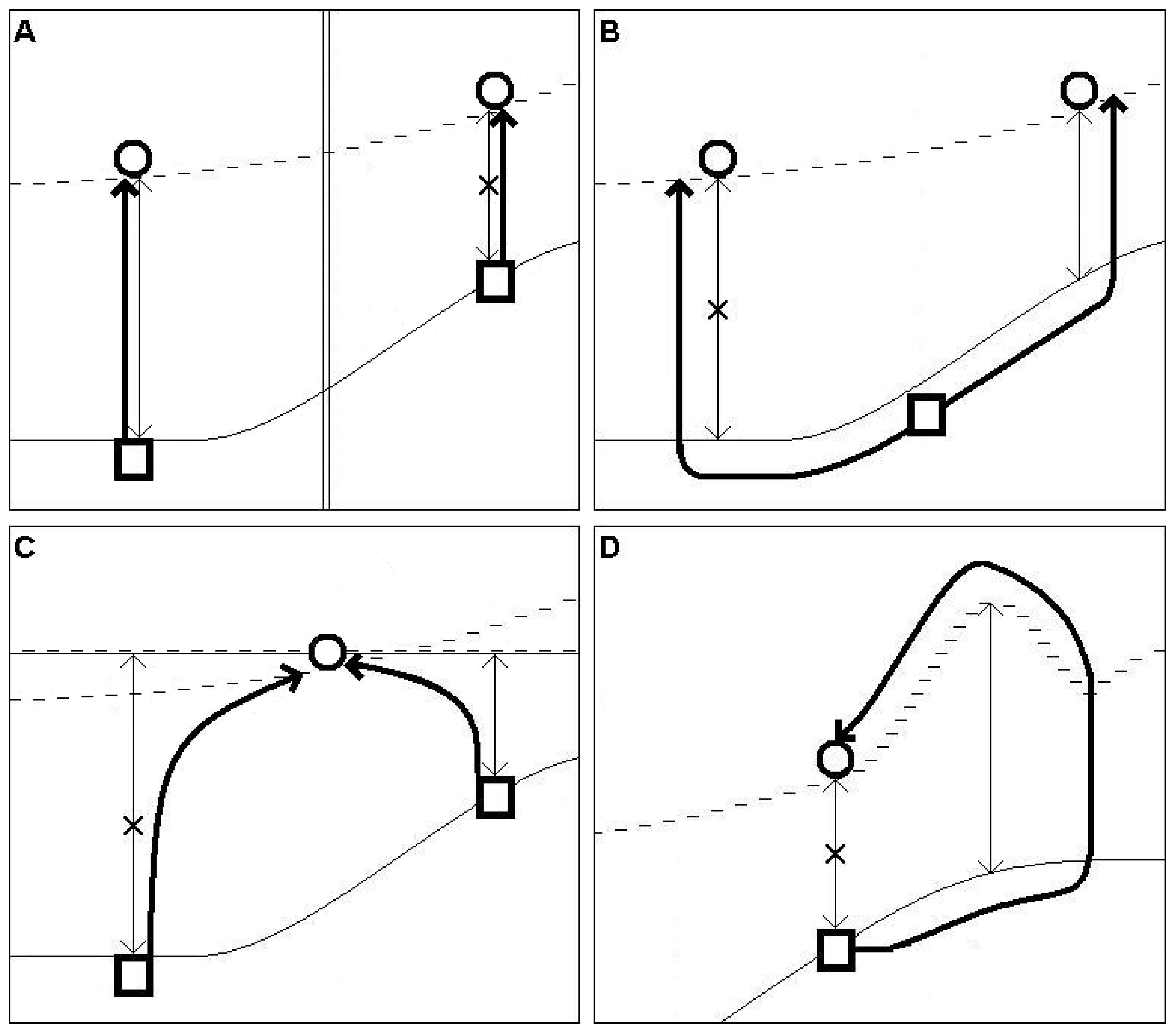
When more than one track is likely (relevant for five species), the identification of probable paths requires a panbiogeographic approach [23], i.e., analyses of the topographical features of the area where the disjunction appears. Possible routes, shaped by barriers and distances to overcome around the connecting zone of the Balkan and the Pannonian region, should be analyzed. The theoretical types of the former migration-routes of these 5 species are introduced in Figure 2 (multiple track-structure). Four different types can be identified. (A) Two possible tracks are separated by an effective N-S directed barrier and two separated isolations have survived. In this case, both tracks had to function because the tracks were continuously separated by the N-S directed barrier and W-E directed crossings never occurred, that is why we select the track with the greater gap-distance as exemplified by Aspleniuim lepidum. (B) Two separated isolations have survived. As there is no barrier between them, it is very possible that the taxon moved through the shorter gap as this was easier. In our study, there is no example of this case. (C) Only one isolation survived and this could be reached via tracks from different directions. Therefore, we select the root which has the smallest gap-distance, exemplified by Aethionema saxatile and Silene flavescens. (D) The taxa in this group have special habitat-requirements, and track these habitats. The necessary habitat-type goes far beyond the recent isolation poleward, and the taxa had to reach the recent isolation through this point. Thus, we consider the gap between the northernmost point of the suitable habitat-belt and the southern distribution of the recently occupied habitats. Note, that all mentioned taxa in categories A or C also fall into category D.
Supplementary Figure S1 demonstrates types C and D for Silene flavescens in detail. The supplementary material also provides detailed information regarding all five species with multiple tracks.
Supplementary Table S2 shows the alternative tracks for these five species and the supplementary text before the table describes the tracks by names of geographical sites.
Table 1 shows the N-SD data with the respective MATD data for all of the 15 species. The rounding is set to 0.1° in latitude, because the relationship between the latitude and temperature is 0.1° ≈ 0.07 °C and so a more accurate distinction is not necessary.
4. Discussion
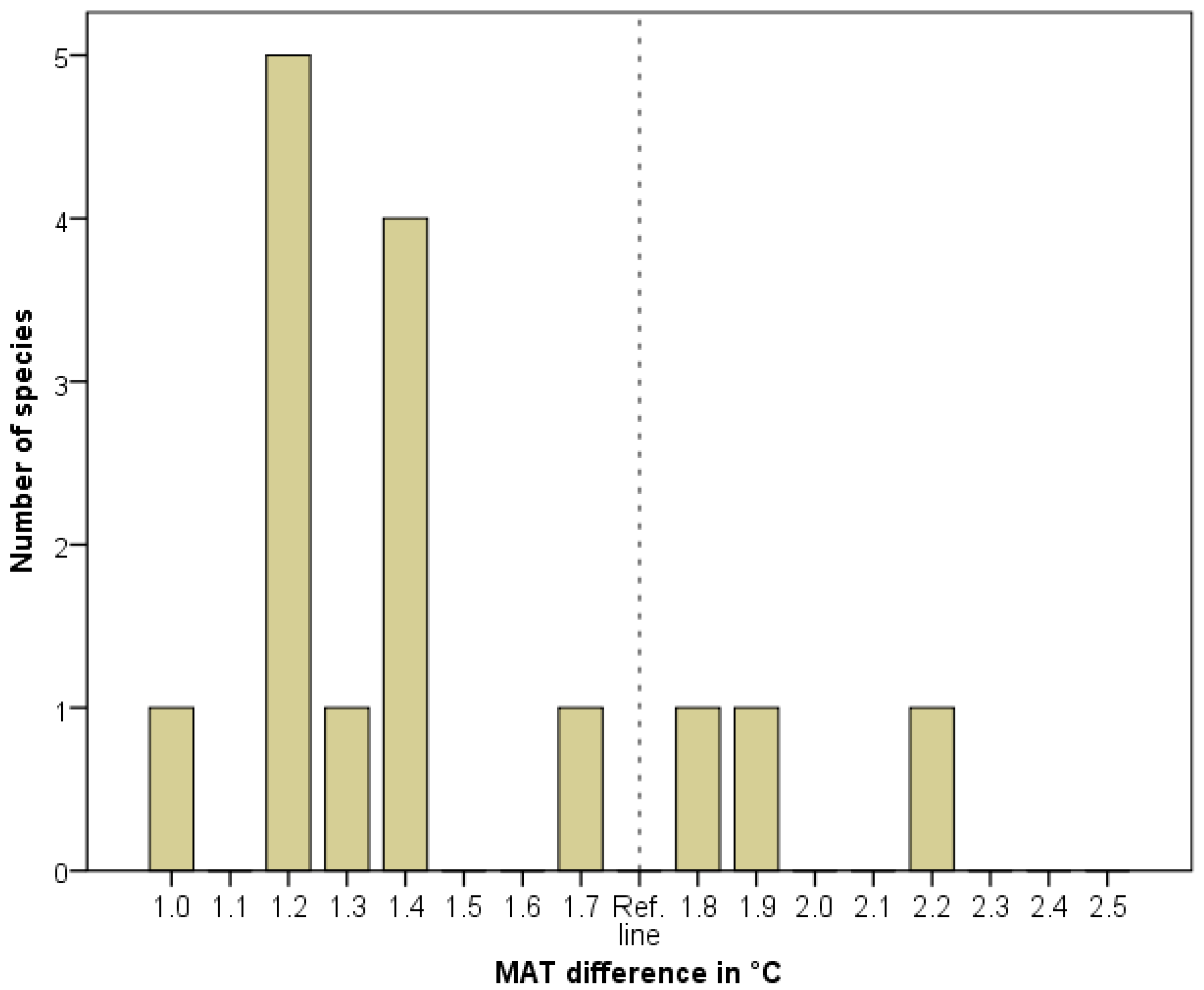
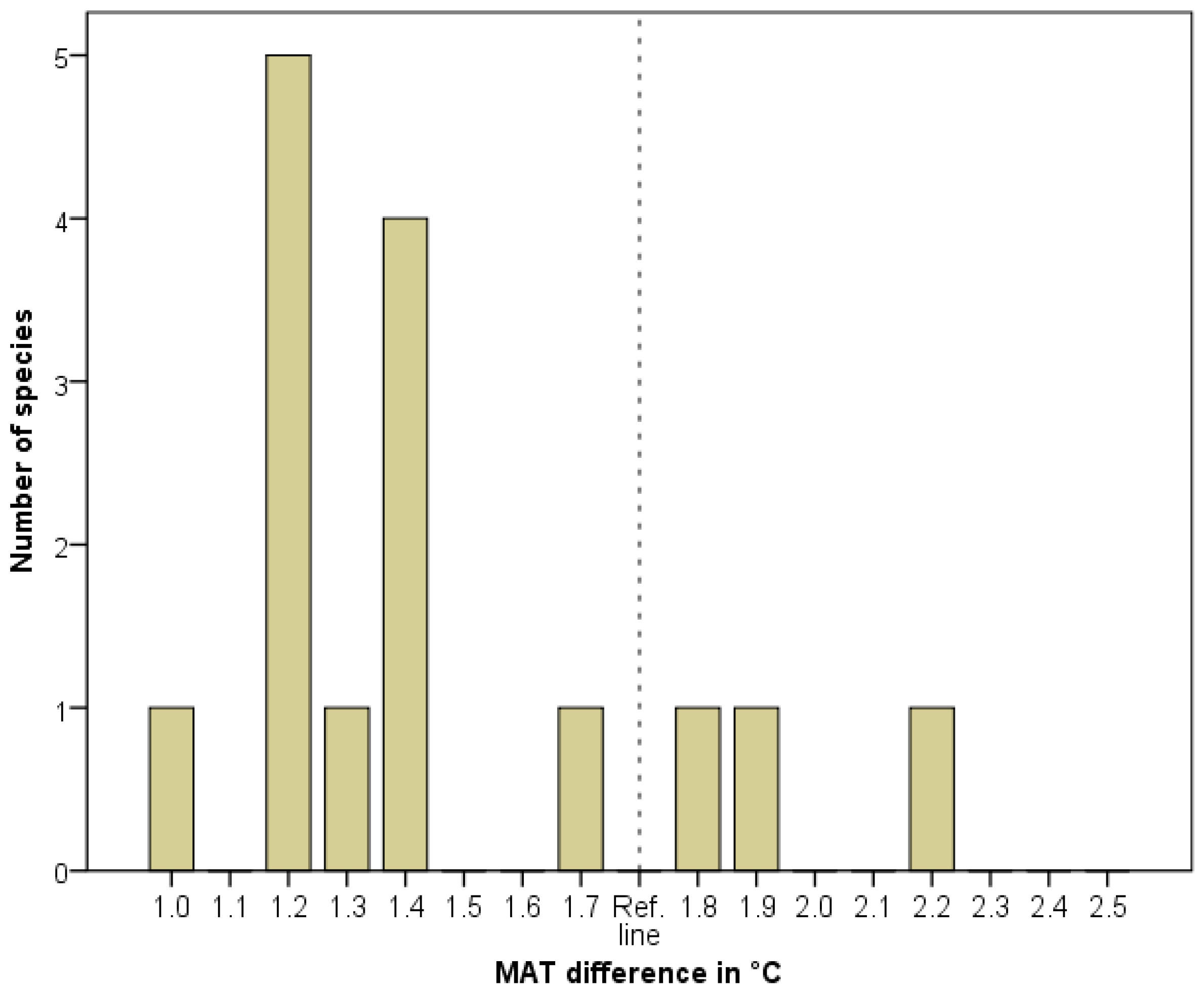
We developed strict criteria for identifying a population as an HTM relict; thus the real number of HTM relicts in a certain flora may be higher. In the Pannonian flora, we identified 15 species fulfilling the HTM relict criteria. Twelve species have an MATD of between 1.0 and 1.7 °C. Three species (Hypericum umbellatum, Physospermum cornubiense and Silene flavescens) show MATD values between 1.8 and 2.5 °C (Figure 4).
We evaluated the distribution of the MATD values using a binomial test. The results show a significantly higher number of species (12) in the lower MATD sub-interval (1.0–1.7 °C), p = 0.04. There are no species with an MATD > 2.2 °C in the whole interval. These findings did not confirm that paleontological estimates with large maximum values of HTM MATD [30,41] were the only supported scenarios. Nevertheless, we could not exclude higher values. The three species indicating high MATD (Hypericum umbellatum, Physospermum cornubiense and Silene flavescens) are potential interglacial relicts in the case of low MATD estimations [29,40], or HTM relicts in the case of high values. This problem may call for the use of molecular genetic approaches.
When the number of plants with a potential HTM origin was high, further statistical analysis of the distribution of the data would be possible to provide additional information on the background of their origin and their recent biogeography. This characteristic could be more specifically measurable when investigating floras with many more relict species than the Pannonian flora, such as, for example, those in the southeastern part of North America, eastern China, Ukraine and SW-Russia or the southern part of the Balkan Peninsula.
Hungarian local botanical literature considers some species interglacial relicts, at least from the Eemian period. Most of them occur west of the River Danube and south of Lake Balaton, including Colchicum hungaricum Janka, Medicago orbicularis (L.) Bartal., Orobanche nana (Reuter) Noë ex G. Beck and Stipa bromoides (L.) Dörfler. The nearest occurrences of these plants are located on the Mediterranean coast of the Adriatic Sea further to the south (not in maritime habitats) or further south-southeast in the Balkan Peninsula but at a much larger distance. To reach the Pannonian region from the Mediterranean, it is necessary to cross the Dinarian Mountains, which are rich in warm rocky slopes. In these mountains, during the period when these species arrived in the Pannonian region, the sub-Mediterranean climate necessarily reached at least to an altitude of 1000–1200 m above sea level (the altitude of the lowest passes), compared to the recent 400–600 m height of the sub-Mediterranean zone in this area. The 400–800 m difference implies 3–5 °C as a minimum value of MATD (considering the adiabatic lapse rate) which is higher than the estimated MATD of the HTM in any formerly mentioned paleontological scenario, and clearly indicates an Eemian (or earlier interglacial) origin of these species. Because MATD estimates are minimum values in our approach, the fact that MATD values for the three species with potential Eemian origin (Hypericum umbellatum, Physospermum cornubiense and Silene flavescens) are lower than 3–5 °C classifying them as species with interglacial origin is still allowed.
Regarding applicability, our approach can be helpful in the selection of candidate taxa for phylogeographic research when various questions emerge, such as, for example, the question of whether it is more typical for Eemian-originated taxa in the Pannonian vascular flora to evolve to species, or, to a lesser extent, to subspecies, or only to irrelevant cladistic differences. In this case, we could propose that in Hungary, investigation of the most critical 4 + 3 taxa (the four interglacial-aged relicts and the three species with a high MATD-value from our list) could be systematically supplemented with interglacial-originated Pannonian endemics [8,62] and compared with those 12 species from our list that are remnants from the HTM with a high probability. On the other hand, the distance analysis can offer reference points in timing slowly evolving relicts, where molecular taxonomy cannot provide convincing results. Oeyen and his colleagues tested the “speciation by distance” (both in space and time) phylogeographic hypothesis in the Pannonian region by combining topographic and molecular genetic approaches on the Mediterranean Banded Centipede (Scolopendra cingulata Latreille) [63]. The authors studied the rediscovered Austrian and the already known Hungarian and Romanian occurrences as examples isolated by great distance, genetically comparing them to the populations of the continuous Mediterranean coastal areas. This study failed to detect a positive correlation between geographic and genetic distance. In this case, the distance variable was derived from the direct distance data between the locations of isolated and main population occurrences, and not from the N-SD values of the occurrences.
If Hypericum umbellatum, Physospermum cornubiense and Silene flavescens are Eemian relicts, they are Pannonian interglacial relicts with a southern continuous distribution not falling into the Mediterranean zone but into the warm-temperate, sub-Mediterranean vegetation and climatic zone. The four established Eemian relicts of the western part of the Pannonian region have distribution and habitats which are rather Mediterranean in character. Stipa bromoides is a Mediterranean grass, but rarely occurs in the sub-Mediterranean climatic zone too. The other three species, Colchicum hungaricum, Medicago orbicularis and Orobanche nana are unambiguously Mediterranean species. Considering current climatic trends, we predict that HTM relicts will expand their range faster than interglacial ones, as warming tendencies can fulfil the higher thermal optimum of the latter taxa only later. However, this process might be significantly altered by human disturbance of critical habitats, in our case, in Central Europe. Species which are less tolerant of disturbance, and prefer habitats, reduced by humans, can utilize the possibilities of the warming less effectively (even if they are younger, HTM-relicts) than disturbance-tolerating, habitat-generalist species (even if they are old, interglacial relicts). A scenario of a lose-lose outcome is also possible if the negative effect of growing climatic fluctuations, extremities and uncertainties overrides the direct impact of warming. The future tracks of expansions are limited by topographic constraints because the species examined are inhabitants of topographically determined, microclimatically affected habitats.
5. Conclusions
In our study, we revealed the importance of the North-South component of distance between northernmost, isolated occurrences of vascular plant species and their closest continuous distributional area, when identifying relict species and dating their origins. Using our method, we found 15 species in the Pannonian flora where there was a fairly high probability that they reached this region during the Holocene Thermal Maximum (HTM) period. Based on the extent of the disjunctions three out of these 15 species may have reached the Pannonian region during even an earlier, interglacial period. Compared to the whole Hungarian flora, woody species are over-represented among these 15 species.
In the near future several of these species may expand their range because of climate warming. HTM relicts are expected to follow this pattern with a higher probability than interglacial ones, as climatic trends are predicted to realize their thermal optimum sooner. The future tracks of expansion are limited by topographic constraints.
Supplementary Materials
The following are available online at www.mdpi.com/1999-4907/8/11/459/s1, Figure S1: Possible (simplified) migration routes (or tracks) for the Silene flavescens until the HTM (or an earlier warmer climatic period), Table S1: MAT values in the meteorological stations of the Pannonian region from north to south, Table S2: The 5 ‘more-than-one-way’ species with their potential migration routes.
Acknowledgments
Research was partly supported by OTKA K 116639 project.
Author Contributions
Attila Molnár, Béla Tóthmérész and Zsolt Végvári conceived and designed the study; Attila Molnár performed the study and analyzed the data. Attila Molnár, Zsolt Végvári and Béla Tóthmérész wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hampe, A.; Jump, A.S. Climate relicts: Past, present, future. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 313–333. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Lomolino, M.V. Biogeography; Cambridge University Press: Sunderland, MA, USA, 1998; p. 300. [Google Scholar]
- Croizat, L. Manual of Phytogeography; Springer-Science + Business Media: Amsterdam, The Netherlands, 1952. [Google Scholar]
- Cox, C.B.; Moore, P.D.; Marquardt, W.C.; Demaree, R.S.; Grieve, R.B. Biogeography: An Ecological and Evolutionary Approach, 6th ed.; Blackwell Publishing: Hoboken, NJ, USA, 2005. [Google Scholar]
- Habel, J.C.; Assmann, T. (Eds.) Relict Species: Phylogeography and Conservation Biology; Springer Science & Business Media: New York, NY, USA, 2009; p. 324. [Google Scholar]
- Hájková, P.; Horsák, M.; Hájek, M.; Jankovská, V.; Jamrichová, E.; Moutelíková, J. Using multi-proxy palaeoecology to test a relict status of refugial populations of calcareous-fen species in the Western Carpathians. Holocene 2015, 25, 702–715. [Google Scholar] [CrossRef]
- Huggett, R.J. Fundamentals of Biogeography, 2nd ed.; Routledge: Abingdon-on-Thames, UK, 2004; pp. 62–63. [Google Scholar]
- Soó, R. The history of the vegetation of Hungary. Collect. Bot. 1968, 7, 1109–1120. [Google Scholar]
- Zahariev, D. Biodiversity of Relict Vascular Plants in Bulgaria. Int. J. Res. Stud. Biosci. 2016, 4, 38–51. [Google Scholar]
- Kaplan, Z. Flora and phytogeography of the Czech Republic. Dedicated to the centenary of the Czech Botanical Society (1912–2012). Preslia 2012, 84, 505–573. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Štěpánková, J.; Bureš, P.; Zázvorka, J.; Hroudová, Z.; Ducháček, M.; Grulich, V.; Řepka, R.; Dančáket, M.; et al. Distributions of vascular plants in the Czech Republic. Part 1. Preslia 2015, 87, 417–500. [Google Scholar]
- Bartha, D.; Király, G. (Eds.) Distribution Atlas of Vascular Plants of Hungary; University of West Hungary Press: Sopron, Hungary, 2015; pp. 1–329. [Google Scholar]
- Futák, J.; Bertová, L. Flóra Slovenska III; VEDA, Vydavateľstvo Slovenskej Akadémie Vied: Bratislava, Slovakia, 1982; p. 519. [Google Scholar]
- Goliašová, K.; Šípošová, H. (Eds.) Flóra Slovenska VI/1; VEDA Vydavateľstvo Slovenskej Akadémie Vied: Bratislava, Slovakia, 2008; p. 170. [Google Scholar]
- Rull, V. Microrefugia. J. Biogeogr. 2009, 36, 481–484. [Google Scholar] [CrossRef]
- Birks, H.J.B. Some reflections on the refugium concept and its terminology in historical biogeography, contemporary ecology and global-change biology. Biodiversity 2015, 16, 196–212. [Google Scholar] [CrossRef]
- Hájková, P.; Roleček, J.; Hájek, M.; Horsák, M.; Fajmon, K.; Polák, M.; Jamrichová, E. Prehistoric origin of the extremely species-rich semi-dry grasslands in the Bílé Karpaty Mts (Czech Republic and Slovakia). Preslia 2011, 83, 185–204. [Google Scholar]
- Riddle, B.R.; Dawson, M.N.; Hadly, E.A.; Hafner, D.J.; Hickerson, M.J.; Mantooth, S.J.; Yoder, A.D. The role of molecular genetics in sculpting the future of integrative biogeography. Prog. Phys. Geogr. 2008, 32, 173–202. [Google Scholar] [CrossRef]
- Roderick, G.K. Geographic structure of insect populations: Gene flow, phylogeography, and their uses. Annu. Rev. Entomol. 1996, 41, 325–352. [Google Scholar] [CrossRef] [PubMed]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, UK, 2000; pp. 3–36. [Google Scholar]
- Collevatti, R.G.; Rabelo, S.G.; Vieira, R.F. Phylogeography and disjunct distribution in Lychnophora ericoides (Asteraceae), an endangered cerrado shrub species. Ann. Bot.-Lond. 2009, 104, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Gentili, R.; Bacchetta, G.; Fenu, G.; Cogoni, D.; Abeli, T.; Rossi, G.; Salvatore, M.C.; Baroni, C.; Citterio, S. From cold to warm-stage refugia for boreo-alpine plants in southern European and Mediterranean mountains: The last chance to survive or an opportunity for speciation? Biodiversity 2015, 16, 247–261. [Google Scholar] [CrossRef]
- Craw, R.C.; Grehan, J.R.; Heads, M.J. Panbiogeography: Tracking the History of Life; Oxford University Press: Oxford, UK, 1999; pp. 4–6. [Google Scholar]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and postglacial trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z. Extra-Mediterranean Refugia, Post-Glacial Vegetation History and Area Dynamics in Eastern Central Europe. In Relict Species: Phylogeography and Conservation Biology; Habel, J.C., Assmann, T., Eds.; Springer Science & Business Media: New York, NY, USA, 2009; pp. 57–87. [Google Scholar]
- Kwiatkowski, P.; Krahulec, F. Disjunct distribution patterns in vascular flora of the Sudetes. Ann. Bot. Fenn. 2016, 53, 91–102. [Google Scholar] [CrossRef]
- Chen, J.; Wan, S.; Henebry, G.; Qi, J.; Gutman, G.; Sun, G.; Kappas, M. (Eds.) Dryland East Asia: Land Dynamics Amid Social and Climate Change; Walter de Gruyter: Berlin, Germany, 2014; p. 251. [Google Scholar]
- Jump, A.; Mátyás, C.; Penuelas, J. The paradox of altitude for latitude comparisons in species range retractions. Trends Ecol. Evol. 2009, 24, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Molnár, A.; Végvári, Z. Reconstruction of early Holocene Thermal Maximum temperatures using present vertical distribution of conifers in the Pannonian region (SE Central Europe). Holocene 2016, 27, 236–245. [Google Scholar] [CrossRef]
- Tóth, M.; Magyari, E.; Buczkó, K.; Braun, M.; Panagiotopoulos, K.; Heiri, O. Chironomid-inferred Holocene temperature changes in the South Carpathians (Romania). Holocene 2015, 25, 569–582. [Google Scholar] [CrossRef] [Green Version]
- Dobbertin, M.; Hilker, N.; Rebetez, M.; Zimmermann, N.E.; Wohlgemuth, T.; Rigling, A. The upward shift in altitude of pine mistletoe (Viscum album ssp. austriacum) in Switzerland—The result of climate warming? Int. J. Biometeorol. 2005, 50, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Rasztovits, E.; Moricz, N.; Berki, I.; Poetzelsberger, E.; Matyas, C. Evaluating the performance of stochastic distribution models for European beech at low-elevation xeric limits. Időjárás 2012, 116, 173–194. [Google Scholar]
- Ruiz-Labourdette, D.; Schmitz, M.F.; Pineda, F.D. Changes in tree species composition in Mediterranean mountains under climate change: Indicators for conservation planning. Ecol. Indic. 2013, 24, 310–323. [Google Scholar] [CrossRef]
- Walter, H.; Lieth, H. Klimadiagramm-Weltatlas; Gustav Fisher Verlag: Vienna, Austria, 1960. [Google Scholar]
- Woodward, F.I. Temperature and the distribution of plant species. Symp. Soc. Exp. Biol. 1988, 42, 59–75. [Google Scholar] [PubMed]
- Hocker, H.W. Certain aspects of climate as related to the distribution of Loblolly Pine. Ecology 1956, 37, 824–834. [Google Scholar] [CrossRef]
- Molnár, A.; Végvári, Z. Bioclimatic constraints of European mistletoe Viscum album at its southern distribution limit at past and present temporal scales, Pannonian Basin, Hungary. Clim. Res. 2017, 71, 237–248. [Google Scholar] [CrossRef]
- Root, T. Energy constraints on avian distributions and abundances. Ecology 1988, 69, 330–339. [Google Scholar] [CrossRef]
- Brown, J.H. Macroecology; The University of Chicago Press: Chicago, IL, USA, 1995. [Google Scholar]
- Feurdean, A.; Perşoiu, A.; Tanţău, I.; Stevens, T.; Magyari, E.K.; Onac, B.P.; Markovićh, S.; Andriči, M.; Connorj, S.; Fărcaş, S.; et al. Climate variability and associated vegetation response throughout Central and Eastern Europe (CEE) between 60 and 8 ka. Quat. Sci. Rev. 2014, 106, 206–224. [Google Scholar] [CrossRef] [Green Version]
- Magyari, E.K.; Jakab, G.; Bálint, M.; Kernd, Z.; Buczkóe, K.; Braunf, M. Rapid vegetation response to Lateglacial and early Holocene climatic fluctuation in the South Carpathian Mountains (Romania). Quat. Sci. Rev. 2012, 35, 116–130. [Google Scholar] [CrossRef]
- Gabler, R.; Petersen, J.; Trapasso, L.; Dorothy, M.S. Physical Geography, 9th ed.; Brooks/Cole, Cengage Learning: Belmont, CA, USA, 2008. [Google Scholar]
- Soó, R. A Magyar Flóra és Vegetáció Rendszertani-Növényföldrajzi Kézikönyve. (Handbook of the Flora and Vegetation of HUNGARY) I–VII; Akadémiai Kiadó: Budapest, Hungary, 1964–1985.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. I; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1952; p. 136.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. II; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1953; p. 155.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. III; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1955; pp. 404–405.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. IV; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1956; pp. 41, 622.
- Savulescu, T. (Ed.) Flora Republicae Popularis Romanicae. (Flora of Romania) Vol. VII; Editio Academiae Rei Publicae Socialisticae Romania: Bucharest, Romania, 1960; pp. 225–226.
- Dihoru, G.; Negran, G. Cartea Roşie a Plantelor Vasculare din România (Red Book of Vascular Plants of Romania); Editura Academiei: Buchureşti, Romania, 2009; pp. 526–527. [Google Scholar]
- Jalas, J.; Suominen, J. (Eds.) Atlas Florae Europaeae. 1. Pteridophyta; Commitee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1972; p. 79. [Google Scholar]
- Jalas, J.; Suominen, J. (Eds.) Atlas Florae Europaeae. 3. Salicaceae to Balanophoraceae; Commitee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1976; p. 63. [Google Scholar]
- Jalas, J.; Suominen, J. (Eds.) Atlas Florae Europaeae. 7. Caryophyllaceae (Silenoideae); Commitee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1986; p. 65. [Google Scholar]
- Jalas, J.; Suominen, J. (Eds.) Atlas Florae Europaeae. 8. Nymphaeaceae to Ranunculaceae; Commitee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1989; pp. 135–136, 148. [Google Scholar]
- Kurtto, A.; Lampinen, R.; Junikka, L. Atlas Florae Europaeae. 13. Rosaceae (Spiraea to Fragaria, Excl. Rubus); Cambridge United Kingdom: Helsinki, Finland, 2004; p. 219. [Google Scholar]
- Bogosavljević, S.; Zlatković, B.; Ranđelović, V. Flora klisure Svrljiškog Timoka Stefan Bogosavljević. In Proceedings of the 9th Symposium on Flora of Southeastern Serbia and Neighbouring Regions, Niš, Serbia, 1–3 September 2007; pp. 41–54. [Google Scholar]
- Marković, S.M.; Stankov, J.V.; Mitić, V. Study of oak forests and scrubs of hornbeam vegetation, metals content of Teucrium chamedrys and soils the first year after wildfire on Vidlic Mountain. Saf. Eng. 2015, 5, 61–68. [Google Scholar]
- Petrova, A.; Vladimirov, V. Balkan endemics in the Bulgarian flora. Phytol. Balc. 2010, 16, 293–311. [Google Scholar]
- Jávorka, S. Magyar Flóra (Flora Hungarica); Magyar Nemzeti Múzeum Növénytára: Budapest, Hungary, 1924; pp. 691, 796. [Google Scholar]
- Stupar, V.; Milanović, D.; Brujić, J.; Čarni, A. Formalized classification and nomenclatural revision of thermophilous deciduous forests (Quercetalia pubescentis) of Bosnia and Herzegovina. Tuexenia 2015, 35, 85–130. [Google Scholar]
- Prodanović, D.; Krivošej, Z.; Lazarević, P.; Amidžić, L. Contribution to the knowledge of serpentine flora in Kosovo’s part of the Ibar river valley. Bot. Serb. 2010, 34, 81–86. [Google Scholar]
- Mártonfi, P.; Kolarcik, V.; Somlyay, L. Lectotypification of Onosma viridis and synonymization of O. tornensis with O. viridis (Boraginaceae). Ann. Bot. Fenn. 2014, 51, 201–206. [Google Scholar]
- Tatár, M. A Pannoniai Flóra Endemikus Fajai. Bölcsészdoktori Értekezés. (Endemic Species of the Pannonian Flora. Dissertation); Debreceni Református Kollégium Tanárképző Intézetének kiadása: Debrecen, Hungary, 1938. [Google Scholar]
- Oeyen, J.P.; Funke, S.; Böhme, W.; Wesener, T. The evolutionary history of the rediscovered Austrian population of the giant centipede Scolopendra cingulata Latreille 1829 (Chilopoda, Scolopendromorpha). PLoS ONE 2014, 9, e108650. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Disjunction between an isolated poleward population and the indicated former climatic optimum, in the case of a gap of North-South distance (N-SD) = 165 km.
Figure 1.
Disjunction between an isolated poleward population and the indicated former climatic optimum, in the case of a gap of North-South distance (N-SD) = 165 km.

Figure 2.
Multiple track structure of the supposed migration during the HTM-period; Explanation for subfigures (A–D) is provided in the main text. Symbols: square = closest recent southern occurrence to the recent isolation; circle = recent northernmost, isolated occurrence; solid line = northern edge of suitable habitats consisting of sporadic or continuous recent occurrences; broken line = hypothetic, supposed minimum of the northernmost expansion of the taxa during the HTM period (this could expand further north but without remnant occurrences poleward this cannot be judged); thin arrows with arrowheads on both ends = (N-SD), distance between the recent isolated and continuous occurrences, or distance between the continuous occurrences and the northernmost part of the suitable habitat-belt (D); thick, simple arrows = hypothetical tracks of the taxa during the HTM period; double solid lines = barrier of migration (A); double solid-broken line = added line (C).
Figure 2.
Multiple track structure of the supposed migration during the HTM-period; Explanation for subfigures (A–D) is provided in the main text. Symbols: square = closest recent southern occurrence to the recent isolation; circle = recent northernmost, isolated occurrence; solid line = northern edge of suitable habitats consisting of sporadic or continuous recent occurrences; broken line = hypothetic, supposed minimum of the northernmost expansion of the taxa during the HTM period (this could expand further north but without remnant occurrences poleward this cannot be judged); thin arrows with arrowheads on both ends = (N-SD), distance between the recent isolated and continuous occurrences, or distance between the continuous occurrences and the northernmost part of the suitable habitat-belt (D); thick, simple arrows = hypothetical tracks of the taxa during the HTM period; double solid lines = barrier of migration (A); double solid-broken line = added line (C).
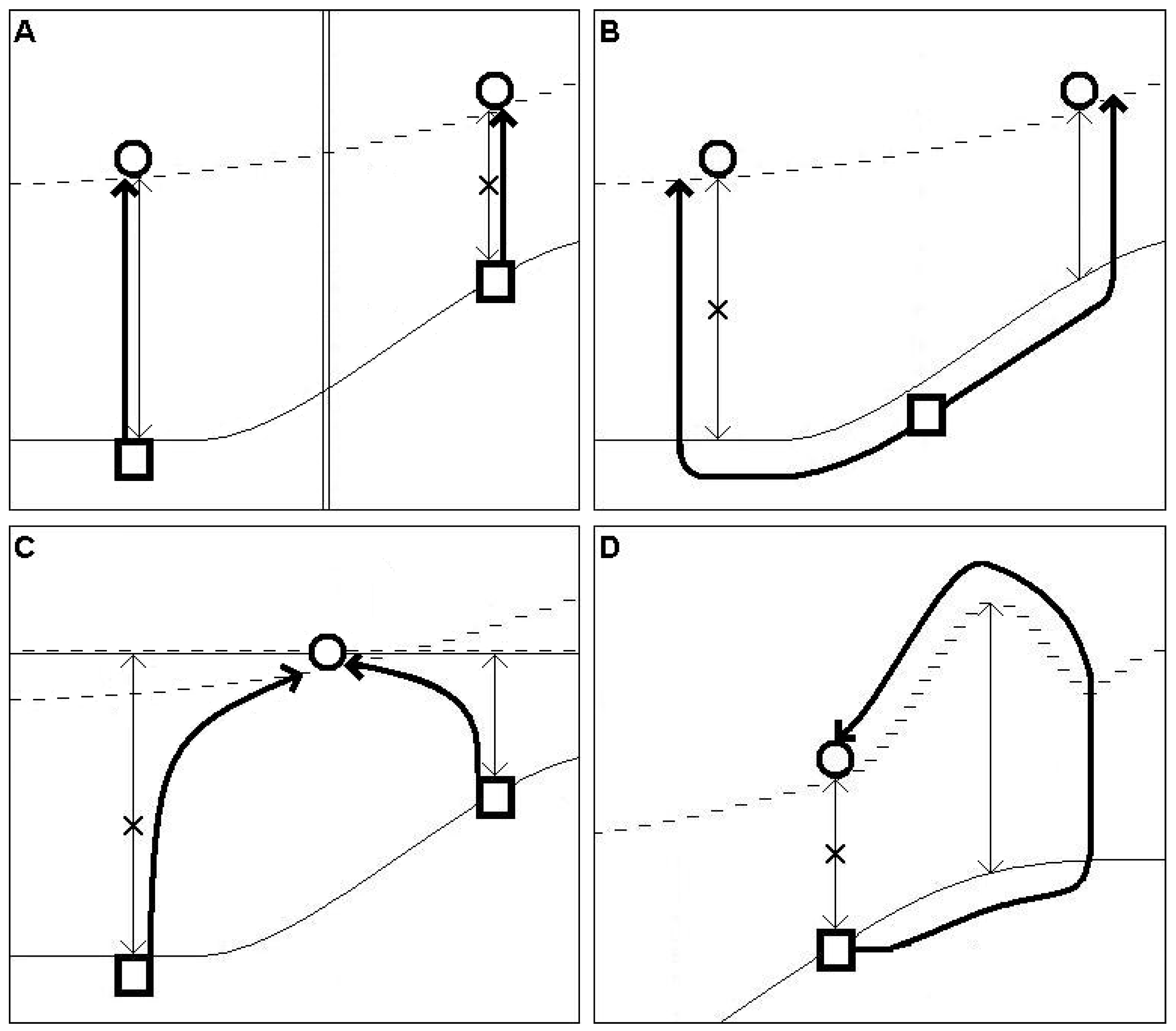
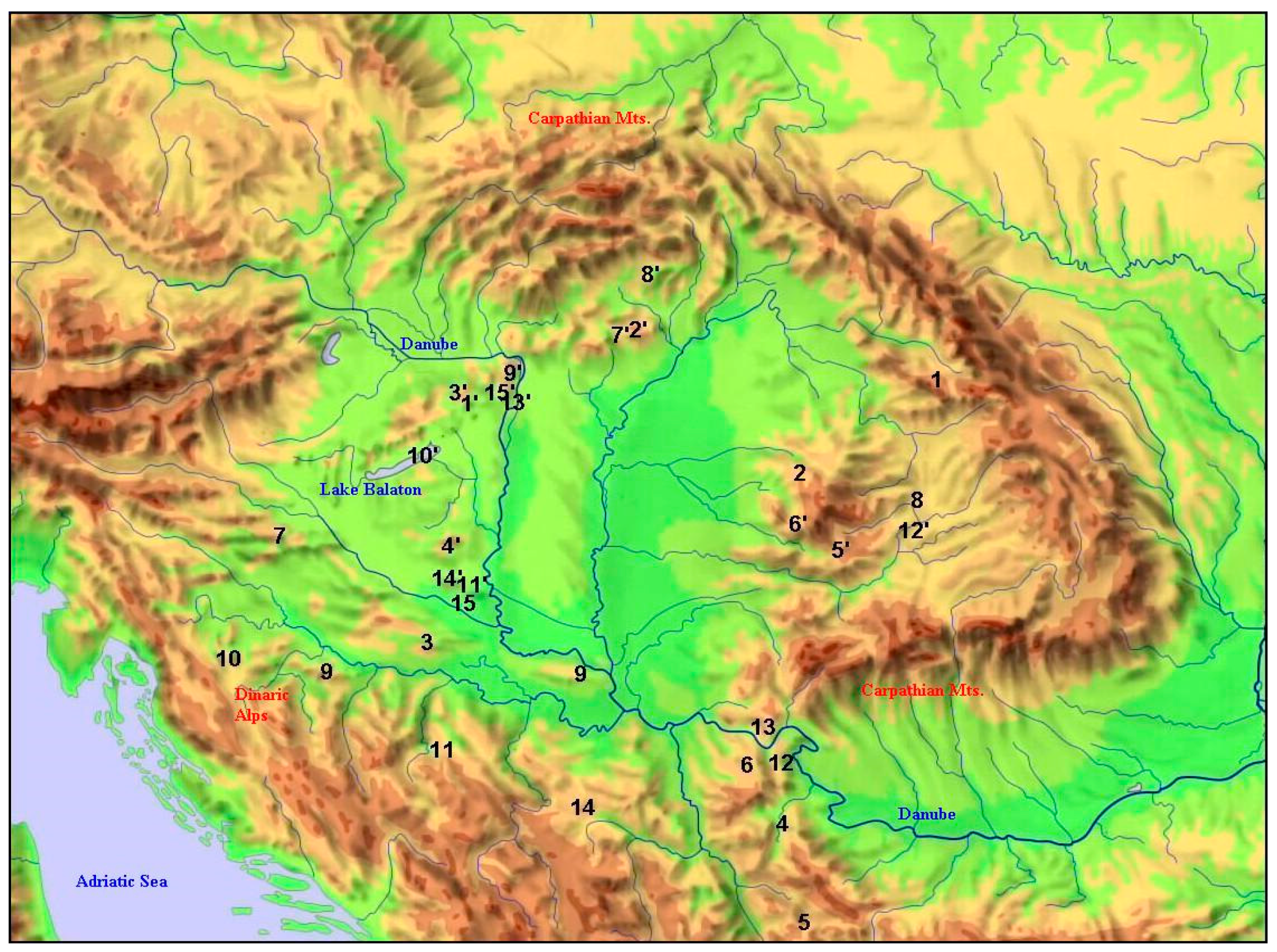
Figure 3.
Distribution of the northernmost and nearest southern occurrence-pairs of the selected 15 species (simplified to the supposed version in the case of more than one track). Numbers refer to species in Table 1.
Figure 3.
Distribution of the northernmost and nearest southern occurrence-pairs of the selected 15 species (simplified to the supposed version in the case of more than one track). Numbers refer to species in Table 1.
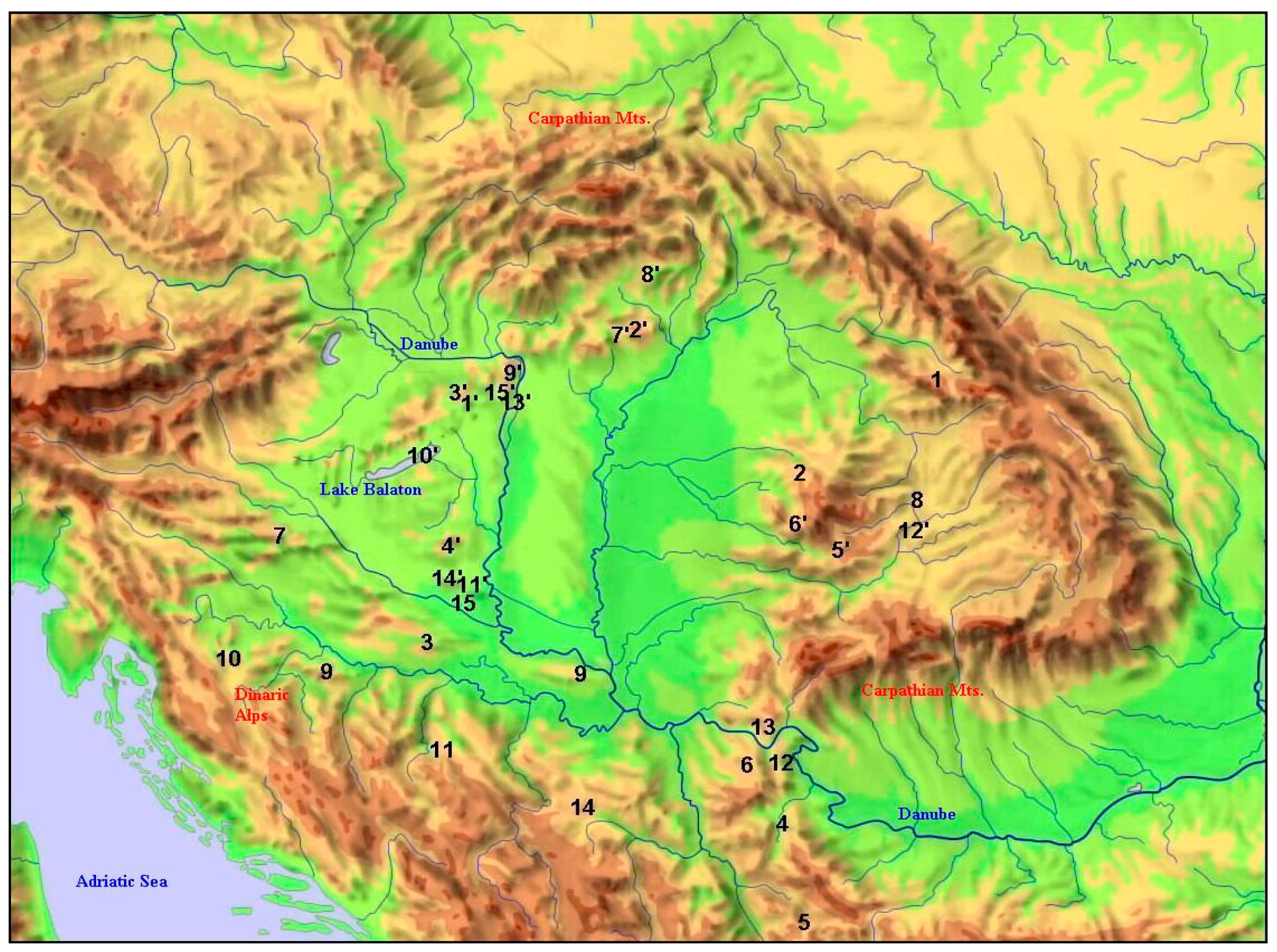
Figure 4.
Distribution of indicated MATD. Frequency = number of species in a certain 0.1 °C interval.
Figure 4.
Distribution of indicated MATD. Frequency = number of species in a certain 0.1 °C interval.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Final list of the selected 15 species.
| Species | Southern, Continuous Occurrence 1 (Degree of Latitude) | Northern, Isolated Occurrence 2 (Degree of Latitude) | N-SD (Distance in Degree of Latitude) | Difference in Temperature as Derived from N-SD (°C) |
|---|---|---|---|---|
| 1. Aethionema saxatile | 47.5 | 49.0 | 1.5 | 1.0 |
| 2. Asplenium lepidum | 47.0 | 49.0 | 2.0 | 1.4 |
| 3. Carpinus orientalis | 45.6 | 47.4 | 1.8 | 1.2 |
| 4. Cuscuta approximata | 44.1 | 46.1 | 2.0 | 1.4 |
| 5. Hypericum umbellatum | 43.1 | 46.3 | 3.2 | 2.2 |
| 6. Ilex aquifolium | 44.5 | 46.3 | 1.8 | 1.3 |
| 7. Micromeria thymifolia | 46.1 | 48.1 | 2.0 | 1.4 |
| 8. Onosma viridis | 46.6 | 49.0 | 2.4 | 1.7 |
| 9. Physospermum cornubiense | 45.2 | 47.8 | 2.6 | 1.8 |
| 10. Potentilla pedata | 45.2 | 46.9 | 1.7 | 1.2 |
| 11. Ranunculus psilostachys | 44.1 | 45.8 | 1.7 | 1.2 |
| 12. Saponaria bellidifolia | 44.5 | 46.5 | 2.0 | 1.4 |
| 13. Silene flavescens | 44.8 | 47.5 | 2.7 | 1.9 |
| 14. Trigonella gladiata | 44.1 | 45.8 | 1.7 | 1.2 |
| 15. Vincetoxicum pannonicum | 45.8 | 47.5 | 1.7 | 1.2 |
1 Northernmost edge of the large southern distribution, on the southern side of the gap of the distribution; 2 Isolated occurrence, on the northern side of the gap of the distribution or the northernmost point of the suitable habitat-belt required to be traversed.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Molnár, A.; Végvári, Z.; Tóthmérész, B. Identification of Floral Relicts Based on Spatial Distance of Isolation. Forests 2017, 8, 459. https://doi.org/10.3390/f8110459
AMA Style
Molnár A, Végvári Z, Tóthmérész B. Identification of Floral Relicts Based on Spatial Distance of Isolation. Forests. 2017; 8(11):459. https://doi.org/10.3390/f8110459
Chicago/Turabian StyleMolnár, Attila, Zsolt Végvári, and Béla Tóthmérész. 2017. "Identification of Floral Relicts Based on Spatial Distance of Isolation" Forests 8, no. 11: 459. https://doi.org/10.3390/f8110459
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.