
Lichen Monitoring Delineates Biodiversity on a Great Barrier Reef Coral Cay

Abstract
:
1. Introduction
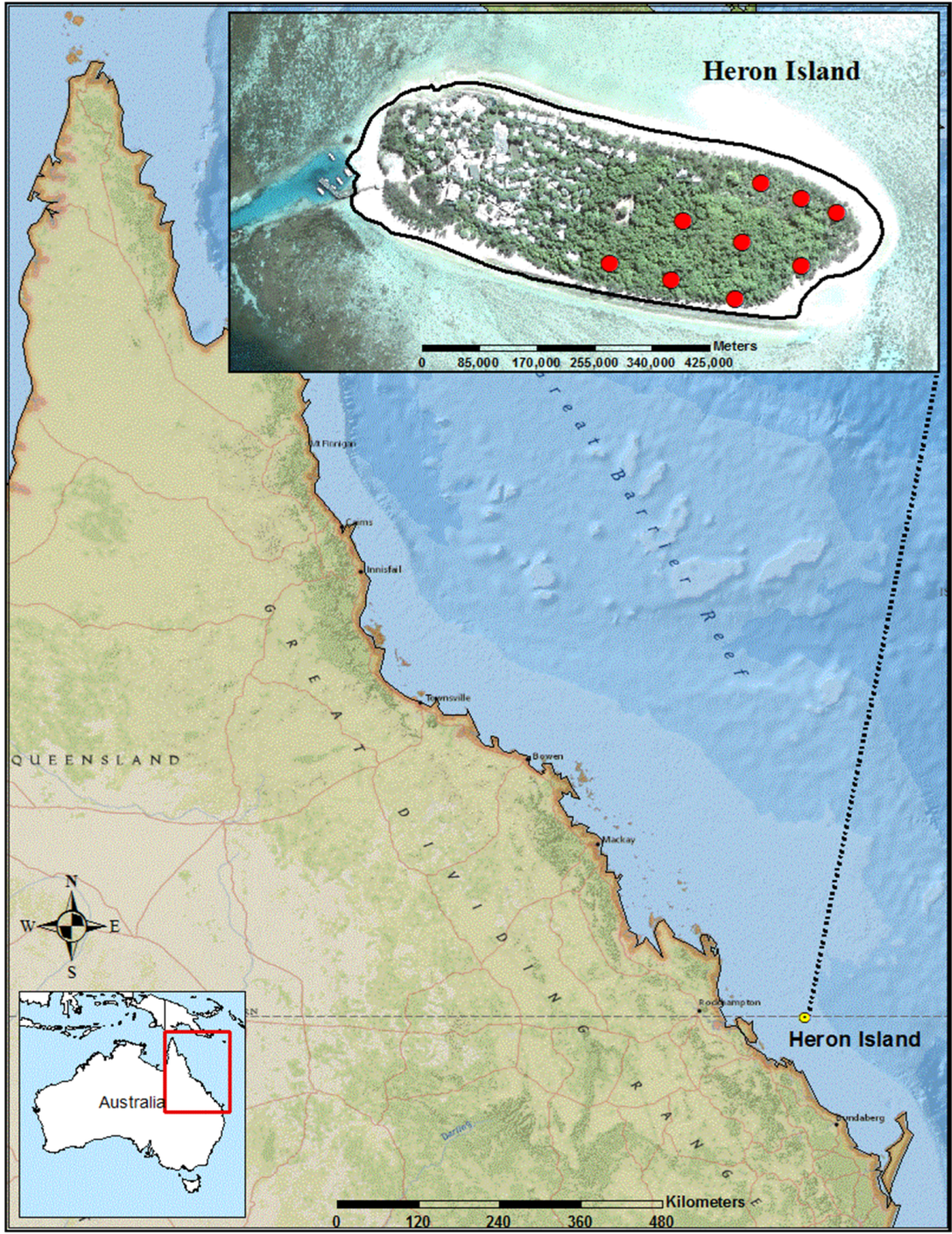
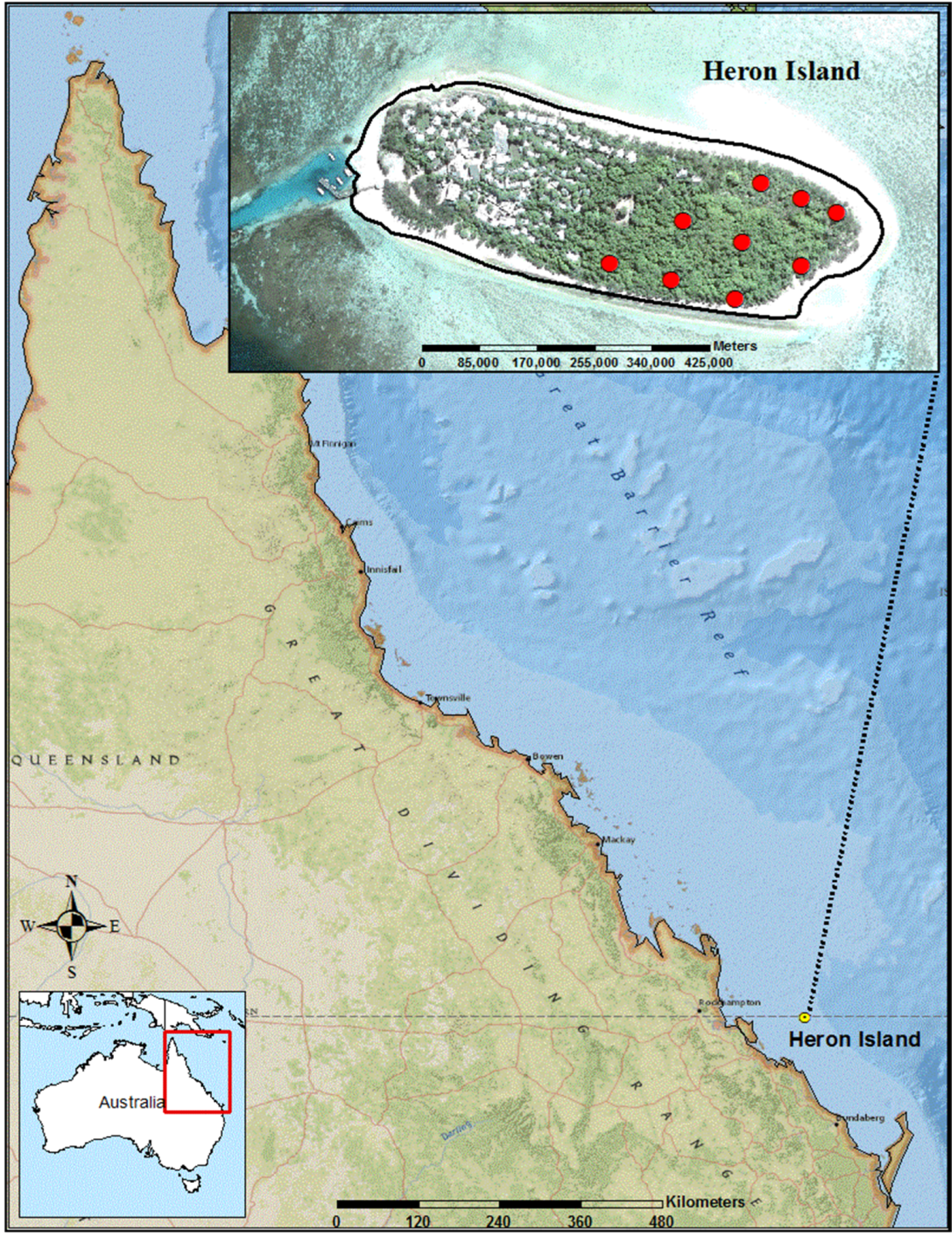
2. Materials and Methods
2.1. Study Area and Field Methods
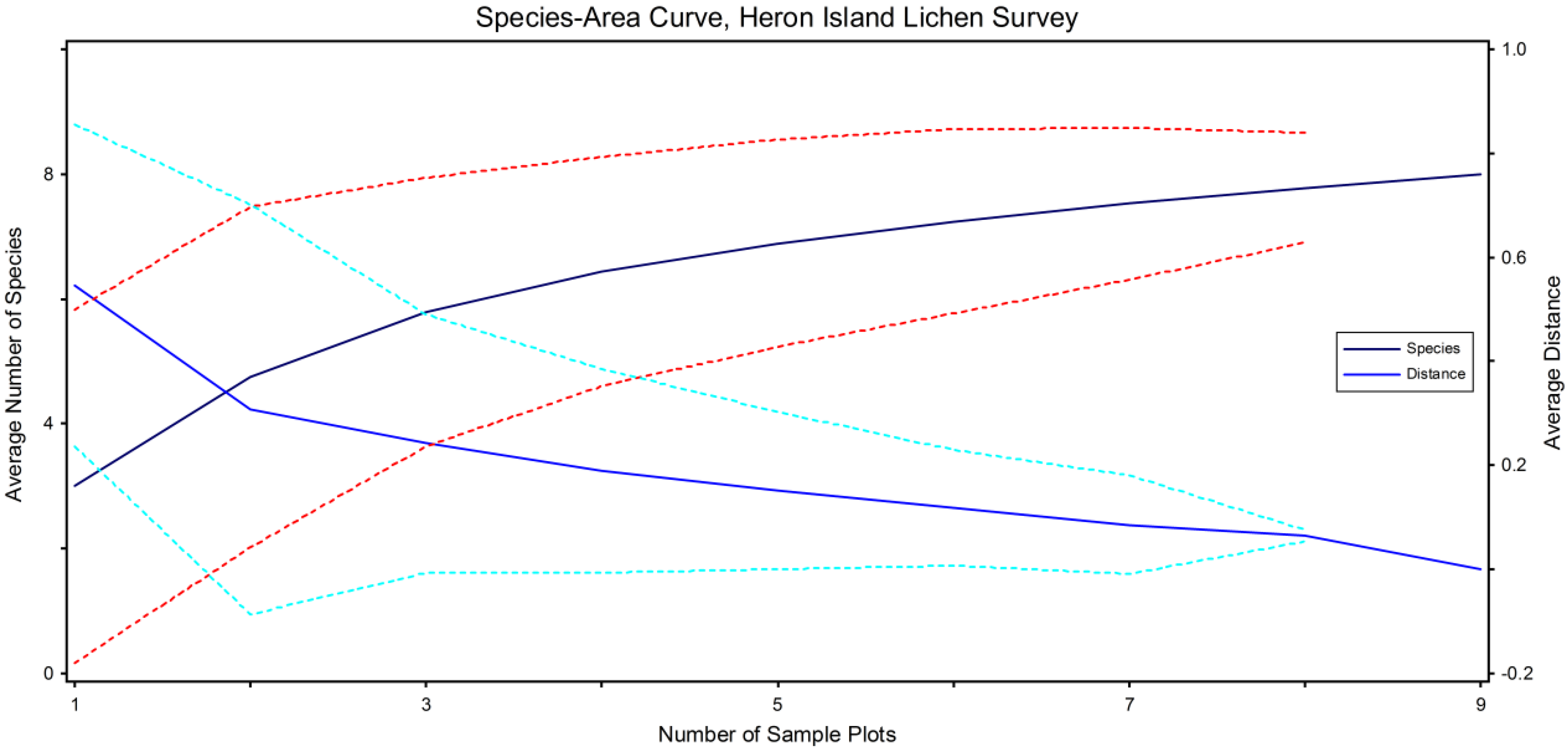
2.2. Derived Variables and Analytical Methods
3. Results
3.1. Forest Conditions and Lichen Species of Heron Island

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot ID | Forest Type | UTM Easting | UTM Northing | Distance to Forest Edge (m) | Trees ha−1 | Total BA (m2/ha−1) | Live BA (m2/ha−1) | Pisonia BA (m2/ha−1) | Tree Species Count | Lichen Species Count | Total Lichen Abundance |
|---|---|---|---|---|---|---|---|---|---|---|---|
| H1 | CAEQ/ARAR | 389,458 | 7,407,117 | 18.80 | 780 | 9.54 | 9.54 | 0.00 | 4 | 4 | 14 |
| H2 | CAEQ/ARAR | 389,405 | 7,407,147 | 8.00 | 325 | 41.17 | 40.66 | 29.39 | 4 | 4 | 12 |
| H3 | CAEQ/ARAR | 389,368 | 7,407,153 | 8.69 | 390 | 32.07 | 31.06 | 0.00 | 3 | 4 | 14 |
| H11 | PIGR mixed | 389,467 | 7,406,983 | 14.70 | 975 | 46.72 | 46.72 | 32.52 | 3 | 3 | 10 |
| H13 | PIGR | 389,387 | 7,407,023 | 83.83 | 1755 | 144.87 | 144.74 | 144.87 | 1 | 1 | 4 |
| H15 | PIGR mixed | 389,285 | 7,407,039 | 103.10 | 1950 | 28.92 | 27.49 | 15.90 | 4 | 2 | 6 |
| H21 | PIGR mixed | 389,342 | 7,406,946 | 14.36 | 1820 | 116.84 | 116.48 | 109.29 | 3 | 5 | 12 |
| H23 | PIGR | 389,261 | 7,406,963 | 25.43 | 1885 | 97.19 | 97.19 | 96.80 | 2 | 3 | 7 |
| H25 | PIGR | 389,133 | 7,407,033 | 31.75 | 1170 | 75.91 | 74.44 | 74.26 | 3 | 1 | 4 |
| Species | Species Code | Form | Frequency of Presence (% Plots) | Landscape Abundance | Substrate Tree Species |
|---|---|---|---|---|---|
| LICHENS | |||||
| Dirinaria picta | DIPI | foliose | 33 | 11 | CAEQ |
| Pyxine cocoes | PYCO | foliose | 56 | 19 | ARAR, CAEQ, PAHE, PIGR |
| Hyperphyscia adglutinata | HYAD | foliose | 66 | 17 | ARAR, CAEQ, PAHE, PIAR, PIGR |
| Coenogonium queenslandicum | COQU | crustose | 22 | 3 | ARAG, PIGR |
| Lecanora arthothelinella | LEAR | crustose | 11 | 2 | ARAR |
| Strangospora ochrophora | STOC | crustose | 11 | 2 | PIGR |
| ASSOCIATES | |||||
| Cyanobacterium | CYANO | crustose | 56 | 13 | ARAR, CAEQ, PIGR |
| Sterile thalli | THALLI | crustose | 44 | 16 | PIGR |
3.2. Lichen Community Differences among Forest Types
| Forest Type Pairs | T | A | p |
|---|---|---|---|
| CAEQ/ARAR vs. PIGR mixed | −2.457 | 0.190 | 0.025 |
| CAEQ/ARAR vs. PIGR | −2.604 | 0.490 | 0.024 |
| PIGR mixed vs. PIGR | −1.364 | 0.235 | 0.097 |
| All types (grand test) | −2.704 | 0.411 | 0.015 |
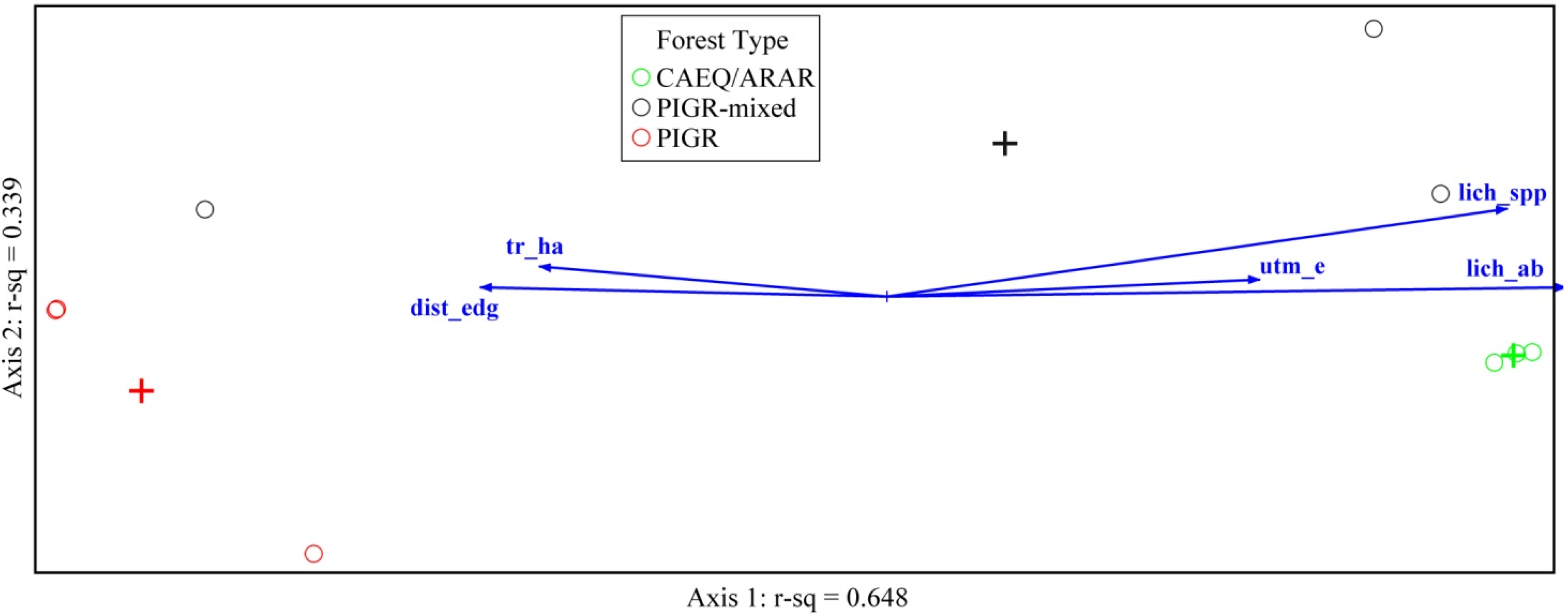
3.3. Environmental Factors Affecting Coral Cay Lichen Communities
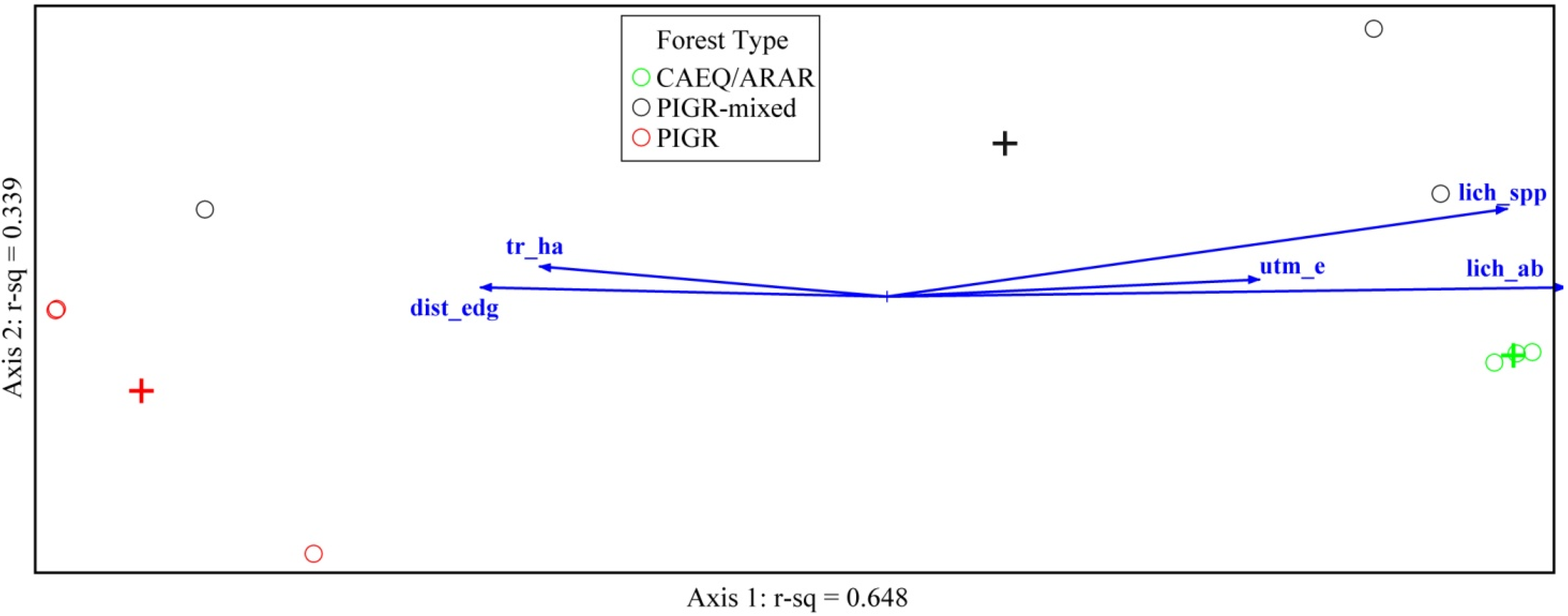

| r Value | |||
|---|---|---|---|
| Code | Axis 1 | Axis 2 | |
| ENVIRONMENTAL VARIABLES | |||
| UTM Easting | utm_e | 0.710 | 0.151 |
| UTM Northing | utm_n | 0.401 | −0.318 |
| Distance to Forest Edge (beach) | dist_edge | −0.741 | 0.109 |
| Number of Trees ha−1 | tr_ha | −0.685 | 0.202 |
| Total Basal Area ha−1 | ba_total | −0.491 | 0.111 |
| Live Basal Area ha−1 | ba_live | −0.485 | 0.108 |
| Pisonia grandis Basal Area ha−1 | ba_pigr | −0.547 | 0.072 |
| Number of Tree Species | tr_spp | 0.499 | 0.216 |
| Number of Lichen Species | lich_spp | 0.916 | 0.344 |
| Total Lichen Abundance (Plot) | lich_ab | 0.959 | 0.113 |
| LICHEN SPECIES | |||
| Dirinaria picta | DIPI | 0.667 | −0.299 |
| Pyxine cocoes | PYCO | 0.974 | 0.355 |
| Hyperphyscia adglutinata | HYAD | 0.978 | 0.145 |
| Coenogonium queenslandicum | COQU | −0.223 | 0.544 |
| Lecanora arthothelinella | LEAR | 0.261 | 0.695 |
| Strangospora ochrophora | STOC | −0.308 | −0.667 |
| Cyanobacteria (unknown) | CYANO | 0.964 | 0.231 |
| Sterile lichen thalli (unknown) | THALLI | −0.992 | −0.319 |
4. Discussion
4.1. Lichens as Indicators of Forest Diversity

4.2. Key Factors Influencing Pisonia-Dominated Lichen Communities
5. Conclusions
Acknowledgments
Author Contributions
%20ary





Conflicts of Interest
References
- Batianoff, G.N.; Naylor, G.C.; Olds, J.A.; Fechner, N.A.; Neldner, V.J. Climate and Vegetation Changes at Coringa-Herald National Nature Reserve, Coral Sea Islands, Australia 1. Pac. Sci. 2010, 64, 73–92. [Google Scholar] [CrossRef]
- Greenslade, P. Climate variability, biological control and an insect pest outbreak on Australia’s Coral Sea islets: Lessons for invertebrate conservation. J. Insect Conserv. 2008, 12, 333–342. [Google Scholar] [CrossRef]
- Ogden, J. On cyclones, Pisonia grandis and the mortality of Black Noddy Anous minutus on Heron Island. Emu 1993, 93, 281–283. [Google Scholar] [CrossRef]
- Woinarski, J. Biodiversity conservation in tropical forest landscapes of Oceania. Biol. Conserv. 2010, 143, 2385–2394. [Google Scholar] [CrossRef]
- Batianoff, G.N. Floristic, vegetation and shoreline changes on Masthead Island, Great Barrier Reef. Proc. R. Soc. Qld. 1999, 108, 1–11. [Google Scholar]
- Batianoff, G.N.; Hacker, J.L.F. Vascular plant protrait of Wilson Island, Great Barrier Reef, Australia. Proc. R. Soc. Qld. 2000, 109, 31–38. [Google Scholar]
- Walker, T.A.; Chaloupka, M.Y.; King, B.R. Pisonia islands of the Great Barrier Reef. Atoll Res. Bull. 1991, 350, 1–23. [Google Scholar] [CrossRef]
- Cribb, A.B.; Cribb, J.W. Plant Life of the Great Barrier Reef and Adjacent Shores; University of Queensland Press: St. Lucia, QLD, Australia, 1985. [Google Scholar]
- Nimis, P.L.; Scheidegger, C.; Wolseley, P. (Eds.) Monitoring with Lichens—Monitoring Lichens; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002.
- Neitlich, P.N.; McCune, B. Hotspots of epiphytic lichen diversity in two young managed forests. Conserv. Biol. 1997, 11, 172–182. [Google Scholar] [CrossRef]
- Rogers, P.C.; Ryel, R.J. Lichen community change in response to succession in aspen forests of the Rocky Mountains, USA. For. Ecol. Manag. 2008, 256, 1760–1770. [Google Scholar] [CrossRef]
- Rosso, A.L.; Rosentreter, R. Lichen diversity and biomass in relation to management practices in forests of northern Idaho. Evansia 1999, 16, 97–104. [Google Scholar]
- Will-Wolf, S.; Esseen, P.A.; Neitlich, P. Monitoring biodiversity and ecosystem function: Forests. In Monitoring with Lichens—Monitoring Lichens; Nimis, P.L., Scheidegger, C., Wolseley, P.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 203–222. [Google Scholar]
- Hedenås, H.; Ericson, L. Aspen lichens in agricultural and forest landscapes: The importance of habitat quality. Ecography 2004, 27, 521–531. [Google Scholar] [CrossRef]
- Barkman, J.J. (Ed.) Phytosociology and Ecology of Cryptogamic Epiphytes; Van Gorcum & Co.: Assen, The Netherlands, 1958.
- Morley, S.E.; Gibson, M. Successional changes in epiphytic rainforest lichens: Implications for the management of rainforest communities. Lichenologist 2010, 42, 311–321. [Google Scholar] [CrossRef]
- Abbott, I.; Williams, M.R. Silvicultural impacts in jarrah forest of Western Australia: Synthesis, evaluation, and policy implications of the Forestcheck monitoring project of 2001–2006. Aust. For. 2011, 74, 350–360. [Google Scholar] [CrossRef]
- Stevens, G.N.; Rogers, R.W. The macrolichens flora from the mangroves of Moreton Bay. Proc. R. Soc. Qld. 1979, 90, 33–49. [Google Scholar]
- Will-Wolf, S.; Scheidegger, C.; McCune, B. Methods for monitoring biodiversity and ecosystem function: Monitoring scenarios, sampling strategies and data quality. In Monitoring with Lichens—Monitoring Lichens; Nimis, P.L., Scheidegger, C., Wolseley, P.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 147–162. [Google Scholar]
- McCune, B. Lichen communities as indicators of Forest Health. New Front. Bryol. Lichenol. 2000, 103, 353–356. [Google Scholar]
- McCune, B.; Lesica, P. The trade-off between species capture and quantitative accuracy in ecological inventory of lichens and bryophytes in forests in Montana. Bryologist 1992, 95, 296–304. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. PC-ORD: Multivariate Analysis of Ecological Data; MjM Software: Gleneden Beach, OR, USA, 2006. [Google Scholar]
- Peck, J.E. Multivariate Analysis of Community Ecologists: Step-by-Step Using PC-ORD; MjM Sortware Design: Gleneden Beach, OR, USA, 2010. [Google Scholar]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM Software: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Kruskal, J.B. Nonmetric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Rogers, P.C.; Rosentreter, R.; Ryel, R. Aspen indicator species in lichen communities in the Bear River Range of Idaho and Utah. Evansia 2007, 24, 34–41. [Google Scholar] [CrossRef]
- Wolseley, P.A.; Hill, D.J. Methods for Monitoring Lichens. In Monitoring with Lichens—Monitoring Lichens; Nimis, P.L., Scheidegger, C., Wolseley, P.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 269–272. [Google Scholar]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Van Herk, C.M.; Mathijssen-Spiekman, A.M.; de Zwart, D. Long distance nitrogen air pollution effects on lichens in Europe. Lichenologist 2003, 35, 347–359. [Google Scholar]
- Jovan, S.; McCune, B. Using epiphytic macrolichen communities for biomonitoring ammonia in forests of the greater Sierra Nevada, California. Water Air Soil Pollut. 2006, 170, 69–93. [Google Scholar] [CrossRef]
- Rogers, P.C.; Moore, K.; Ryel, R.J. Aspen succession and nitrogen loading: A case for epiphytic lichens as bioindicators in the Rocky Mountains, USA. J. Veg. Sci. 2009, 20, 498–510. [Google Scholar] [CrossRef]
- Allaway, W.G.; Ashford, A.E. Nutrient input by seabirds to the forest on a coral island of the Great Barrier Reef. Mar. Ecol. Prog. Ser. 1984, 19, 297–298. [Google Scholar] [CrossRef]
- Schmidt, S.; Dennison, W.C.; Moss, G.J.; Stewart, G.R. Nitrogen ecophysiology of Heron Island, a subtropical coral cay of the Great Barrier Reef, Australia. Funct. Plant Biol. 2004, 31, 517–528. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y. Dynamics of epiphytic macrolichen abundance, diversity and composition in boreal forest. J. Appl. Ecol. 2015, 52, 181–189. [Google Scholar] [CrossRef]
- Kikkawa, J. Birds recorded at Heron Island. Sunbird 1970, 1, 34–48. [Google Scholar]
- Flood, P.G.; Jell, J.S. The effect of cyclone ‘David’ (January 1976) on the sediment distribution patterns on Heron Reef, Great Barrier Reef. In Proceedings of the Third International Coral Reef Symposium, University of Miami, Miami, FL, USA, May 1977; pp. 119–125.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rogers, P.C.; Rogers, R.W.; Hedrich, A.E.; Moss, P.T. Lichen Monitoring Delineates Biodiversity on a Great Barrier Reef Coral Cay. Forests 2015, 6, 1557-1575. https://doi.org/10.3390/f6051557
Rogers PC, Rogers RW, Hedrich AE, Moss PT. Lichen Monitoring Delineates Biodiversity on a Great Barrier Reef Coral Cay. Forests. 2015; 6(5):1557-1575. https://doi.org/10.3390/f6051557
Chicago/Turabian StyleRogers, Paul C., Roderick W. Rogers, Anne E. Hedrich, and Patrick T. Moss. 2015. "Lichen Monitoring Delineates Biodiversity on a Great Barrier Reef Coral Cay" Forests 6, no. 5: 1557-1575. https://doi.org/10.3390/f6051557