
Decomposition of Leaves and Fine Roots in Three Subtropical Plantations in China Affected by Litter Substrate Quality and Soil Microbial Community

1
Key Laboratory on Forest Ecology and Environmental Sciences of State Forestry Administration, Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China
2
Research Institute of Economic Forestry, Xinjiang Academy of Forestry Science, Urumqi 830063, China
3
Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China
4
Tree and Timber Institute, National Research Council of Italy, Via Madonna del Piano 10, 50019 Sesto Fiorentino (FI), Italy
5
College of Forestry and Horticulture, Xinjiang Agricultural University, Urumqi 830052, China
*
Author to whom correspondence should be addressed.
Forests 2017, 8(11), 412; https://doi.org/10.3390/f8110412
Submission received: 9 August 2017
/
Revised: 19 October 2017
/
Accepted: 23 October 2017
/
Published: 30 October 2017
(This article belongs to the Special Issue Forest Fine Roots in Changing Climate)
Abstract
:Leaf and root litter decomposition has been a major research focus. However, the possible effects of belowground microbial community structure and diversity on this process are poorly understood. Understanding the biochemical mechanisms controlling aboveground decomposition processes is important to predict the changes of soil carbon and nutrient cycling in response to changes of forest management regimes. Here, we explore the biochemical controls of leaf and fine root decomposition in three subtropical plantations (Ford Erythrophleum (Erythrophleum fordii Oliver), Masson Pine (Pinus massoniana Lamb.)), and a mixed plantation containing both species) using the litterbag method, and soil microbial communities were determined using phospholipid fatty acid profiles. Overall, leaves decomposed more rapidly than fine roots, potentially due to the faster degradation of their cellulose component, but not lignin. In addition, leaf and fine root decomposition rates varied among plantations, being higher in E. fordii and lower in P. massoniana. Substrate quality such as N, Ca, lignin concentration, and C/N ratio were responsible for the decomposition rate changes among plantation types. Moreover, we used redundancy analysis to examine the relationships between litter decomposition and soil microbial community composition and diversity. Results revealed that actinobacteria and arbuscular mycorrhizal fungi community were the key determinants affecting leaf and fine root litter decomposition, respectively. Our work demonstrates that litter decomposition was linked to substrate quality and to the structure of soil microbial communities, and evidences the probable role of E. fordii in increasing soil nutrient availability, especially N, P and Ca. Additional data on phospholipid fatty acid (PLFA) or DNA marker groups within the litterbags over time may provide insights into litter decomposition dynamics, which represents potential objectives for future long-term decomposition studies.
1. Introduction
Litter decomposition is a crucial process influencing carbon (C) turnover and nutrient cycling in soils [1]. In forest ecosystems, plant litter is mainly non-woody is composition, derived from aboveground productivity such as leaves and twigs, and belowground productivity such as fine roots, and roots productivity can be similar in magnitude to leaf productivity [2]. During the past decades, research on leaf litter decomposition has been carried out extensively in different forest ecosystems [3,4,5,6]. However, root decomposition is an often ignored, yet potentially an important process for soil organic matter formation and nutrient cycling [7,8]. Understanding the mass loss dynamics and accompanying nutrient release patterns during root decomposition, and the factors that control these processes, are rather more limited when compared with leaf decomposition. In the few studies that compared decomposition processes of leaves and fine roots simultaneously, there appears to be no consistent trend in mass loss dynamics and nutrient release patterns [9,10]. Leaves have been reported to decompose both slower [10,11] and faster than fine roots [12,13,14]. To gain a comprehensive understanding of nutrient cycling in forest ecosystems, more studies comparing decomposition of leaves and fine roots are needed.
Substrate quality has been considered as one of the major factors regulating litter decomposition processes [1,15]. Particularly, high lignin concentrations usually have a strong rate-reducing influence on litter decomposition [10,16]. Previous studies, that compared initial chemical properties of leaves and roots, some reported lower concentrations of lignin in leaves than in roots [10,13,17]; usually considering this litter trait as a main cause for faster decomposition of leaves [12,13,17]. When the decomposition of different tree species (e.g., broadleaves vs. conifers, N-fixing vs. non-N-fixing) was compared, other initial litter substrate quality, such as nitrogen (N), phosphorus (P) and calcium (Ca) concentrations, C/N and lignin/N ratios, have also been identified as regulators of mass loss and nutrient cycling rates [5,18,19]. Generally, plant litter with lower C/N or lignin/N ratios decomposes at a relatively higher rate [11,14,20].
Besides substrate quality, litter decomposition is also strongly influenced by the soil microbial communities [20,21]. Decomposition is mediated by microbes that use plant primary production from above- and belowground litter, and soil, as C sources [22]. Therefore, alterations in the composition of surface litter resources, which modify the characteristics of the decomposer communities, could result in significant changes on the chemistry of decomposing litter [23]. How the soil decomposer communities respond to changes in litter species composition can have significant implications on the soil organic matter formation and nutrients dynamics, because soil decomposer communities are responsible for releasing up to 30% of the nutrients available to plants [4,24]. However, past research has demonstrated that litter substrate quality strongly controls rates of litter decomposition, how these factors indirectly influence the soil microbial communities, and hence, litter decomposition, is still poorly understood.
In subtropical China, commercial forests are extensively managed through afforestation and reforestation. However, the large-scale selection and planting of single coniferous (e.g., Pinus massoniana Lamb. and Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.)) or exotic (e.g., Eucalyptus (Eucalyptus spp.)) tree species, has caused a number of ecological problems, such as reduction in soil fertility, biodiversity and ecosystem stability [14]. Many plantations of indigenous valuable broadleaf trees are increasingly established to supply valuable timber, increase biodiversity and improve other ecosystem services [25]. Therefore, studies on litter decomposition of the main tree species used in forest management can provide a good opportunity to improve the understanding of C patterns and nutrient cycling in China’s subtropical plantations.
In this study, three representative subtropical plantation types, Erythrophleum fordii, Pinus massoniana, and a mixed plantation of the two, hereafter refer to as mixed forest, were chosen to carry out litter decomposition experiments. Erythrophleum fordii and P. massoniana are the dominant tree species used for afforestation and reforestation in the study area. We compared decomposition processes of leaves and fine roots in terms of mass loss rates, changes in organic chemical components (lignin and cellulose) and nutrients (C, N, P, potassium (K), sodium (Na), Ca, magnesium (Mg), manganese (Mn)) dynamics, including the soil microbial community structure. Our main objectives were (i) to compare litter decomposition rates and nutrient release patterns between the two substrates, and among tree species; and (ii) to identify the chemical and biological factors controlling litter decomposition processes in the tree species.
2. Materials and Methods
2.1. Study Description
The study site is located at the Daqingshan forest farm of Experimental Center of Tropical Forestry, Chinese Academy of Forestry (106°42′ E, 22°10′ N), Pingxiang City, Guangxi Zhuang Autonomous Region, China. The selected site is representative of regional afforestation and reforestation features in southern China. Altitude ranges from 120 to 210 m above sea level (a.s.l.) and the slope is approximately 10°. This region has a typical subtropical monsoon climate, with an annual average temperature of 21.0 °C and a relative humidity of 80% to 84%. Annual mean rainfall is approximately 1400 mm, concentrated between April and September, with an average evaporation ranging between 1261 and 1388 mm year−1. The soil is formed from weathered granite and is classified as lateritic red soil, according to the Chinese system of soil classification, with an organic layer about 8 cm thick (approximately, L: 2 cm; F: 2 cm; H: 4 cm). The average soil organic carbon (SOC), total nitrogen (TN), NH4+-N, NO3−-N, and pH of 0~10 cm soil is 17.84 g kg−1, 1.24 g kg−1, 6.91 mg kg−1, 1.95 mg kg−1, and 4.58, respectively. Litterfall and fine root biomass is about 360.17 g m−2 year−1 and 143.80 g m−2, respectively. For more details please refer to [25].
Vegetation within the Daqingshan forest farm is a mosaic of different plantation patches. Historically, this area was vegetated with a single coniferous species, P. massoniana; seedlings were planted in 1983 on a deforested hill. In 2006, the study plantations were designed with the same density of 2500 trees ha−1 after clear-cutting the former P. massoniana forest. Configuration of the mixed forest is 1:3 (i.e., 25% E. fordii and 75% P. massoniana), and the undergrowth vegetation is characterized by Japanese Climbing Fern (Lygodium japonicum (Thunb.) Sw.), Old World Forked Fern (Dicranopteris linearis (Burm.f.) Underw.), Spreading Cyrtococcum (Cyrtococcum patens (L.) A.Camus), and Common Lophantherum (Lophatherum gracile Brongn.).
2.2. Experimental Design and Mass Loss Estimation
For each plantation type, three patches were randomly selected, based on their similar topography, soil texture, stand age, management history, and litter horizon, for sampling and in situ litter decomposition estimations (i.e., nine patches in total). From July to August 2012, freshly abscised leaves of Erythrophleum fordii, Pinus massoniana and mixed forest were collected from litter fall-traps (1 m × 1 m nylon net, with a mesh size of 1 mm2) placed under the trees on the forest floor [14]. Simultaneously, fresh fine roots (<2 mm diameter) of E. fordii, P. massoniana and mixed forest were collected from a 0–10 cm soil layer. We opted to use fresh fine roots because they best represent roots that have not yet begun to decompose [26]. During collecting, the twigs, cones and other materials were removed from leaf samples, and adherent soil particles and other extraneous materials were carefully removed from fine root samples. All samples were then placed in polyethylene bags in duplicate and transported to the laboratory [27]. One was air-dried for initial chemical analyses, and the other was prepared for litter decomposition experiment. The decomposition process of leaves and fine roots was determined using the litterbag method [28]. Then, 10 g of air-dried leaves were placed in a 250 × 250 mm, 1 mm2-mesh polyethylene bag, and 2 g of air-dried fine roots were placed in a 100 × 100 mm, 1 mm2-mesh polyethylene bag [14]. For the mixed forest, the original litter mass of the two species in the litterbag was based on field composition, and the proportion of E. fordii and P. massoniana was 27.8% and 72.2%, respectively, for leaf litter, while it was 32% and 68% for fine roots.
To determine in situ litter decomposition, in each plantation patch, 42 leaf litterbags were placed back under seven randomly selected trees [29] and attached with metal pins to prevent movement [5]. Concomitantly, 42 fine root litterbags were buried into the soil at a depth of 5 cm [29], inserted at an angle of about 45°, and anchored to the soil with metal pins [5]. This was carried out on 26 August 2012. The litterbags of the two substrates were harvested at three-month intervals and total harvesting six points in time (i.e., 18-month decomposition period). In total, 756 litterbags were prepared, 378 for each. After harvesting the litter was transferred to the laboratory, adherent soil particles and other extraneous materials were carefully removed with a brush and tweezer, respectively. The residual leaves and fine roots were dried in an oven at 50 °C to constant weight to estimate remaining mass [29].
For several litter types and species, decomposition proceeds progressively more slowly as decay progresses, and the rate may even approach 0. Howard and Howard [30] found that the amount remaining after decomposition of some litter types approached a steady level. Berg [31] modified their function and created a new one.
where Lt is the accumulated mass loss (in percent), t is time in years or in days, and kA is the decomposition rate. The kA should be highest at t = 0 and decrease with increasing accumulated mass loss. Finally, m represents the asymptotic level that the accumulated mass loss will ultimately reach, normally not 100% and often considerably less.
2.3. Litter Chemical Analysis
To obtain sufficient material for chemical analyses, the seven litterbags of each substrate harvested from each patch were pooled as a residual sample after estimate remaining mass. Initial (Table 1) and residual samples of leaves and fine roots, taken at each sampling time, were dried at 70 °C and finely ground in a laboratory mill to perform the following chemical analyses. C and N concentrations were determined using a C/N autoanalyzer (vario MAX CN, Elementar Analysensysteme GmbH, Langenselbold, Germany). After microwave digestion, P, K, Na, Ca, Mg and Mn were measured by inductively coupled plasma (ICP) mass spectrometry analysis (IRIS Intrepid II XSP, Thermo Electron Corporation, Waltham, MA, USA); and lignin and cellulose concentrations were determined according to Van Soest and Wine [32].
2.4. Soil Microbial Community Analysis
To determine the structure of the soil microbial community, in each patch, soil samples (0–10 cm) were collected from seven cores (6 cm diameter) after 3, 6, 9, and 12 months. In the field, cores were passed through a 2 mm sieve and pooled together to get one composite sample for each patch. Fresh samples were immediately transported to the laboratory with a portable ice box and stored at −20 °C prior to analysis.
Soil microbial community structure was assessed using phospholipid fatty acid (PLFA) profiles [33,34]. Chemically similar lipid indicators were used to represent ecological groups of microorganisms. These included the following: Gram-positive (Gram+) bacteria (sum of 14:0 iso, 15:0 iso, 15:0 anteiso, 16:0 iso, 17:0 iso, and 17:0 anteiso), Gram-negative (Gram−) bacteria (sum of 16:1 ω11c, 16:1 ω7c, 17:1 ω8c, 16:1 2OH, and 18:1 ω7c), anaerobic bacteria (sum of 17:0 cyclo and 19:0 cyclo), actinobacteria (sum of 16:0 10-methyl, 17:0 10-methyl, and 19:0 10-methyl), arbuscular mycorrhizal (AM) fungi (16:1 ω5c), and saprotrophic and ectomycorrhizal (SEM) fungi (18:2ω6,9c and 18:1ω9c). Ratios of fungal:bacterial (F:B ratio), and Gram+:Gram− bacterial lipids (Gram+:Gram− ratio) were also included in the data analyses [35].
Microbial diversity indices were also calculated as soil microbial community attributes. According to Frostegård et al. [36], we treated each PLFA as a “species” and the size of the chromatogram peak as equivalent to the frequency of that species. Then, species richness (S) index was expressed as the number of species present in each plantation type, and Shannon’s diversity (H’) and Pielou’s evenness (J) indices were also calculated.
2.5. Statistical Analysis
Mixed effect model was used to test differences in dynamics of dry weight, organic chemical components and nutrient quantities of leaves and fine roots during decomposition. Sampling times were considered as a random factor while plantation types and litter types were used as fixed factors. Statistical tests were performed using SPSS 22.0 (IBM Crop., Armonk, NY, USA). Statistical significance was established at p values < 0.05. Figures were generated using SigmaPlot version 10.0. Redundancy analysis (RDA) was used to analyze the responses of litter decomposition (kA, C, N, P, K, Na, Ca, Mg, Mn, lignin and cellulose) to soil microbial community attributes (S, H’, J, Gram+ bacteria, Gram− bacteria, anaerobic bacteria, actinobacteria, AM fungi, SEM fungi, Gram+:Gram− ratio, F:B ratio) using CANOCO software (version 4.5, Microcomputer Power, Inc., Ithaca, NY, USA) for Windows. Automatic selection of means by Monte Carlo permutations was used to test the significance of the variables (p values < 0.05).
3. Results
3.1. Initial Chemical Properties
Initial nutrient concentration and organic chemical components significantly differed between leaves and fine roots and according to plantation types (Table 1). Concentrations of N, P, K, Na, Ca, Mg and Mn in the leaves were all significantly higher than in fine roots (p < 0.05, p < 0.01 or p < 0.001), whereas C was lower in the leaves compared to fine roots (p < 0.05). With regard to the organic chemical component, lignin concentration was higher in the fine roots than in leaves (p < 0.05). However, cellulose was similar in both litter substrates except for E. fordii leaves. The ratios of C/N, C/P, and lignin/N, were significantly higher in the fine roots when compared to the leaves (p < 0.01 or p < 0.001), however, the N/P ratio was higher in leaves than in fine roots (p < 0.001). For both leaves and fine roots, Erythrophleum fordii showed the highest concentrations of N, Ca, cellulose, C/P and N/P ratios, followed by mixed forest and Pinus massoniana. However, P, Mn, lignin, and lignin/N ratio in P. massoniana were greater than those in E. fordii and mixed forest.
3.2. Dry Mass Loss during Decomposition
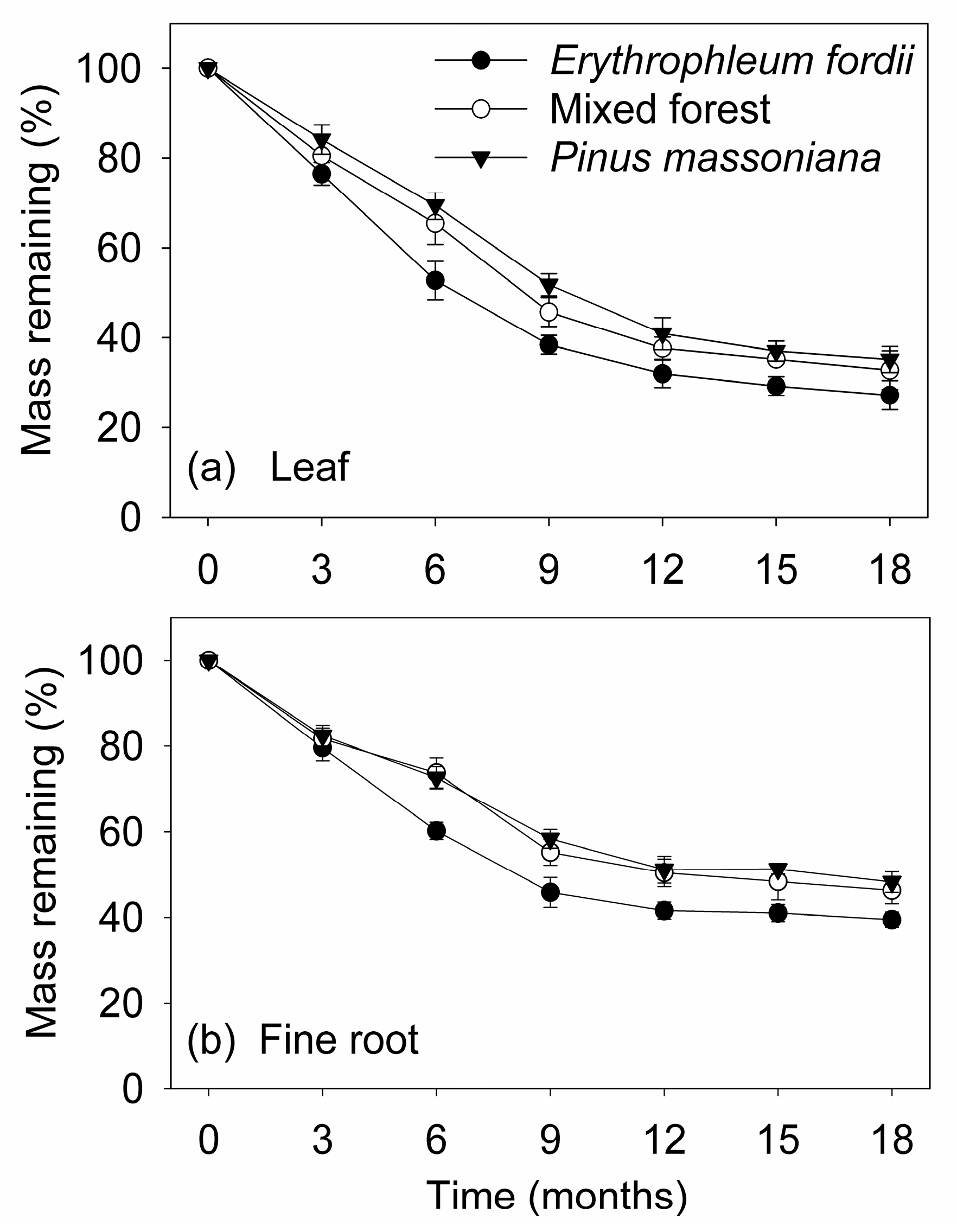
The mass loss of dry weight in the two substrates followed the same trend over the 18-month period studied. The greatest mass loss of the two substrates occurred in the initial nine months, decreasing gradually thereafter. Although, the initial chemical properties of the two substrates were highly variable among plantation types (Table 1), mass loss rates of leaves were significantly faster than those of fine roots during decomposition (p < 0.01; Figure 1 and Table 2, Table 3 and Table 4). The final mass remaining in the leaves at 18 months was 27–35% of the original compared with 40–48% in fine roots. Similarly, decomposition rate (kA) was higher in the leaves, ranging between 0.19 and 0.24 d−1, compared to fine roots, which ranged between 0.13 and 0.17 d−1 (Table 2).
Plantation type also showed a significant effect on the mass loss rates of leaves and fine roots during decomposition (p < 0.05; Figure 1 and Table 2, Table 3 and Table 4). The slowest decomposition was found for Pinus massoniana, while the fastest was observed for Erythrophleum fordii. After 18 months of decomposition, the lost mass of leaves and fine roots was 73% and 60% for E. fordii, 67% and 54% for mixed forest, and 65% and 52% for P. massoniana, respectively.
3.3. Organic Chemical Components during Decomposition
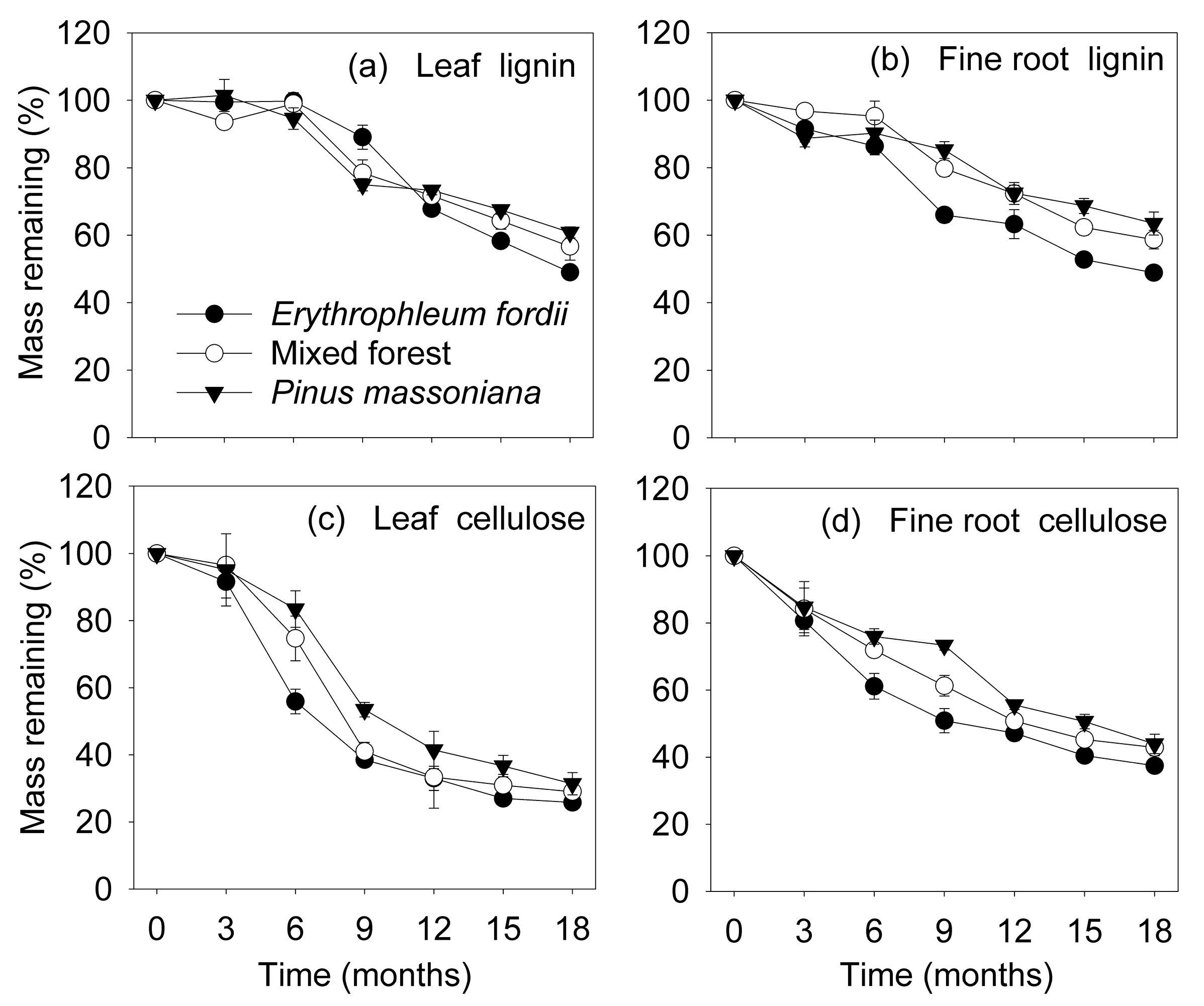
The remaining lignin mass did not change during the initial 6 months, but decreased thereafter (Figure 2a,b). As decomposition proceeded, the remaining mass of lignin varied significantly among plantation types (p < 0.001), but no effect of litter types (p = 0.144) was detected (Table 3 and Table 4). However, the remaining cellulose mass decreased during the initial 12 months, staying relatively constant thereafter (Figure 2c,d). The remaining mass of cellulose varied significantly among plantation types (p < 0.01), and between litter types (p < 0.001) during decomposition, being lower in leaves than fine roots (Table 3 and Table 4).
3.4. Nutrient Dynamics during Decomposition
Depending on the element, patterns of nutrient release differed between litter types and among plantations during decomposition (Figure 3, Figure 4 and Figure 5 and Table 3 and Table 4). During decomposition, plantation type significantly affected the remaining mass of C, N, P, Ca and Mn (p < 0.01 or (p < 0.001), but had no effect for K, Na and Mg. However, litter type significantly affected the mass remaining of C, N, Na, Ca, Mg and Mn (p < 0.01 or (p < 0.001)), but had no effect for P and K. Overall, nutrients were released faster in the leaves than in fine roots, and Erythrophleum fordii showed the fastest nutrient release, followed by mixed forest, and Pinus massoniana showed the slowest (Figure 4).
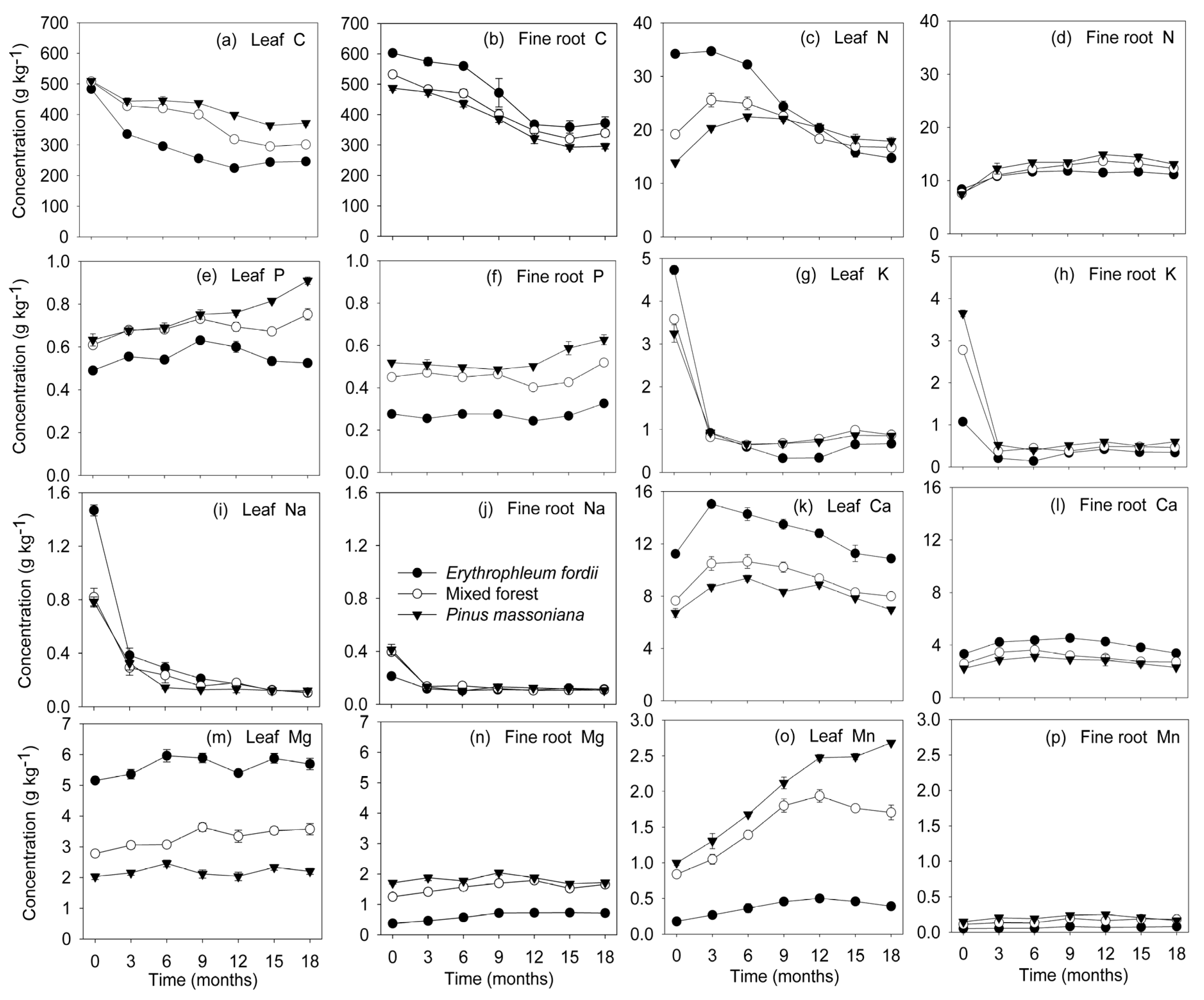
Initial C and Mg concentration declined gradually, staying relatively constant thereafter in all decomposing litter (Figure 3a,b,m,n). Change in remaining C mass (Figure 4a,b) was similar to that of dry weight (Figure 1). At the end of the investigation, about 14–26% of the initial C was retained in the leaves and 24–29% in fine roots.
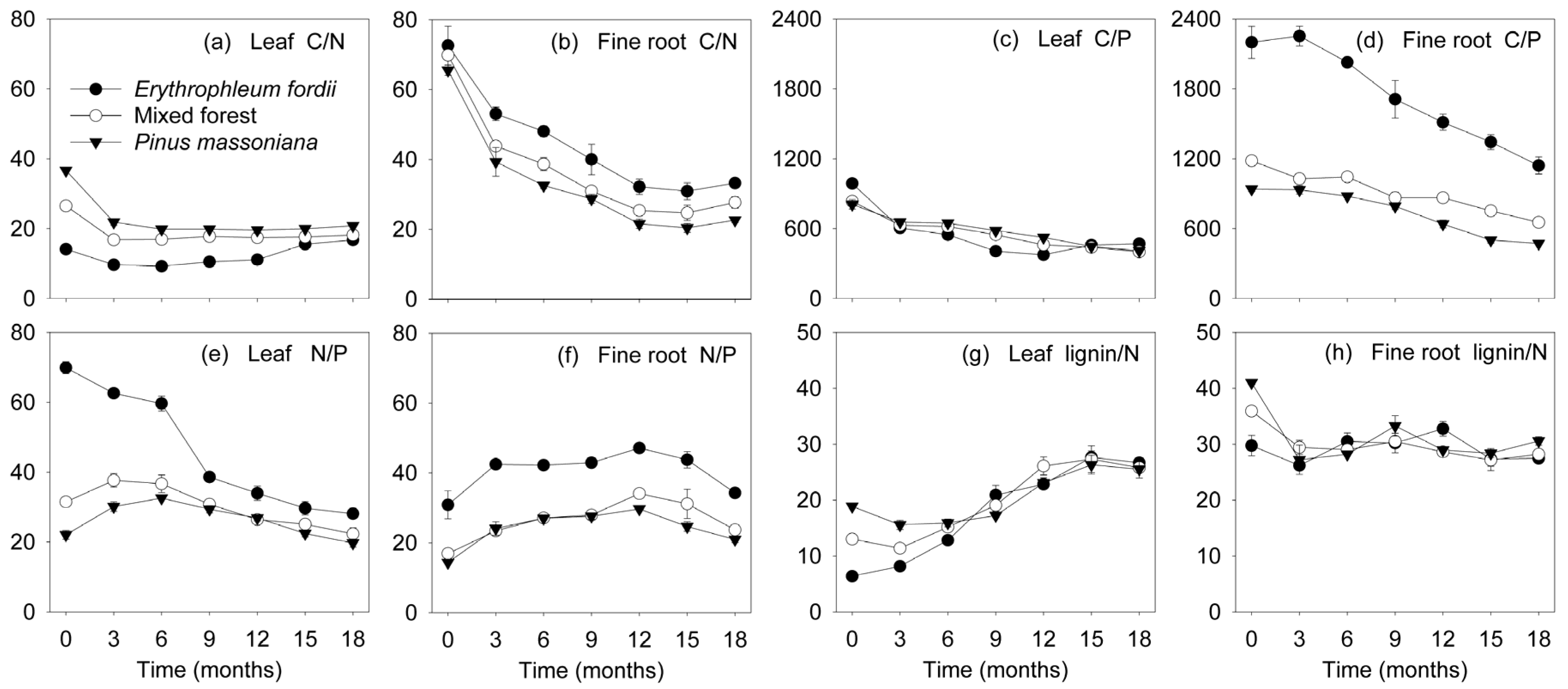
N and Ca concentration in the leaves exhibited a notable increase initially in most of the decomposing litter, followed by a decrease towards the end of the study (Figure 3c,k). An exception was the Erythrophleum fordii leaves, in which N concentration significantly decreased from 34.2 to 14.7 g kg−1. For fine roots, N and Ca concentration in all plantations increased with time (Figure 3d,l). However, remaining N and Ca masses eventually decreased, despite a temporal immobilization phase was shown (Figure 4c,d,k,l). The C/N ratio dropped initially and then remained constant (Figure 5a,b), whereas lignin/N was variable at first, eventually stabilizing (Figure 5g,h).
P concentration gradually increased during leaf and fine root decomposition (Figure 3e,f). Conversely, the C/P ratio declined gradually through time (Figure 5c,d). Change in the N/P ratio (Figure 5e,f) was similar to that of N concentration (Figure 3c,d). P was released rapidly at first, subsequently increasing in both leaves and fine roots. The exception was for the Erythrophleum fordii leaves, which tended to release P continuously (Figure 4e,f).
K and Na were the most rapidly lost elements from all decomposing litter. Finally, about 90–96% of the initial K and 94–98% of the initial Na were lost from leaves (Figure 4g,i), and 87–92% and 79–88% from fine roots (Figure 4h,j), respectively. This was matched by a sharp decrease in concentration (Figure 3g–j).
Mn concentration appeared to increase slightly through time, except for mixed forest and Pinus massoniana leaves, in which Mn concentrations increased significantly from 0.84 to 1.71 g kg−1 and from 1.00 to 2.68 g kg−1, respectively (Figure 3o,p). Mn eventually decreased in variable amounts during decomposition, with more than 50% retained in both leaves and fine roots (Figure 4o,p).
3.5. Soil Microbial Community
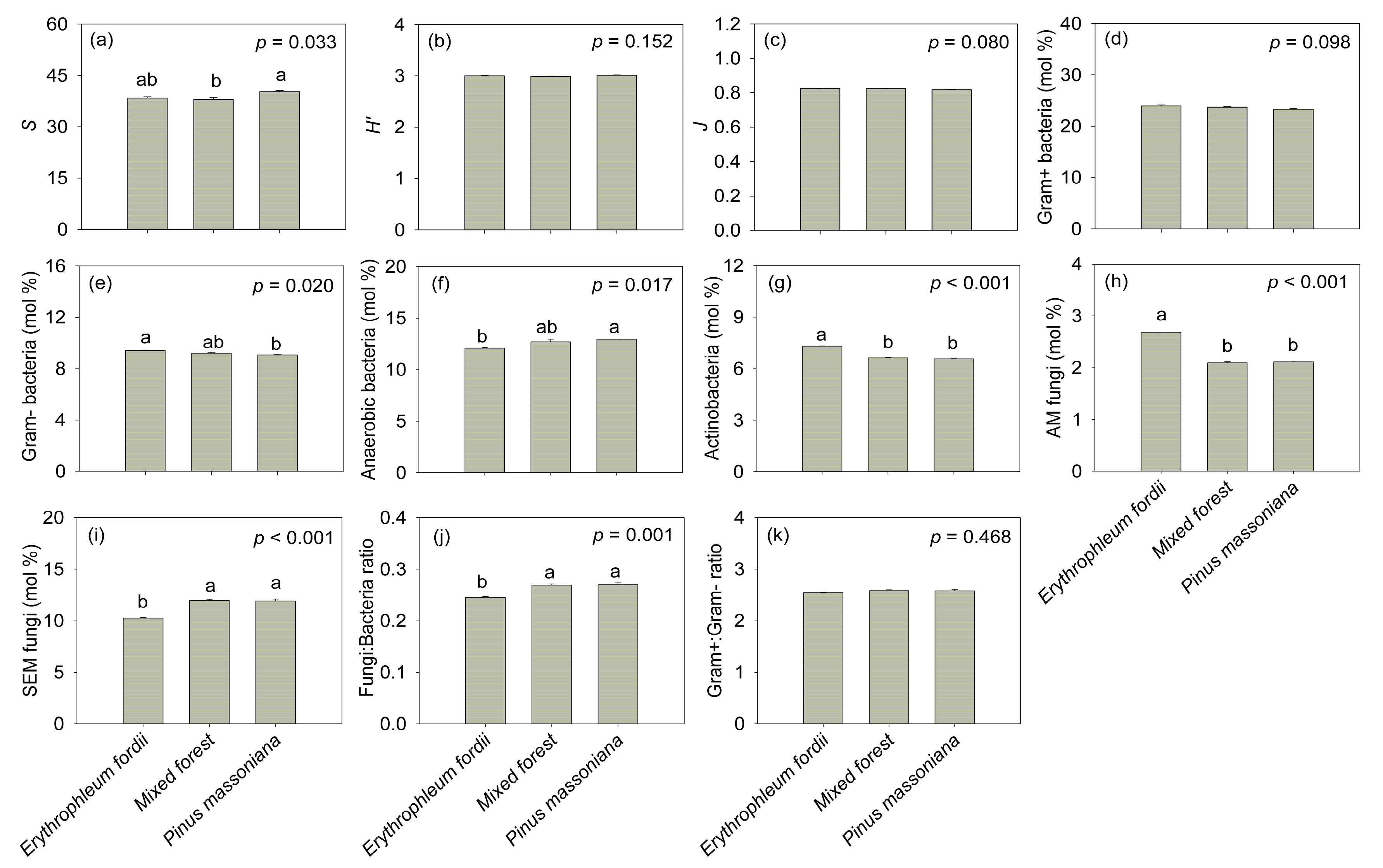
Plantation types significantly affected the S of PLFA signatures (F = 6.381, p < 0.05, Figure 6a) and the relative abundance of Gram− bacteria (F = 8.117, p < 0.05, Figure 6e), anaerobic bacteria (F = 8.645, p < 0.05, Figure 6f), actinobacteria (F = 198.224, p < 0.001, Figure 6g), AM fungi (F = 502.789, p < 0.001, Figure 6h), SEM fungi (F = 52.71, p < 0.001, Figure 6i), and the fungi:bacteria ratio (F = 29.487, p < 0.01, Figure 6j). Pinus massoniana soil was associated with the highest S and anaerobic bacteria abundances. The Gram− bacteria, actinobacteria, and AM fungi were significantly higher in Erythrophleum fordii soil than in mixed forest and P. massoniana soils. Conversely, SEM fungi and fungi:bacteria ratio in E. fordii soil was significantly lower than in the soils of other plantations. There was no significant difference among plantation types for the other microbial community attributes examined (p > 0.05; Figure 6b–d,k).
Redundancy analysis (RDA) showed that the first axes (RD 1) and second axes (RD 2) explained 77.3% and 9.0% of the total variance for the relationship between leaf litter decomposition and soil microbial community attributes (Figure 7a), while the RD 1 and RD 2 explained 66.1% and 14.1% of the total variance for the relationship between fine root litter decomposition and soil microbial community attributes (Figure 7b), respectively. The significance of soil microbial community attributes (S, H’, J, Gram+ bacteria, Gram− bacteria, anaerobic bacteria, actinobacteria, AM fungi, SEM fungi, fungi:bacteria ratio, and Gram+:Gram− ratio) present in the ordination was determined by Monte Carlo permutation tests, which demonstrated that actinobacteria (p = 0.006, F = 11.06) and AM fungi (p = 0.004, F = 9.23) were the key factors in influencing the leaf and fine root litter decomposition, respectively, under these forests assayed in this study.
4. Discussion
4.1. Decomposition of Leaves and Fine Roots
We found that fine roots decayed more slowly than leaves (Figure 1). This coincides with previous studies that have compared leaf and fine root decomposition processes simultaneously, which reported that a lower initial concentration of lignin in leaves results in a faster decay [12,17]. However, few studies also showed that leaves decomposed slower than fine roots, despite initial lignin concentration [10,11]. Lignin, which is in itself recalcitrant to degradation and can delay litter decomposition processes, was generally lower in leaves (Table 1). Nevertheless, we found there was no significant difference in remaining lignin mass between litter types during decomposition (Table 4), and the lost mass of the lignin component from leaves (39–51%) was similar to that of the fine roots (37–51%) at the end of the study (Figure 2a,b and Table 3). Therefore, observed differences in decomposition rates between leaves and fine roots cannot be directly ascribed to lignin concentrations. Regarding cellulose, that showed similar concentrations between the two litter types (Table 1) while the remaining mass significantly differed during decomposition (Table 4), and it decayed slower in fine roots than in leaves (Figure 2c,d). Therefore, differences in the decomposition of cellulose could be the main factor affecting decomposition of leaves and fine roots. A previous study [9] also reported that one of the most important regulators determining decay rates of the two litter substrates was the cellulose component, but not the acid-insoluble fraction (AIS), which is the less-decomposable fraction containing lignin.
However, apart from the litter substrate quality, the decomposition environment could also be a main factor controlling the decomposition processes [9,11]. The belowground environment, where the decomposing fine roots were buried, was considered more conducive to mass loss than leaves aboveground [11]. For example, previous studies proposed that moister conditions belowground could conduct rapid decay by promoting microbial activity and/or leaching, having a closer proximity to mineralized nutrients than aboveground, or being the decomposing material, more rapidly colonized by microbes [10,11]. Nevertheless, in our study, fine roots decomposed slower in better conditions as compared to leaves (Figure 1 and Table 3 and Table 4). It thus appears reasonable to suggest that the decomposition environment did not affect decomposition as strongly as litter quality [37].
4.2. Effects of Tree Species on Litter Decomposition
In our study, the lost mass of litter varied significantly among plantation types during the 18-month decomposition (Figure 4 and Figure 5 and Table 4), where the low-nutrient and high-lignin Pinus massoniana litter decomposed slowly, while the high-nutrient and low-lignin Erythrophleum fordii litter decomposed faster. This is consistent with previous findings that tree species with low litter quality have lower decomposition than those with higher litter quality [38], and further confirms that litter substrate quality is a major factor regulating decomposition processes [15]. Faster decomposition on E. fordii litter in our study could be related to its high-quality decomposition materials (i.e., high N and Ca, low C/N and lignin; Table 1). Previous studies have shown a significant and positive effect of litter initial N concentration on decomposition rates in most forest litter [39]. Litter N is an important factor known to stimulate microbial activity, and thus, affecting decomposition processes [40,41], and has been usually identified as a rate increaser during the initial period of decomposition [42,43]. As a good indicator of nutritional balance, litter C/N ratio has also been found to regulate soil and litter microbial communities, influencing litter nutrient dynamics [44]. In general, litter with low initial C/N ratio encourages rapid decomposition [20,45]. However, in our study, the decay rate of fine roots seemed less related to its C/N ratio (Table 1 and Figure 2). The observed decrease in decomposition rates of both leaves and fine roots with increasing lignin concentration in our study parallels the results obtained by Cizungu et al. [6]. Also, the concentration of litter lignin was negatively related to decomposition rate, as has been reported several times [12,13]. An additional factor explaining the relatively fast decomposition of Erythrophleum fordii litter may be its high Ca concentration (Table 1). Litter Ca has been identified as an essential co-factor of the ligninolytic enzymes of the decomposer microflora and it is favorable to the growth of white rot fungal species [5,46]. Moreover, litter Ca has also been related to increased activity of microorganisms, abundance and diversity of fungi and earthworms, and removal rates of the forest floor [47,48,49].
Litter mixtures usually decompose faster compared with the expected decomposition rates derived from the individual mono-specific litter types in the mixture [6,50]. This may result from niche differentiation due to an increased variety of substrates yielding increased microbial biomass and faster decomposition [4]. In this study, however, the mixed litter decomposed slower than monoculture litter of Erythrophleum fordii (Figure 1 and Table 2 and Table 4). Potentially, the relatively low contribution of the high quality E. fordii litter in mixed forest may have suppressed its additive effect on the litter mixture decomposition, indicating that a mixed litter is not necessarily better than a monoculture one. The lack of an additive effect in forest litter mixture was also observed in the initial decomposition in a central African tropical mountain forest [6].
4.3. Nutrient Dynamics during Decomposition
In the study, no significant difference in remaining mass of each nutrient element was found through time (Table 4), which indicates that they exhibited similar relative mobility. In general, distinct patterns of particular nutrient elements in the decomposition dynamics of various forest ecosystems reflect different nutrients availabilities for decomposer communities [5]. Therefore, those nutrient elements, with concentrations below the limiting threshold for decomposer communities, would be immobilized in the litter [1,51].
In our study, we found that N and Ca, early rate-controlling factors, were immobilized at the beginning of the study in most decomposing litter, followed by a rapid decrease towards the end of the study (Figure 4c,d,k,l), whilst P was immobilized in the litter at a late stage of decomposition (Figure 4e,f). Temporal variations of these nutrient elements may reflect changes in the factors controlling decomposition as decomposition progresses, litter quality changes and decomposers’ requirements vary [5]. The pattern of C release (Figure 4a,b) was similar to that of the dry weight loss (Figure 1) because C is a structural component of plant litter. The observed nutrient release patterns of N, P and Ca during decomposition are somewhat similar to those from other litter decomposition studies [5,11,52]. However, these elements have also showed particular dynamics in other studies. For example, N was continuously lost [53] or immobilized [11] during decomposition, and P was immobilized at the early stages of decomposition and subsequently released [53]. Conversely, release patterns of K and Na in all studied litter were distinctly different from those of other chemical elements (Figure 4g–j). K and Na showed the most rapid release during early decomposition. This pattern is characteristic of K and Na, because they are non-structural components of plant litter, and are subjected to physical removal through leaching without microbial activity [54,55]. Mg was also highly mobile, but declined more slowly than K and Na in all decomposing litter (Figure 4m,n). Mn is essential for the Mn peroxidase activity, a ligninolytic enzyme [56]. The remaining Mn mass eventually decreased at variable quantities (Figure 4o,p). This decay pattern is inconsistent with that in Mediterranean oak forests, where a Mn immobilization occurred during the late stages of decomposition [5].
4.4. Soil Microbial Influence on Decomposition
Recently, litter decomposition experiments have been used to directly explore the potential effects of microbial community composition and diversity on multiple ecosystem processes. Some studies have identified correlations between microbial community and litter decomposition processes as they are considered part of the same continuum. For example, Strickland et al. [57] assessed the effects of microbial community composition on the decomposition of rhododendron, pine, and grass litter sampled from three different temperate ecosystems. The authors concluded that microbial community composition explained a significant proportion of the variation in litter C mineralization rates (with community explaining 22.3–86.2% of rate variation). Esperschütz et al. [21] found that the composition and activity of the microbial community in the detritusphere were strongly linked to the amount of litter N. Krumins et al. [58] showed strong effects of eukaryote species richness on decomposition process. Consistently, in the present study, there were significant relationships from the RDA to confirm that litter decomposition was related to soil microbial community attributes (Figure 7). For example, arbuscular mycorrhizal fungi were the key determinants in influencing the fine root litter decomposition. Arbuscular mycorrhizal fungi, which form mycorrhizal symbioses with two out of three of all plant species, are believed to be obligate biotrophs that are wholly dependent on the plant partner for their carbon supply [59]. It is thought that they possess no degradative capability and that they are unable to decompose complex organic molecules. Some studies [60,61,62], however, suggested that arbuscular mycorrhizal fungi play an invaluable role in organic matter decomposition, as they can decay complex organic material through releasing extracellular enzymes and organic acids. For example, mycorrhizal fungi have been found to regulate the degradation of litter in tropical forest [60] or grass leaves in soil [61]. Also, mycorrhizal fungi has been reported to promote the decomposition of complex organic materials, such as C, N, P, in plant residues by secreting phosphatase, protease, cellulase, chitinase, and so forth [62]. Although our RDA revealed that actinobacteria was responsible for the leaf litter decomposition, the mechanisms needed to be studied more.
Nevertheless, other studies have also identified disconnections between microbial community and litter decomposition processes even though they are considered part of the same continuum. For example, Ball et al. [4] showed a correlation between N loss and the size and mass of microbial community in the litter but not in the soil, indicating the responses for the litter and soil are not the same. The lack of correlation between the litter decomposition processes and the soil microbial community suggests that the microbial community in the litter were different from the soil, perhaps because soil microbes are preferentially selected for different litter types [63], and/or because the microbial community is affected by soil fauna [21]. Additional data on PLFA or DNA marker groups within the litterbags over time may provide insights into litter decomposition dynamics. Such data are not available for our study, however represents potential objectives for future long-term decomposition studies.
5. Conclusions
In this study, we have compared differences of leaf and fine root litter decomposition and explored the possible effects of belowground microbial community composition and diversity on this process. We found that differences in the decomposition of cellulose, but not lignin, could be the main factor affecting decomposition of the two substrates. Faster decomposition on Erythrophleum fordii litter could be related to its high-quality decomposition materials (i.e., high N and Ca, low C/N and lignin). From redundancy analysis, we have identified that actinobacteria and arbuscular mycorrhizal fungi community are the main regulators affecting the leaf and fine root litter decomposition, respectively. Our work demonstrates that litter decomposition was linked to substrate quality and to the structure of soil microbial communities, and evidences the probable role of E. fordii in increasing soil nutrient availability, especially N, P and Ca. Although we have explored soil microbial communities as main factors controlling aboveground litter decomposition in these plantations, further studies, like PLFA or DNA marker group analyses within the litterbags over time, are needed to provide insights into litter decomposition processes.
Acknowledgments
We would like to thank the Guangxi Youyiguan Forest Ecosystem Research Station for their support. We are grateful to Shenglei Fu and Lixia Zhou for their help with soil microbial community analyses. We also appreciate two anonymous reviewers for their valuable comments and suggestions on the manuscript. This work was jointly funded by the National Key Research and Development Program (2016YFC0502104-02), and the National Natural Science Foundation of China (31290223, 31570240).
Author Contributions
D.L., R.C., and Z.S. participated in the discussion and experimental designs; W.W. prepared the experimental materials; D.L. undertook laboratory analyses and drafted the manuscript. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare that they have no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Norby, R.J.; Ledford, J.; Reilly, C.D.; Miller, N.E.; O’Neill, E.G. Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichment. Proc. Natl. Acad. Sci. USA 2004, 101, 9689–9693. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Su, J.; Janssens, I.A.; Zhou, G.; Xiao, C. Fine root and litterfall dynamics of three Korean pine (Pinus koraiensis) forests along an altitudinal gradient. Plant Soil 2014, 374, 19–32. [Google Scholar] [CrossRef]
- Ball, B.A.; Carrillo, Y.; Molina, M. The influence of litter composition across the litter-soil interface on mass loss, nitrogen dynamics and the decomposer community. Soil Biol. Biochem. 2014, 69, 71–82. [Google Scholar] [CrossRef]
- Aponte, C.; García, L.V.; Maranon, T. Tree species effect on litter decomposition and nutrient release in mediterranean oak forests changes over time. Ecosystems 2012, 15, 1204–1218. [Google Scholar] [CrossRef] [Green Version]
- Cizungu, L.; Staelens, J.; Huygens, D.; Walangululu, J.; Muhindo, D.; Van Cleemput, O.; Boeckx, P. Litterfall and leaf litter decomposition in a central African tropical mountain forest and Eucalyptus plantation. For. Ecol. Manag. 2014, 326, 109–116. [Google Scholar] [CrossRef]
- Lai, Z.; Zhang, Y.; Liu, J.; Wu, B.; Qin, S.; Fa, K. Fine-root distribution, production, decomposition, and effect on soil organic carbon of three revegetation shrub species in northwest China. For. Ecol. Manag. 2016, 359, 381–388. [Google Scholar] [CrossRef]
- Ma, C.; Xiong, Y.; Li, L.; Guo, D. Root and leaf decomposition become decoupled over time: Implications for below- and above-ground relationships. Funct. Ecol. 2016, 30, 1239–1246. [Google Scholar] [CrossRef]
- Fujii, S.; Takeda, H. Dominant effects of litter substrate quality on the difference between leaf and root decomposition process above- and belowground. Soil Biol. Biochem. 2010, 42, 2224–2230. [Google Scholar] [CrossRef]
- Hobbie, S.E. Temperature and plant species control over litter decomposition in Alaskan tundra. Ecol. Monogr. 1996, 66, 503–522. [Google Scholar] [CrossRef]
- Ostertag, R.; Hobbie, S.E. Early stages of root and leaf decomposition in Hawaiian forests: Effects of nutrient availability. Oecologia 1999, 121, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, J.; Vogt, K.A.; Vogt, D.J. Decay rate and substrate quality of fine roots and foliage of two tropical tree species in the Luquillo Experimental Forest, Puerto Rico. Plant Soil 1993, 150, 233–245. [Google Scholar] [CrossRef]
- Lehmann, J.; Schroth, G.; Zech, W. Decomposition and nutrient release from leaves, twigs and roots of three alley-cropped tree legumes in central Togo. Agrofor. Syst. 1995, 29, 21–36. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Mo, J. Correlation between leaf litter and fine root decomposition among subtropical tree species. Plant Soil 2010, 335, 289–298. [Google Scholar] [CrossRef]
- Couteaux, M.M.; Bottner, P.; Berg, B. Litter decomposition, climate and liter quality. Trends Ecol. Evol. 1995, 10, 63–66. [Google Scholar] [CrossRef]
- Osono, T.; Takeda, H. Decomposition of organic chemical components in relation to nitrogen dynamics in leaf litter of 14 tree species in a cool temperate forest. Ecol. Res. 2005, 20, 41–49. [Google Scholar] [CrossRef]
- Fujimaki, R.; Takeda, H.; Wiwatiwitaya, D. Fine root decomposition in tropical dry evergreen and dry deciduous forests in Thailand. J. For. Res. 2008, 13, 338–346. [Google Scholar] [CrossRef]
- Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship. Oikos 1997, 79, 439–449. [Google Scholar] [CrossRef]
- Li, S.; Liu, W.; Li, D.; Li, Z.; Song, L.; Chen, K.; Fu, Y. Slower rates of litter decomposition of dominant epiphytes in the canopy than on the forest floor in a subtropical montane forest, southwest China. Soil Biol. Biochem. 2014, 70, 211–220. [Google Scholar] [CrossRef]
- Esperschütz, J.; Zimmermann, C.; Dümig, A.; Welzl, G.; Buegger, F.; Elmer, M.; Munch, J.C.; Schloter, M. Dynamics of microbial communities during decomposition of litter from pioneering plants in initial soil ecosystems. Biogeosciences 2013, 10, 5115–5124. [Google Scholar] [CrossRef]
- Smyth, C.E.; Macey, D.; Trofymow, J.A. Long-term litter decay in Canadian forests and the influence of soil microbial community and soil chemistry. Soil Biol. Biochem. 2015, 80, 251–259. [Google Scholar] [CrossRef]
- Brant, J.B.; Myrold, D.D.; Sulzman, E.W. Root controls on soil microbial community structure in forest soils. Oecologia 2006, 148, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Wickings, K.; Grandy, A.S.; Reed, S.C.; Cleveland, C.C. The origin of litter chemical complexity during decomposition. Ecol. Lett. 2012, 15, 1180–1188. [Google Scholar] [CrossRef] [PubMed]
- Setälä, H.; Huhta, V. Soil fauna increase Betula pendula growth: Laboratory experiments with coniferous forest floor. Ecology 1991, 72, 665–671. [Google Scholar] [CrossRef]
- Luo, D.; Cheng, R.; Shi, Z.; Wang, W.; Xu, G.; Liu, S. Impacts of nitrogen-fixing and non-nitrogen-fixing tree species on soil respiration and microbial community composition during forest management in subtropical China. Ecol. Res. 2016, 31, 683–693. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Oleksyn, J.; Eissenstat, D.M.; Reich, P.B. Fine root decomposition rates do not mirror those of leaf litter among temperate tree species. Oecologia 2010, 162, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Brown, S.; Xue, J.; Fang, Y.; Li, Z. Response of litter decomposition to simulated N deposition in disturbed, rehabilitated and mature forests in subtropical China. Plant Soil 2006, 282, 135–151. [Google Scholar] [CrossRef]
- Berg, B.; Berg, M.P.; Bottner, P.; Box, E.; Breymeyer, A.; de Anta, R.C.; Couteaux, M.; Escudero, A.; Gallardo, A.; Kratz, W. Litter mass loss rates in pine forests of Europe and Eastern United States: Some relationships with climate and litter quality. Biogeochemistry 1993, 20, 127–159. [Google Scholar] [CrossRef]
- Ostertag, R.; Marín-Spiotta, E.; Silver, W.L.; Schulten, J. Litterfall and decomposition in relation to soil carbon pools along a secondary forest chronosequence in Puerto Rico. Ecosystems 2008, 11, 701–714. [Google Scholar] [CrossRef]
- Howard, P.J.A.; Howard, D.M. Microbial decomposition of tree and shrub leaf litter. Oikos 1974, 25, 341–352. [Google Scholar] [CrossRef]
- Berg, B. Decomposition patterns for foliar litter—A theory for influencing factors. Soil Biol. Biochem. 2014, 78, 222–232. [Google Scholar] [CrossRef]
- Van Soest, P.; Wine, R. Determination of lignin and cellulose in acid-detergent fiber with permanganate. J. Assoc. Off. Anal. Chem. 1968, 51, 780–785. [Google Scholar]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Frostegård, Å.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fert. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Fichtner, A.; Von Oheimb, G.; Härdtle, W.; Wilken, C.; Gutknecht, J. Effects of anthropogenic disturbances on soil microbial communities in oak forests persist for more than 100 years. Soil Biol. Biochem. 2014, 70, 79–87. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol. Biochem. 2011, 43, 1621–1625. [Google Scholar] [CrossRef]
- McClaugherty, C.A.; Pastor, J.; Aber, J.D.; Melillo, J.M. Forest litter decomposition in relation to soil nitrogen dynamics and litter quality. Ecology 1985, 66, 266–275. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Krashevska, V.; Sandmann, D.; Maraun, M.; Scheu, S. Consequences of exclusion of precipitation on microorganisms and microbial consumers in montane tropical rainforests. Oecologia 2012, 170, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Hättenschwiler, S.; Tiunov, A.V.; Scheu, S. Biodiversity and litter decomposition in terrestrial ecosystems. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Aber, J.D.; Melillo, J.M.; McClaugherty, C.A. Predicting long-term patterns of mass loss, nitrogen dynamics, and soil organic matter formation from initial fine litter chemistry in temperate forest ecosystems. Can. J. Bot. 1990, 68, 2201–2208. [Google Scholar] [CrossRef]
- Berg, B. Litter decomposition and organic matter turnover in northern forest soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Eddy, W.C.; Buyarski, C.R.; Adair, E.C.; Ogdahl, M.L.; Weisenhorn, P. Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment. Ecol. Monogr. 2012, 82, 389–405. [Google Scholar] [CrossRef]
- Enriquez, S.; Duarte, C.M.; Sand-Jensen, K. Patterns in decomposition rates among photosynthetic organisms: The importance of detritus C: N: P content. Oecologia 1993, 94, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Hirata, E. Decomposition patterns of leaf litter of seven common canopy species in a subtropical forest: N and P dynamics. Plant Soil 2005, 273, 279–289. [Google Scholar] [CrossRef]
- Eriksson, K.E.L.; Blanchette, R.; Ander, P. Microbial and Enzymatic Degradation of Wood and Wood Components; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Berg, B.; De Santo, A.V.; Rutigliano, F.A.; Fierro, A.; Ekbohm, G. Limit values for plant litter decomposing in two contrasting soils-influence of litter elemental composition. Acta Oecol. 2003, 24, 295–302. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Pérez, C.A.; Hedin, L.O.; Armesto, J.J. Nitrogen mineralization in two unpolluted old-growth forests of contrasting biodiversity and dynamics. Ecosystems 1998, 1, 361–373. [Google Scholar]
- Staff, H.; Berg, B. Accumulation and release of plant nutrients in decomposing Scots pine needle litter. II. Long-term decomposition in a Scots pine forest. Can. J. Bot. 1982, 60, 1561–1568. [Google Scholar] [CrossRef]
- Osono, T.; Azuma, J.; Hirose, D. Plant species effect on the decomposition and chemical changes of leaf litter in grassland and pine and oak forest soils. Plant Soil 2014, 376, 411–421. [Google Scholar] [CrossRef]
- Lisanework, N.; Michelsen, A. Litterfall and nutrient release by decomposition in three plantations compared with a natural forest in the Ethiopian highland. For. Ecol. Manag. 1994, 65, 149–164. [Google Scholar] [CrossRef]
- O’Connell, A. Nutrient dynamics in decomposing litter in karri (Eucalyptus diversicolor F. Muell.) forests of south-western Australia. J. Ecol. 1988, 76, 1186–1203. [Google Scholar] [CrossRef]
- Sharma, E.; Ambasht, R.S. Litterfall, decomposition and nutrient release in an age sequence of Alnus nepalensis plantation stands in the eastern Himalaya. J. Ecol. 1987, 75, 997–1010. [Google Scholar] [CrossRef]
- Berg, B.; Steffen, K.T.; McClaugherty, C. Litter decomposition rate is dependent on litter Mn concentrations. Biogeochemistry 2007, 82, 29–39. [Google Scholar] [CrossRef]
- Strickland, M.; Lauber, C.; Fierer, N.; Bradford, M.A. Testing the functional signifcance of microbial community composition. Ecology 2009, 90, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Krumins, J.A.; Long, Z.T.; Steiner, C.F.; Morin, P.J. Indirect effects of food web diversity and productivity on bacterial community function and composition. Funct. Ecol. 2006, 20, 514–521. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Went, F.W.; Stark, N. Mycorrhiza. BioScience 1968, 18, 1035–1039. [Google Scholar] [CrossRef]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Pritsch, K.; Garbaye, J. Enzyme secretion by ECM fungi and exploitation of mineral nutrients from soil organic matter. Ann. For. Sci. 2011, 68, 25–32. [Google Scholar] [CrossRef]
- Norris, C.E.; Quideau, S.A.; Macey, D.E. Processing of 13C glucose in mineral soil from aspen, spruce and novel ecosystems in the Athabasca oil sands region. Appl. Soil Ecol. 2013, 71, 24–32. [Google Scholar] [CrossRef]
Figure 1.
Average percent of leaf and fine root litter original remaining mass (mean ± standard error, n = 3) of three subtropical plantations in China.
Figure 1.
Average percent of leaf and fine root litter original remaining mass (mean ± standard error, n = 3) of three subtropical plantations in China.
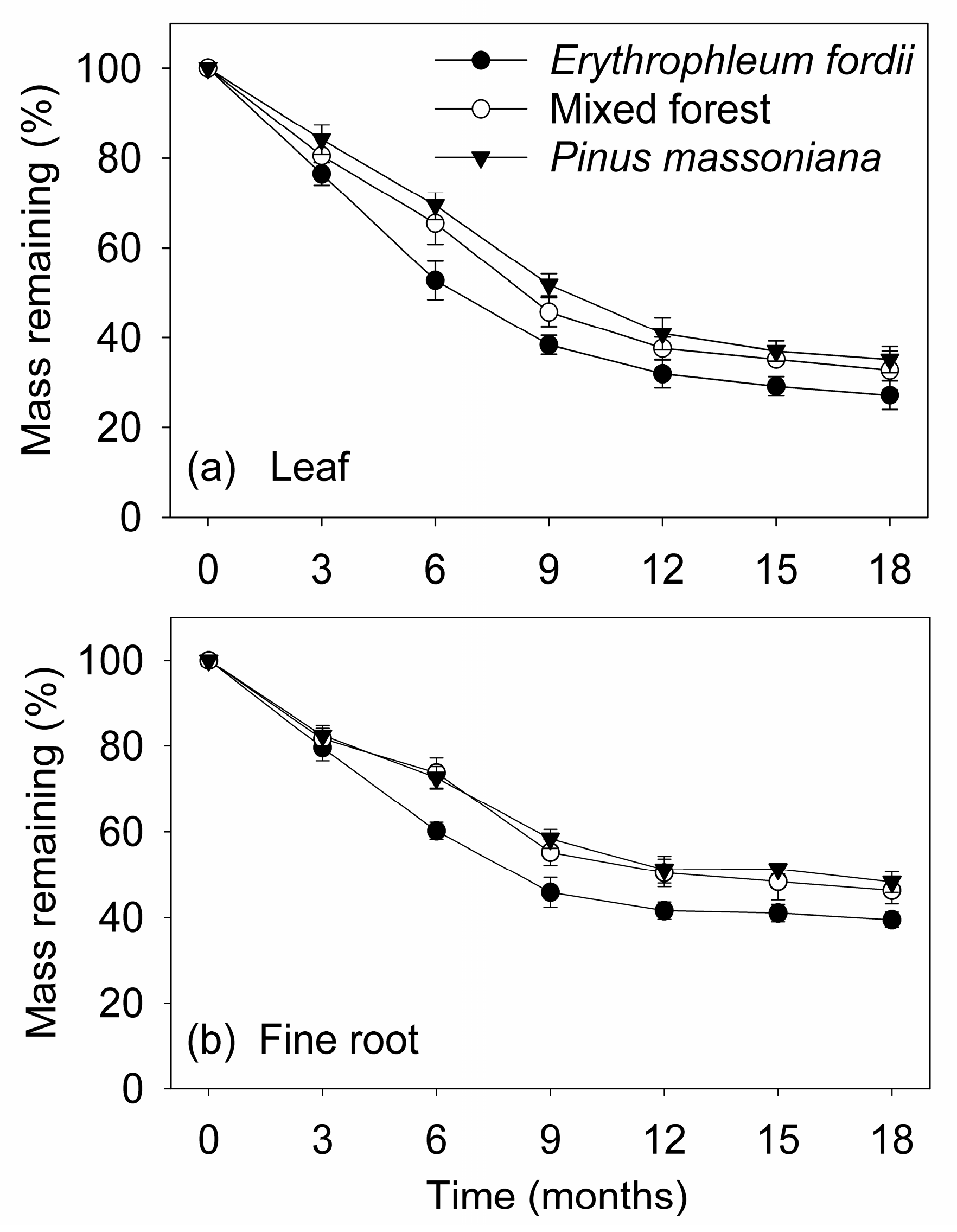
Figure 2.
Dynamics of lignin and cellulose remaining mass (mean ± standard error, n = 3) in leaf and fine root litter of three subtropical plantations in China.
Figure 2.
Dynamics of lignin and cellulose remaining mass (mean ± standard error, n = 3) in leaf and fine root litter of three subtropical plantations in China.
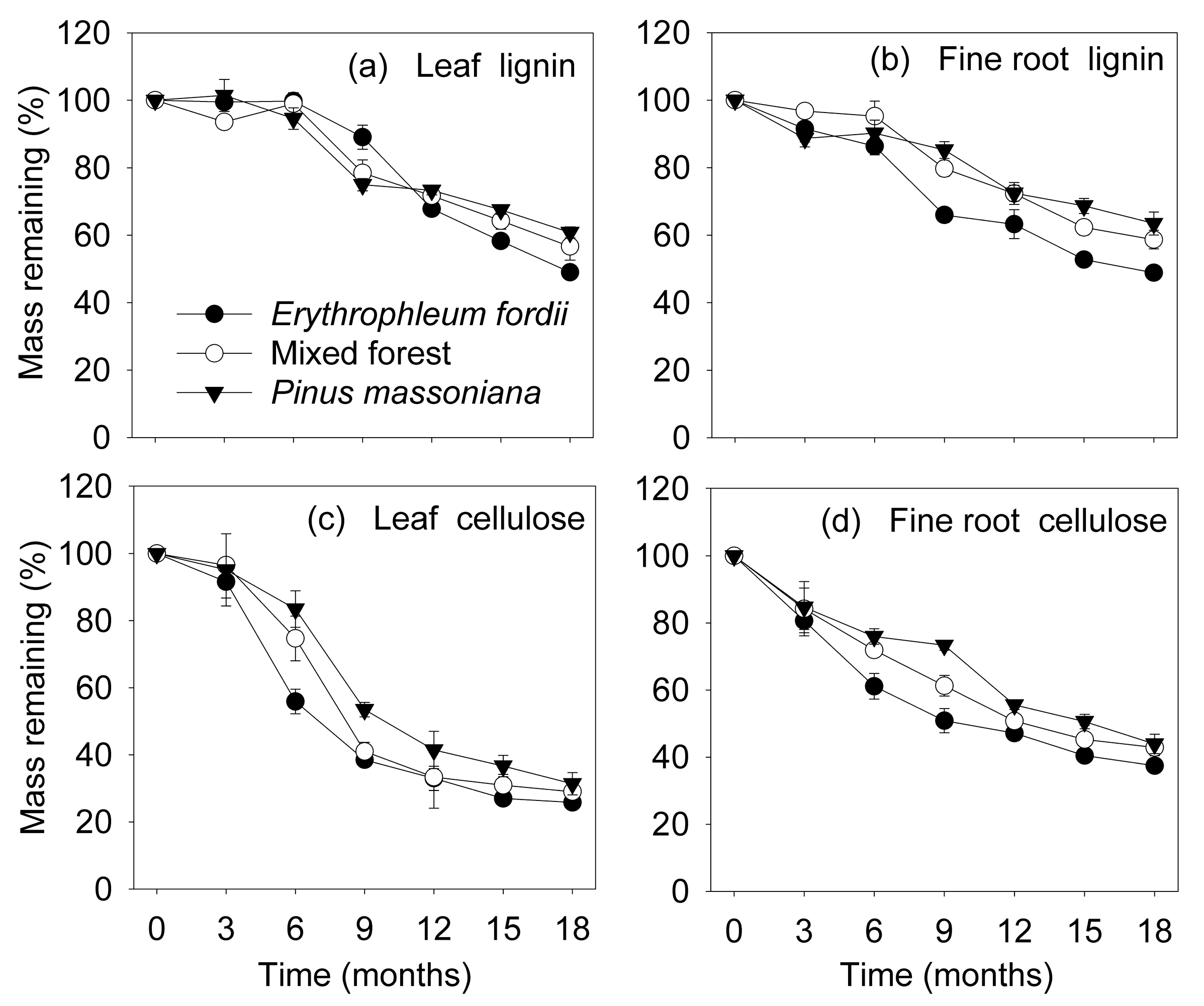
Figure 3.
Nutrient concentration dynamics (mean ± standard error, n = 3) in leaf and fine root litter of three subtropical plantations in China.
Figure 3.
Nutrient concentration dynamics (mean ± standard error, n = 3) in leaf and fine root litter of three subtropical plantations in China.

Figure 4.
Nutrient quantity dynamics (mean ± standard error, n = 3) in leaf and fine root litter of three subtropical plantations in China.
Figure 4.
Nutrient quantity dynamics (mean ± standard error, n = 3) in leaf and fine root litter of three subtropical plantations in China.
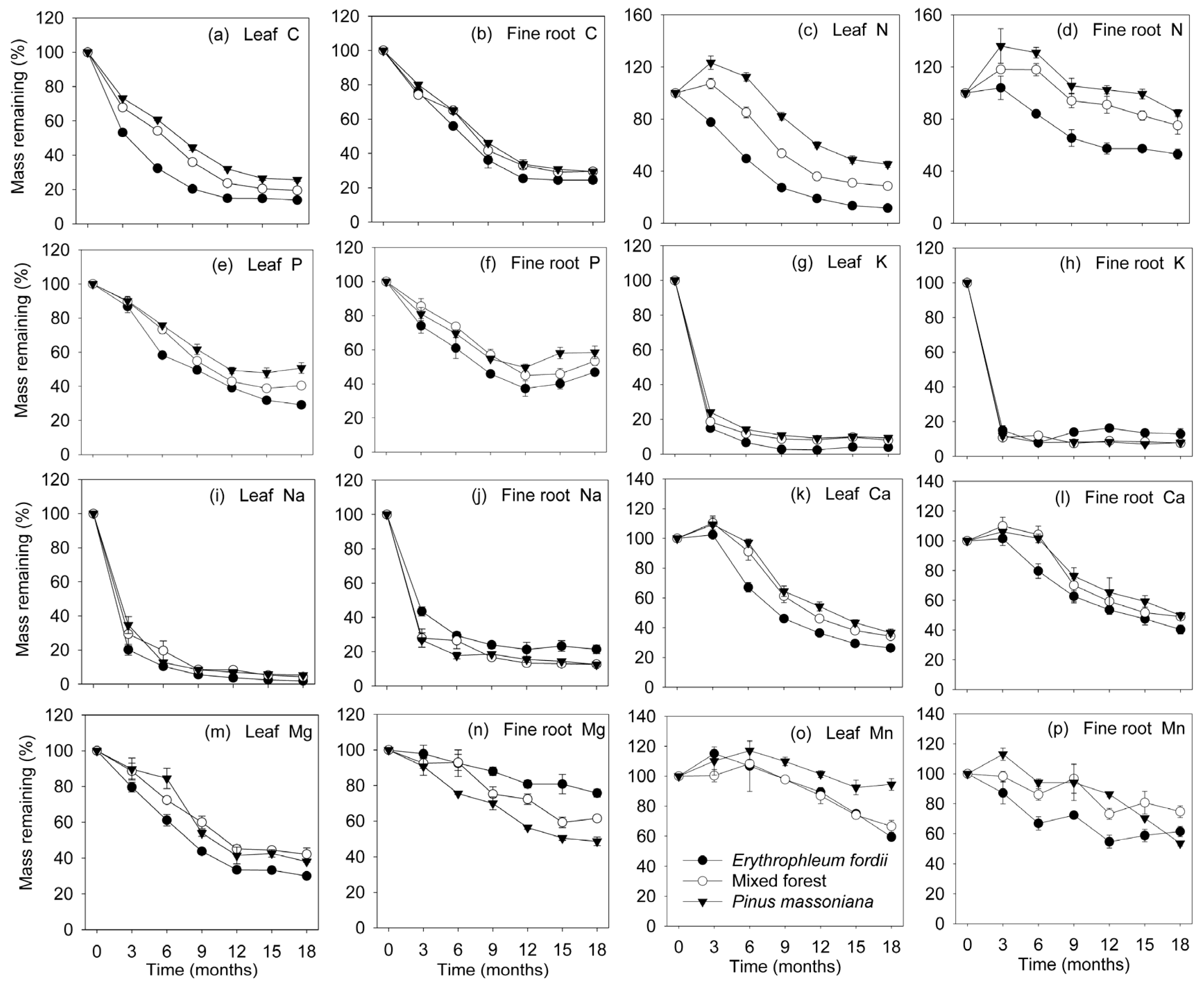
Figure 5.
Dynamics of C/N, C/P, N/P, and lignin/N ratios (mean ± standard error, n = 3) in leaf and fine root litter of three subtropical plantations in China.
Figure 5.
Dynamics of C/N, C/P, N/P, and lignin/N ratios (mean ± standard error, n = 3) in leaf and fine root litter of three subtropical plantations in China.
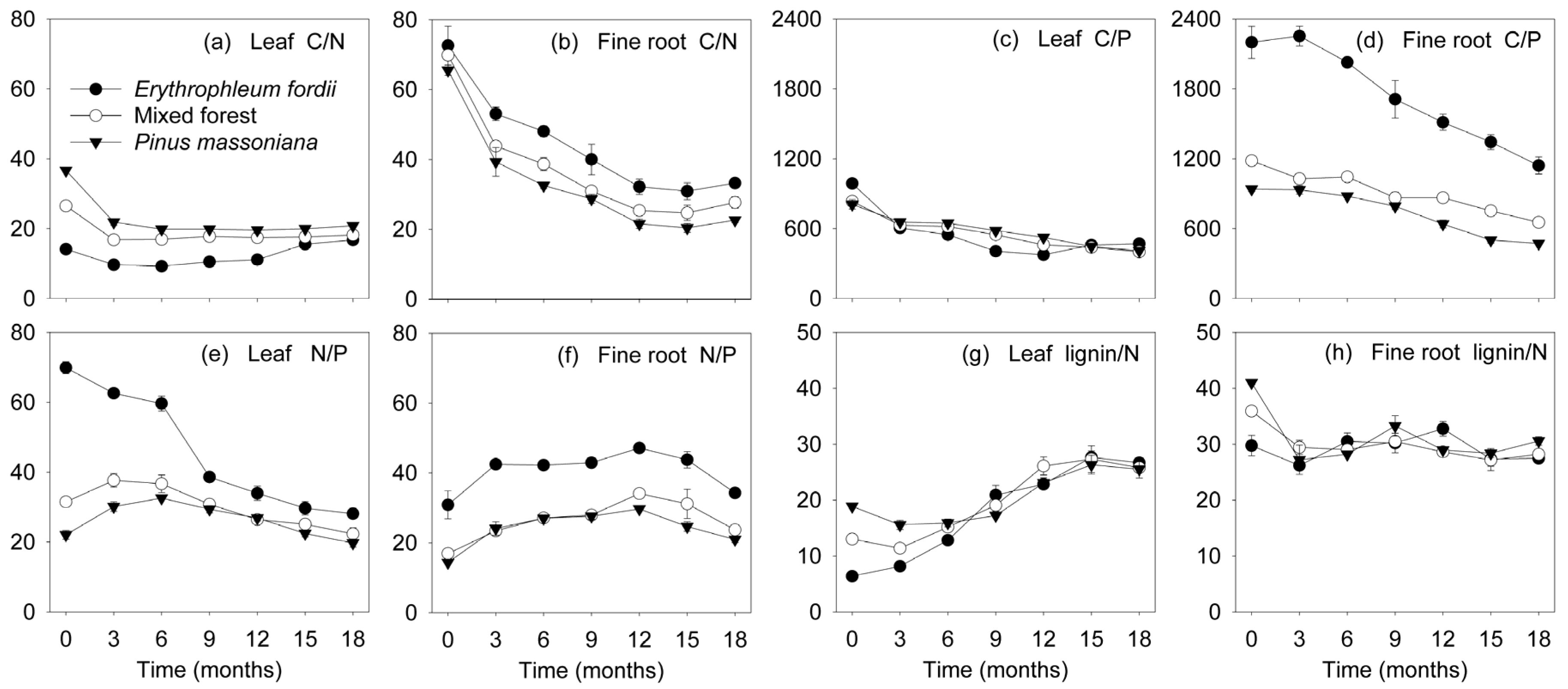
Figure 6.
Soil microbial community attributes determined from three subtropical plantation soils in China. S: species richness index; H’: Shannon’s diversity index; J: Pielou’s evenness index; AM fungi: arbuscular mycorrhizal fungi; SEM fungi: saprotrophic and ectomycorrhizal fungi. Different letters indicate statistically significant differences among plantation types. Error bars represent the standard error of the mean (n = 3).
Figure 6.
Soil microbial community attributes determined from three subtropical plantation soils in China. S: species richness index; H’: Shannon’s diversity index; J: Pielou’s evenness index; AM fungi: arbuscular mycorrhizal fungi; SEM fungi: saprotrophic and ectomycorrhizal fungi. Different letters indicate statistically significant differences among plantation types. Error bars represent the standard error of the mean (n = 3).
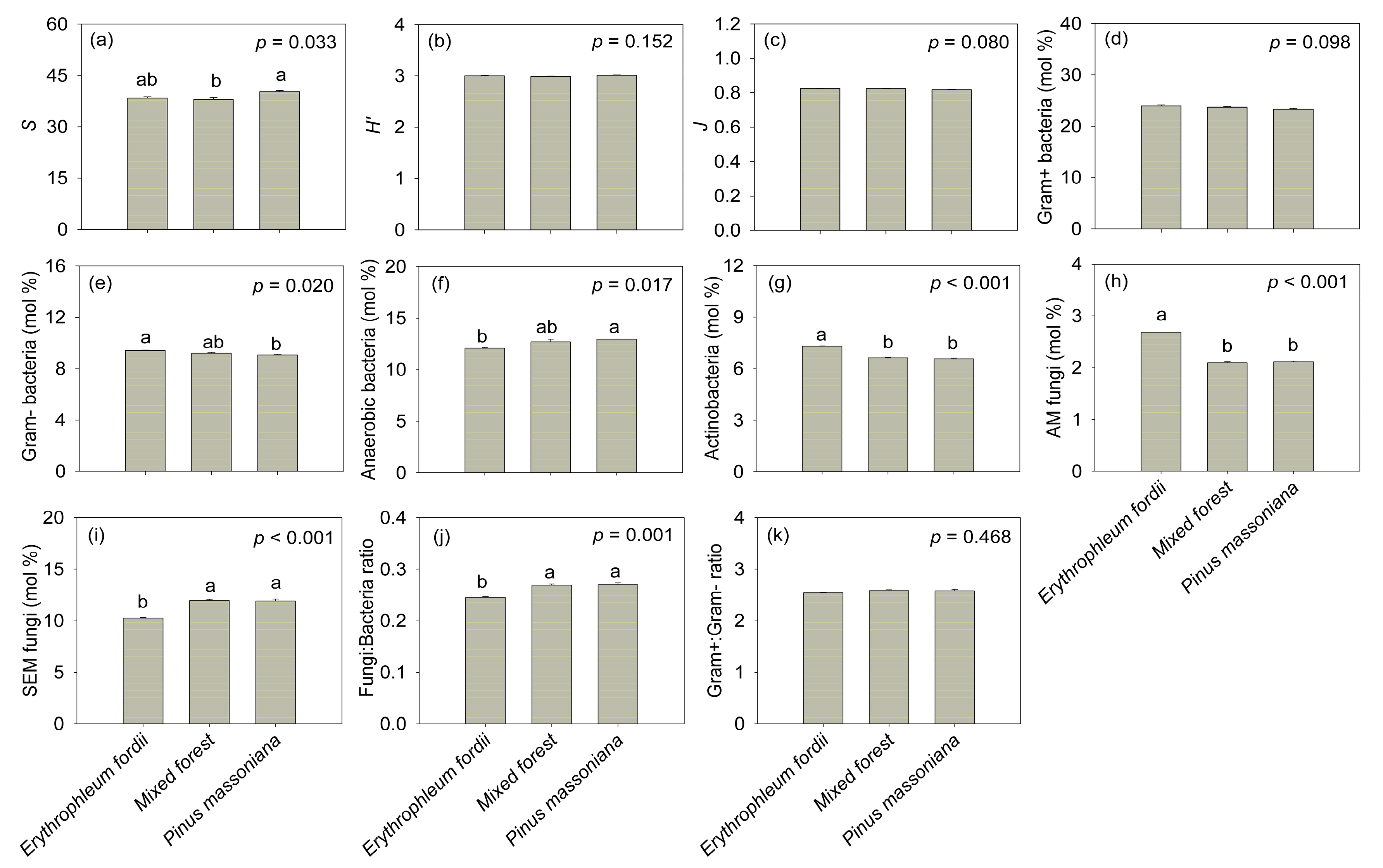
Figure 7.
Redundancy analysis (RDA) of relationships between litter decomposition and soil microbial community attributes. The dashed lines with blue color represent the litter decomposition rates and the solid lines with red color represent the soil microbial community attributes. S: species richness index; H’: Shannon’s diversity index; J: Pielou’s evenness index; G+: gram+ bacteria; G−: gram− bacteria; An: anaerobic bacteria; Ac: actinobacteria; AM: arbuscular mycorrhizal fungi; SEM: saprotrophic and ectomycorrhizal fungi; F/B: fungi:bacteria ratio; G+/G−: gram+ bacteria:gram− bacteria ratio. The p values < 0.05 presented in figure resulted from the Monte Carlo permutation test.
Figure 7.
Redundancy analysis (RDA) of relationships between litter decomposition and soil microbial community attributes. The dashed lines with blue color represent the litter decomposition rates and the solid lines with red color represent the soil microbial community attributes. S: species richness index; H’: Shannon’s diversity index; J: Pielou’s evenness index; G+: gram+ bacteria; G−: gram− bacteria; An: anaerobic bacteria; Ac: actinobacteria; AM: arbuscular mycorrhizal fungi; SEM: saprotrophic and ectomycorrhizal fungi; F/B: fungi:bacteria ratio; G+/G−: gram+ bacteria:gram− bacteria ratio. The p values < 0.05 presented in figure resulted from the Monte Carlo permutation test.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Initial chemical properties of leaf and fine root litter of three subtropical plantations in China. Values are mean ± standard error (n = 3).
Table 1.
Initial chemical properties of leaf and fine root litter of three subtropical plantations in China. Values are mean ± standard error (n = 3).
| Chemical Property (g kg−1) | Leaf | Fine Root | Litter Type | Plantation Type | ||||
|---|---|---|---|---|---|---|---|---|
| Erythrophleum fordii | Mixed Forest | Pinus massoniana | Erythrophleum fordii | Mixed Forest | Pinus massoniana | |||
| C | 483.6 ± 6.89 | 508.0 ± 5.06 | 509.7 ± 5.90 | 602.7 ± 8.48 | 532.5 ± 9.44 | 487.4 ± 7.55 | p < 0.05 | p = 0.130 |
| N | 34.2 ± 0.16 | 19.2 ± 0.33 | 13.9 ± 0.22 | 8.38 ± 0.52 | 7.62 ± 0.17 | 7.44 ± 0.05 | p < 0.001 | p < 0.01 |
| P | 0.49 ± 0.01 | 0.61 ± 0.01 | 0.63 ± 0.03 | 0.28 ± 0.02 | 0.45 ± 0.01 | 0.52 ± 0.00 | p < 0.001 | p < 0.001 |
| K | 4.73 ± 0.11 | 3.58 ± 0.02 | 3.25 ± 0.21 | 1.07 ± 0.04 | 2.78 ± 0.07 | 3.65 ± 0.08 | p < 0.05 | p = 0.604 |
| Na | 1.47 ± 0.04 | 0.82 ± 0.07 | 0.78 ± 0.04 | 0.21 ± 0.01 | 0.40 ± 0.01 | 0.41 ± 0.04 | p < 0.001 | p = 0.182 |
| Ca | 11.2 ± 0.06 | 7.66 ± 0.21 | 6.72 ± 0.33 | 3.33 ± 0.12 | 2.58 ± 0.12 | 2.23 ± 0.07 | p < 0.001 | p < 0.001 |
| Mg | 5.15 ± 0.08 | 2.78 ± 0.04 | 2.03 ± 0.10 | 0.37 ± 0.02 | 1.25 ± 0.02 | 1.71 ± 0.04 | p < 0.01 | p = 0.331 |
| Mn | 0.18 ± 0.00 | 0.84 ± 0.02 | 1.00 ± 0.04 | 0.05 ± 0.00 | 0.11 ± 0.00 | 0.15 ± 0.00 | p < 0.001 | p < 0.01 |
| Lignin | 218.3 ± 0.34 | 250.1 ± 5.21 | 262.4 ± 1.26 | 247.5 ± 0.02 | 273.6 ± 0.02 | 305.2 ± 0.04 | p < 0.001 | p < 0.001 |
| Cellulose | 172.8 ± 5.48 | 133.7 ± 5.20 | 125.7 ± 9.39 | 140.7 ± 1.35 | 140.8 ± 2.27 | 135.4 ± 5.94 | p = 0.418 | p < 0.05 |
| C/N | 14.1 ± 0.21 | 26.5 ± 0.71 | 36.7 ± 0.23 | 72.6 ± 5.55 | 69.9 ± 0.49 | 65.5 ± 1.45 | p < 0.001 | p = 0.138 |
| C/P | 987.7 ± 14.9 | 834.0 ± 18.4 | 808.6 ± 40.5 | 2199 ± 138 | 1182 ± 36.6 | 940.2 ± 20.8 | p < 0.01 | p < 0.01 |
| N/P | 69.9 ± 1.75 | 31.5 ± 0.15 | 22.1 ± 1.25 | 30.9 ± 3.98 | 16.9 ± 0.51 | 14.3 ± 0.13 | p < 0.001 | p < 0.001 |
| Lignin/N | 6.38 ± 0.03 | 13.0 ± 0.21 | 18.9 ± 0.35 | 29.8 ± 1.84 | 35.9 ± 0.75 | 41.0 ± 0.37 | p < 0.001 | p < 0.001 |
Table 2.
Decomposition rate (kA, n = 3) of leaf and fine root litter of three subtropical plantations in China.
Table 2.
Decomposition rate (kA, n = 3) of leaf and fine root litter of three subtropical plantations in China.
| Litter Type | Asymptotic Function | |||
|---|---|---|---|---|
| m | Std. Err. | kA (d−1) | R2adj | |
| Erythrophleum fordii | ||||
| Leaf | 97.96 | 4.33 | 0.24 | 0.97 |
| Fine root | 95.65 | 5.37 | 0.17 | 0.92 |
| Mixed forest | ||||
| Leaf | 99.21 | 3.51 | 0.20 | 0.97 |
| Fine root | 96.74 | 3.85 | 0.14 | 0.94 |
| Pinus massoniana | ||||
| Leaf | 99.60 | 2.85 | 0.19 | 0.98 |
| Fine root | 96.27 | 3.64 | 0.13 | 0.94 |
m: asymptotic level; Std. Err.: standard error; kA: decomposition rate.
Table 3.
Remaining mass of dry mass, organic chemical components, and nutrient elements at the end of the decomposition.
Table 3.
Remaining mass of dry mass, organic chemical components, and nutrient elements at the end of the decomposition.
| Remaining Mass (%) | Leaf | Fine Root | ||||
|---|---|---|---|---|---|---|
| Erythrophleum fordii | Mixed Forest | Pinus massoniana | Erythrophleum fordii | Mixed Forest | Pinus massoniana | |
| Dry mass | 27.1 ± 1.30 | 32.7 ± 0.75 | 35.1 ± 1.10 | 39.5 ± 1.01 | 46.3 ± 1.82 | 48.3 ± 1.39 |
| Lignin | 49.0 ± 1.33 | 56.7 ± 4.08 | 60.8 ± 1.65 | 48.9 ± 0.58 | 58.7 ± 2.79 | 63.5 ± 3.40 |
| Cellulose | 25.8 ± 2.20 | 29.0 ± 0.78 | 31.4 ± 3.32 | 37.5 ± 1.16 | 42.9 ± 2.44 | 44.0 ± 2.90 |
| C | 13.9 ± 0.79 | 19.5 ± 0.83 | 25.6 ± 0.78 | 24.5 ± 2.19 | 29.5 ± 1.00 | 29.3 ± 1.07 |
| N | 11.7 ± 0.21 | 28.7 ± 2.21 | 45.3 ± 2.41 | 53.1 ± 3.79 | 75.1 ± 6.63 | 85.0 ± 2.34 |
| P | 29.1 ± 0.85 | 40.4 ± 0.73 | 50.7 ± 3.11 | 46.9 ± 1.96 | 53.4 ± 2.51 | 58.5 ± 37.4 |
| K | 3.87 ± 0.12 | 8.04 ± 0.40 | 9.34 ± 0.91 | 1.29 ± 3.05 | 7.71 ± 040 | 7.93 ± 0.53 |
| Na | 1.94 ± 0.02 | 4.35 ± 0.46 | 5.36 ± 0.23 | 21.4 ± 2.49 | 12.7 ± 0.11 | 12.5 ± 0.84 |
| Ca | 26.3 ± 0.39 | 34.3 ± 2.49 | 36.6 ± 2.45 | 40.4 ± 2.95 | 49.0 ± 1.93 | 49.9 ± 1.16 |
| Mg | 30.1 ± 1.60 | 42.2 ± 3.53 | 38.1 ± 0.37 | 75.7 ± 2.15 | 61.5 ± 1.64 | 48.7 ± 2.34 |
| Mn | 59.4 ± 0.84 | 66.5 ± 3.95 | 94.5 ± 3.88 | 61.4 ± 3.46 | 74.7 ± 3.92 | 53.7 ± 1.76 |
Values are mean ± standard error (n = 3).
Table 4.
Results of mixed effect model on litter decay (dry mass, organic chemical components, and nutrient elements) for the effects of sampling time, plantation type and litter type.
Table 4.
Results of mixed effect model on litter decay (dry mass, organic chemical components, and nutrient elements) for the effects of sampling time, plantation type and litter type.
| Litter Decay | Factor | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Fixed Effect | Random Effect | ||||||||
| Plantation Type | Litter Type | Time | |||||||
| Num. d.f. | F-Value | p-Value | Num. d.f. | F-Value | p-Value | Estimates | Std. Err. | Wald Z | |
| Dry mass | 2 | 62.966 | 0.013 | 1 | 332.844 | 0.002 | 72.008 | 102.682 | 0.701 |
| Lignin | 2 | 28.687 | <0.001 | 1 | 2.443 | 0.144 | 57.582 | 81.578 | 0.706 |
| Cellulose | 2 | 569.811 | 0.001 | 1 | 3379.03 | <0.001 | 40.041 | 59.682 | 0.671 |
| C | 2 | 19.753 | 0.007 | 1 | 38.391 | 0.003 | 123.823 | 175.675 | 0.705 |
| N | 2 | 146.613 | <0.001 | 1 | 593.284 | <0.001 | 65.290 | 94.215 | 0.693 |
| P | 2 | 30.759 | 0.004 | 1 | 0.476 | 0.529 | 175.069 | 248.144 | 0.706 |
| K | 2 | 0.283 | 0.756 | 1 | 1.461 | 0.238 | 0.292 | 0.630 | 0.464 |
| Na | 2 | 1.343 | 0.285 | 1 | 85.441 | <0.001 | 4.389 | 6.747 | 0.650 |
| Ca | 2 | 69.423 | <0.001 | 1 | 230.415 | <0.001 | 88.551 | 125.928 | 0.703 |
| Mg | 2 | 0.241 | 0.791 | 1 | 18.312 | 0.002 | 89.507 | 127.637 | 0.701 |
| Mn | 2 | 11.660 | <0.001 | 1 | 33.851 | <0.001 | 41.871 | 60.447 | 0.693 |
Num. d.f.: numerator degrees of freedom; Std. Err.: standard error; Wald Z: Wald statistic value.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Luo, D.; Cheng, R.; Shi, Z.; Wang, W. Decomposition of Leaves and Fine Roots in Three Subtropical Plantations in China Affected by Litter Substrate Quality and Soil Microbial Community. Forests 2017, 8, 412. https://doi.org/10.3390/f8110412
AMA Style
Luo D, Cheng R, Shi Z, Wang W. Decomposition of Leaves and Fine Roots in Three Subtropical Plantations in China Affected by Litter Substrate Quality and Soil Microbial Community. Forests. 2017; 8(11):412. https://doi.org/10.3390/f8110412
Chicago/Turabian StyleLuo, Da, Ruimei Cheng, Zuomin Shi, and Weixia Wang. 2017. "Decomposition of Leaves and Fine Roots in Three Subtropical Plantations in China Affected by Litter Substrate Quality and Soil Microbial Community" Forests 8, no. 11: 412. https://doi.org/10.3390/f8110412
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.