
Regional Instability in the Abundance of Open Stands in the Boreal Forest of Eastern Canada

Abstract
:1. Introduction
2. Materials and Methods
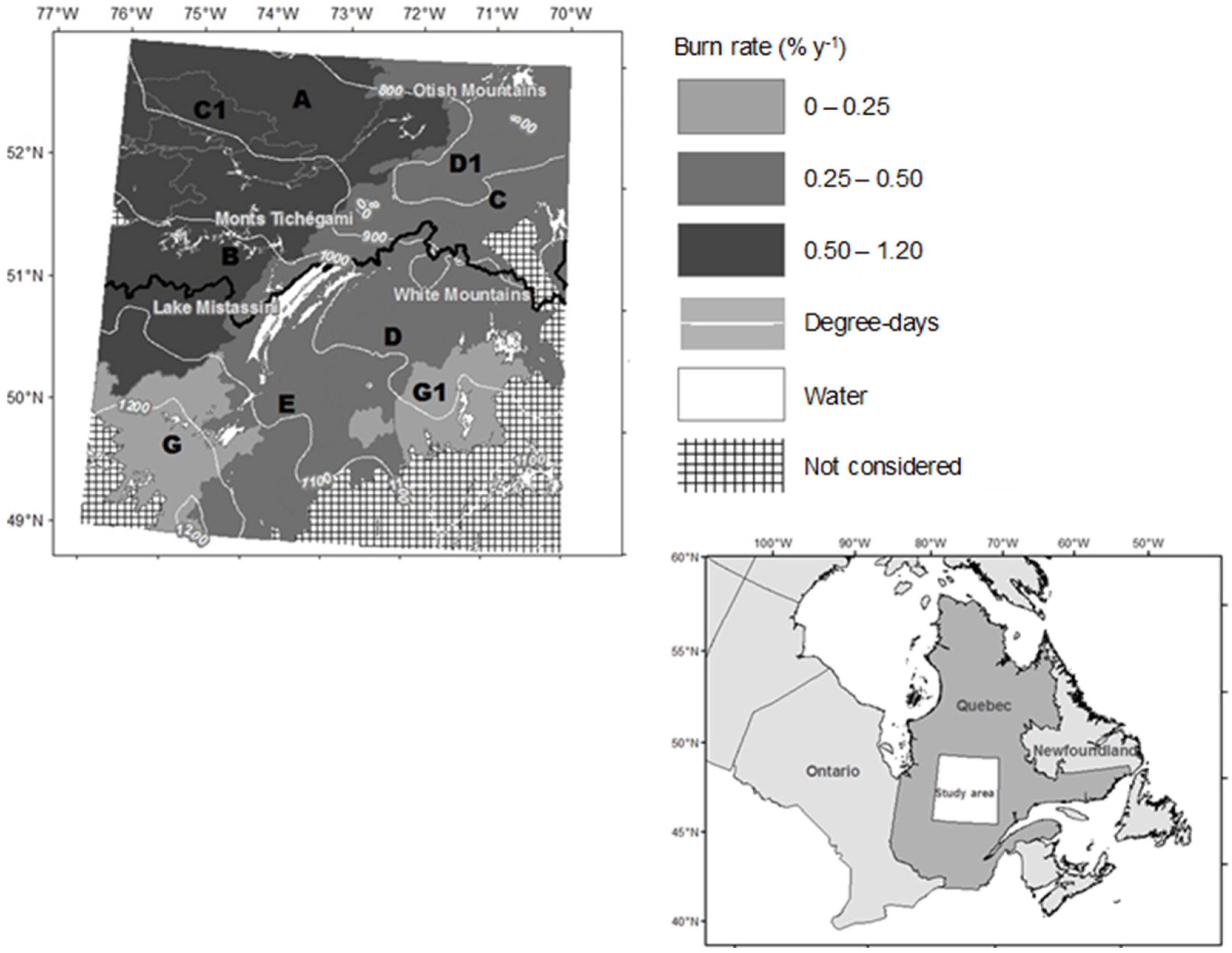
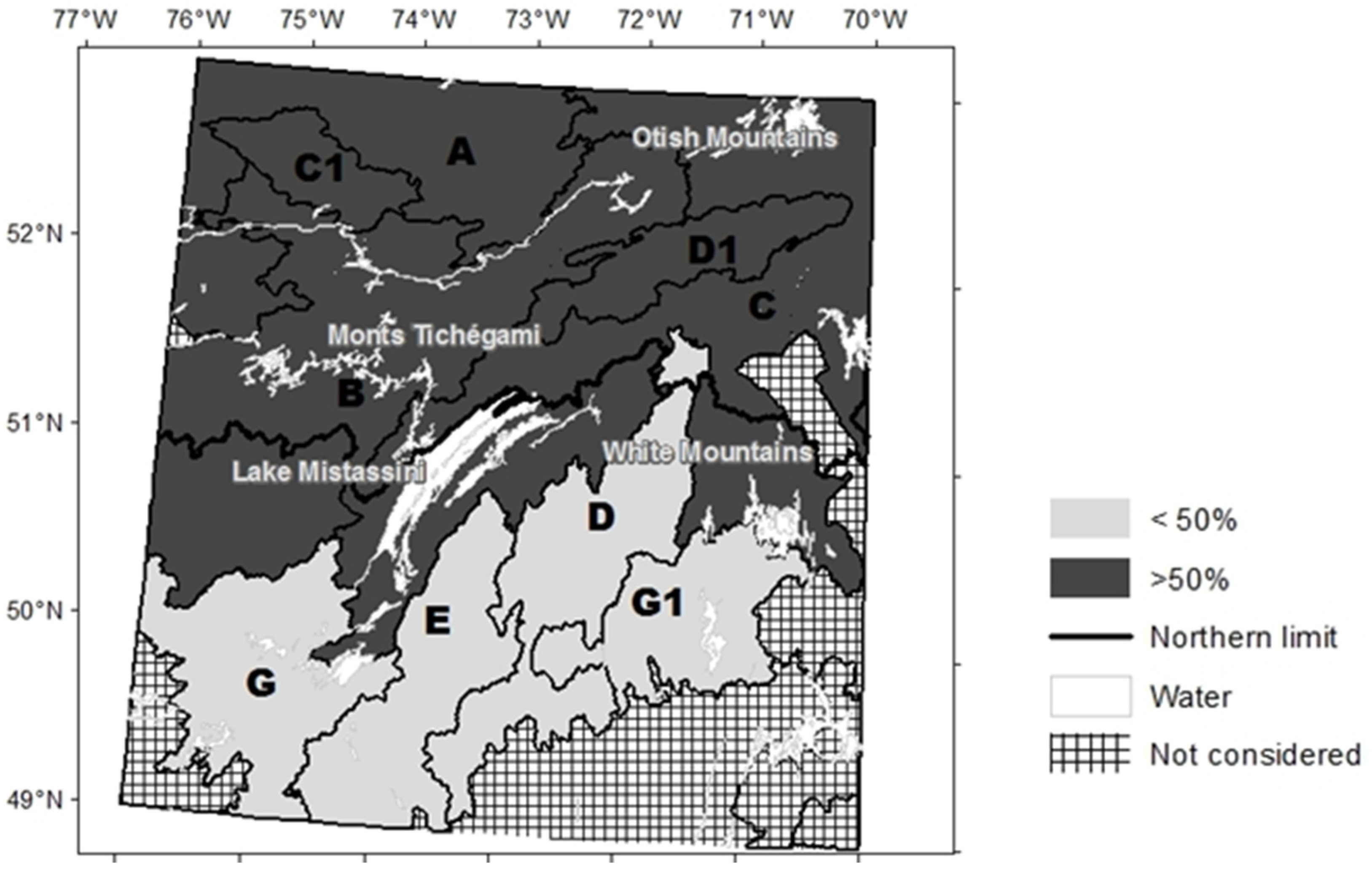
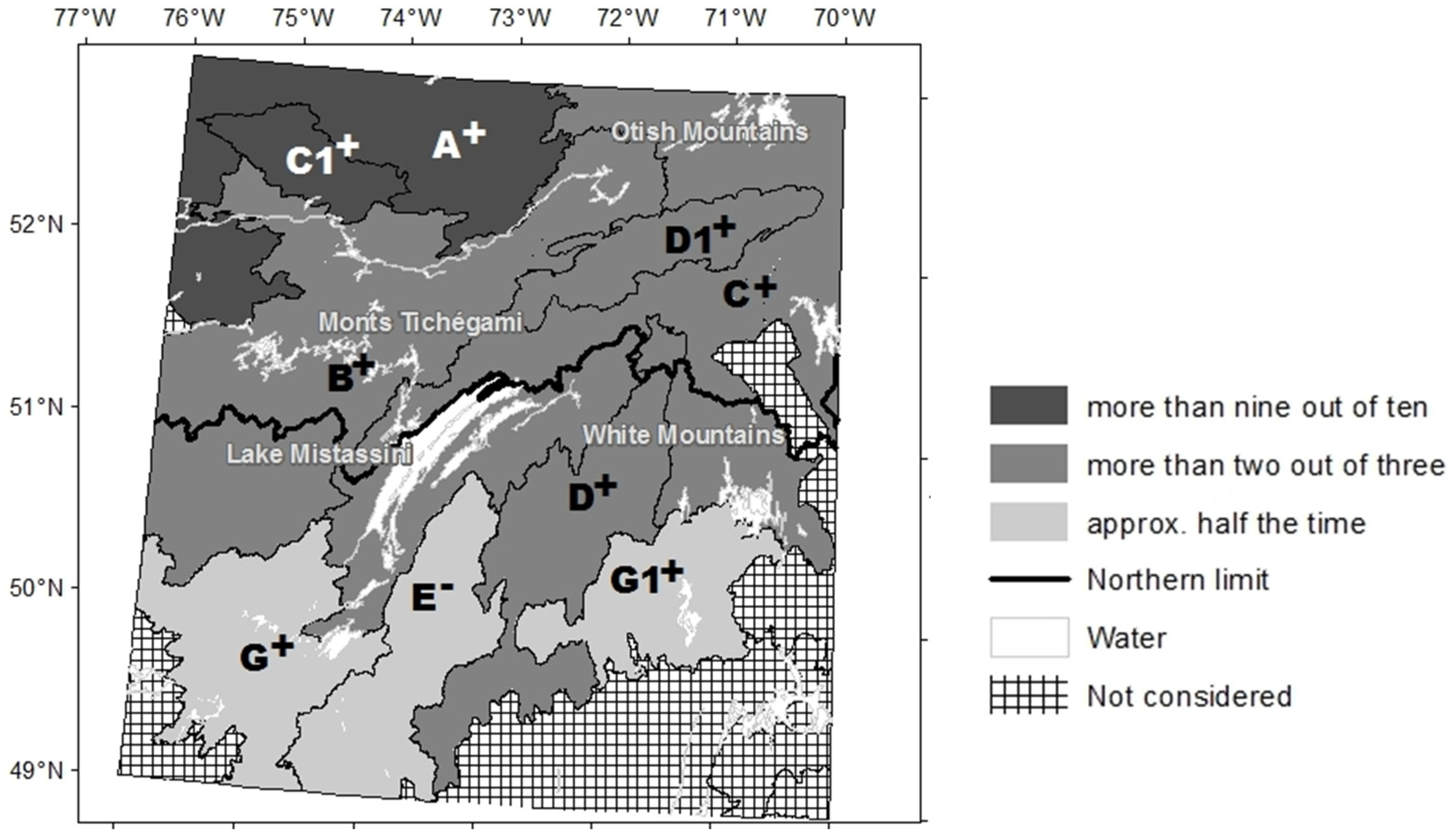
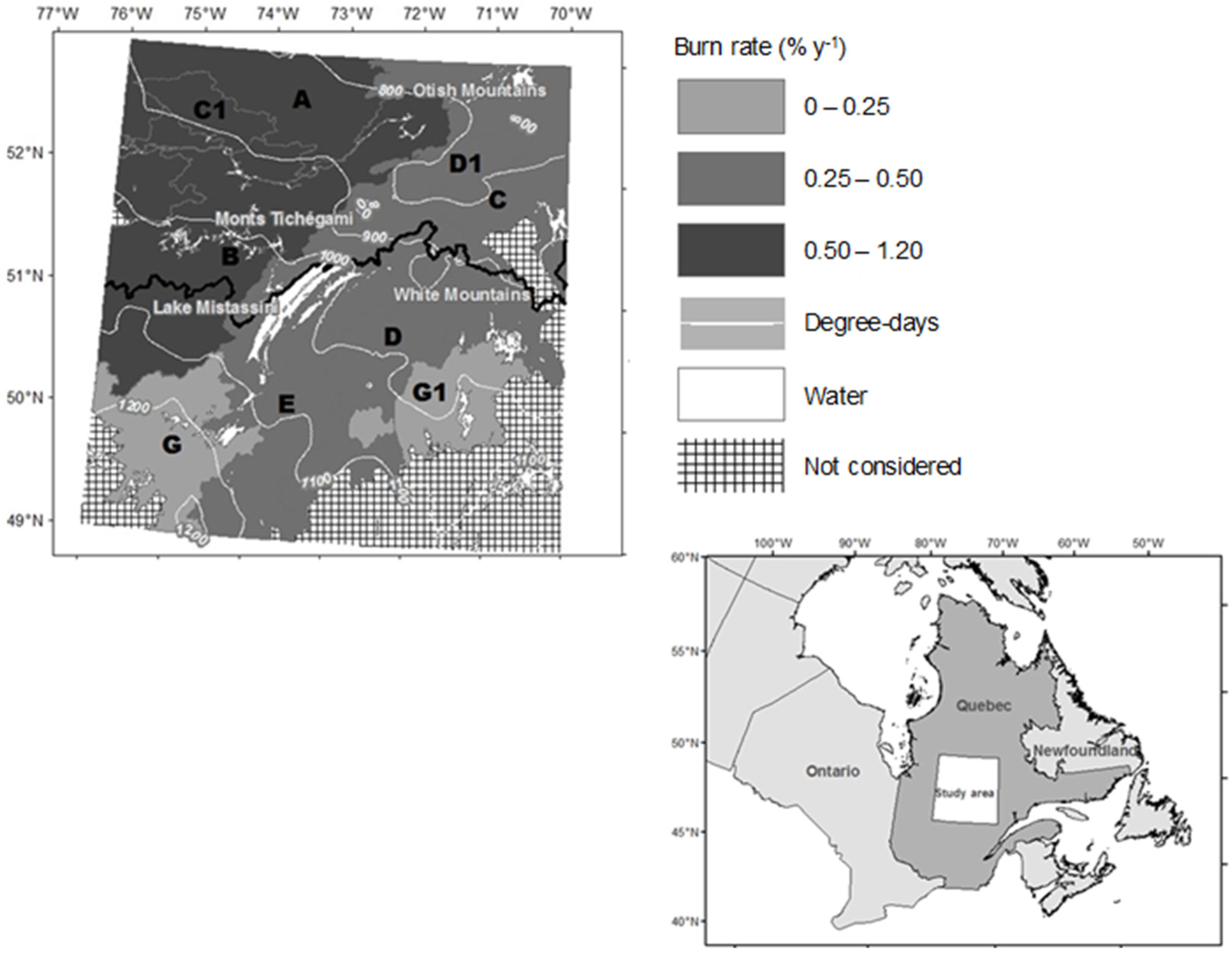
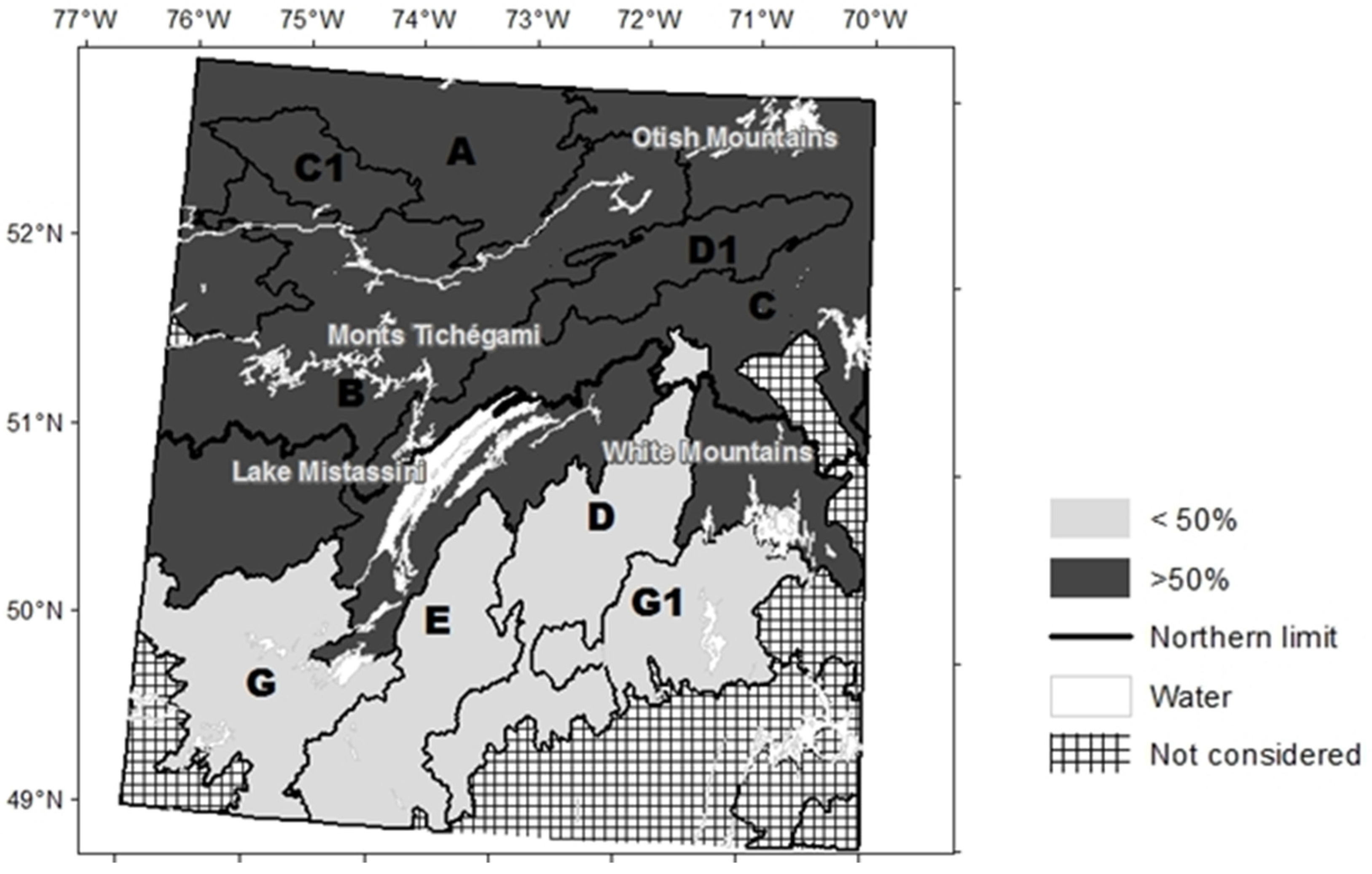
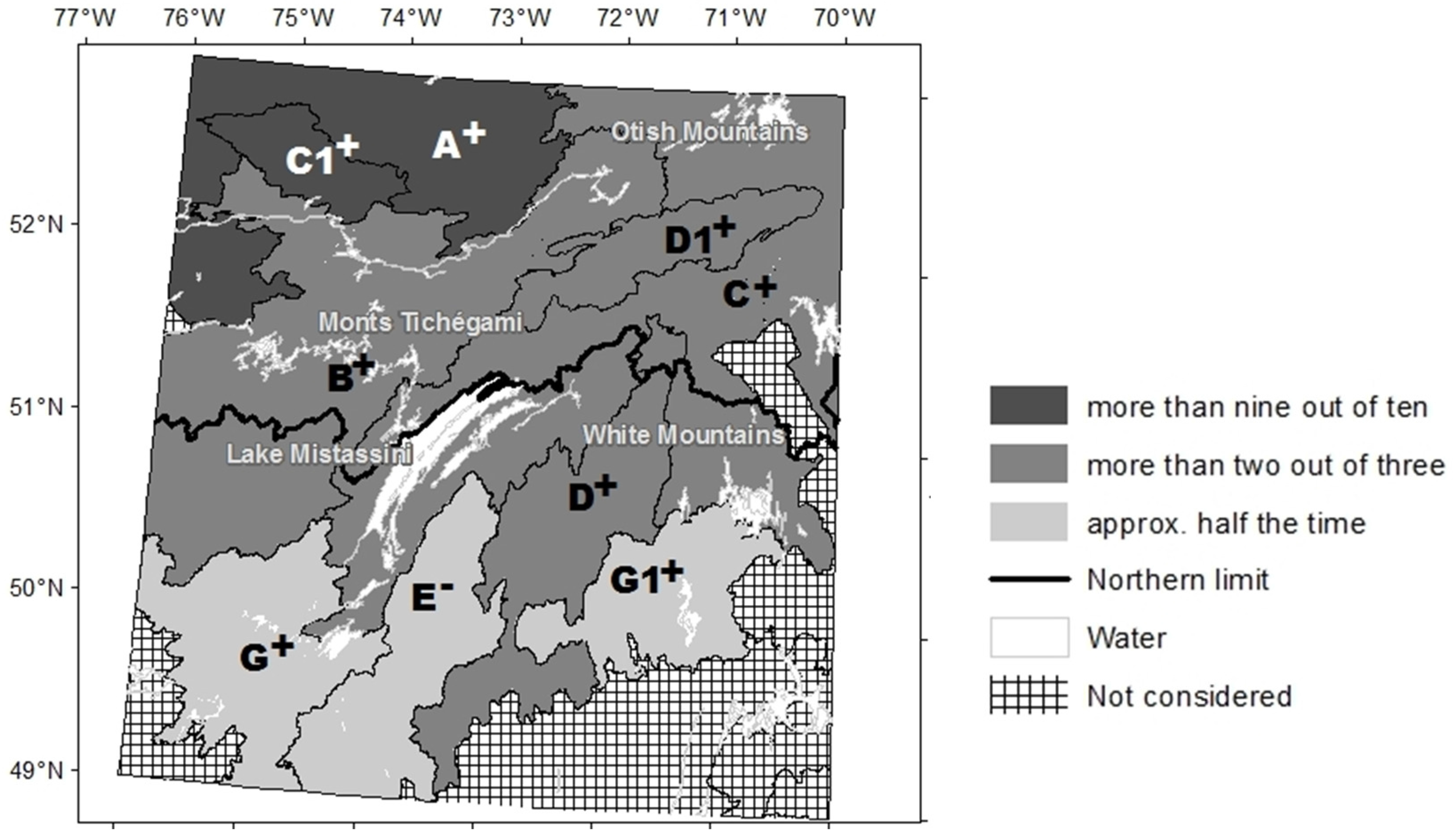
2.1. Study Area
2.2. Forest Data
2.3. Data on Forest Fires
2.4. Description of the Landscape Dynamics Model
2.4.1. Landscape Description in VLM
2.4.2. Succession Submodel
2.4.3. Fire Submodel
2.4.4. Simulation Runs
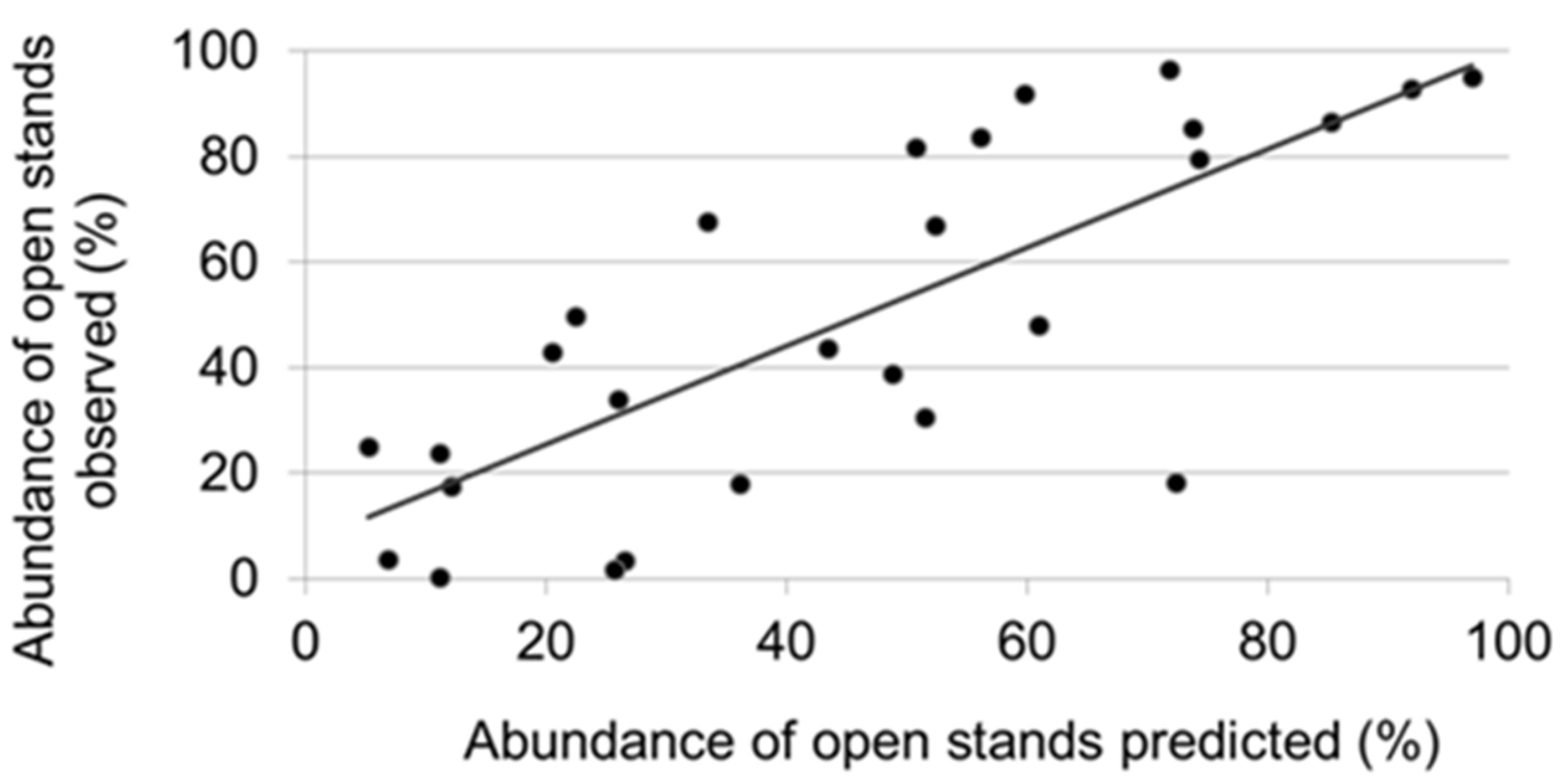
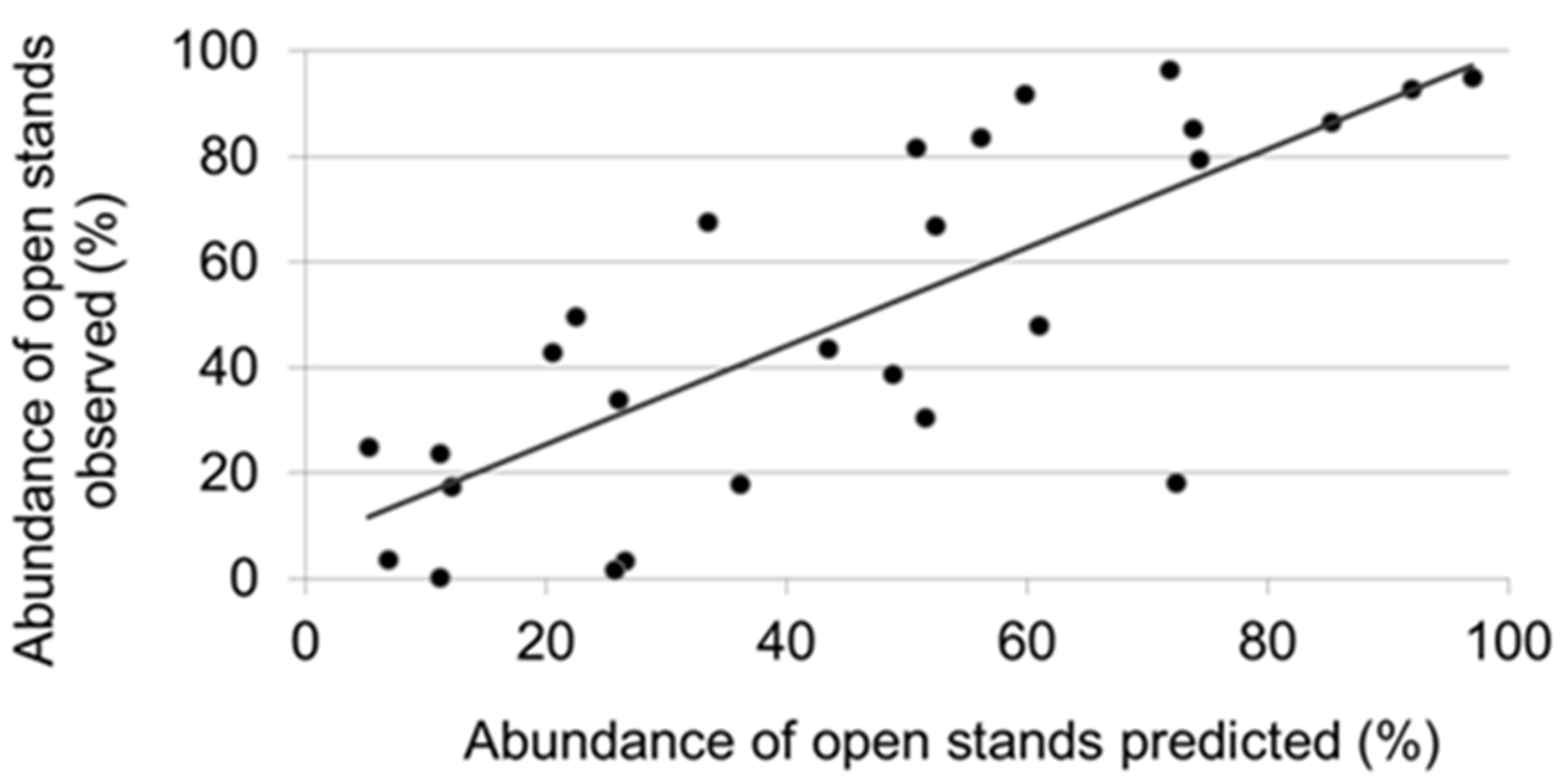
2.5. Interpretation of Simulation Results
3. Results
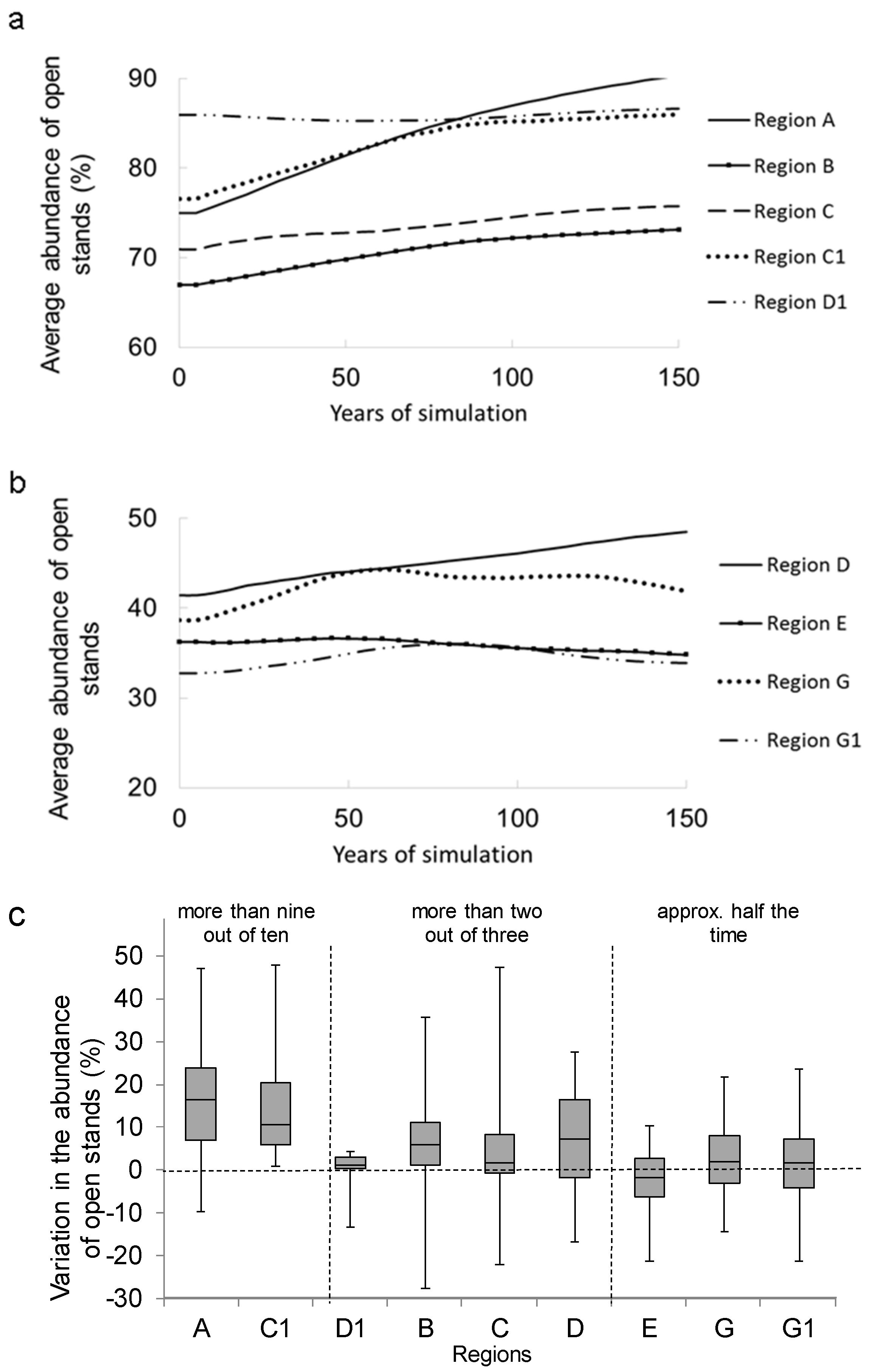
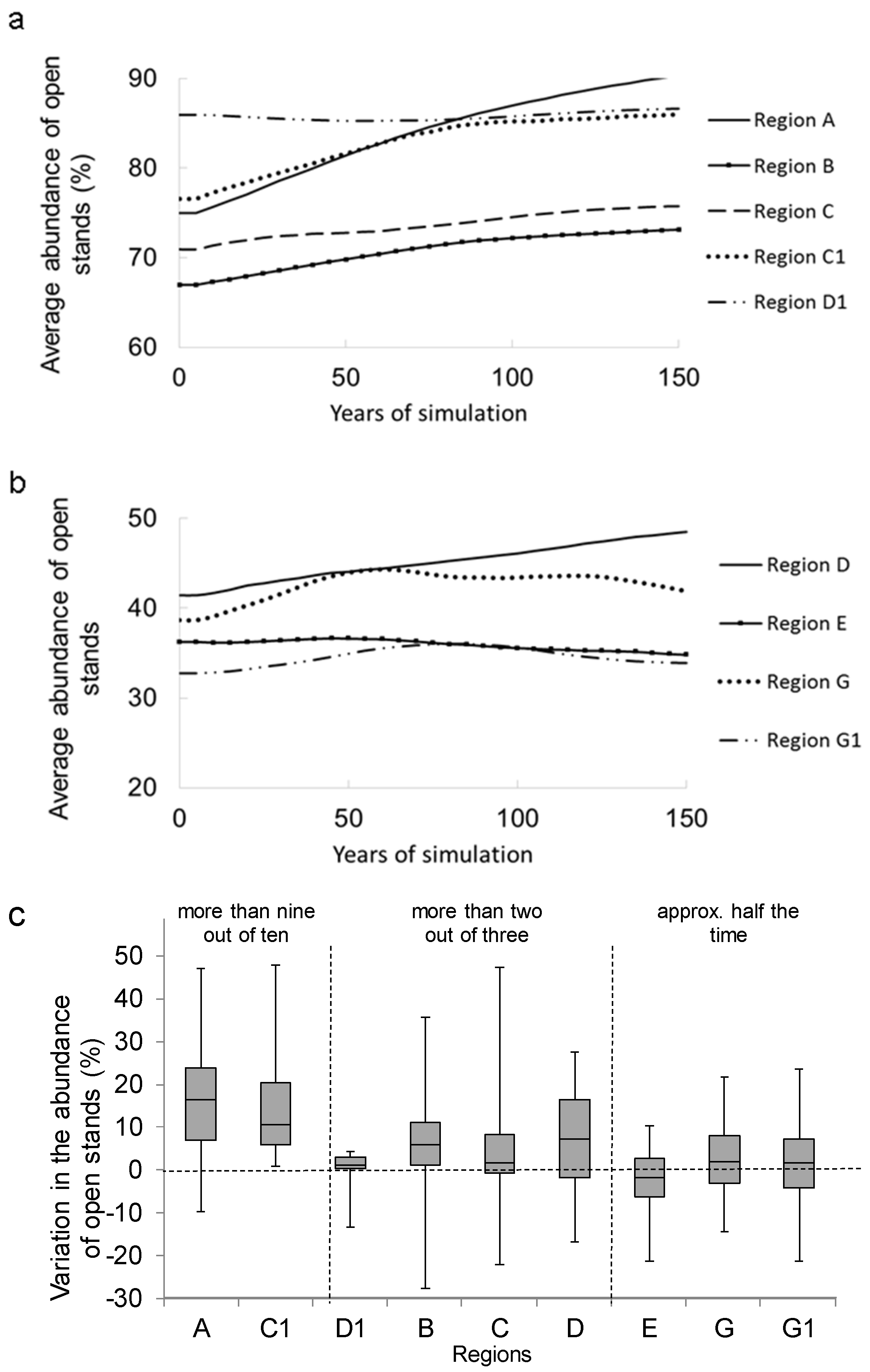
3.1. Temporal Variation in the Abundance of Open Stands
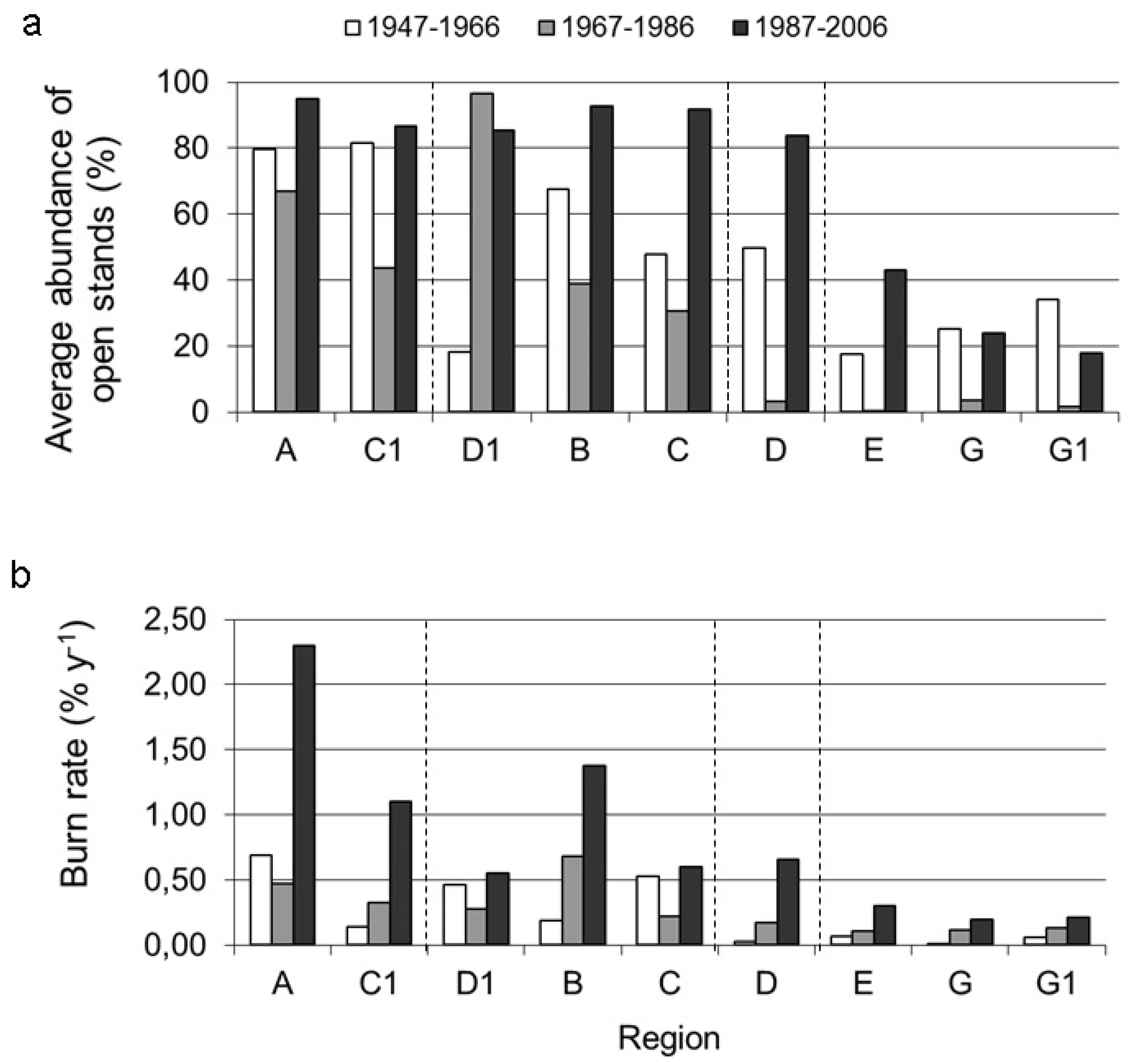
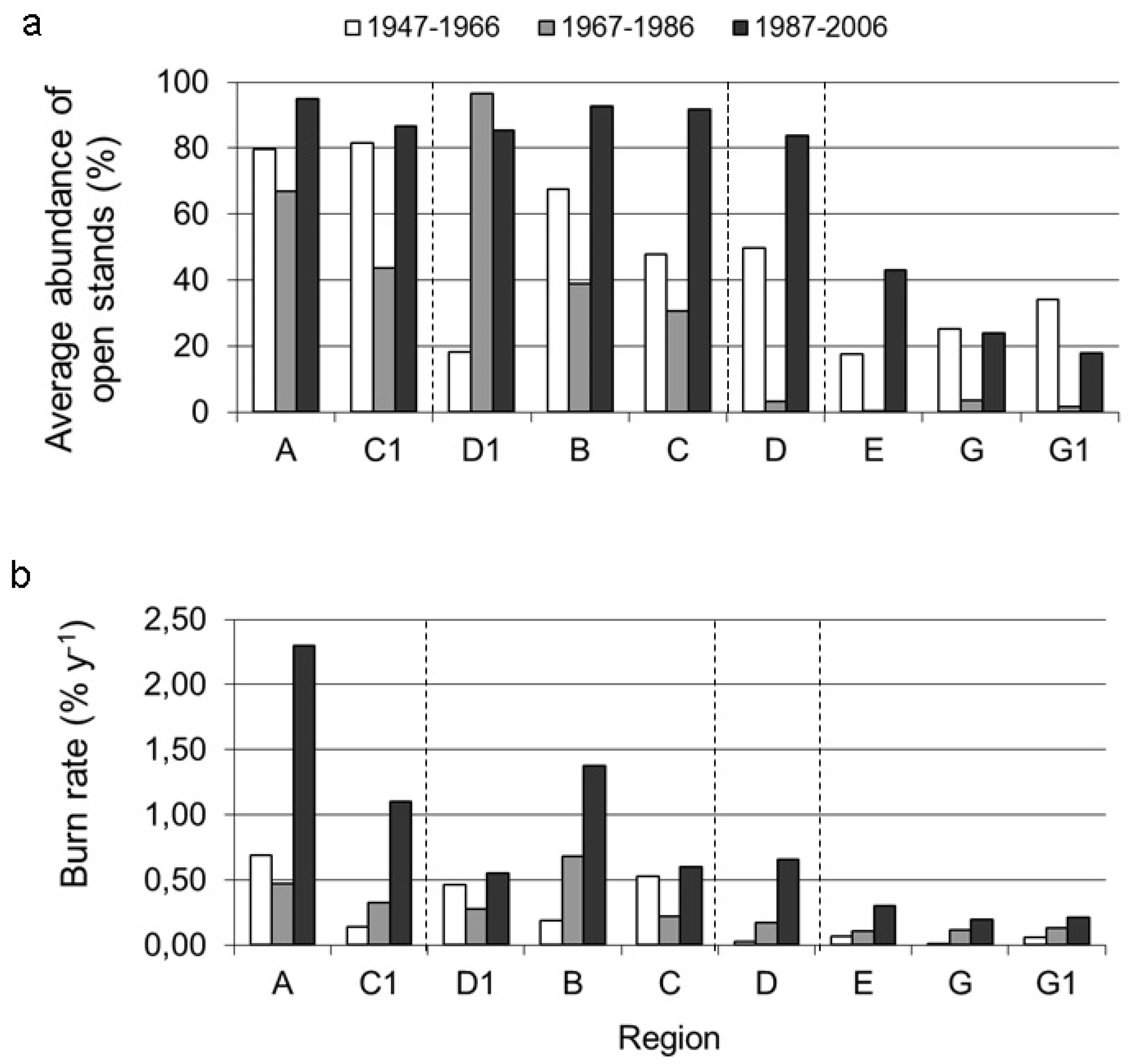
3.2. Long-Term Change in Abundance of Open Stands
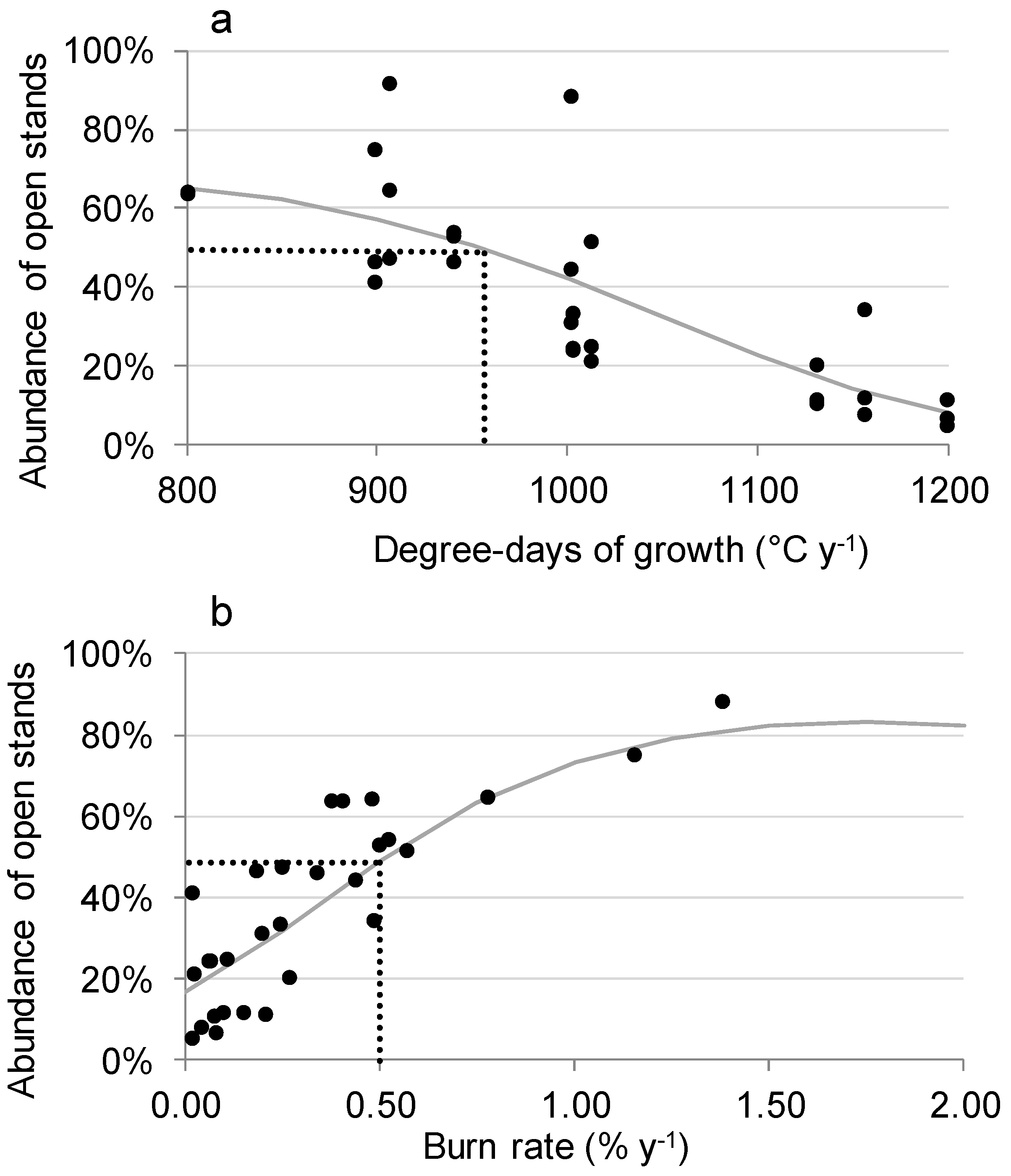
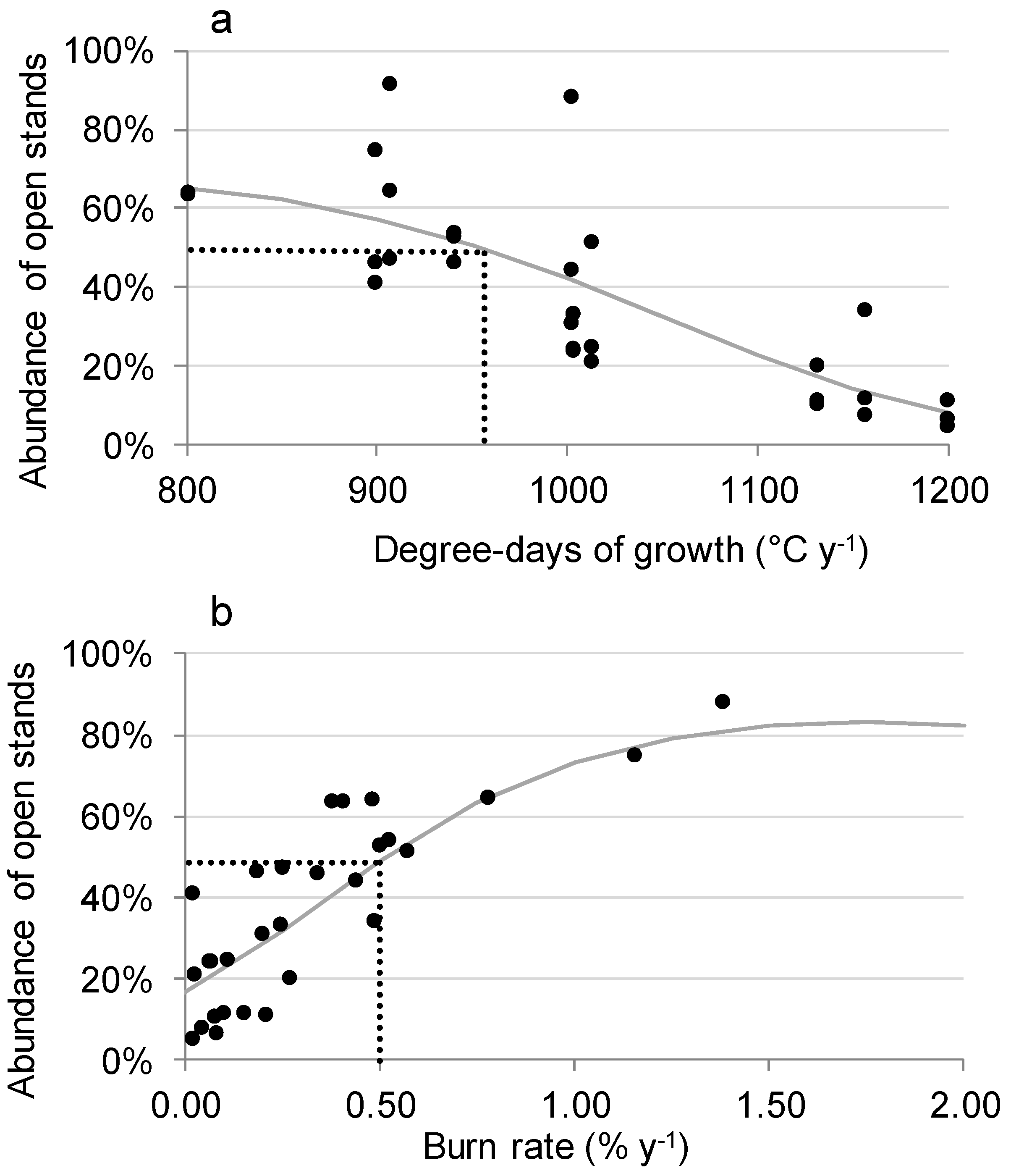
3.3. Factors Responsible for the Variation in the Abundance of Open Stands
4. Discussion
4.1. Correlation of Fire Frequency with the Abundance of Open Stands
4.2. Natural Dynamics of the Spruce-Moss Forest and Lichen Woodland
4.3. Vulnerability and Adaptation to Disturbance Regimes
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pollock, S.L.; Payette, S. Stability in the patterns of long-term development and growth of the Canadian spruce-moss forest. J. Biogeogr. 2010, 37, 1684–1697. [Google Scholar] [CrossRef]
- Payette, S.; Bhiry, N.; Delwaide, A.; Simard, M. Origin of the lichen woodland at its southern range limit in eastern Canada: The catastrophic impact of insect defoliators and fire on the spruce-moss forest. Can. J. For. Res. 2000, 30, 288–305. [Google Scholar] [CrossRef]
- Ministère des Ressources Naturelles et de la Faune. Programme d’Inventaire Écoforestier Nordique. Norme de Cartographie Écoforestière. Direction des Inventaires Forestiers; Gouvernement du Québec: Québec, QC, Canada, 2008.
- Payette, S.; Delwaide, A. Shift of conifer boreal forest lichen-heath parkland caused by successive stand disturbances. Ecosystems 2003, 6, 540–550. [Google Scholar] [CrossRef]
- Simard, M.; Lecomte, N.; Bergeron, Y.; Bernier, P.Y.; Paré, D. Forest productivity decline caused by successional paludification of boreal soils. Ecol. Appl. 2007, 17, 1619–1637. [Google Scholar] [CrossRef] [PubMed]
- Girard, F.; Payette, S.; Gagnon, R. Rapid expansion of lichen woodlands within the closed-crown boreal forest zone over the last 50 years caused by stand disturbances in eastern Canada. J. Biogeogr. 2008, 35, 529–537. [Google Scholar] [CrossRef]
- Scheffer, M.; Hirota, M.; Holmgren, M.; Nes, E.H.V.; Chapin, F.S. Thresholds for boreal biome transitions. Proc. Natl. Acad. Sci. USA 2012, 109, 21384–21389. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.G.; Romme, W.H.; Gardner, R.H.; O’Neill, R.V.; Kratz, T.K. A revised concept of landscape equilibrium: Disturbance and stability on scaled landscapes. Landsc. Ecol. 1993, 3, 213–227. [Google Scholar] [CrossRef]
- Robitaille, A.; Saucier, J.P. Paysages Régionaux du Québec Méridional. Ministère des Ressources Naturelles, Gouvernement du Québec; Les publications du Québec: Québec, QC, Canada, 1998. [Google Scholar]
- Bergeron, Y.; Dansereau, P.R. Fire history in the southern boreal forest of north-western Quebec. Can. J. For. Res. 1993, 23, 25–32. [Google Scholar]
- Le Goff, H.; Sirois, L. Black spruce and jack pine dynamics simulated under varying fire cycles in the northern boreal forest of Quebec, Canada. Can. J. For. Res. 2004, 34, 2399–2409. [Google Scholar] [CrossRef]
- Lavoie, L.; Sirois, L. Vegetation changes caused by recent fires in the northern boreal forest of eastern Canada. J. Veg. Sci. 1998, 9, 483–492. [Google Scholar] [CrossRef]
- Boucher, D.; Gauthier, S.; De Grandpré, L. Structural changes in coniferous stands along a chronosequence and a productivity gradient in the northeastern boreal forest of Québec. Ecoscience 2006, 13, 172–180. [Google Scholar] [CrossRef]
- Rapanoela, R.; Raulier, F.; Gauthier, S.; Ouzennou, H.; Saucier, J.P.; Bergeron, Y. Contrasting current and potential productivity and the influence of fire and species composition in the boreal forest: A case study in eastern Canada. Can. J. For. Res. 2015, 45, 541–552. [Google Scholar] [CrossRef]
- Van Wagner, C.E. Age-class distribution and the forest fire cycle. Can. J. For. Res. 1978, 8, 220–227. [Google Scholar] [CrossRef]
- Gauthier, S.; Leduc, A.; Bergeron, Y.; Le Goff, H. La fréquence des feux et l’aménagement forestier inspiré des perturbations naturelles. In Aménagement Écosystémique en Forêt Boréale; Gauthier, S., Vaillancourt, M.-A., Leduc, A., de Granpré, L., Kneeshaw, D., Morin, H., Drapeau, P., Bergeron, Y., Eds.; Presses de l’Université du Québec: Montréal, QC, Canada, 2008; pp. 61–78. [Google Scholar]
- Cyr, D.; Gauthier, S.; Bergeron, Y.; Carcaillet, C. Forest management is driving the eastern North American boreal forest outside its natural range of variability. Front. Ecol. Environ. 2009, 7, 519–524. [Google Scholar] [CrossRef]
- Liang, J.; Picard, N. Matrix model of forest dynamics: An overview and outlook. For. Sci. 2013, 59, 359–378. [Google Scholar] [CrossRef]
- Landres, P.; Morgan, P.; Swanson, F. Overview of the use of natural variability concepts in managing ecological systems. Ecol. Appl. 1999, 9, 1179–1188. [Google Scholar]
- Johnson, E.A.; Gutsell, S.L. Fire frequency models, methods and interpretations. Adv. Ecol. Res. 1994, 25, 239–287. [Google Scholar]
- Bergeron, Y.; Gauthier, S.; Kafka, V.; Lefort, P.; Lesieur, D. Natural fire frequency for the eastern Canadian boreal forest: Consequences for sustainable forestry. Can. J. For. Res. 2001, 31, 384–391. [Google Scholar] [CrossRef]
- Bergeron, Y.; Flannigan, M.; Gauthier, S.; Leduc, A.; Lefort, P. Past, current and future fire frequency in the Canadian boreal forest: Implications for sustainable forest management. Ambio 2004, 33, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, Y.; Gauthier, S.; Flannigan, M.; Kafka, V. Fire regimes at the transition between mixedwood and coniferous boreal forest in northwestern Quebec. Ecology 2004, 85, 1916–1932. [Google Scholar] [CrossRef]
- Stocks, B.J.; Mason, J.A.; Todd, J.B.; Bosch, E.M.; Wotton, B.M.; Amiro, B.D.; Flannigan, M.D.; Hirsch, K.G.; Logan, K.A.; Martell, D.L.; et al. Large forest fires in Canada, 1959–1997. J. Geophys. Res. 2003, 108D1, FFR-5. [Google Scholar] [CrossRef]
- Mansuy, N.; Gauthier, S.; Robitaille, A.; Bergeron, Y. The effects of surficial deposit-drainage combinations on spatial variations of fire cycles in the boreal forest of eastern Canada. Int. J. Wildland Fire 2010, 19, 1083–1098. [Google Scholar] [CrossRef]
- Metsaranta, J.M.; Kurz, W.A. Inter-annual variability of ecosystem production in boreal jack pine forests (1975–2004) estimated from tree-ring data using CBM-CFS3. Ecol. Model. 2012, 224, 111–123. [Google Scholar] [CrossRef]
- Mouillot, F.; Field, C.B. Fire history and the global carbon budget: A 1 × 1 fire history reconstruction for the 20th century. Glob. Chang. Biol. 2005, 11, 398–420. [Google Scholar] [CrossRef]
- Kasischke, E.S.; Turetsky, M.R. Recent changes in the fire regime across the North American boreal region—Spatial and temporal patterns of burning across Canada and Alaska. Geophys. Res. Lett. 2006, 33, L09703. [Google Scholar] [CrossRef]
- Morris, W.; Doak, D.; Groom, M.; Kareiva, P.; Fieberg, J.; Gerber, L.; Murphy, P.; Thomson, D. A Practical Handbook for Population Viability Analysis; The Nature Conservancy: Arlington, VA, USA, 1999. [Google Scholar]
- Buongiorno, J.; Gilless, J.K. Decision Methods for Forest Resource Management; Academic Press: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Zhou, M.; Buongiorno, J. Forest landscape management in a stochastic environment, with an application to mixed loblolly pine-hardwood forest. For. Ecol. Manag. 2006, 223, 170–182. [Google Scholar] [CrossRef]
- Caswell, H. Matrix Population Models: Construction, Analysis and Interpretation; Sinauer: Sunderland, MA, USA, 1989. [Google Scholar]
- Yemshanov, D.G.; Perera, A.H. A spatially explicit stochastic model to simulate boreal forest cover transitions: General structure and properties. Ecol. Model. 2002, 150, 189–209. [Google Scholar] [CrossRef]
- Régnière, J.; Saint-Amant, R. BioSIM 9—Manuel de l’Utilisateur; Rapport d’information LAU-X-134F; Ressources Naturelles Canada, Service Canadien des Forêts, Centre de Foresterie des Laurentides: Québec, QC, Canada, 2008. Available online: ftp://ftp.cfl.scf.rncan.gc.ca/regniere/software/BioSIM/ (accessed on 11 December 2013).
- Ministère des Ressources Naturelles et de la Faune (MRNFQ). Limite Nordique des Forêts Attribuables; Rapport Final du Comité; Ministère des Ressources Naturelles et de la Faune: Québec, QC, Canada, 2000. Available online: http://www.mrn.gouv.qc.ca/publications/forets/consultation/partie1.pdf (accessed on 12 November 2013).
- Létourneau, J.P.; Matajek, S.; Morneau, C.; Robitaille, A.; Roméo, T.; Brunelle, J.; Leboeuf, A. Norme de Cartographie Écoforestière du Programme d’Inventaire Écoforestier Nordique; Ministère des Ressources Naturelles et de la Faune: Québec, QC, Canada, 2008.
- Gauthier, S.; Raulier, F.; Ouzennou, H.; Saucier, J.-P. Strategic analysis of forest vulnerability to risk related to fire: An example from the coniferous boreal forest of Quebec. Can. J. For. Res. 2015, 45, 553–565. [Google Scholar] [CrossRef]
- Fall, A.; Fortin, M.-J.; Kneeshaw, D.D.; Yamasaki, S.H.; Messier, C.; Bouthillier, L.; Smyth, C. Consequences of various landscape-scale ecosystem management strategies and fire cycles on age-class structure and harvest in boreal forests. Can. J. For. Res. 2004, 34, 310–322. [Google Scholar] [CrossRef] [Green Version]
- Didion, M.; Fortin, M.-J.; Fall, A. Forest age structure as indicator of boreal forest sustainability under alternative management and fire regimes: A landscape level sensitivity analysis. Ecol. Model. 2007, 200, 45–58. [Google Scholar] [CrossRef]
- James, P.M.A.; Fortin, M.-J.; Sturtevant, B.R.; Fall, A.; Kneeshaw, D.D. Modelling spatial interactions among fire, spruce budworm, and logging in the boreal forest. Ecosystems 2011, 14, 60–75. [Google Scholar] [CrossRef]
- Tittler, R.; Messier, C.; Fall, A. Concentrating anthropogenic disturbance to balance ecological and economic values: Applications to forest management. Ecol. Appl. 2012, 22, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Fall, A.; Fall, J. A domain-specific language for models of landscape dynamics. Ecol. Model. 2001, 141, 1–18. [Google Scholar] [CrossRef]
- Raulier, F.; Dhital, N.; Racine, P.; Tittler, R.; Fall, A. Increasing resilience of timber supply: How a variable buffer stock of timber can efficiently reduce exposure to shortfalls caused by wildfires. For. Pol. Econ. 2014, 46, 47–55. [Google Scholar] [CrossRef]
- McRoberts, R.E.; Nelson, M.D.; Wendt, D.G. Stratified estimation of forest area using satellite imagery, inventory data, and the k-nearest neighbors technique. Remote Sens. Environ. 2002, 82, 457–468. [Google Scholar] [CrossRef]
- McRoberts, R.E.; Tomppo, E.O.; Finley, A.O. Heikkinen, J. Estimating areal means and variances of forest attributes using the k-Nearest Neighbors technique and satellite imagery. Remote Sens. Environ. 2007, 111, 466–480. [Google Scholar] [CrossRef]
- Mansuy, N.; Gauthier, S.; Robitaille, A.; Bergeron, Y. Regional patterns of post fire canopy recovery in the northern boreal forest of Quebec: Interactions between surficial deposit, climate, and fire cycle. Can. J. For. Res. 2012, 42, 1328–1343. [Google Scholar] [CrossRef]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–871. [Google Scholar] [CrossRef]
- Tuominen, S.; Fish, S.; Poso, S. Combining remote sensing, data from earlier inventories, and geostatistical interpolation in multisource forest inventory. Can. J. For. Res. 2003, 33, 624–634. [Google Scholar] [CrossRef]
- Dunne, T.; Leopold, L.B. Water in Environmental Planning, 1st ed.; W.H. Freeman & Company: New York, NY, USA, 1978. [Google Scholar]
- Sutherland, G.D.; Eng, M.; Fall, S.A. Effects of uncertainties about stand-replacing natural disturbances on forest-management projections. J. Ecosyst. Manag. 2004, 4, 1–18. Available online: http://www.forrex.org/jem/2004/vol4/no2/art5.pdf (accessed on 9 May 2016). [Google Scholar]
- Hatcher, R.J. A Study of Black Spruce Forests in Northern Quebec; Department of Forests: Ottawa, ON, Canada, 1963.
- Bouchard, M.; Pothier, D.; Gauthier, S. Fire return intervals and tree species succession in the North Shore region of eastern Quebec. Can. J. For. Res. 2008, 38, 1621–1633. [Google Scholar] [CrossRef]
- Confais, J.; le Guen, M. Premiers pas en Régression Linéaire avec SAS; Revue MODULAD; SAS Institute Inc.: Cary, NC, USA, 2006. [Google Scholar]
- Chapin, F.S., III; Callaghan, T.V.; Bergeron, Y.; Fukuda, M.; Johnstone, J.F.; Juday, G.; Zimov, S.A. Global change and the boreal forest: Thresholds, shifting states or gradual changes. Ambio 2004, 33, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Sirois, L.; Bonan, G.B.; Shugart, H.H. Development of a simulation model of forest-tundra transition zone of northeastern Canada. Can. J. For. Res. 1994, 24, 697–706. [Google Scholar] [CrossRef]
- Price, D.T.; Alfaro, R.I.; Brown, K.J.; Flannigan, M.D.; Fleming, R.A.; Hogg, E.H.; Girardin, M.P.; Lakusta, T.; Johnston, M.; McKenney, D.W.; et al. Anticipating the consequences of climate change for Canada’s boreal forest ecosystems. Environ. Rev. 2013, 21, 322–365. [Google Scholar] [CrossRef]
- Johnson, E.A.; Van Wagner, C.E. The theory and use of two fire history models. Can. J. For. Res. 1985, 15, 214–220. [Google Scholar] [CrossRef]
- Metsaranta, J.M. Potentially limited detectability of short-term changes in boreal fire regimes: A simulation study. Int. J. Wildland Fire 2010, 19, 1140–1146. [Google Scholar] [CrossRef]
- Li, C. Estimation of fire frequency and fire cycle: A computational perspective. Ecol. Model. 2002, 154, 103–120. [Google Scholar] [CrossRef]
- Ter-Mikaelian, M.T.; Colombo, S.J.; Chen, J. Estimating natural forest fire return interval in northeastern Ontario, Canada. For. Ecol. Manag. 2009, 258, 2037–2045. [Google Scholar] [CrossRef]
- Boulanger, Y.; Gauthier, S.; Burton, P.J. A refinement of models projecting future Canadian fire regimes using homogeneous fire regime zones. Can. J. For. Res. 2014, 44, 365–376. [Google Scholar] [CrossRef]
- Le Goff, H.; Flannigan, M.D.; Bergeron, Y.; Girardin, M.P. Historical fire regime shifts related to climate teleconnections in the Waswanipi area, central Quebec, Canada. Int. J. Wildland Fire 2007, 16, 607–618. [Google Scholar] [CrossRef]
- Sirois, L. Impact of fire on Picea mariana and Pinus banksiana seedlings in subarctic lichen woodlands. J. Veg. Sci. 1993, 4, 795–802. [Google Scholar] [CrossRef]
- Barrett, K.; Kasischke, E.S.; McGuire, A.D.; Turetsky, M.R.; Kane, E.S. Modeling fire severity in black spruce stands in the Alaskan boreal forest using spectral and non-spectral geospatial data. Remote Sens. Environ. 2010, 114, 1494–1503. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Kasischke, E.S. Stand-level effects of soil burn severity on postfire regeneration in a recently burned black spruce forest. Can. J. For. Res. 2005, 35, 2151–2163. [Google Scholar] [CrossRef]
- Lecomte, N.; Simard, M.; Bergeron, Y. Effects of fire severity and initial tree composition on stand structural development in the coniferous boreal forest of northwestern Québec, Canada. Ecoscience 2006, 13, 152–163. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Chapin, F.S. Effects of soil burn severity on post-fire tree recruitment in boreal forest. Ecosystems 2006, 9, 14–31. [Google Scholar] [CrossRef]
- Jin, Y.; Randerson, J.T.; Goetz, S.J.; Beck, P.S.A.; Loranty, M.M.; Goulden, M.L. The influence of burn severity on post-fire vegetation recovery and albedo change during early succession in North American boreal forests. J. Geophys. Res. 2012, 117, G01036. [Google Scholar] [CrossRef]
- Johnson, E.A.; Miyanishi, K. Testing the assumptions of chronosequences in succession. Ecol. Lett. 2008, 11, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.R.; Chen, H.Y.H. Multiple successional pathways of boreal forest stands in central Canada. Ecography 2011, 34, 208–219. [Google Scholar] [CrossRef]
- Frelich, L.E.; Reich, P.B. Neighborhood effects, disturbance severity, and community stability in forests. Ecosystems 1999, 2, 151–166. [Google Scholar] [CrossRef]
- Miller, J.D.; Skinner, C.N.; Safford, H.D.; Knapp, E.E.; Ramirez, C.M. Trends and causes of severity, size, and number of fires in northwestern California, USA. Ecol. Appl. 2012, 22, 184–203. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, S.; Bernier, P.; Kuuluvainen, T.; Shvidenko, A.Z.; Schepaschenko, D.G. Boreal forest health and global change. Science 2015, 349, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Rowe, J.S. Forest Regions of Canada. Bulletin 123; Government of Canada, Department of Northern Affairs and National Resources, Forestry Branch, Headquarters: Ottawa, ON, Canada, 1959.
- Hare, F.K.; Ritchie, J.C. The boreal Bioclimates. Geogr. Rev. 1972, 62, 333–65. [Google Scholar] [CrossRef]
- Heinselman, M.L. The natural role of fire in northern conifer forests. In Proceedings of the Fire in the Northern Environment: A symposium, Fairbanks, AK, USA, 13–14 April 1971; Slaughter, C.W., Barney, R.J., Hansen, G.M., Eds.; USDA, Forest Service, Pacific Northwest Research Station: Portland, OR, USA; pp. 61–72.
- Payette, S. Fire as a controlling process in the North American boreal forest. In Systems Analysis of the Global Boreal Forest; Shugart, H.H., Leemans, R., Bonan, G.B., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 144–169. [Google Scholar]
- Jasinski, J.P.P.; Payette, S. The creation of alternative stable states in the southern boreal forest, Québec, Canada. Ecol. Monogr. 2005, 75, 561–583. [Google Scholar] [CrossRef]
- Beisner, B.E.; Haydon, D.T.; Cuddington, K. Alternative stable states in ecology. Front. Ecol. Environ. 2003, 1, 376–382. [Google Scholar] [CrossRef]
- Gagnon, R.; Morin, H. Les forêts d'épinette noire au Québec: Dynamique, perturbations et biodiversité. Nat. Can. 2001, 125, 26–35. [Google Scholar]
- Holling, C.S. Resilience and stability of ecological systems. Ann. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Groot, A.; Horton, B.J. Age and size structure of natural and second-growth peatland Picea mariana stands. Can. J. For. Res. 1994, 24, 225–233. [Google Scholar] [CrossRef]
- Garet, J.; Raulier, F.; Pothier, D.; Cumming, S.G. Forest age class structures as indicators of sustainability in boreal forest: Are we measuring them correctly? Ecol. Indic. 2012, 23, 202–210. [Google Scholar] [CrossRef]
- Van Bogaert, R.K.; Gauthier, S.; Raulier, F.; Saucier, J.P.; Boucher, D.; Robitaille, A.; Bergeron, Y. Exploring forest productivity at an early age after fire: A case study at the northern limit of commercial forests in Quebec. Can. J. For. Res. 2015, 45, 579–593. [Google Scholar] [CrossRef]
- Irulappa Pillai Vijayakumar, D.B.; Raulier, F.; Bernier, P.; Paré, D.; Gauthier, S.; Bergeron, Y.; Pothier, D. Cover density recovery after fire disturbance controls landscape aboveground biomass carbon in the boreal forest of eastern Canada. For. Ecol. Manag. 2016, 360, 170–180. [Google Scholar] [CrossRef]
- Girard, F.; Payette, S.; Gagnon, R. Origin of the lichen-spruce woodland in the closed-crown forest zone of eastern Canada. Glob. Ecol. Biogeogr. 2009, 18, 291–303. [Google Scholar] [CrossRef]
- Hogg, E.H.; Bernier, P.Y. Climate change impacts on drought-prone forests in western Canada. For. Chron. 2005, 81, 675–682. [Google Scholar] [CrossRef]
- Gauthier, S.; Bernier, P.; Burton, P.J.; Edwards, J.; Isaac, K.; Isabel, N.; Jayen, K.; Le Goff, H.; Nelson, E.A. Climate change vulnerability and adaptation in the managed Canadian boreal forest. Environ. Rev. 2014, 22, 256–285. [Google Scholar] [CrossRef]
- Imbeau, L.; St-Laurent, M.-H.; Marzell, L.; Brodeur, V. Current capacity to conduct ecologically sustainable forest management in northeastern Canada reveals challenges for conservation of biodiversity. Can. J. For. Res. 2015, 45, 567–578. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Area (km2) | Degree Day (°C·year−1) | Burn Rate (%·year−1) [25] | Abundance of Open Stands (Median) in 2006 (%) |
|---|---|---|---|---|
| A | 18,525 | 900 | 1.11 | 89 |
| B | 35,157 | 1000 | 0.78 | 88 |
| C1 | 4265 | 900 | 0.67 | 54 |
| C | 42,468 | 940 | 0.48 | 85 |
| D1 | 6492 | 800 | 0.42 | 81 |
| D | 14,291 | 1010 | 0.37 | 93 |
| E | 11,522 | 1130 | 0.28 | 44 |
| G1 | 15,043 | 1000 | 0.16 | 34 |
| G | 13,509 | 1200 | 0.14 | 28 |
| Abundance of Open Stands | Probability of Increase of Open Stands | ||||
|---|---|---|---|---|---|
| Variables | Estimate (SE) | Wald χ2 | Variables | Estimate (SE) | Wald χ2 |
| Degree day | 0.0078 (0.0007) | 106.7 | Last frost day | 0.0595 (0.0109) | 29.8 |
| Degree day × burn rate | −1.058 (0.217) | 23.9 | Total precipitation | −0.00886 (0.0019) | 22.0 |
| Burn rate | 794 (207) | 14.7 | Burn rate | 1.248 (0.269) | 21.6 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rapanoela, R.; Raulier, F.; Gauthier, S. Regional Instability in the Abundance of Open Stands in the Boreal Forest of Eastern Canada. Forests 2016, 7, 103. https://doi.org/10.3390/f7050103
Rapanoela R, Raulier F, Gauthier S. Regional Instability in the Abundance of Open Stands in the Boreal Forest of Eastern Canada. Forests. 2016; 7(5):103. https://doi.org/10.3390/f7050103
Chicago/Turabian StyleRapanoela, Rija, Frédéric Raulier, and Sylvie Gauthier. 2016. "Regional Instability in the Abundance of Open Stands in the Boreal Forest of Eastern Canada" Forests 7, no. 5: 103. https://doi.org/10.3390/f7050103