1. Introduction
Pitch canker disease, caused by
Fusarium circinatum (F. circinatum) Nirenberg and O’Donnell, is an emergent disease that is threatening
Pinus trees in Europe. Since it was first recognized in Southeastern USA [
1], the disease has been reported in other parts of the world [
2]. In many of these reports, the disease appeared first in forest nurseries, like in South Africa [
3], Chile [
4], Spain [
5], Portugal [
6] and Brazil [
7]. In trees,
F. circinatum causes branch dieback and resinous cankers that progressively girdle the wood causing branch death. Canker infection of multiple branches of the main stem can cause extensive dieback in the tree crown and tree mortality.
Pine seeds are considered one of the major pathways of introduction of
F. circinatum into new countries, as it occurred in Chile [
8], South Africa [
9], and Spain [
10]. Pine seed is also one of the potential inoculum sources of
F. circinatum in nurseries [
2,
11,
12]. This fungus is a seed-borne pathogen in
Pinus radiata [
11]; it has also been detected in seeds of
P. taeda [
13] and
P. elliottii [
14].
Fusarium circinatum may infest seeds superficially in cones growing on healthy branches [
11]. The fungus can also infect seeds internally, where it causes damage to embryo and gametophyte tissues [
13]. The mechanisms by which
F. circinatum infects seed are unknown, but three types of infection have been identified [
11]: superficial, active internal and dormant internal, though the latter is not detectable on plate medium. The extent of internal and/or external seed contamination depends on the pine species and environmental conditions [
2].
Fusarium circinatum was isolated from up to 83% of seeds of
P. radiata [
11]. Anderson
et al. [
15] determined that 34% of
P. elliottii infested seed was internal.
In forest nurseries,
F. circinatum causes pre- and post-emergence damping-off of seedlings [
11] and mortality of established seedlings [
3]. Main symptoms are collar rot and wilting [
2], but under certain conditions, seedlings can be asymptomatic and may develop the disease after they are transplanted to plantations [
2,
11]. From an epidemiological point of view, host growth stage is an important parameter influencing disease intensity that should be taken into account for disease management [
16]. Viljoen
et al. [
3] found that mortality of established seedlings tends to be lower than that of newly germinated seedlings, and Storer
et al. [
11] determined that seedling emergence from infested Monterey pine seed was 9% compared with 67% for non-infested seeds. Another important epidemiological parameter is the infection rate [
16], for which there are no available data, comparing infection of newly germinated seedlings and established seedlings.
Little is known about this pitch canker disease in forest nurseries in Spain.
Fusarium circinatum may be detected in any of the usual species found in these nurseries (
P. pinaster,
P. sylvestris,
P. nigra and
P. radiata) [
5,
17], although disease has only been recorded in plantations of
P. radiata [
18,
19] and recently, in
P. pinaster [
19]. Disease outbreaks in nurseries are sporadic, which suggests an external source of inoculum. On the other hand, routine seed health tests performed following the specific method for the detection of
F. circinatum on
Pinus spp. as recommended by ISTA [
20] reveal that number of seeds with presence of the pathogen is almost null. This standard procedure is based on placing seeds on semi-selective media, and it does not distinguish internal from external presence of
F. circinatum. Moreover, the presence of the fungus does not indicate necessarily that it has been transmitted from tree to seed, since fungus could have contaminated seeds during commercial processes of harvesting, cleaning and storage. The objectives of this work are to (1) determine the frequency of seeds with presence of
F. circinatum collected from
P. radiata trees, and evaluate the importance of seed contamination during its storage; (2) study persistence of the pathogen in seedlings, to demonstrate transmission of seed borne inoculum to the seedling; (3) study the effect of seed inoculum location and dose on infection and development of different host growth stages, including germination; and (4) perform a temporal analysis of the disease with different doses of inoculum. These evaluations will be performed under optimal conditions of temperature [
21] on
P. radiata, one of the most susceptible species [
18,
22], to determine the maximum disease infection rate.
2. Experimental Section
2.1. Natural Presence of Fusarium circinatum in/on Seeds of Symptomatic Trees
In eight plantations of
P. radiata located in Northern Spain (Pais Vasco), cones on damaged stems and branches were collected in 2009 from each of 10 trees with symptoms of pitch canker disease. Disease was visually evaluated by the number of cankers and percentage of defoliation. Cones from each tree were pooled and then 100 seeds were obtained by exposing cones to a temperature of 35 °C–40 °C for 24 h to cause them to open. Seeds (after hand-removing their wings) were dissected into separate fractions consisting of seed coat, gametophyte and embryo, following the protocol described by Aitken and Iturritxa [
23] and then cultivated separately in FSM (Fusarium Selective Medium) [
24]. Presence of
Fusarium circinatum was evaluated after incubation for 14 day at 20 °C ± 2 °C.
Logistic regression was used to analyze the relation between the proportion of seeds with the pathogen and the number of cankers per tree (as a measure of severity disease). All statistical analyses were conducted using SAS, Version 9.3 (SAS Institute Inc., Cary, NC, USA).
2.2. Seed Disinfestation and Inoculation with Fusarium circinatum—Determination of Inoculum Dose on/in the Seed
Pathogen free-seed of P. radiata from New Zealand was used both in seed contamination and artificial inoculation assays. Seeds were surface disinfested by immersion and shaking in: (1) 10% commercial bleach plus 0.1% Tween 20 for 15 min; (2) purified water (RIOs Type 3 RO, Millipore, Madrid, Spain) for 5 min, twice; (3) hydrogen peroxide 33% for 10 min; (4) sterile distilled water for 10 min, three times; and (5) dried in an air flow cabinet for 2 h.
Fusarium circinatum (CECT20759) was isolated from a
P. radiata seedling in Guipuzkoa, Spain. This isolate was identified to be in vegetative compatibility group (VCG) A and Mating type 2 (Mat-2), the only mating type present in the Basque Country [
25]. This isolate is stored on small pieces of sterile filter paper (Whatman no.1) at −20 °C. For inoculation purposes, it was cultured for 10 days at 22 °C on Potato Dextrose Agar (PDA) (Pronadisa, Madrid, Spain). The inoculum suspension was prepared by adding 10 mL of sterile purified water on the surface of a PDA plate where
F. circinatum has grown for about 10 days at 22 °C, scraping with a spatula, and then filtering with glass wool washed (Panreac Química, Barcelona, Spain). The spore concentration was determined using a haemocytometer and the suspension diluted to the desired concentration with sterile purified water. Inoculation was done by immersion and shaking at 150 rpm for 5 min in a spore suspension of
F. circinatum. Seeds inoculated with water were used as control. Inoculated seeds were dried for 2 h in an air flow cabinet.
The viability of the inoculum was determined by plating dilution series onto PDA plates. The inoculum actually present in the seed after inoculation was estimated by grinding individually inoculated seeds (at various dose) in 1 mL sterile distilled water and plating on PDA plates. Also, when this estimate was below the detection limit, presence of inoculum was visualized growing the seeds on SFA (Selective Fusarium Agar) [
26] plates.
2.3. Seed Contamination by Fusarium circinatum during Seed Storage
Differences during storage were tested at three conditions: 4 °C and 20% ± 3% RH, 20 °C and 25% ± 5% RH and 20 °C 65% ± 1% RH and at four sampling times. For each condition and sampling time, three plates with 101 surface disinfested P. radiata seeds per plate were used. One of these 101 seeds was inoculated by immersion in a suspension of 103 spore/mL and plated on a 50 mm-diameter Petri dish with the other 100 non-inoculated seeds. Plates were incubated at the three conditions. Every week during four weeks, three plates of each condition were sampled and their seeds were plated on FSM to detect the number of seeds contaminated with F. circinatum.
2.4. In Vitro Effect of Fusarium circinatum on Seed Germination and Germling Growth
To assess the effects of inoculum location and doses on seed germination and germling growth, 50 surface-disinfested seeds of
P. radiata were inoculated by immersion as described in
Section 2.2, at doses of 10
2 or 10
4 spores/mL: (1) protrusion-stage pregerminated seeds on water agar that mimic embryo, gametophyte and external infestation; (2) carborundum rubbed seeds to crack their coat and favor fungal penetration into seeds (imitating gametophyte and external infestation); and (3) no previous seed treatment to imitate external infestation of seed coats. Inoculation with water was done for each respective inoculation method. Each group of 50 seeds were germinated on a tray, covered with plastic bags to maintain high humidity and incubated at 22 °C with 12 h of light. Periodically until 48 days post inoculation (dpi), germlings were evaluated in a scale (0–2) for growth stage, where 0 = no germination, 1 = radicle present, and 2 = radicle and hypocotyl present; and for presence/absence of
F. circinatum mycelia in stages 1 and 2.
Logit analysis was performed to test effects of inoculation method and inoculum dose on germination percentage at 20 dpi. Likewise, proportion of damaged germlings in growth stages 1 and 2 at the end of the experiment was analyzed by logistic regression on inoculation method and inoculum dose and their interaction.
2.5. Temporal Analysis of Pre- and Post- Emergence Damping-Off, and of Death of Established Seedlings
Pinus radiata surface-disinfested seeds were externally inoculated at doses of 104 or 106 spores/mL, stored at room temperature for one day until sowing in forest sowing 35-trays containing substrate composed of 80% peat plus 20% coconut fiber, and incubated at constant temperature 20 °C, and 12 h day/night light. Each treatment was applied to three trays, and trays were arranged in a randomized block design. The same conditions were repeated in a second experiment at the dose of 104 spores/mL.
There was a periodical monitoring and recording of seedling emergence and appearance of symptoms produced by
F. circinatum seedling infection, such as girdling of root collar, wilting and die-back. Dead plants were sampled and some sections were plated on SFA medium to corroborate the presence of the pathogen. Identification of
F. circinatum was confirmed morphologically on SNA [
26] and PDA media based on morphological features in pure culture [
27]. The last day of recording pre-emergence damping-off was at the date of the last emergence event. Post-emergence damping-off was recorded until 281 dpi, considering the first two months after emergence as damping-off and later on, damage of established seedlings. Analysis of variance was used to estimate the effect of inoculum dose on the proportion of pre- and post- emergence damping-off as well as proportion of dead plants at the end of the experiment. Data were transformed prior to the analysis using arc sine of the square root of the proportion. Mean values were compared by Student-Newman-Keuls multiple range tests.
Disease progress for the low inoculum dose (10
4 spores/mL) was regressed using different models (linear, monomolecular, logistic and Gompertz) [
28]. The best model was chosen according to the lowest mean square error value (MSE) and the highest coefficient of determination value (
R2); then, disease infection rate was obtained from the slope parameter of the selected model [
28]. Best fit of data was achieved using the linear model, that is, disease incidence regressed with time (dpi). Two lines were adjusted, one for post-emergence damping-off, and other for damage of established seedlings. The disease infection rate is the average of both replicates.
2.6. Persistence of Fusarium circinatum in Seedlings
As seedlings died from 77–281 dpi in the experiment described above, quantity of F. circinatum was estimated in a sample of them. Colony forming units (cfu) were calculated in five seedlings seed-inoculated with 106 and in 10 seedlings seed-inoculated with 104 spores/mL. Plants were surface disinfested by dipping them in 10% commercial bleach for 15 s, washed 3 times in sterile purified water, cut in pieces and ground in sterile purified water. Dilution series of these extracts were plated on SFA medium.
By the end of the same experiment (at 281 dpi), there were 30 seedlings apparently healthy. In order to further explore symptom development of these seedlings, they were transplanted to pots and grown in a greenhouse at 18 ± 5 °C with relative humidity of 55%–60%. Before this, seven of these seedlings were randomly chosen and processed as described above to estimate cfu of F. circinatum. The remaining 23 seedlings were monitored until 458 dpi. As seedlings died, plant parts (roots, stems and tips) were cut in transversal sections, plated on SFA and checked for presence of F. circinatum. Two plants survived at the end of this period.
3. Results
3.1. Natural Presence of Fusarium circinatum in/on Seeds of Symptomatic Trees
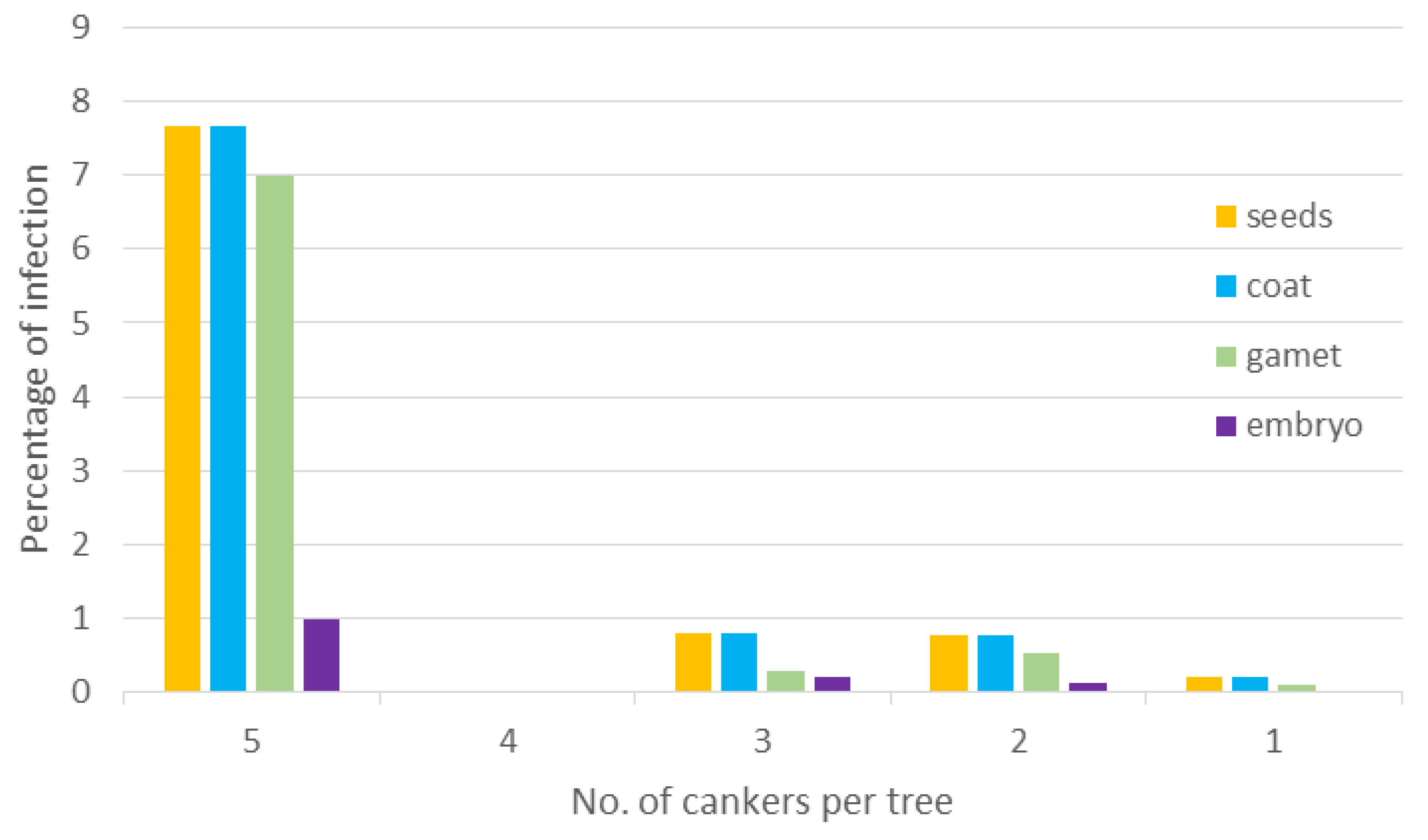
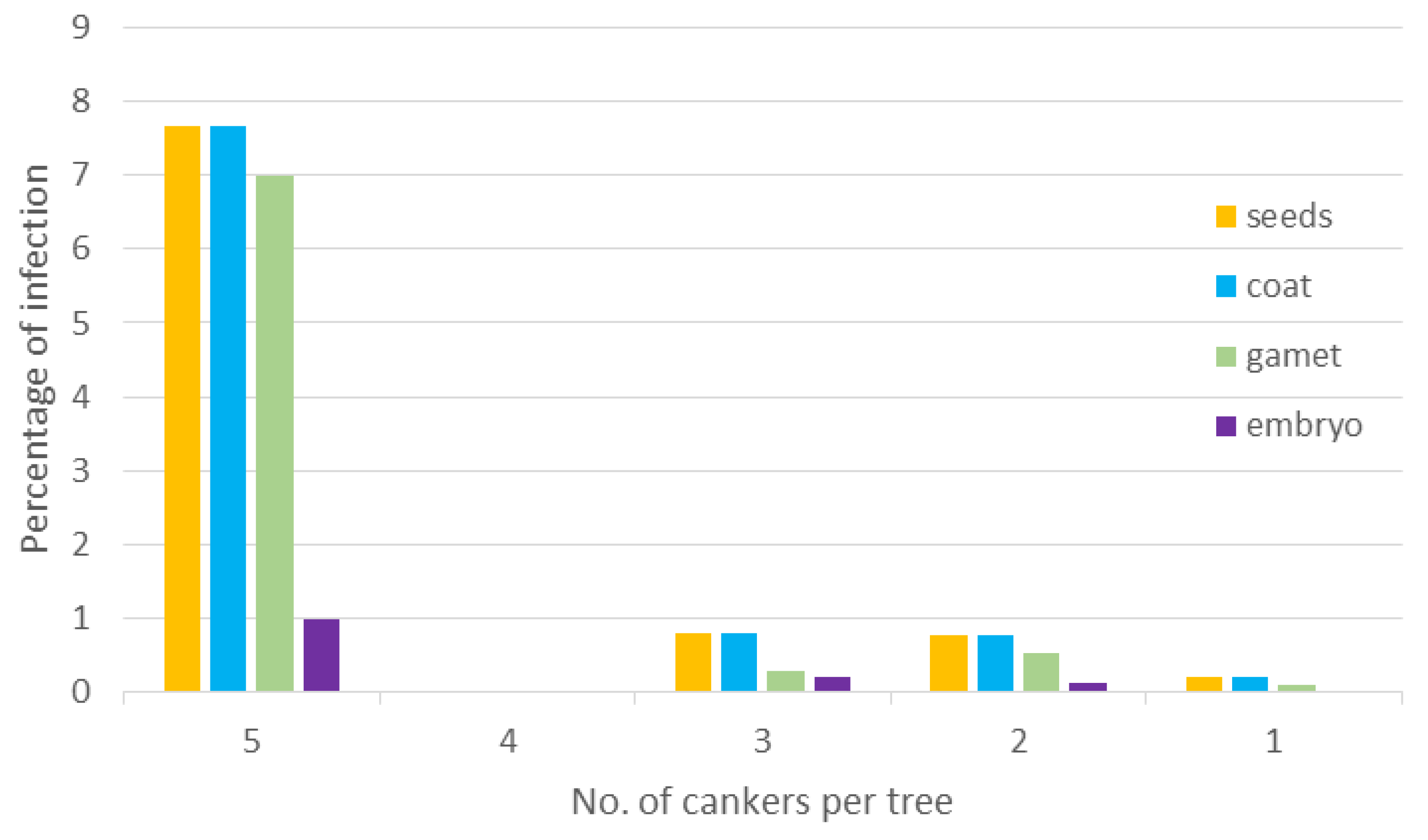
Total number of
P. radiata seeds collected in eight plantations was 7100 from 71 trees. Out of these seeds, less than 1% (52 seeds from 12 trees) were infested with
F. circinatum (mean of infested seeds = 0.73%, standard error ± 0.244%). The pathogen was present on the coat of all infested seeds and in some of them,
F. circinatum was also infecting the gametophyte (mean percentage of infection 0.52 ± 0.219). In a small proportion, the embryo was also infected (0.10 ± 0.05). All seeds with an infected embryo had an infected gametophyte and seed coat as well (
Figure 1).
Figure 1.
Percentage of plantation-collected Pinus radiata (P. radiata) seeds with the presence of F. circinatum and their location on coat, gametophyte and embryo in relation to the tree disease severity (measured by the number of cankers) where seeds were collected.
Figure 1.
Percentage of plantation-collected Pinus radiata (P. radiata) seeds with the presence of F. circinatum and their location on coat, gametophyte and embryo in relation to the tree disease severity (measured by the number of cankers) where seeds were collected.
Logistic regression analysis between percentage of infested seeds and tree disease severity (evaluated as number of cankers in a tree) showed a strong relationship (chi-square for wald test = 101.54, df = 4, p-value < 0.001). Odds ratio (OR) of number of infested seeds from trees with more than 1 canker compared to trees with only 1 canker was calculated from the logit model. According to this analysis, the odds that seeds would be infested was 40 times greater if seeds were collected from trees with five cankers (OR = 40.4, Wald chi-square < 0.001), and almost four times greater if seeds were collected from trees with two and three cankers (respectively, OR = 3.7, Wald chi-square = 0.0033; and OR = 3.9, Wald chi-square = 0.0064).
3.2. Seed Contamination by Fusarium circinatum during Storage
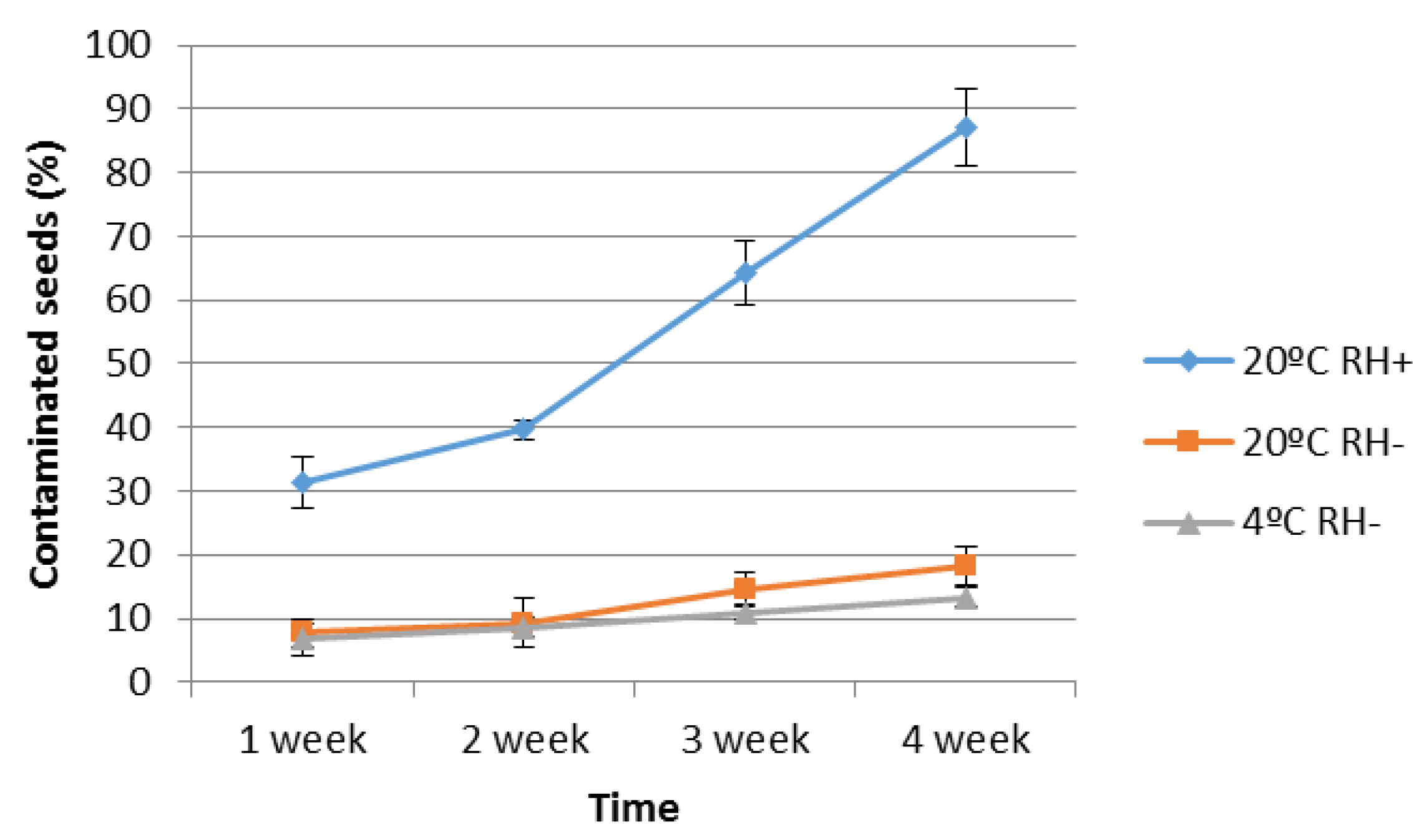
High relative humidity had a great effect on seed contamination at 20 °C, and this was true from the first week (
Figure 2). There were no differences between number of contaminated seeds at 4 °C and 20 °C with low RH (20%). After four weeks, final percentage of contaminated seeds was 87% when incubated at 20 °C and 65% RH, while that value was 18.3% at 20 °C and 13.3% at 4 °C with low RH.
Figure 2.
Percentage of P. radiata seeds contaminated by F. circinatum after incubating 100 disinfested seeds with one superficially inoculated seed. Incubation conditions were 20 °C with 25% or 65% of RH; and 4 °C and 20% RH. Each point is the average of three replicates. Bars are standard deviation.
Figure 2.
Percentage of P. radiata seeds contaminated by F. circinatum after incubating 100 disinfested seeds with one superficially inoculated seed. Incubation conditions were 20 °C with 25% or 65% of RH; and 4 °C and 20% RH. Each point is the average of three replicates. Bars are standard deviation.
3.3. In Vitro Effect of Fusarium circinatum on Seed Germination and Germling Growth
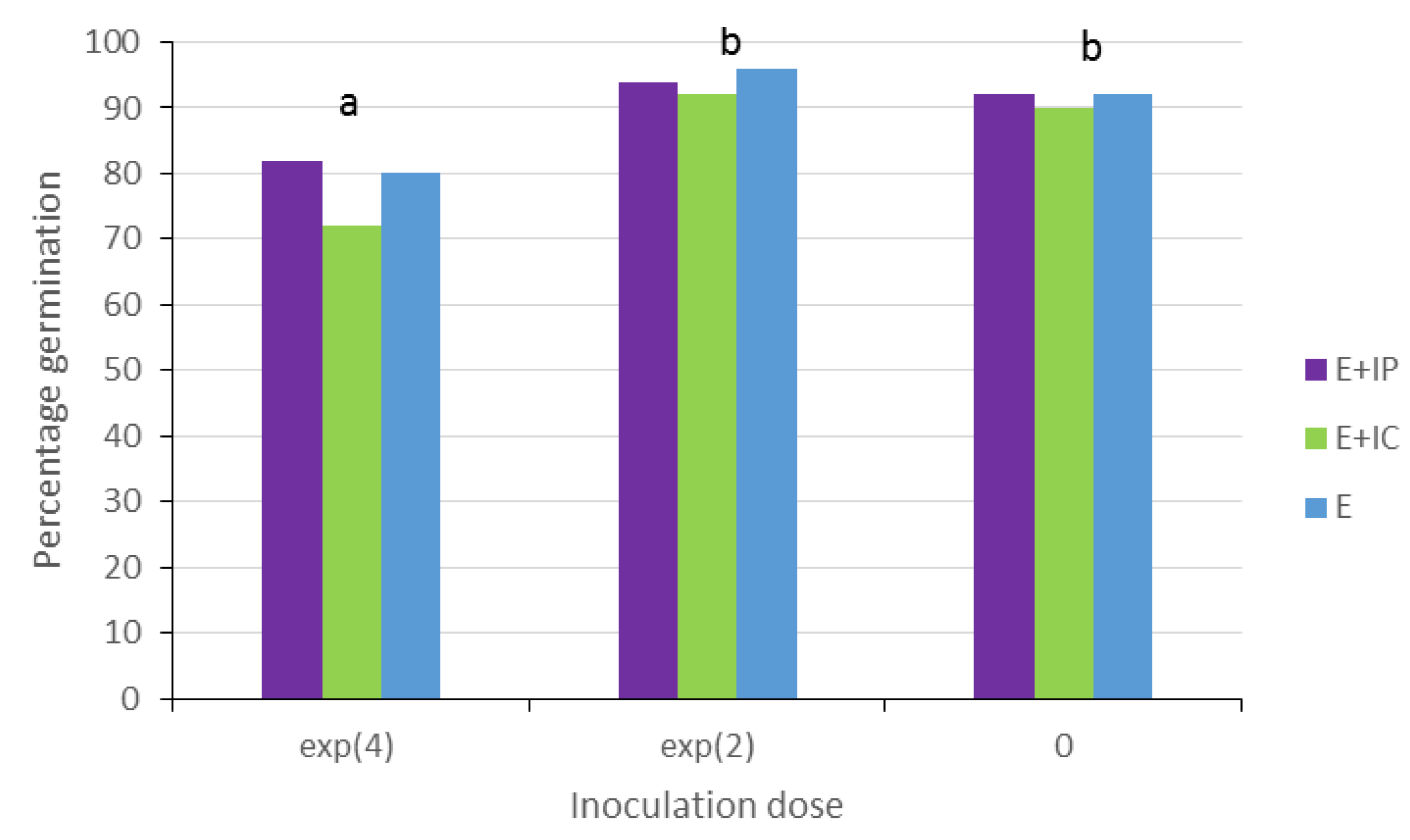
Germination of
P. radiata seeds occurred until 20 dpi. At this date, percentage of total inoculated-seed germination was 86% (
Figure 3). This value depended on inoculum dose (chi-square for wald test = 18.73,
df = 2,
p-value < 0.0001), and not on inoculation method (chi-square for wald test = 2.11,
df = 2,
p-value = 0.349), according to logit regression analysis of both factors. Germination mean was 78% ± 3.74% (standard error) for the dose of 10
4 spores/mL, and 94% ± 1.41% for the dose of 10
2 spores/mL. This germination mean one did not differ significantly from the mean of untreated seeds (91% ± 0.82%) (
Figure 3).
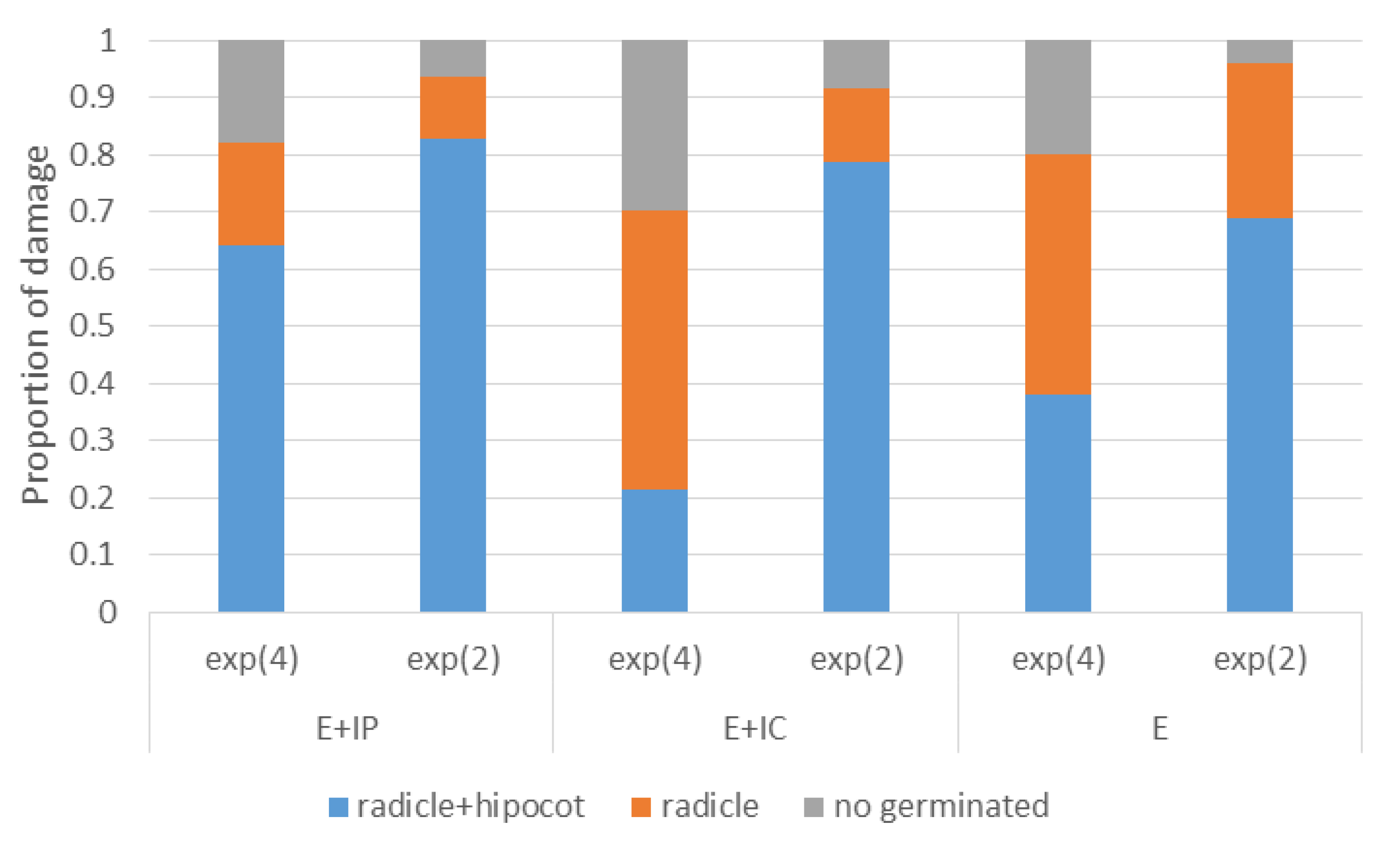
Figure 3.
Total percentage germination of P. radiata seeds at 20 dpi for different inoculation doses and inoculation method: exp (4) = 104 spores/mL; exp (2) = 102 spores/mL; E + IP = internal by immersion of pre-germinated seeds; E + IC = internal by immersion of seeds previously rubbed with carborundum: E = external by immersion. Treatments with different letters differ significantly at P = 0.05. No significant differences were among inoculation methods.
Figure 3.
Total percentage germination of P. radiata seeds at 20 dpi for different inoculation doses and inoculation method: exp (4) = 104 spores/mL; exp (2) = 102 spores/mL; E + IP = internal by immersion of pre-germinated seeds; E + IC = internal by immersion of seeds previously rubbed with carborundum: E = external by immersion. Treatments with different letters differ significantly at P = 0.05. No significant differences were among inoculation methods.
Total seed-inoculated germlings of
P. radiata with only radicle grown (stage 1) at the end of the experiment (48 dpi) was 77 of total 300 seeds (equivalent to 26.3%) (
Table 1). Of these, only two germlings did not show lesions or signs of infection by
F. circinatum (
Table 1). Final proportion of germlings in stage 1 depended on inoculation method (
p-value = 0.0035) and on inoculum dose (
p-value = 0.0001), but not on their interaction (
p-value = 0.3).
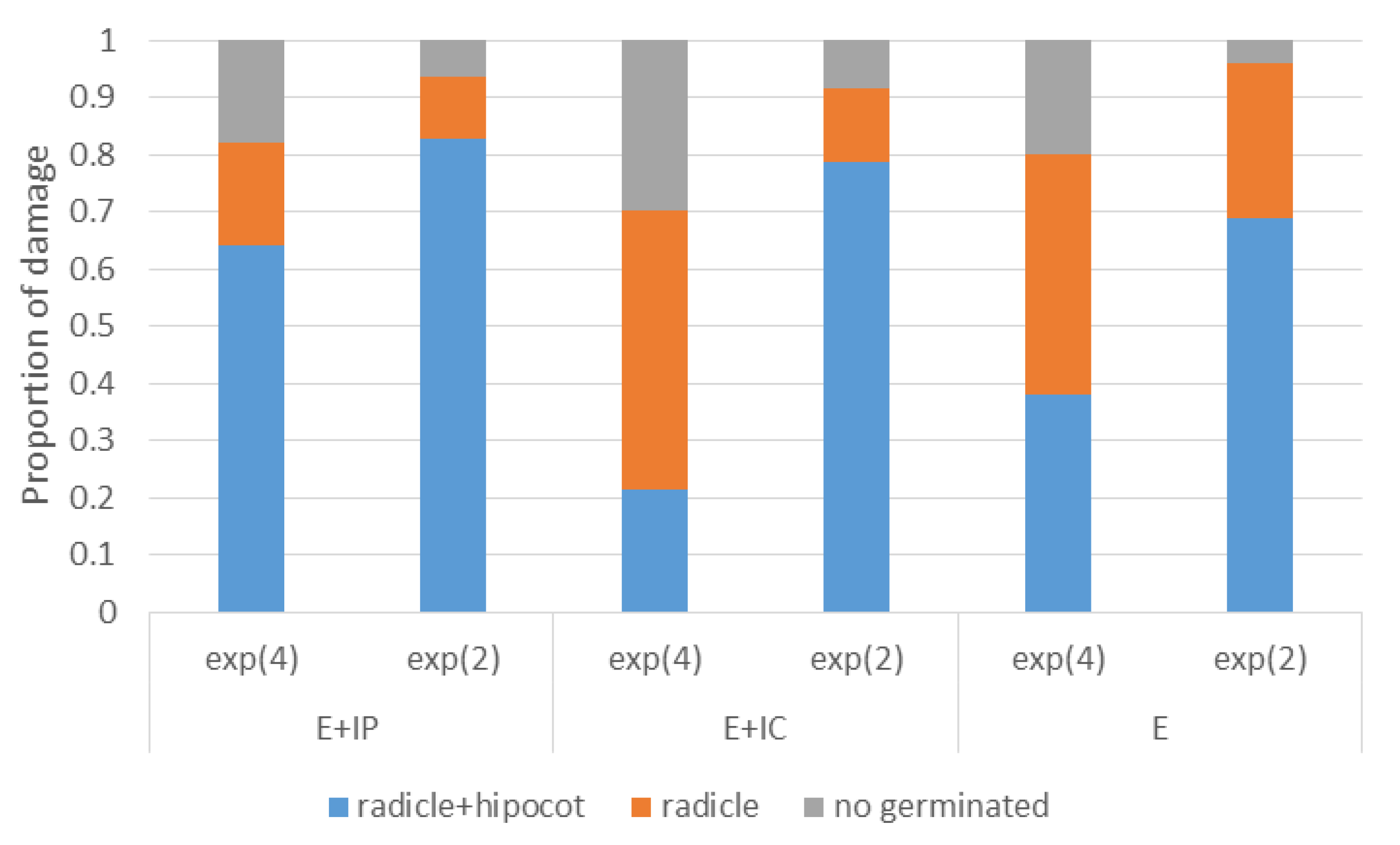
According to the estimated odds ratio (OR) (
Table 2), there were more germlings remaining in growth stage 1 when presence of
F. circinatum was external than if the fungus was within the embryo (
Figure 4) (in a proportion of 1 to 0.3; OR = 0.3,
p-value = 0.001). There were not differences when the pathogen was external or infecting the gametophyte (
p-value = 0.433). Regarding the dose, seeds inoculated with 10
4 spores/mL were three times more likely to remain in that stage than seeds inoculated with 10
2 spores/mL (
Table 2). Proportion of damaged germlings at growth stage 2 depended on inoculation method (
p-value = 0.0016) and dose (
p-value < 0.0001). Estimated odds ratio (
Table 2) showed that seeds with embryo and coat infected (protrusion-stage infection) were 2.4 times more likely to express symptoms than seeds with only coat infected (external infection).
Table 1.
Number of Pinus radiata (P. radiata) germlings in each category of growth stage at 48 dpi, with presence or absence of Fusarium circinatum (F. circinatum) mycelia for each inoculation method and dose applied to surface-disinfested seeds.
Table 1.
Number of Pinus radiata (P. radiata) germlings in each category of growth stage at 48 dpi, with presence or absence of Fusarium circinatum (F. circinatum) mycelia for each inoculation method and dose applied to surface-disinfested seeds.
| Growth stage (1) | Fc (2) | E + IP (3) | E + IC (3) | E (3) | Sum | Sum of inoculated |
|---|
| | | 104 (4) | 102 | wt | 104 | 102 | wt | 104 | 102 | wt | | |
| 2 | 0 | 0 | 4 | 43 | 1 | 3 | 40 | 0 | 2 | 46 | 139 | 10 |
| 2 | 1 | 32 | 38 | 0 | 10 | 37 | 0 | 19 | 33 | 0 | 169 | 169 |
| Total 2 | | 32 | 42 | 43 | 11 | 40 | 40 | 19 | 35 | 46 | 308 | 179 |
| 1 | 0 | 0 | 0 | 3 | 2 | 0 | 5 | 0 | 0 | 0 | 10 | 2 |
| 1 | 1 | 9 | 5 | 0 | 23 | 6 | 0 | 21 | 13 | 0 | 77 | 77 |
| Total 1 | | 9 | 5 | 3 | 25 | 6 | 5 | 21 | 13 | 0 | 87 | 79 |
| Total 0 | | 9 | 3 | 4 | 14 | 4 | 5 | 10 | 2 | 4 | 55 | 42 |
| Total | | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 450 | 300 |
Table 2.
Odds ratio estimated by logistic regression of proportion of P. radiata damaged germlings on inoculation method and inoculum dose.
Table 2.
Odds ratio estimated by logistic regression of proportion of P. radiata damaged germlings on inoculation method and inoculum dose.
| | Growth 1 stage 1 | Growth stage 2 |
|---|
| | OR | p-value | 95% IC | OR | p-value | 95% IC |
|---|
| IP 2 vs. E | 0.30 | 0.001 | 0.145, 0.612 | 2.35 | 0.0064 | 1.272, 4.351 |
| IC vs. E | 0.78 | 0.4328 | 0.421, 1.448 | 0.80 | 0.4536 | 0.443, 1.439 |
| IP vs. IC | 0.38 | 0.0095 | 0.185, 0.790 | 2.95 | 0.0006 | 1.589, 5.463 |
| 104 vs. 102 | 3.0 | 0.0001 | 1.707, 5.257 | 0.25 | <0.0001 | 0.152, 0.411 |
Figure 4.
Proportion of P. radiata germlings in each category of growth stage at 48 dpi, with presence of F. circinatum mycelia for each inoculation method and dose applied to surface-disinfested seeds. Exp (4) = 104 spores/mL; exp (2) = 102 spores/mL; E + IP = internal by immersion of pre-germinated seeds; E + IC = internal by immersion of seeds previously rubbed with carborundum: E = external by immersion.
Figure 4.
Proportion of P. radiata germlings in each category of growth stage at 48 dpi, with presence of F. circinatum mycelia for each inoculation method and dose applied to surface-disinfested seeds. Exp (4) = 104 spores/mL; exp (2) = 102 spores/mL; E + IP = internal by immersion of pre-germinated seeds; E + IC = internal by immersion of seeds previously rubbed with carborundum: E = external by immersion.
The percentage of germlings growing from inoculated seeds that developed radicle and hypocotyl at the end of the experiment (stage 2) was 60%. Of these germlings, all but 10 showed symptoms of
F. circinatum, whereas germlings grown from non-inoculated seed showed no lesions (
Table 1). The transmission rate from seed to germinating seeds and seedlings showing symptoms was determined and results are presented in
Table 3. The rate of transmission from seed to germinating seeds causing pre-emergence death was always greater for the high inoculum dose whereas this transmission rate at post-emergence was always greater for the low dose. Infection after inoculation at the protrusion stage (E + IP) with 10
4 spores/mL was lower than those rates for other methodsat pre-emergence and higher at post-emergence.
Table 3.
Transmission rate of F. circinatum from inoculated seeds to germinating seeds and symptomatic seedlings at 48 days after inoculation as determined in vitro and in potting soil assays.
Table 3.
Transmission rate of F. circinatum from inoculated seeds to germinating seeds and symptomatic seedlings at 48 days after inoculation as determined in vitro and in potting soil assays.
| Assay | Inoculation method (1) | Inoculum dose | Pre-emergence death | Post-emergence death | Total disease development |
|---|
| “in vitro” | E + IP | 104 | 0.36 (2) | 0.64 | 1 |
| | | 102 | 0.16 | 0.76 | 0.92 |
| | E + IC | 104 | 0.78 | 0.2 | 0.98 |
| | | 102 | 0.2 | 0.74 | 0.94 |
| | E | 104 | 0.62 | 0.38 | 1 |
| | | 102 | 0.3 | 0.66 | 0.96 |
| | average | 104 | 0.59 | 0.41 | 1 |
| | average | 102 | 0.22 | 0.72 | 0.94 |
| potting soil | E | 106 | 0.33 | 0.47 | 0.8 |
| | | 104 | 0.15 | 0.13 | 0.28 |
3.4. Temporal Analysis of Pre- and Post- Emergence Damping-Off, and of Death of Established Seedlings
Inoculum on seeds was checked and it was present in all of them with estimates of 180–400 cfu/seed when inoculated with 106 spores/mL, and ≤10 cfu/seed when inoculum was 104 or 102 spores/mL. Highest emergence occurred respectively at 23 and 24 dpi for each repetition of the experiment, and disease symptoms became visible one week later. The major symptom was root collar rot, where mycelia could be observed. For older seedlings, the more generalized symptom was a wilting, starting at the top of the seedling and progressing down the stem until the seedling collapsed. Roots were underdeveloped.
Pre-emergence damping-off was 33.3% and 15.7% (
Table 4) when seeds of
P. radiata were inoculated respectively with 10
6 and 10
4 spores/mL, but only the higher dose had a significant effect compared to the low inoculum dose and the untreated control (
Table 4). In post-emergence, inoculum dose had a significant effect on death of seedlings, with differences between both doses being greater in post-emergence (2-month-old seedlings) than in older seedlings (approximately 9-month-old seedlings) (
Table 4).
Table 4.
Effect of inoculum doses on percentage of pre- and post-emergence damping off, and death of established seedlings caused by F. circinatum on externally inoculated seeds of P. radiata.
Table 4.
Effect of inoculum doses on percentage of pre- and post-emergence damping off, and death of established seedlings caused by F. circinatum on externally inoculated seeds of P. radiata.
| Inoculum dose (spore/mL) | Pre-emergence damping-off (%) | Post-emergence damping-off (%) (3) | Death of seedlings (%) (3) |
|---|
| 106 (1) | 33.3 (4.67) (2) a (4) | 71.4 (5.07) a | 88.4 (4.10) a |
| 104 (1) | 15.7 (3.10) b | 24.7 (3.69) b | 66.0 (5.73) b |
| 0 (control) | 7.6 (5.08) b | 0 | 0 |
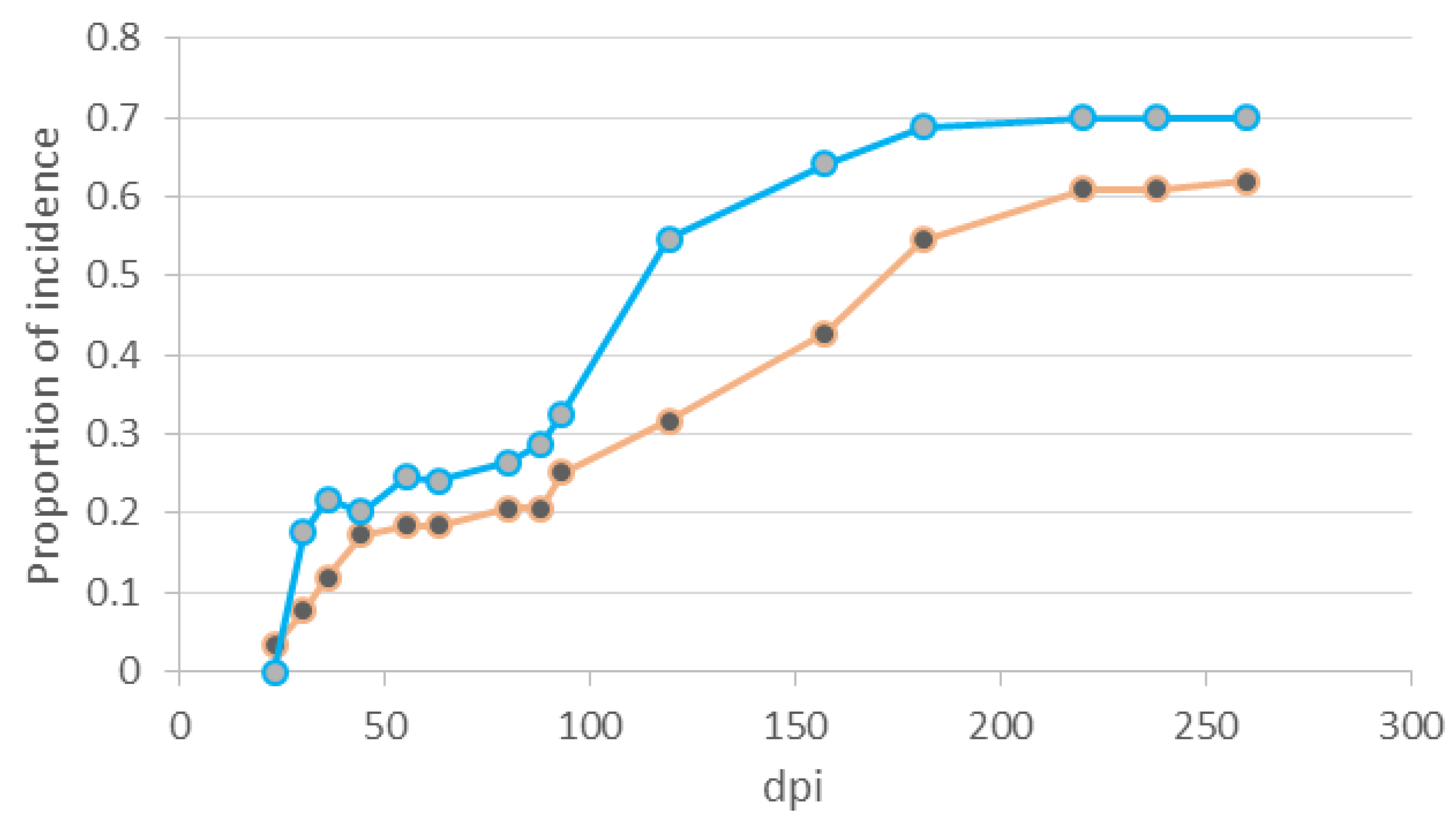
Disease incidence progressed throughout the experiment (until 281 dpi) with two peaks for which disease rate was highest (
Figure 5). One peak was during post-emergence stage from 30–36 dpi, and the other was during the period of established seedlings, at 93 dpi for the first repetition, and at 114 dpi for the second one. (
Figure 5). Although disease rate was not uniform in each of the two periods, the best model describing the disease progress curve was the lineal model (disease incidence regressed on time) being the slope of that line an estimate of the disease infection rate. The rate of disease incidence per day was 0.0025 ± 0.00067 (standard error) for damping-off in post-emergence and 0.0016 ± 0.00024 during the period of established seedlings. However, these rates were not significantly different.
Figure 5.
Disease progress curve on seedlings of P. radiata grown from seeds externally inoculated with 104 spores/mL of F. circinatum. Each curve is the mean of three replicates in each experiment repetition (repetition 1 for orange curve and repetition 2 for blue curve). Dpi means days post inoculation.
Figure 5.
Disease progress curve on seedlings of P. radiata grown from seeds externally inoculated with 104 spores/mL of F. circinatum. Each curve is the mean of three replicates in each experiment repetition (repetition 1 for orange curve and repetition 2 for blue curve). Dpi means days post inoculation.
Specific values of disease incidence at 48 dpi were used to determine the transmission rate from seed to symptomatic seedlings (
Table 3). Transmission rates at this time of the disease development are similar in pre- and post-emergence for the low inoculum dose (10
4 spores/mL), and higher in post-emergence for the high inoculum dose.
3.5. Persistence of Fusarium circinatum in Seedlings
Fusarium circinatum was quantified in a sample of P. radiata dead seedlings (five and 10 seedlings from seeds inoculated respectively with 106 and 104 spores/mL). Populations in roots were in the range of 105–107 cfu/gr fresh weight for both seed inoculation doses, and in stems between 102 and 105 cfu/gr fresh weight. F. circinatum was not detected in the stem of three dead plants, however, it could be present since detection level was about 120 cfu/gr. Estimates of F. circinatum cfu were in those ranges regardless of whether seedlings came from inoculated seeds with either 106 or 104 as it was observed in root populations.
At 281 dpi, 30 seedlings remained symptomless. On seven of these, F. circinatum was estimated and it was detected in roots of five plants, with populations up to 104 cfu/gr. In stems of these plants, F. circinatum was only detected in one plant (105 cfu/gr). The other 23 seedlings were moved to a greenhouse and grown in pots from 281 dpi to 479 dpi. All plants (from both seed inoculation doses) except two of them died in this period, and F. circinatum was detected in all dead plants, in roots and stems, except in the stem of one of these plants. The two plants remaining at the end of the experiment were symptomless; however, F. circinatum was detected in their roots, stems and tips. The final percentage of symptomless seedlings at 281 dpi was 14% and at 481 dpi was 1%.
4. Discussion
F. circinatum was isolated from
P. radiata seeds from trees with cankers of pitch canker disease, but at a frequency less than 1%. This is a very low rate compared to previous studies. In California,
F. circinatum was isolated from up to 83% of seeds collected from
P. radiata cones on infected branches [
11]. For other
Pinus species, the pathogen was also isolated from 98% of
P. palustris seeds [
29] of which 84% was internal infection; in
P. taeda seeds, external presence varied from 7%–61%, and internal infection from 1%–34% [
30]; in
P. elliottii seeds, internal presence of
F. circinatum was up to 30% [
15].
Besides
Pinus species, environmental conditions also influence the extent of infection [
31]. Seeds analyzed here were collected in one year from eight different locations, and this short sampling time could influence the low level of
F. circinatum detected. However, these results are in agreement with those obtained in routine seed health testing that has been performed in Spain since
F. circinatum was detected in 2005, which are very low (personal communication). In this case, it is assumed that seeds are collected from orchard trees with no pitch canker symptoms, but
F. circinatum may also be present on seeds collected from cones of apparently healthy branches [
11] coming from airborne spores that enter the cone when it is open.
The low seed infection rate could be also due to the location of sampled cones, which may be on branches without symptoms, especially in those trees sampled with one or two cankers. For these trees, percentage of seeds with presence of
F. circinatum was less than 1% for each plantation (
Figure 1), but again this is very low compared to reports from two locations in California whereby 9.2% and 11.7% of seeds were infected from symptomless branches [
11]. Another reason to explain the low infection could be the method used here for detecting the pathogen on seeds. Plating seeds prior to germination does not detect cryptic infections, which are only detected from emerging seedlings [
11].
Fusarium circinatum was mainly on the seed coat and less frequently in the gametophyte. This reveals that infection, at least mainly, is not systemically transmitted from the mother plant to the embryo, but is transmitted indirectly by air-borne spores entering cones at time of pollination or/and penetrating mature cones through small cracks. No coincidence of pollination and dispersal of fungal spores or low amount of them could explain the low seed infection found.
The present study showed that external presence of the pathogen on the seed can increase due to contamination during storage, and then seeds may become an important inoculum source in nurseries. Management of storage conditions, especially low relative humidity level, can help to prevent contamination and therefore it is highly recommended to store seeds under low relative humidity. Further studies are required to optimize storage conditions, and to determine the best treatments to reduce or eliminate the external presence of the fungus, or at least to reduce chances of contamination.
Our work showed that transmission from seed to seedling is accomplished even with presence of F. circinatum limited to the seed coat (external inoculation) with an inoculum dose lower than 10 spores per seed, and these seeds can be a source of primary inoculum for pitch canker disease in nurseries. The fungus was transmitted to seedlings causing death in pre- or post- emergence at different rates depending on the inoculum dose. Rates of transmission were determined from seeds to symptomatic seedlings, but we showed that fungus was also transmitted to the remaining symptomless seedlings in potting soil. Of these, some developed disease later and others did not, but the fungus was detected in all seedlings coming from seeds originally inoculated with F. circinatum. Seed transmission rate was higher in vitro than in potting soil, since the humid environment created in germination trays was more conducive for fungus growth and disease development. This suggests that humidity level in soil during seed germination may influence pathogen transmission in nurseries.
When the disease outbreak occurs in nurseries, the pathogen could be disseminated to new plantations through symptomless seedlings. Seedlings progressively developed pitch canker symptoms during the first 15 months, but a few did not develop symptoms at the end of the experiment (two of a total 203 seedlings that reached post-emergence). It is unknown if these established seedlings will show disease once grown in a plantation, but this could be one of the reasons why pitch canker disease remains latent for years, as has been observed in South Africa [
32] and in Spain. The pathogen was detected in the roots of both types of seedlings, with and without symptoms, as was previously reported by Storer
et al. [
11]. We did not detect the pathogen in two symptomless seedlings (inoculated with low dose) but this could be due to either
F. circinatum population being less than the detection level (13–20 cfu/mL) or simply an infection escape. We found that fungus population present in the roots of seedlings with symptoms was always higher than in seedlings without symptoms, this population being lower than 10
4 cfu/g. Pathogen population in stems of the seedlings with symptoms was always lower than in roots. Because studies on colonization and symptoms of
F. circinatum in established plants have focused on the aerial part of the tree (see e.g., [
33,
34]), the role of root colonization on persistence and dissemination of
F. circinatum may have been underestimated.
Pre-emergence damping-off is caused by germination failure or damages to the radicle. Results from the germling growth assay indicated that final germination
in vitro was not affected by whether the inoculum was located in embryo, gametophyte or coat, as suggested when comparing different inoculation methods (
Figure 3). On the contrary, damages in the radicle preventing further growth were caused by presence of
F. circinatum in the gametophyte or on the seed coat (
Table 3). The reason why there was less damage when the embryo was infected is unknown, but could be influenced by the time the embryo was infected, in the protrusion stage, which may not stop seeds from growing. It may suggest that
F. circinatum inhibited hypocotyl development since the number of germlings that remained in stage 1 after inoculation at the high dose was about double in the treatment without protrusion pretreatment. Higher inoculum dose led to greater damage at the germination plus radicle stage (OR = 3,
Table 2) than at the stage of radicle plus hypocotyl (OR = 0.25,
Table 2). This is in agreement with damages quantified in pre-emergence, which was only significant for seeds inoculated with a high dose (
Table 3).
Once seedlings emerged, high inoculum doses caused greater mortality in the first two months, while low dose caused damage that progressed steadily until 281 dpi (
Table 3,
Figure 5). Viljoen
et al. [
3] observed that mortality of three-year-old seedlings of
P. patula was lower than that of newly germinated seedlings. So, our results corroborate a lower mortality for established seedlings, but only for high inoculum doses. We determined the infection rate in optimal conditions for the pitch canker disease at low inoculum doses, during post-emergence and established seedlings, from lineal models adjusted to the disease progress curve during each growth period (
Figure 5). Disease infection rates were not significantly different, corroborating that mortality in post-emergence and in established seedlings is similar when seeds were inoculated with 10
4 spores/mL.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}