
Diameter Growth, Biological Rotation Age and Biomass of Chinese Fir in Burning and Clearing Site Preparations in Subtropical China


Abstract
:1. Introduction
2. Experimental Section
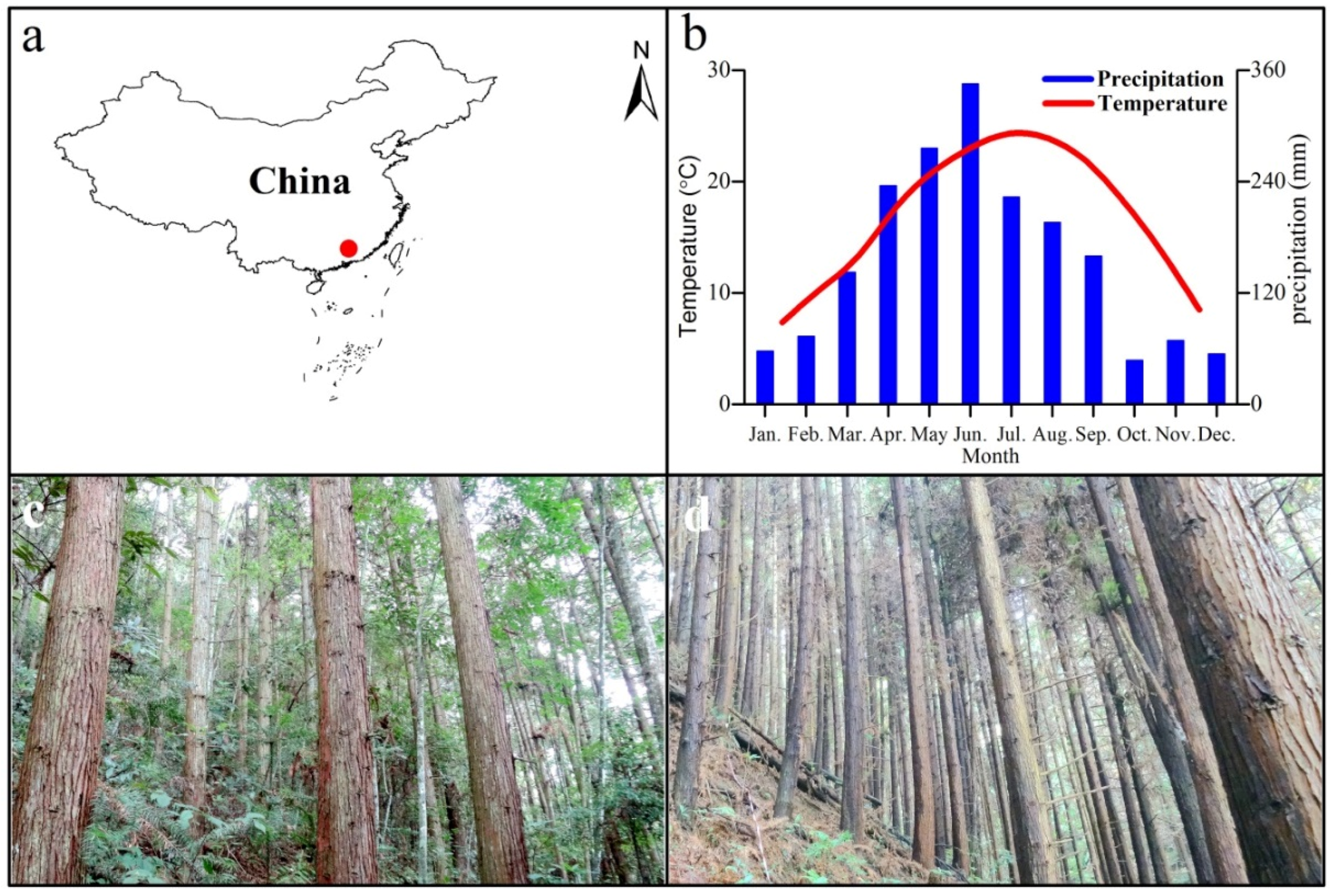
2.1. Study Area
2.2. Site Preparation in the Region
2.3. Experimental Design and Data Collection
2.4. Tree-Ring Analysis
2.5. Analysis of Growth Changes
2.6. Bark Thickness
2.7. Biomass Estimate
3. Results
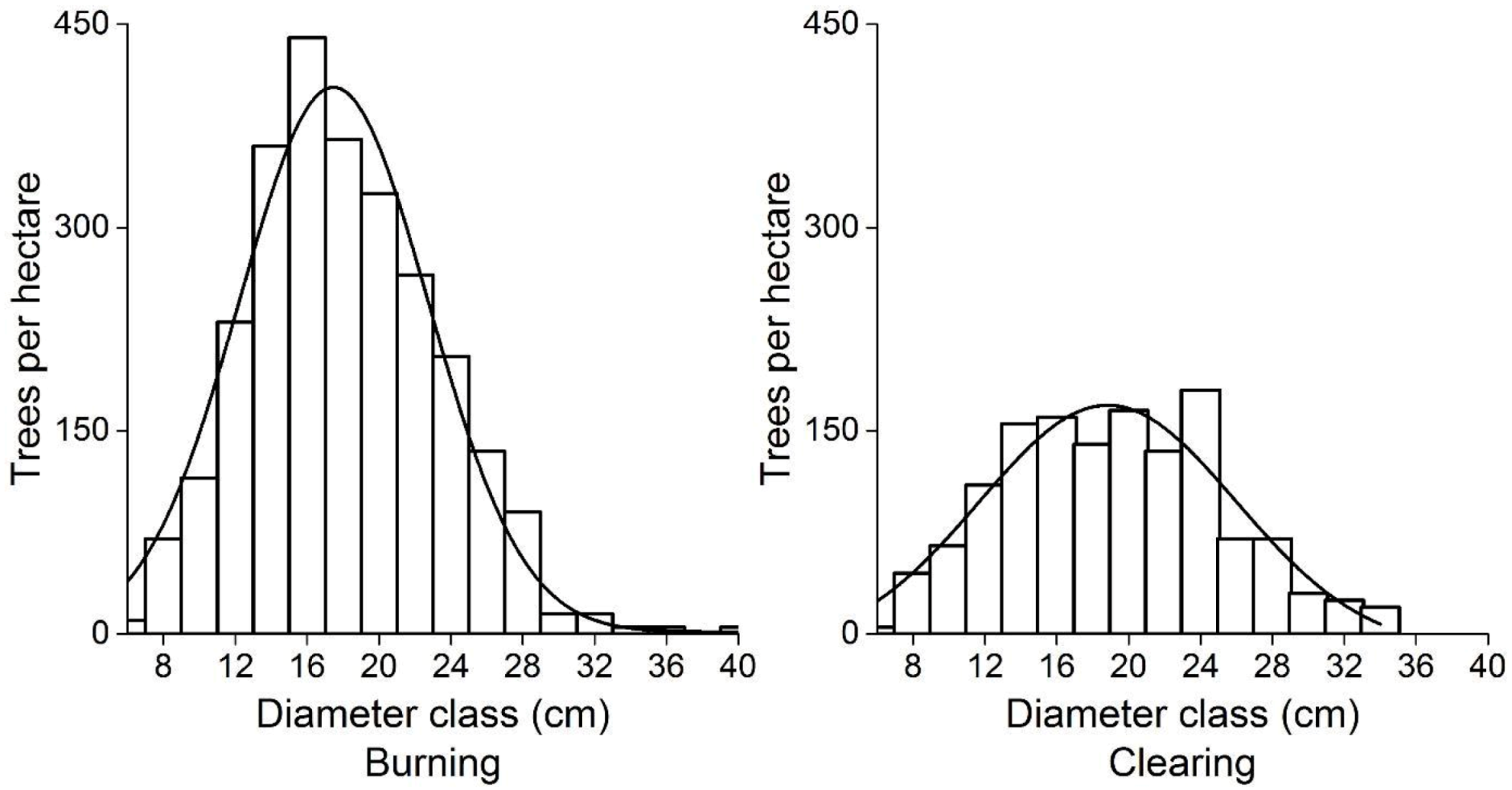
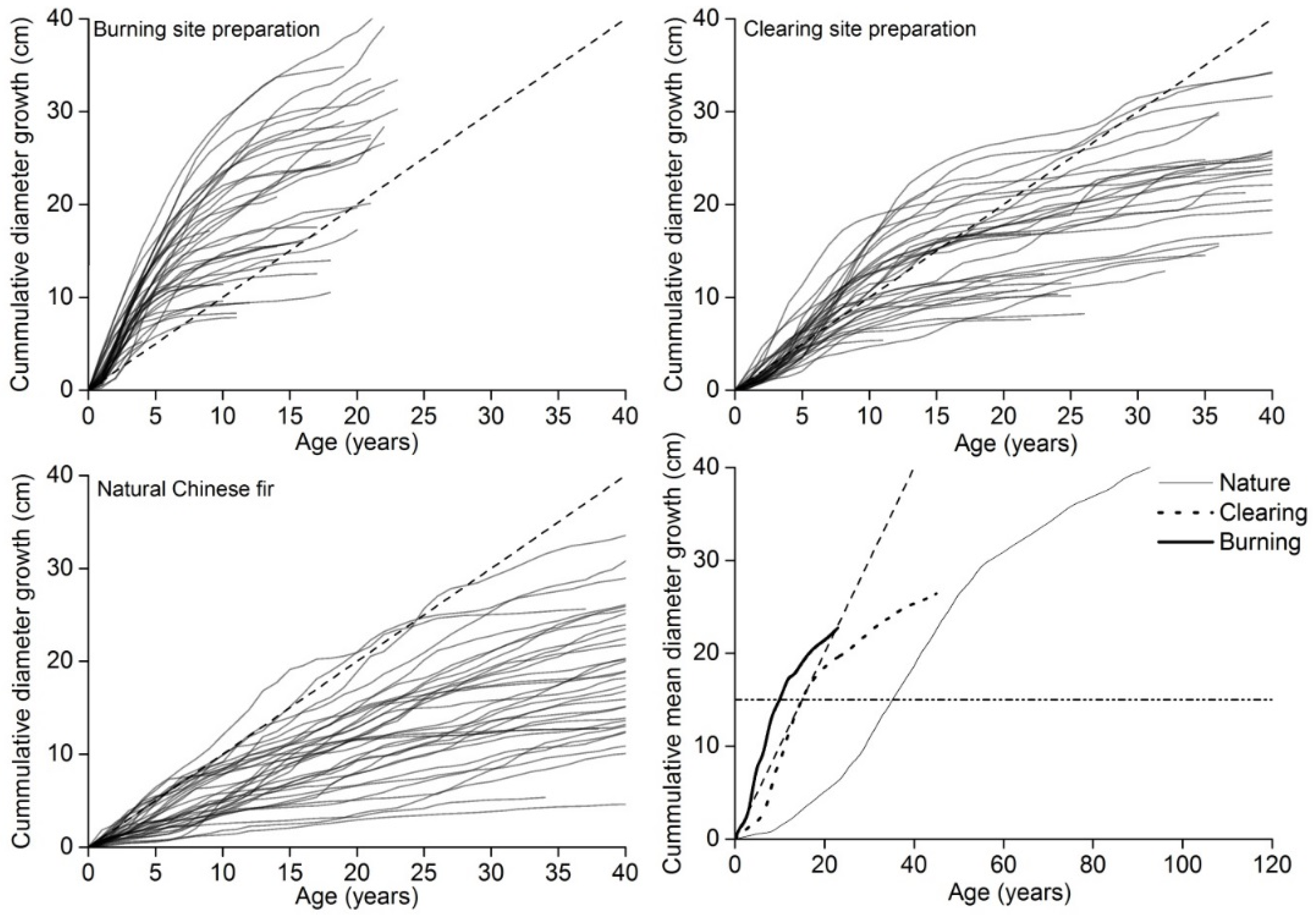
3.1. Structural and Growth Characteristics of Planted and Natural Chinese Fir
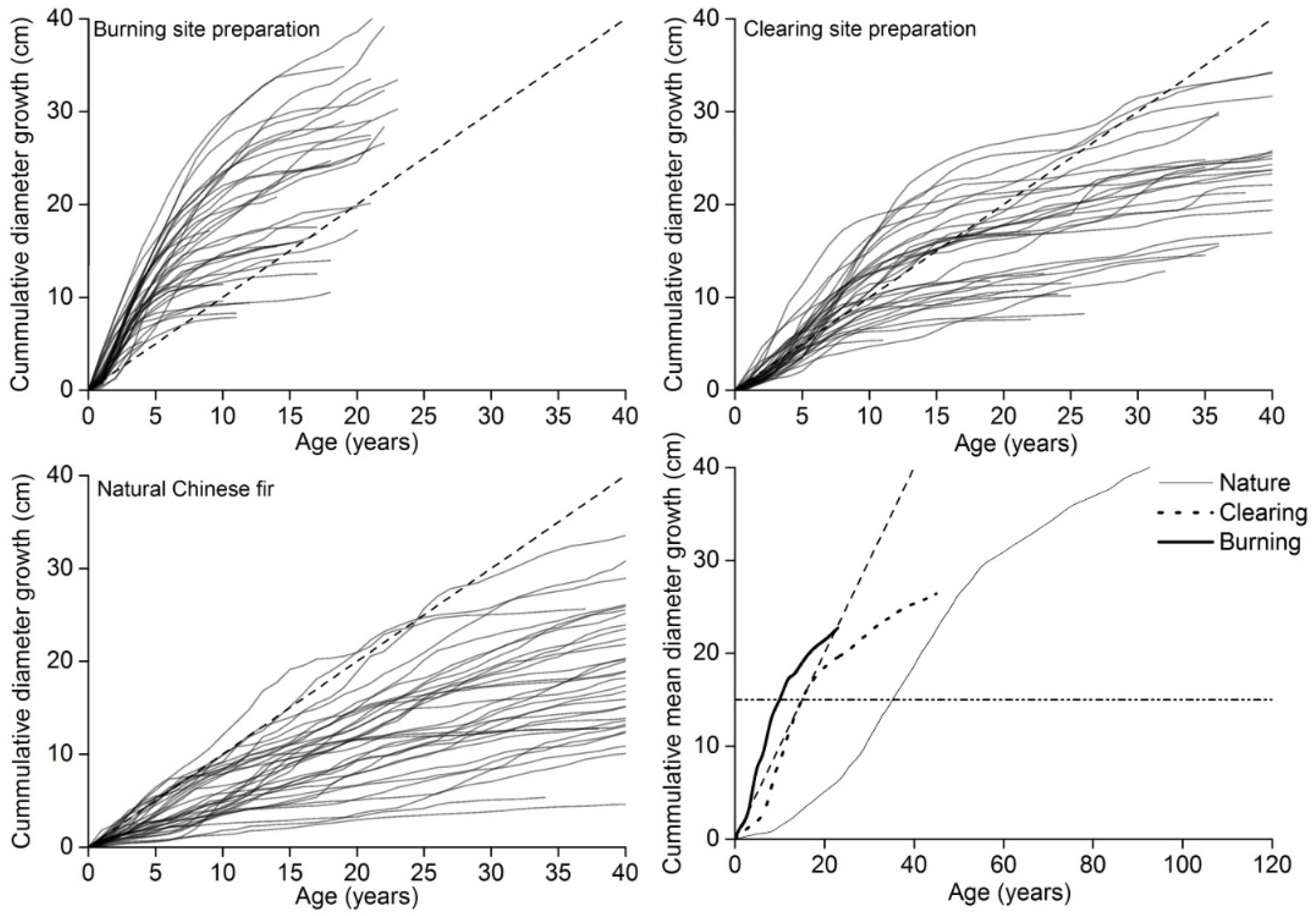
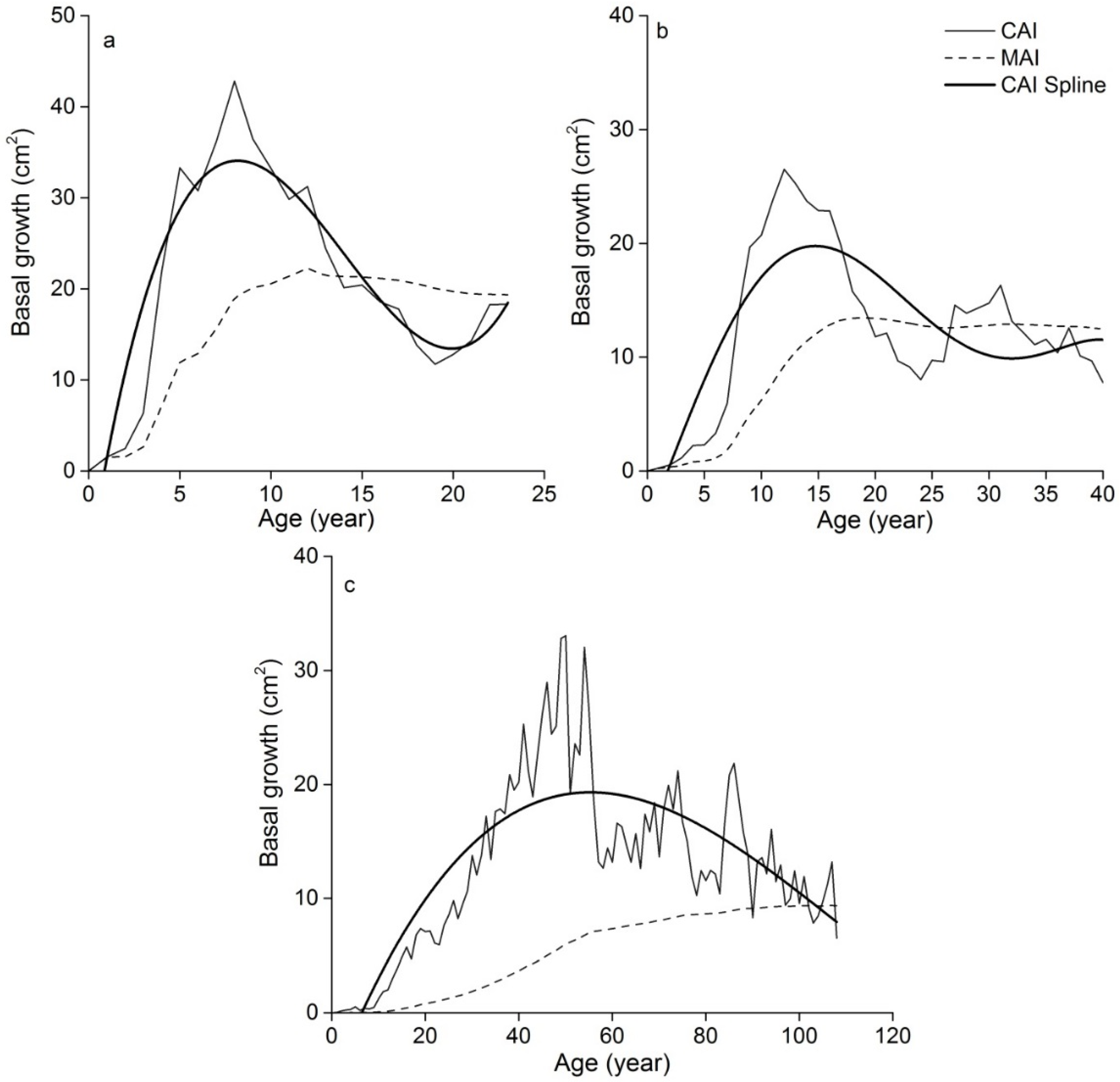
3.2. BRAs
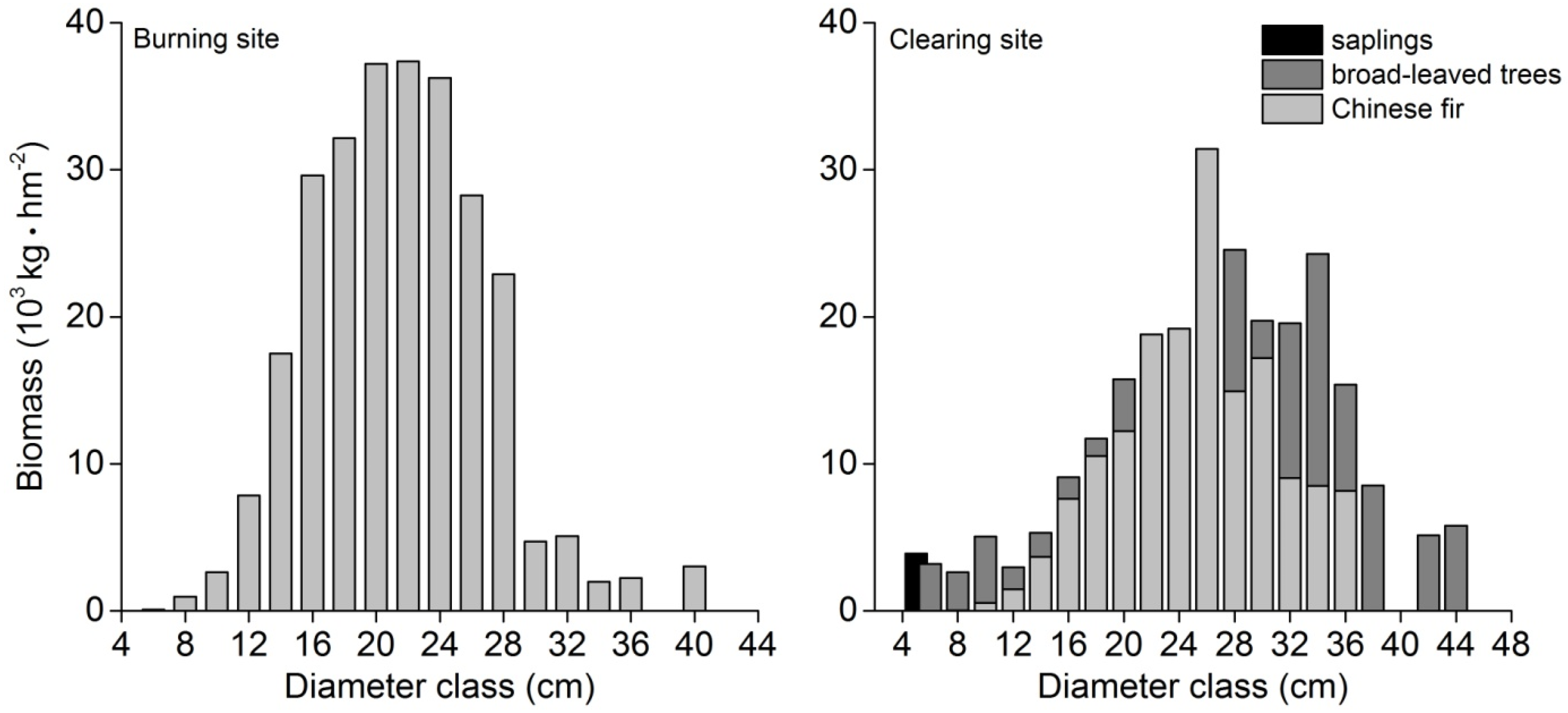
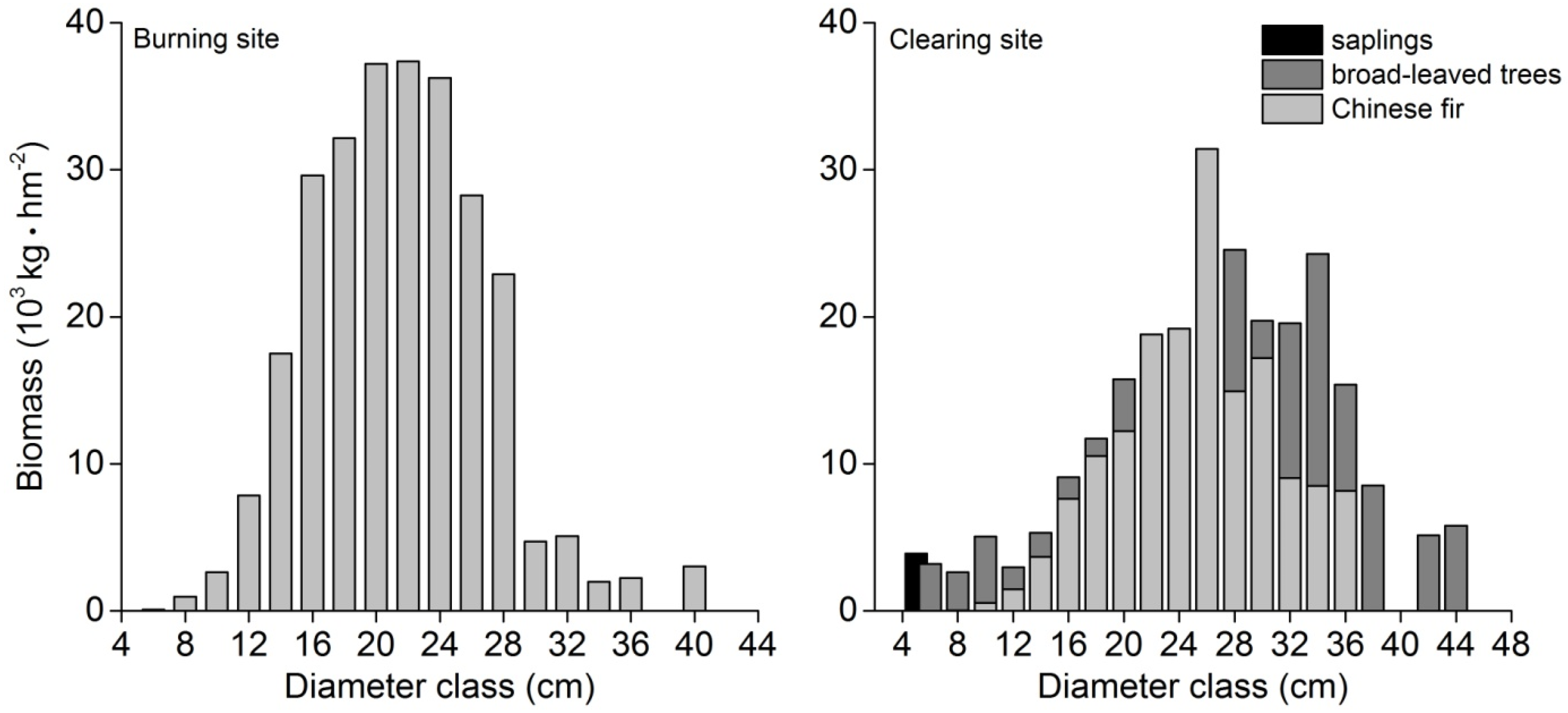
3.3. Biomass
4. Discussion
4.1. Evaluation of the Mean Diameter and BRA
4.2. Evaluation of the Aboveground Biomass
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sample, V.A. Sustainability in Forestry: Origins, Evolution and Prospects; Pinchot Institute for Conservation: Washington, DC, USA, 2004. [Google Scholar]
- Borders, B.; Bailey, R. Loblolly pine—Pushing the limits of growth. South. J. Appl. For. 2001, 25, 69–74. [Google Scholar]
- Jones, P.D.; Edwards, S.L.; Demarais, S.; Ezell, A.W. Vegetation community responses to different establishment regimes in loblolly pine (Pinus taeda) plantations in southern Mississippi, USA. For. Ecol. Manag. 2009, 257, 553–560. [Google Scholar] [CrossRef]
- Zutter, B.; Miller, J. Eleventh-year response of loblolly pine and competing vegetation to woody and herbaceous plant control on a Georgia flatwoods site. South. J. Appl. For. 1998, 22, 88–95. [Google Scholar]
- Clark, D.A.; Clark, D.B. Life-history diversity of canopy and emergent trees in a neotropical rain-forest. Ecol. Monogr. 1992, 62, 315–344. [Google Scholar] [CrossRef]
- Clark, D.A.; Clark, D.B. Getting to the canopy: Tree height growth in a neotropical rain forest. Ecology 2001, 82, 1460–1472. [Google Scholar] [CrossRef]
- Condit, R. Research in large, long-term tropical forest plots. Trends Ecol. Evolut. 1995, 10, 18–22. [Google Scholar] [CrossRef]
- Lieberman, D.; Lieberman, M.; Hartshorn, G.S.; Peralta, R. Growth rates and age-size relationships of tropical wet forest trees in Costa Rica. J. Tropical Ecol. 1985, 1, 97–109. [Google Scholar] [CrossRef]
- Brienen, J.W.; Zuidema, P.A. Lifetime growth patterns and ages of Bolivian rain forest trees obtained by tree ring analysis. J. Ecol. 2006, 94, 481–493. [Google Scholar] [CrossRef]
- Canham, C.D. Suppression and release during canopy recruitment in Acer saccharum. Bull. Torrey Bot. Club 1985, 112, 134–145. [Google Scholar] [CrossRef]
- Landis, R.M.; Peart, D.R. Early performance predicts canopy attainment across life histories in subalpine forest trees. Ecology 2005, 86, 63–72. [Google Scholar] [CrossRef]
- Lorimer, C.G.; Frelich, L.E. A methodology for estimating canopy disturbance frequency and intensity in dense temperate forests. Can. J. For. 1989, 19, 651–663. [Google Scholar] [CrossRef]
- Lusk, C.H.; Smith, B. Life history differences and tree species coexistence in an old-growth New Zealand rain forest. Ecology 1998, 79, 795–806. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. Radial-growth averaging criteria for reconstructing disturbance histories from presettlement-origin oaks. Ecol. Monogr. 1997, 67, 225–249. [Google Scholar] [CrossRef]
- Karlsson, A. Site preparation of abandoned fields and early establishment of planted small-sized seedlings of silver birch. New For. 2002, 23, 159–175. [Google Scholar] [CrossRef]
- Lane, V.R.; Miller, K.V.; Castleberry, S.B.; Miller, D.A.; Wigley, T.B.; Marsh, G.M.; Mihalco, R.L. Plant community responses to a gradient of site preparation intensities in pine plantations in the Coastal Plain of North Carolina. For. Ecol. Manag. 2011, 262, 370–378. [Google Scholar] [CrossRef]
- Miller, D.A.; Chamberlain, M.J. Plant community response to burning and herbicide site preparation in eastern Louisiana, USA. For. Ecol. Manag. 2008, 255, 774–780. [Google Scholar] [CrossRef]
- Thornton, D.H.; Wirsing, A.J.; Roth, J.D.; Murray, D.L. Complex effects of site preparation and harvest on snowshoe hare abundance across a patchy forest landscape. For. Ecol. Manag. 2012, 280, 132–139. [Google Scholar] [CrossRef]
- Yildiz, O.; Esen, D.; Karaoz, O.M.; Sarginci, M.; Toprak, B.; Soysal, Y. Effects of different site preparation methods on soil carbon and nutrient removal from Eastern beech regeneration sites in Turkey’s Black Sea region. Appl. Soil Ecol. 2010, 45, 49–55. [Google Scholar] [CrossRef]
- Miller, D.A.; Wigley, T.B.; Miller, K.V. Managed Forests and Conservation of Terrestrial Biodiversity in the Southern United States. J. For. 2009, 107, 197–203. [Google Scholar]
- Wang, J.L.; Wang, H.M.; Fu, X.L.; Xu, M.; Wang, Y. Effects of site preparation treatments before afforestation on soil carbon release. For. Ecol. Manag. 2016, 361, 277–285. [Google Scholar] [CrossRef]
- Fernández, I.; Cabaneiro, A.; Carballas, T. Carbon mineralization dynamics in soils after wildfires in two Galician forests. Soil Bio. Biochem. 1999, 31, 1853–1865. [Google Scholar] [CrossRef]
- Phillips, R.L.; Zak, D.R.; Holmes, W.E.; White, D.C. Microbial community composition and function beneath temperate trees exposed to elevated atmospheric carbon dioxide and ozone. Oecologia 2002, 131, 236–244. [Google Scholar]
- Hubbard, R.M.; Vose, J.M.; Clinton, B.D.; Elliott, K.J.; Knoepp, J.D. Stand restoration burning in oak–pine forests in the southern Appalachians effects on aboveground biomass and carbon and nitrogen cycling. For. Ecol. Manag. 2004, 190, 311–321. [Google Scholar] [CrossRef]
- Wüthrich, C.; Schaub, D.; Weber, M.; Marxer, P.; Conedera, M. Soil respiration and soil microbial biomass after fire in a sweet chestnut forest in southern Switzerland. Catena 2002, 48, 201–215. [Google Scholar] [CrossRef]
- Boerner, R.E.J.; Decker, K.L.M.; Sutherland, E.K. Prescribed burning effects on soil enzyme activity in a southern Ohio hardwood forest: A landscape-scale analysis. Soil Biol. Biochem. 2000, 32, 899–908. [Google Scholar] [CrossRef]
- McCarthy, D.R.; Brown, K.J. Soil respiration responses to topography, canopy cover, and prescribed burning in an oak-hickory forest in southeastern Ohio. For. Ecol. Manag. 2006, 237, 94–102. [Google Scholar] [CrossRef]
- López, L.; Villalba, R.; Bravo, F. Cumulative diameter growth and biological rotation age for seven tree species in the Cerrado biogeographical province of Bolivia. For. Ecol. Manag. 2013, 292, 49–55. [Google Scholar] [CrossRef]
- Campos, J.J.; Finegan, B.; Villalobos, R. Diversified forest management: Improvement of goods and services from the biodiversity of a neotropical forest. Revista For. Centroamericana 2001, 36, 6–13. [Google Scholar]
- Ni, J. Impacts of climate change on Chinese ecosystems, key vulnerable regions and potential thresholds. Reg. Environ. Chang. 2011, 11, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.Y. Vegetation of China; Science Press: Beijing, China, 1980; p. 825. [Google Scholar]
- Xu, Y.; Shen, Z.H.; Ying, L.X.; Ciais, P.; Liu, H.Y.; Piao, S.L.; Wen, C.; Jiang, Y.X. The exposure, sensitivity and vulnerability of natural vegetationin China to climate thermal variability (1901–2013): An indicator-based approach. Ecol. Indic. 2016, 63, 258–272. [Google Scholar] [CrossRef]
- Sheng, H.; Yang, Y.S.; Yang, Z.J.; Chen, G.S.; Xie, J.S.; Guo, J.F.; Zou, S.Q. The dynamic response of soil respiration to land-use changes in subtropical China. Glob. Chang. Biol. 2010, 16, 1107–1121. [Google Scholar] [CrossRef]
- Xu, J. China’s new forests aren’t as green as they seem. Nature 2011, 477, 370. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.S.; Yang, Z.J.; Gao, R.; Xie, J.S.; Guo, J.F.; Huang, Z.Q.; Yang, Y.S. Carbon storage in a chronosequence of Chinese fir plantations in southern China. For. Ecol. Manag. 2013, 300, 68–76. [Google Scholar] [CrossRef]
- FAO. Global Forest Resource Assessment 2005. Available online: ftp://ftp.fao.org/docrep/fao/008/A0400E/A0400E00.pdf (accessed on 12 August 2015).
- West, P.W. Growing Plantation Forests; Springer-Verlag Berlin Heidelberg: Berlin, Germany, 2006. [Google Scholar]
- Liu, X.Z.; Xiao, Z.Y.; Ma, J.H. Scientific Survey and Study on the Forest Ecosystem in Jiangxi Nature Reserve; China Forestry Publishing House: Beijing, China, 2002; pp. 14–53. [Google Scholar]
- MacDonald, R.L.; Chen, H.Y.H.; Palik, B.P.; Prepas, E.E. Influence of harvesting on understory vegetation along a boreal riparian-upland gradient. For. Ecol. Manag. 2014, 312, 138–147. [Google Scholar] [CrossRef]
- Primicia, I.; Camarero, J.J.; Janda, P.; Cada, V.; Morrissey, R.C.; Trotsiuk, V.; Bace, R.; Teodosiu, M.; Svoboda, M. Age, competition, disturbance and elevation effects on tree and stand growth response of primary Picea abies forest to climate. For. Ecol. Manag. 2015, 354, 77–86. [Google Scholar] [CrossRef]
- De Ridder, M.; Trouet, V.; Van den Bulcke, J.; Hubau, W.; Van Acker, J.; Beeckman, H. A tree-ring based comparison of Terminalia superba climate–growth relationships in West and Central Africa. Trees 2013, 27, 1225–1238. [Google Scholar] [CrossRef]
- Yamaguchi, D.K. A simple method for cross-dating increment cores from living trees. Can. J. For. Res. 1991, 21, 414–416. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree-ring Res. 2001, 57, 205–221. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 44, 69–75. [Google Scholar]
- Stahle, D.W.; Mushove, P.T.; Cleaveland, M.K.; Roig, F.; Haynes, G.A. Management implications of annual growth rings in Pterocarpus angolensis from Zimbabwe. For. Ecol. Manag. 1999, 124, 217–229. [Google Scholar] [CrossRef]
- Brienen, J.W.; Zuidema, P.A.; During, H.J. Autocorrelated growth of tropical forest trees: Unraveling patterns and quantifying consequences. For. Ecol. Manag. 2006, 237, 179–190. [Google Scholar] [CrossRef]
- Biondi, F.; Qeadan, F. A theory-driven approach to tree-tring standardization: Defining the biological trend from expected basal area increment. Tree Ring Res. 2008, 64, 81–96. [Google Scholar] [CrossRef]
- Wang, Z.C.; Du, H.; Song, T.Q.; Peng, W.X.; Zeng, F.P.; Zeng, Z.X.; Zhang, H. Allometric models of major tree species and forest biomass in Guangxi. Acta Ecol. Sinica 2015, 35, 4462–4472. [Google Scholar]
- Therrell, M.D.; Stahle, D.W.; Mukelabai, M.M.; Shugart, H.H. Age, and radial growth dynamics of Pterocarpus angolensis in southern Africa. For. Ecol. Manag. 2007, 244, 24–31. [Google Scholar] [CrossRef]
- Cai, C.; Qiu, S.; Cai, S. A preliminary study on shortening rotation age of Chinese fir plantation. South China For. Sci. 1993, 2, 27–30. (In Chinese) [Google Scholar]
- Yang, Y.; He, Z.; Qiu, R.; Yu, X.; Huang, B. Growth pattern of 29-year-old Chinese fir grown from seed in different rotations. Sci. Silvae Sinicae 1999, 35, 32–36. (In Chinese) [Google Scholar]
- Zhang, Z.; Cai, X.; Zhu, B. Study on the fixing of cutting age of the Chinese fir man-made forest. Acta Agric. Univ. Jiangxiensis 1992, 6, 46–51. (In Chinese) [Google Scholar]
- Jie, J.; Zhan, Y.; Wu, K. Study on propor rotation period for artificial stands of Cunninghamia lanceolata in Jiangxi province. South China For. Sci. 2000, 6, 4–8. (In Chinese) [Google Scholar]
- De Ridder, M.; Van den Bulcke, J.; Van Acker, J.; Beeckman, H. Tree-ring analysis of an African long-lived pioneer species as a tool for sustainable forest management. For. Ecol. Manag. 2013, 304, 417–426. [Google Scholar] [CrossRef]
- Pan, J.; Zheng, L.; Chen, H.; Ye, H.; Wu, L. Growth with intermediate cuttings in mature Chinese fir plantations. J. Zhejiang A F Univ. 2014, 31, 291–295. (In Chinese) [Google Scholar]
- Hemery, G.E. Forest management and silvicultural responses to projected climate change impacts on European broadleaved trees and forests. Intern. For. Rev. 2008, 10, 591–607. [Google Scholar] [CrossRef]
- Way, D.A.; Oren, R. Differential responses to changes in growth temperature between trees from different functional groups and biomes: A review and synthesis of data. Tree Physiol. 2010, 30, 669–688. [Google Scholar] [CrossRef] [PubMed]
- Doley, D. The response of forests to climate change: The role of silviculture in conserving threatened species. Aust. For. 2010, 73, 115–125. [Google Scholar] [CrossRef]
- Leoni, J.M.; Da Fonseca Junior, S.F.; Sch Ngart, J. Growth and population structure of the tree species Malouetia tamaquarina (Aubl.) (Apocynaceae) in the central Amazonian floodplain forests and their implication for management. For. Ecol. Manag. 2011, 29, 597–611. [Google Scholar] [CrossRef]
- Nebel, G.; Dragsted, J.; Simonsen, T.R.; Vanclay, J.K. The Amazon flood plain forest tree Maquira coriacea (Karsten) C.C. Berg: Aspects of ecology and management. For. Ecol. Manag. 2001, 150, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Schöngart, J. Growth-Oriented Logging (GOL): A new concept towards sustainable forest management in Central Amazonian várzea floodplains. For. Ecol. Manag. 2008, 256, 46–58. [Google Scholar] [CrossRef]
- Bogino, S.M.; Villalba, R. Radial growth and biological rotation age of Prosopis caldenia Burkart in Central Argentina. J. Arid Environ. 2008, 72, 16–23. [Google Scholar] [CrossRef]
- Dorado, M.; Astini, E.; Verzino, G.; Di Rienzo, J.; Perpi, A.E. Growth curves for Pinus elliottii, Pinus taeda and Pinus radiata in two areas of the Calamuchita Valley(Córdoba, Argentina). For. Ecol. Manag. 1997, 95, 173–181. [Google Scholar] [CrossRef]
- Verzino, G.; Ingaramo, P.; Joseau, J.; Astini, E.; Di Rienzo, J.; Dorado, M. Basal area growth curves for Pinus patula in two areas of the Calamuchita Valley, Córdoba, Argentina. For. Ecol. Manag. 1999, 124, 185–192. [Google Scholar] [CrossRef]
- Sist, P.; Fimbel, R.; Sheil, D. Towards sustainable management of mixed dipterocarp forests of Southeast Asia: Moving beyond minimum diameter cutting limits. Environ. Conserv. 2003, 30, 364–374. [Google Scholar] [CrossRef]
- Brienen, J.W.; Zuidema, P.A. The use of tree rings in tropical forest management: Projecting timber yields of four Bolivian tree species. For. Ecol. Manag. 2006, 226, 256–267. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, J.F.; Chen, G.S.; Xie, J.S.; Gao, R.; Zhen, L.; Zhao, J. Carbon and nitrogen pools in Chinese fir and evergreen broadleaved forests and changes associated with felling and burning in mid-subtropical China. For. Ecol. Manag. 2005, 216, 216–226. [Google Scholar] [CrossRef]
- Yang, M.; Wang, S.L.; Zhang, W.D.; Wang, Q.K. Dynamics of biomass and nutrient accumulation in a Chinese-fir plantation. Chin. J. Appl. Ecol. 2010, 21, 1674–1680. (In Chinese) [Google Scholar]
- Yu, Y.F.; Song, T.Q.; Zeng, F.P.; Peng, W.X.; Wen, Y.G.; Huang, C.B.; Wu, Q.B.; Zeng, Z.X.; Yu, Y. Dynamic changes of biomass and its allocation in Cunninghamia lanceolata plantations of different stand ages. Chin. J. Ecol. 2013, 32, 1660–1666. (In Chinese) [Google Scholar]
- Wu, J.; Wang, Y.; Yang, Y.; Zhu, T.; Zhu, X. Effects of crop tree release on stand growth and stand structure of Cunninghamia lanceolata plantation. Chin. J. Appl. Ecol. 2015, 26, 340–348. (In Chinese) [Google Scholar]
- Passauer, D.P.; Aust, W.M.; Bolding, M.C.; Strahm, B.D.; Burger, J.M.; Patterson, S.C.; Vance, E. Potential above-ground biomass losses from severe soil rutting during wet weather timber harvests of coastal plain loblolly pine (Pinus taeda) plantations mitigated by mechanical site preparation. For. Ecol. Manag. 2013, 307, 266–273. [Google Scholar] [CrossRef]
- Fox, T.R.; Kyle, K.H.; Andrews, L.J.; Aust, W.M.; Burger, J.A.; Hansen, G.H. Long-term effects of drainage, bedding, and fertilization on growth of loblolly pine (Pinus taeda L.) in the coastal plain of Virginia. South. J. Appl. For. 2005, 29, 205–214. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plantation | Nature Chinese Fir Forest | ||
|---|---|---|---|
| Burning Site Preparation | Clearing Site Preparation | ||
| Latitude | 24°32′25″ N | 24°32′09″ N | 24°32′20″ N |
| Longitude | 114°28′19″ E | 114°27′56″ E | 114°27′25″ E |
| Altitude (m.a.s.l.) | 765 | 695 | 793 |
| Planting density (trees·hm−2) | 2500~3000 | 1500~2000 | No data |
| Planting pit size (cm) | 50 × 50 × 40 | 50 × 50 × 40 | No data |
| Slope | <30° | <25° | <25° |
| Age of stand | 26 | 36 | >80 |
| Mean diameter of stand (cm) | 18 ± 5.2 | 19 ± 6.0 | No data |
| No. of increment cores | 37 | 34 | 34 |
| MAI (cm) | 1.10 ± 0.38a | 0.80 ± 0.25b | 0.56 ± 0.15c |
| BA-MAI (cm2·year−1) | 4.28 ± 1.37a | 3.03 ± 0.95b | 2.24 ± 0.95c |
| Biological rotation age (BRA, year) | 15 | 26 | >100 |
| Diameter at BRA (cm) | 19 | 23 | >50 |
| Local management guides for cutting cycle and minimum cutting diameter | |||
| Cutting cycle (CC, year) | 25 | 25 | No data |
| Minimum cutting diameter (MCD, cm) | 15 | 15 | No data |
| Stand Characteristics | Burning Site Preparation | Clearing Site Preparation |
|---|---|---|
| Chinese fir biomass (ton·hm−2) | 269.8 | 163.4 |
| Sapling biomass (ton·hm−2) | 0 | 3.9 |
| Broad leaved trees biomass (ton·hm−2) | 0 | 84.7 |
| Total biomass (ton·hm−2) | 269.8 | 252.0 |
| Chinese fir density (trees·hm−2) | 2655 ± 442 | 1610 ± 477 |
| Sapling density (trees·hm−2) (1–5 cm·dbh) | 0 | 2295 ± 655 |
| other tree density (trees·hm−2) (>5 cm·dbh) | 0 | 695 ± 320 |
| Total trees per hectare (>5 cm·dbh) | 2655 ± 442 | 2305 ± 539 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Meng, S.; Liu, Q. Diameter Growth, Biological Rotation Age and Biomass of Chinese Fir in Burning and Clearing Site Preparations in Subtropical China. Forests 2016, 7, 177. https://doi.org/10.3390/f7080177
Zhou H, Meng S, Liu Q. Diameter Growth, Biological Rotation Age and Biomass of Chinese Fir in Burning and Clearing Site Preparations in Subtropical China. Forests. 2016; 7(8):177. https://doi.org/10.3390/f7080177
Chicago/Turabian StyleZhou, Hua, Shengwang Meng, and Qijing Liu. 2016. "Diameter Growth, Biological Rotation Age and Biomass of Chinese Fir in Burning and Clearing Site Preparations in Subtropical China" Forests 7, no. 8: 177. https://doi.org/10.3390/f7080177