Large-Scale Regeneration Patterns of Pinus nigra Subsp. salzmannii: Poor Evidence of Increasing Facilitation Across a Drought Gradient



Abstract
:1. Introduction
2. Materials and Methods
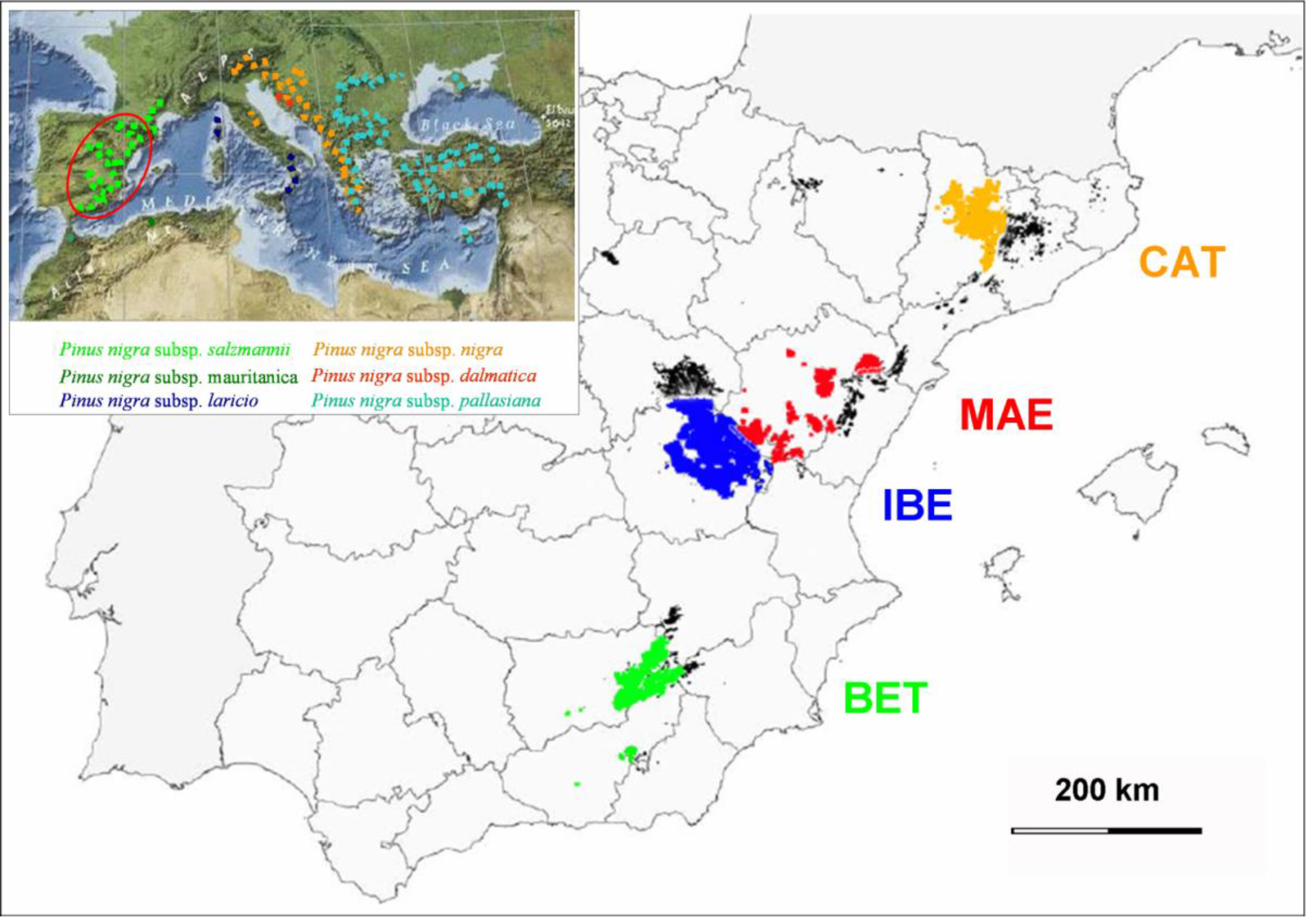
2.1. Species, Data Set, and Study Variables
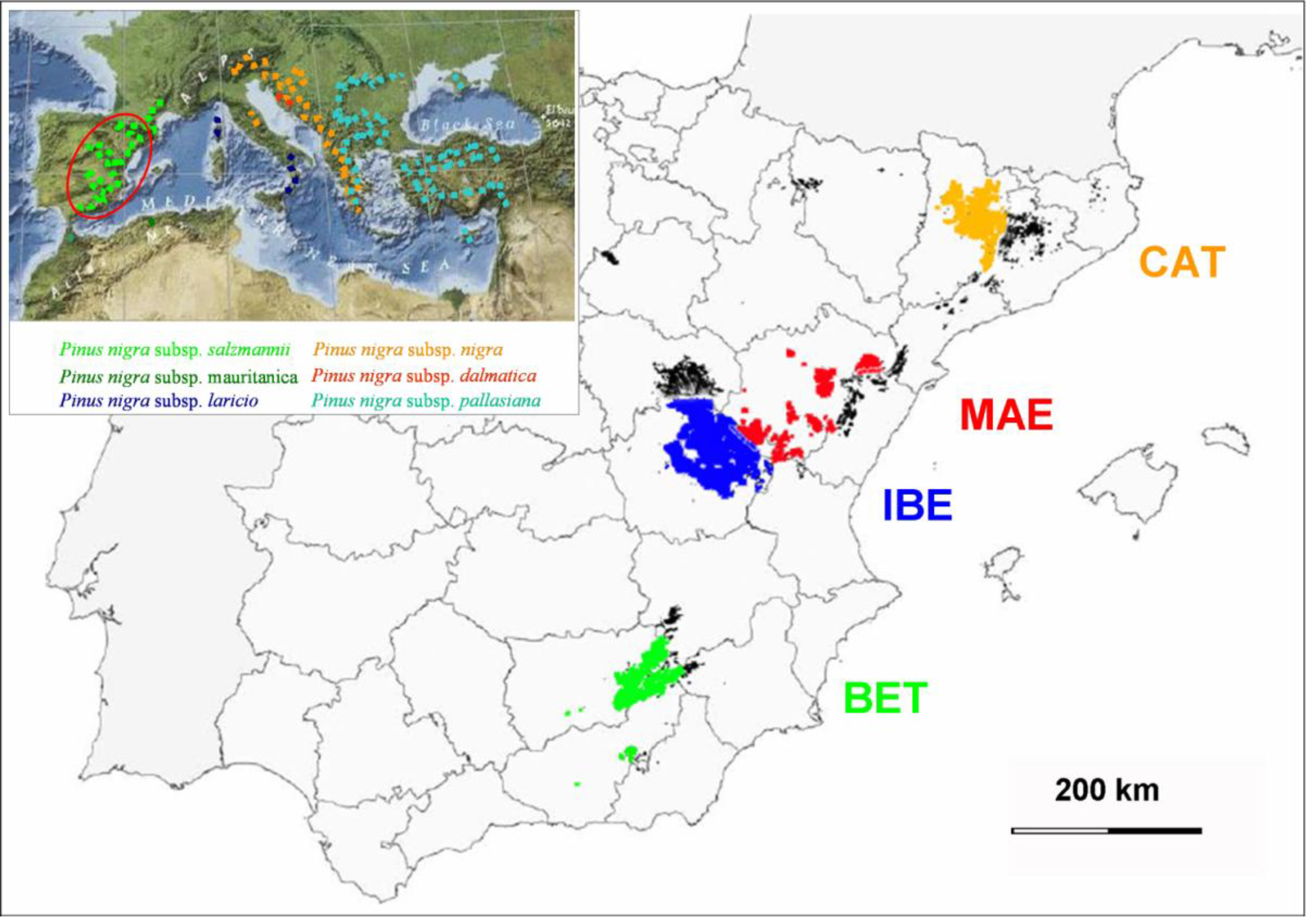
2.3. Statistical Analysis
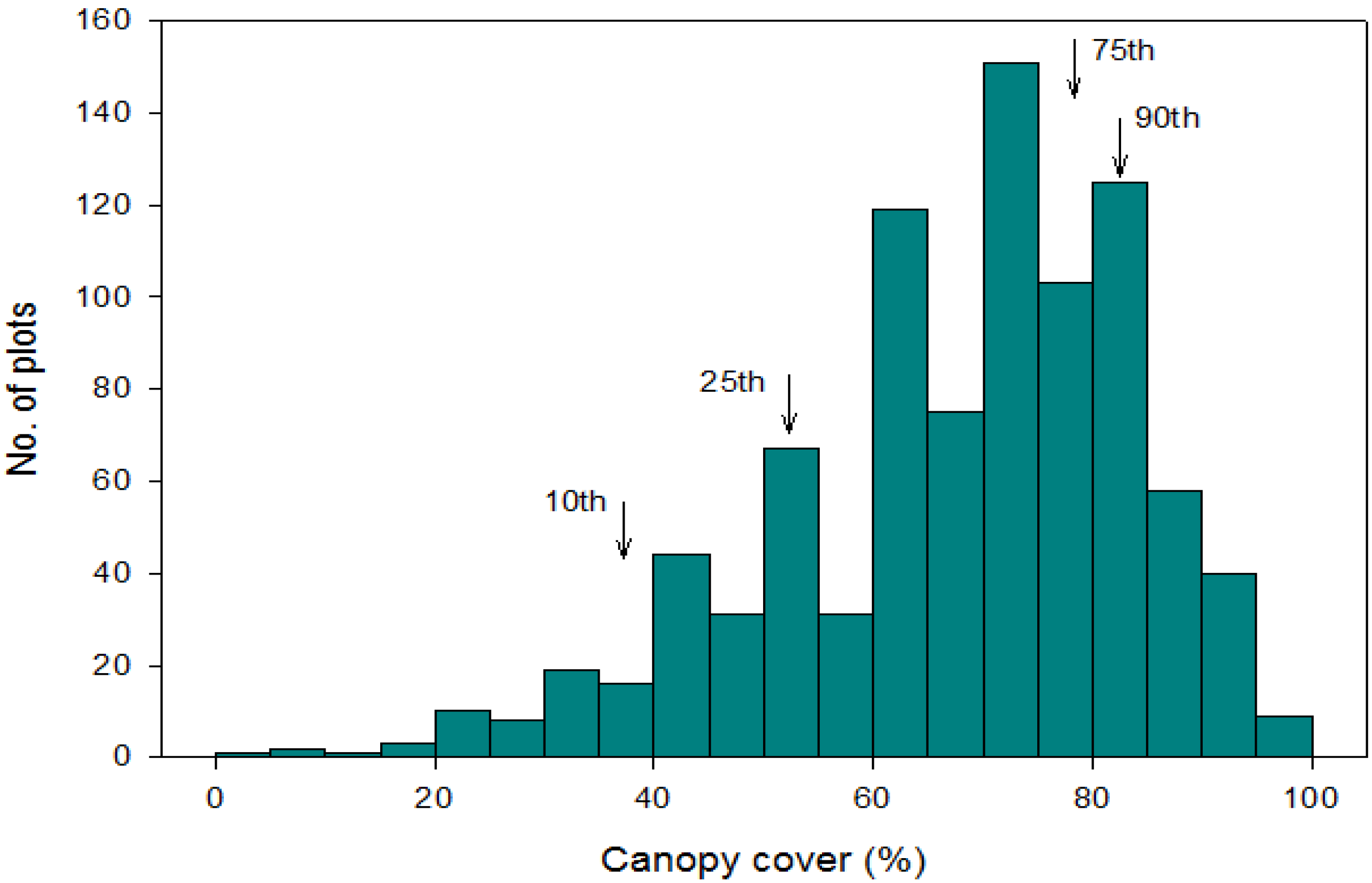
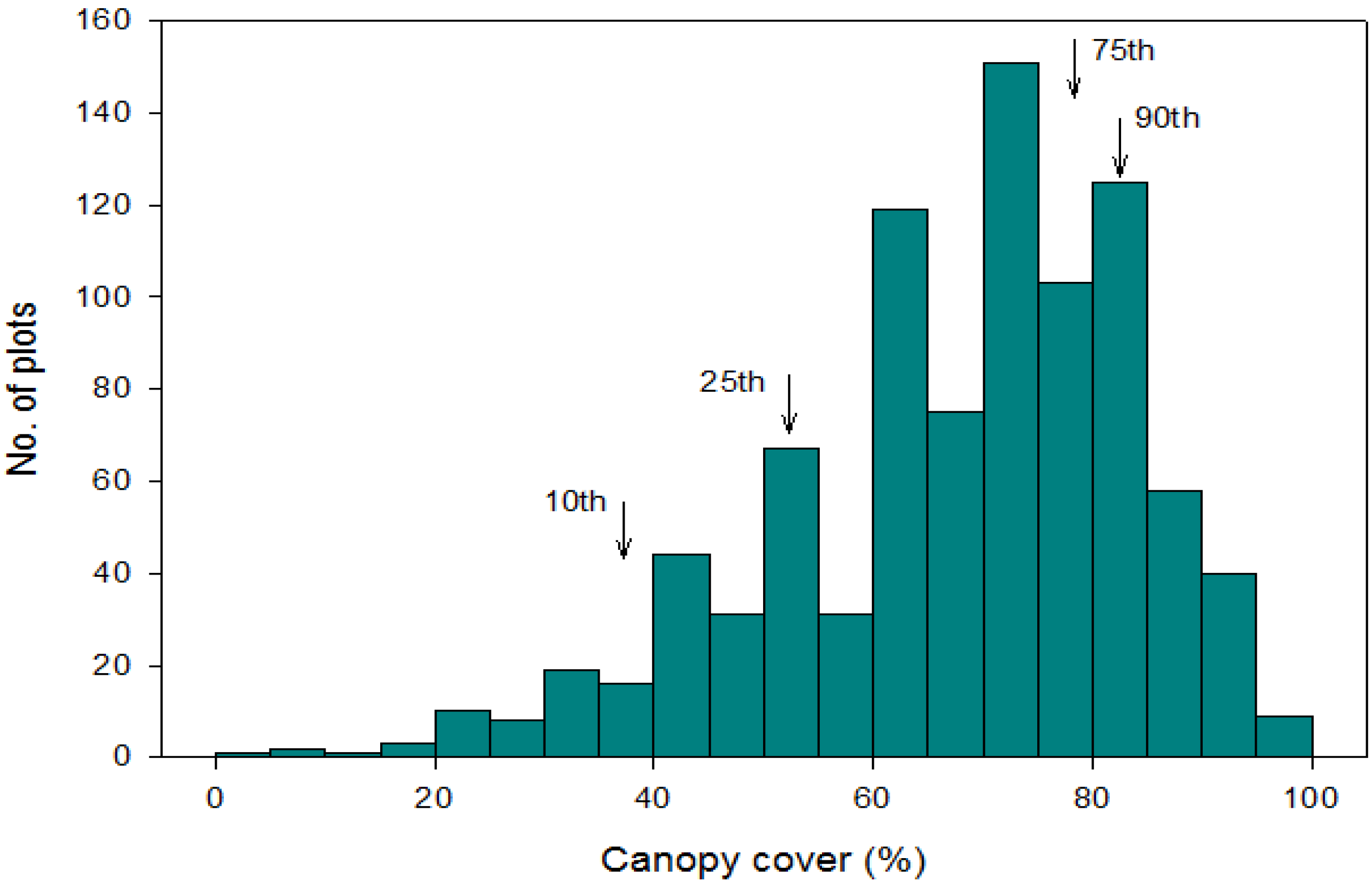
3. Results
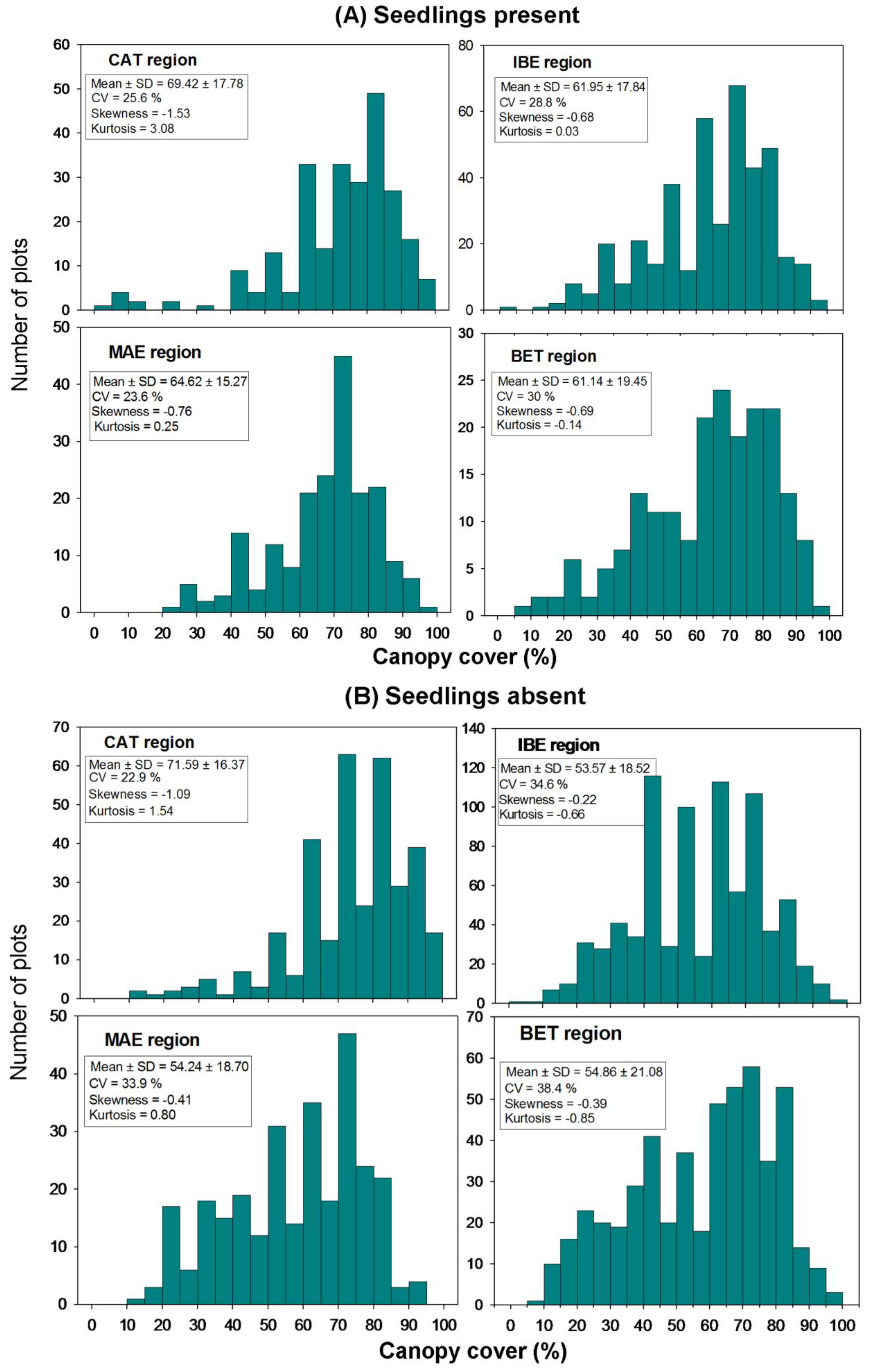
3.1. Climate and Soil Gradients
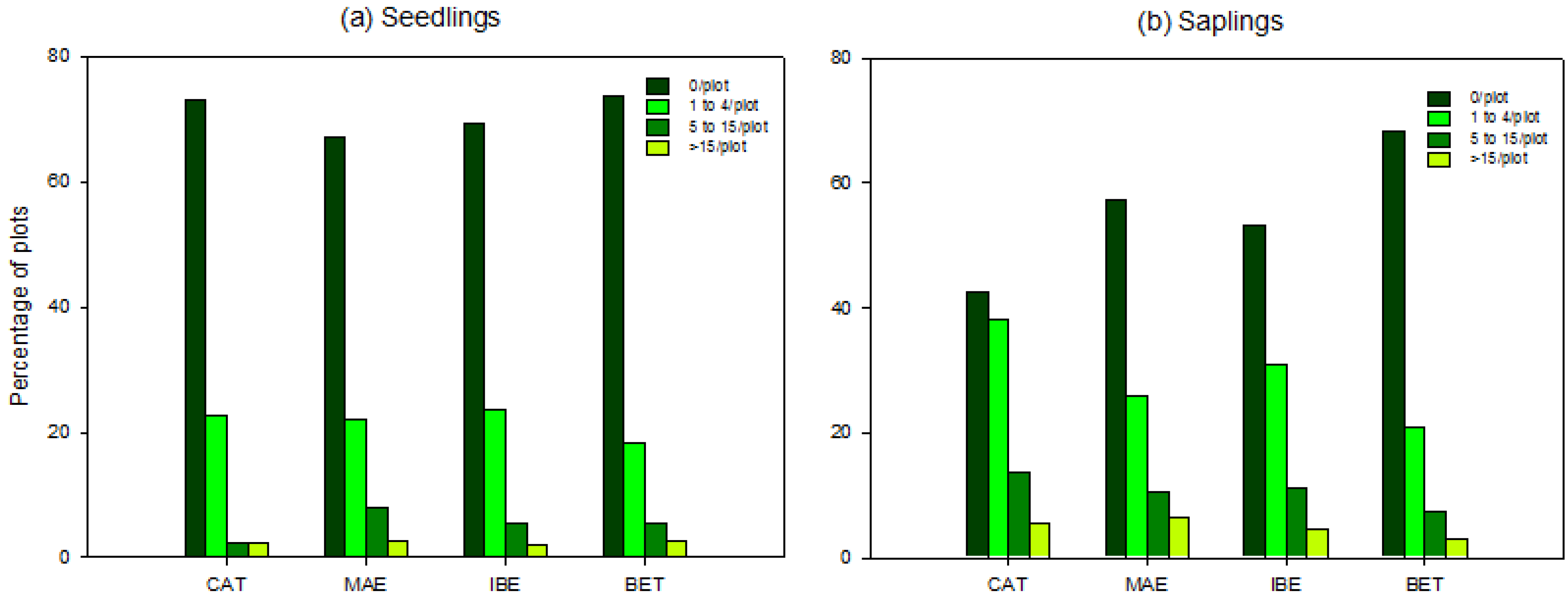
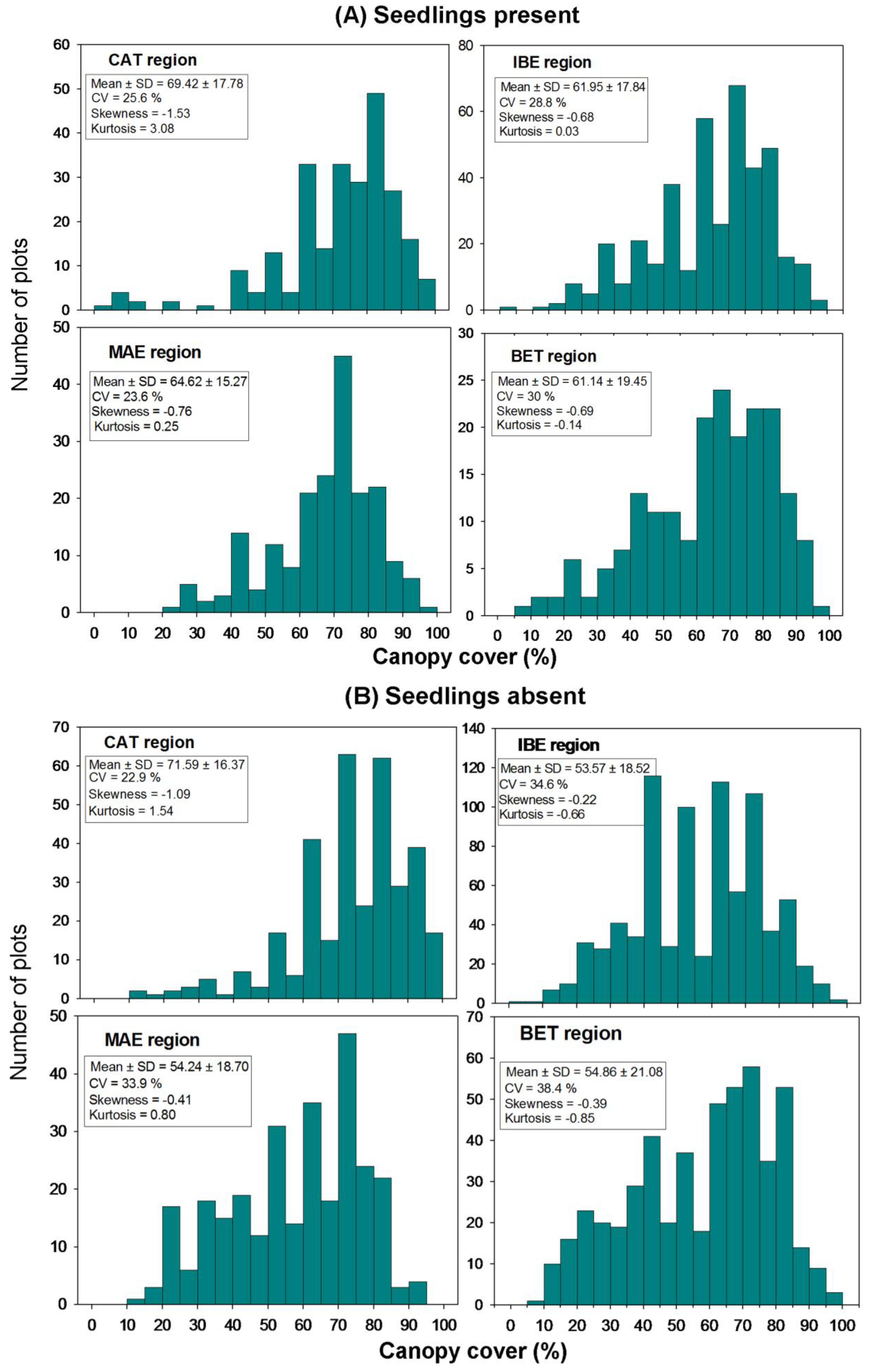
3.2. Scale of Regeneration Failure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Study Region | |||
|---|---|---|---|---|
| CAT | MAE | IBE | BET | |
| Summer precipitation (mm) | 187 ± 1.8(a) | 134 ± 0.8(b) | 103 ± 0.8(c) | 71 ± 0.7(d) |
| Mean summer temperature (°) | 19.7 ± 0.1(a) | 18.2 ± 0.1(b) | 19.7 ± 0.1(a) | 20.0 ± 0.1(a) |
| Aridity Index | 0.51 ± 0.01(a) | 0.41 ± 0.00(b) | 0.29 ± 0.00(c) | 0.20 ± 0.00(d) |
| Altitude (m) | 828 ± 15(d) | 1269 ± 17(b) | 1174 ± 12(c) | 1405 ± 13(a) |
| Slope (°) | 9.19 ± 0.02(a) | 7.45 ± 0.21(b) | 5.36 ± 0.21(c) | 8.58 ± 0.20(a) |
| Rockiness | 1.73 ± 0.04(d) | 2.70 ± 0.05(b) | 2.36 ± 0.05(c) | 2.96 ± 0.04(a) |
| Soil texture *** | 2.26 ± 0.03 | 2.14 ± 0.02 | 2.26 ± 0.03 | 2.21 ± 0.03 |
| Organic Matter content *** | 2.11 ± 0.02 | 2.06 ± 0.02 | 2.10 ± 0.03 | 1.89 ± 0.03 |
| Canopy cover (%) | 69.9 ± 0.9(a) | 60.6 ± 0.9(b) | 58.2 ± 0.9(b) | 61.0 ± 1.1(b) |
| Shrub cover (%) | 38.6 ± 1.4(b) | 37.6 ± 1.5(b) | 51.7 ± 1.7(a) | 23.6 ± 1.2(c) |
| Basal Area (m2 ha−1) | 17.6 ± 0.5(bc) | 15.8 ± 0.6(cd) | 13.4 ± 0.6(d) | 19.3 ± 0.6(ab) |
| Litter depth (cm) | 3.09 ± 0.10(a) | 1.50 ± 0.06(c) | 2.04 ± 0.10(b) | 2.01 ± 0.08(b) |
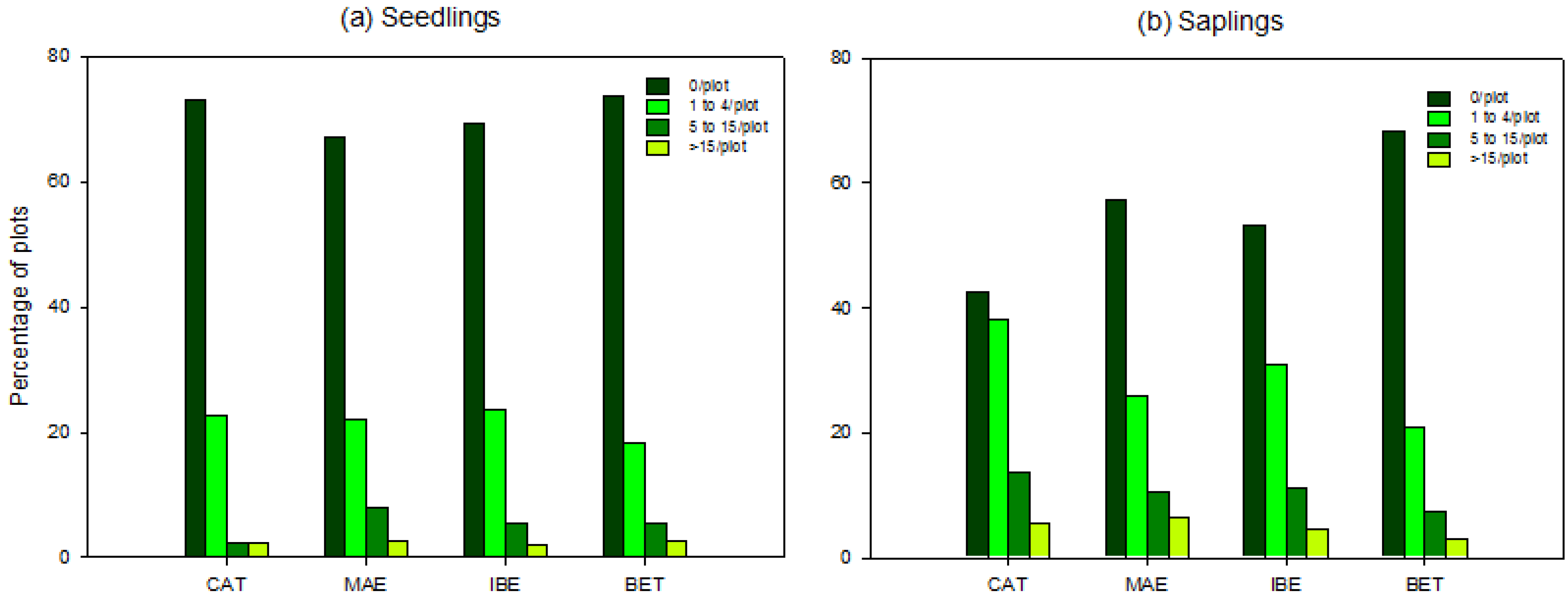
3.3. Biotic and Abiotic Factors Determining Regeneration
| Variables | Region | ||||
|---|---|---|---|---|---|
| CAT | MAE | IBE | BET | All | |
| Canopy cover | −0.050 | 0.265 *** | 0.206 ** | 0.105 | 0.125 *** |
| Shrub cover | 0.066 | −0.067 | −0.004 | 0.391 *** | 0.084 * |
| Stand basal area | −0.130 | 0.244 *** | 0.256 *** | 0.194 ** | 0.145 *** |
| Litter depth | −0.042 | 0.138 | 0.170 * | −0.047 | 0.041 |
| Rockiness | −0.008 | −0.201 ** | −0.116 | −0.178 ** | −0.109 *** |
| Soil texture | 0.114 | −0.001 | −0.025 | −0.143 * | −0.018 |
| Organic Matter | 0.067 | −0.019 | 0.101 | 0.052 | 0.050 |
| Aspect | 0.010 | −0.049 | 0.057 | 0.049 | 0.017 |
| Slope | −0.019 | 0.009 | −0.022 | 0.006 | −0.009 |
| Aridity Index | −0.114 | −0.016 | −0.106 | −0.006 | −0.019 |
| Region | Seedlings | Classes of canopy cover | |||
|---|---|---|---|---|---|
| Low | Moderate | High | Total | ||
| CAT | Present | 21 | 147 | 42 | 210 |
| Absent | 104 | 343 | 114 | 561 | |
| MAE | Present | 32 | 124 | 15 | 171 |
| Absent | 144 | 192 | 12 | 348 | |
| IBE | Present | 98 | 232 | 31 | 361 |
| Absent | 409 | 448 | 40 | 897 | |
| BET | Present | 51 | 101 | 19 | 171 |
| Absent | 193 | 290 | 24 | 507 | |
| Total | 1052 | 1877 | 297 | 3226 | |
| df | Deviance | Resid. df | Resid. Dev. | P(χ2 test) | |
|---|---|---|---|---|---|
| Null | 1359 | 1282.4 | |||
| Canopy cover poly (2) | 2 | 42.91 | 1357 | 1239.5 | 0.00000 |
| Rockiness | 1 | 13.48 | 1353 | 1205.8 | 0.00024 |
| Shrub cover poly (2) | 2 | 10.39 | 1355 | 1229.1 | 0.00553 |
| Basal Area | 1 | 9.85 | 1354 | 1219.2 | 0.00170 |
| Aridity | 1 | 5.18 | 1352 | 1200.6 | 0.02284 |
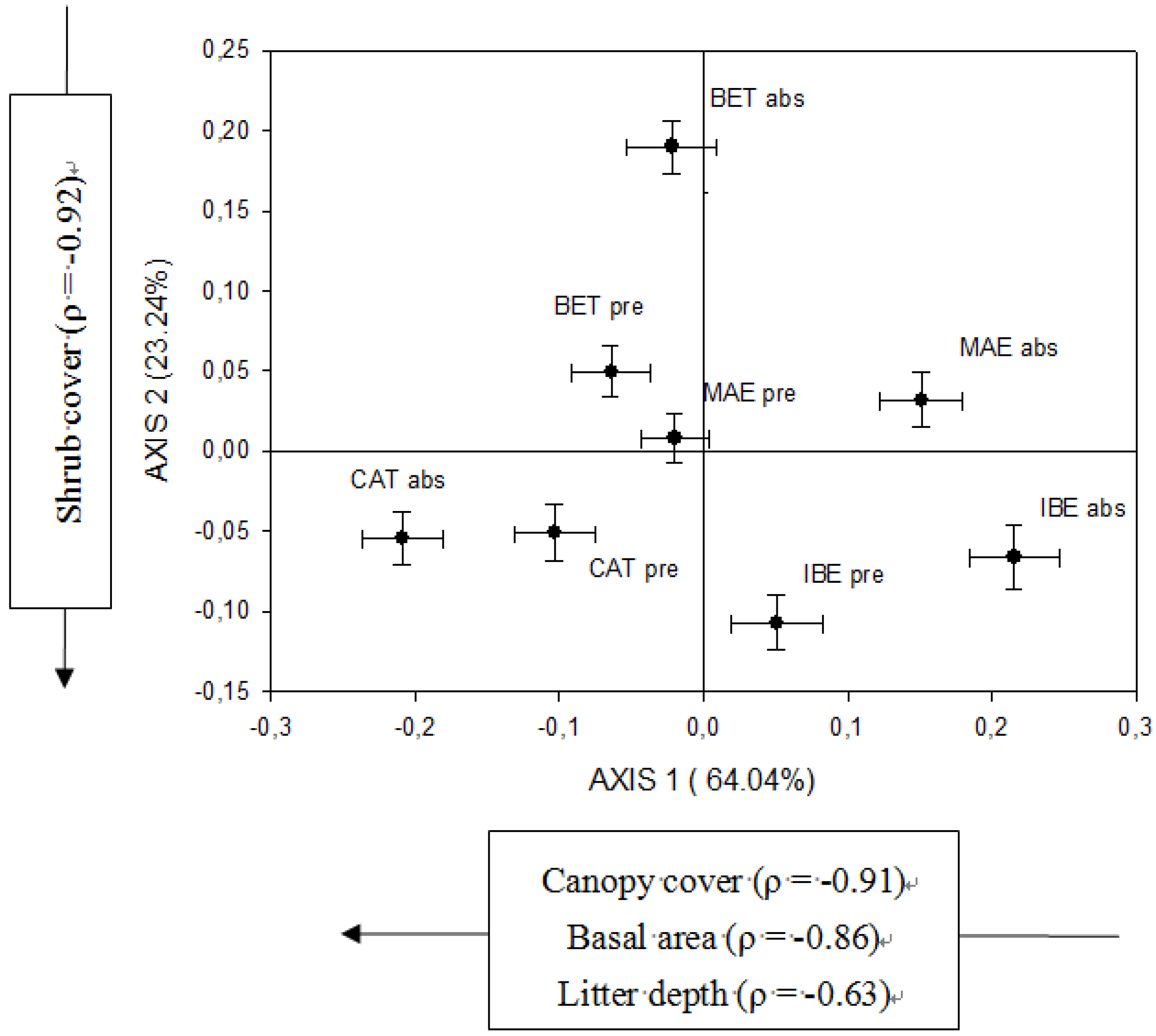
3.4. Regional Patterns of Stand-Structural Effects on Regeneration

4. Discussion
4.1. Scale of Regeneration Failure
4.2. Biotic and Abiotic Factors Determining Regeneration
4.3. Patterns of Facilitation across P. nigra Distribution Area
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Herrera, C.M.; Jordano, P.; López-Soria, L.; Amat, J.A. Recruitment of a mast-fruiting, bird-dispersed tree: Bridging frugivore activity and seedling establishment. Ecol. Monogr. 1994, 64, 315–344. [Google Scholar] [CrossRef] [Green Version]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Seedling establishment of a boreal tree species (Pinus sylvestris) at its southernmost distribution limit: Consequences of being in a marginal Mediterranean habitat. J. Ecol. 2004, 92, 266–277. [Google Scholar] [CrossRef]
- Rey, P.J.; Alcántara, J. Recruitment dynamics of a fleshy-fruited plant (Olea europaea): Connecting patterns of seed dispersal to seedling establishment. J. Ecol. 2000, 88, 622–633. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Alleviation of summer drought boosts establishment success of Pinus sylvestris in a Mediterranean mountain: An experimental approach. Plant Ecol. 2005, 181, 191–202. [Google Scholar] [CrossRef]
- Granda, E.; Escudero, A.; de la Cruz, M.; Valladares, F. Juvenile-adult tree associations in a continental Mediterranean ecosystem: No evidence for sustained and general facilitation at increased aridity. J. Veg. Sci. 2012, 23, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Tíscar, P.A.; Linares, J.C. Structure and regeneration patterns of Pinus nigra ssp. salzmannii natural forests: A basic knowledge for adaptive management in a changing climate. Forests 2011, 4, 1013–1030. [Google Scholar]
- Valladares, F.; Zaragoza-Castells, J.; Sánchez-Gómez, D.; Matesanz, S.; Alonso, B.; Portsmuth, A. Is shade beneficial for Mediterranean shrubs experiencing periods of extreme drought and late-winter frosts? Ann. Bot. 2008, 102, 923–933. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Zamora, R.; Castro, J.; Hódar, J.A. Facilitation of tree saplings by nurse plants: Microhabitat amelioration or protection against herbivores? J. Veg. Sci. 2008, 19, 161–172. [Google Scholar] [CrossRef]
- Tíscar, P.A. Regeneration dynamics of Pinus nigra subsp. salzmannii in its southern distribution area: Stages, processes and implied factors. For. Syst. 2007, 16, 124–135. [Google Scholar]
- Mendoza, I.; Zamora, R.; Castro, J. A seeding experiment for testing tree-community recruitment under variable environments: Implications for forest regeneration and conservation in Mediterranean habitats. Biol. Conserv. 2009, 149, 1491–1499. [Google Scholar] [CrossRef]
- Lookingbill, T.; Zavala, M.A. Spatial pattern of Quercus ilex and Quercus pubescens recruitment in Pinus halepensis dominated woodlands. J. Veg. Sci. 2000, 11, 607–612. [Google Scholar] [CrossRef]
- Callaway, R.M. Positive interactions among plants. Bot Rev. 1995, 61, 306–349. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R.M. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Zamora, R.; Gómez, J.M.; Hódar, J.A.; Castro, J.; Baraza, E. Applying plant facilitation to forest restoration: A meta-analysis of the use of shrubs as nurse plants. Ecol. Appl. 2004, 14, 1128–1238. [Google Scholar]
- Sthultz, C.M.; Gehring, C.A.; Whitham, T.G. Shifts from competition to facilitation between a foundation tree and a pioneer shrub across spatial and temporal scales in a semiarid woodland. New Phytol. 2007, 173, 135–145. [Google Scholar] [CrossRef]
- Lucas-Borja, M.L.; Fidalgo, T.; Lousada, J.L.; Silva-Santos, P.; Martínez García, E.; Andrés, M. Natural regeneration of Spanish black pine [Pinus nigra Arn. ssp. salzmannii (Dunal) Franco] at contrasting altitudes in a Mediterranean mountain area. Ecol. Res. 2012, 27, 913–921. [Google Scholar]
- Maestre, F.T.; Valladares, F.; Reynolds, J.F. Is the change of plant-plant interactions with abiotic stress predictable? A meta-analysis of field results in arid environments. J. Ecol. 2005, 93, 748–757. [Google Scholar]
- Maestre, F.T.; Cortina, J. Are P.halepensis plantations useful as a restoration tool in semiarid Mediterranean areas? For. Ecol. Manag. 2004, 198, 303–317. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Gómez, J.M.; Hódar, J.A.; Gómez-Aparicio, L. Benefits of using shrubs as nurse plants for reforestation in Mediterranean mountains: A 4-year study. Restor. Ecol. 2004, 12, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Verdú, M.; Gómez-Aparicio, L.; Valiente-Banuet, A. Phylogenetic relatedness as a tool in restoration ecology: A meta-analysis. Proc. R. Soc. B. 2011. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.; Huston, M. A theory of the spatial and temporal dynamics of plant communities. Vegetation 1989, 83, 49–69. [Google Scholar]
- Aranda, I.; Pardos, M.; Puértoloas, J.; Jiménez, M.D.; Pardos, J.A. Water-use efficiency in cork oak (Quercus suber) is modified by the interaction of water and light availabilities. Tree Physiol. 2007, 27, 671–677. [Google Scholar] [CrossRef]
- Valladares, F.; Pearcy, R.W. Drought can be more critical in the shade than in the sun: A field study of carbon gain and photoinhibition in a Californian shrub during a dry El Niño year. Plant Cell Environ. 2002, 25, 749. [Google Scholar] [CrossRef]
- Holmgren, M.; Gómez-Aparicio, L.; Quero, J.L.; Valladares, F. Non-linear effects of drought under shade: Reconciling physiological and ecological models in plant communities. Oecologia 2012, 169, 293–305. [Google Scholar] [CrossRef] [Green Version]
- Michalet, R.; Broker, R.W.; Cavieres, L.A.; Kikvidze, Z.; Lortie, C.J.; Pugnaire, F.I.; Valiente-Banuet, A.; Callaway, R.M. Do biotic interactions shape both sides of the humped-back model of species richness in plant communities? Ecol. Lett. 2006, 9, 767–773. [Google Scholar]
- Linares, J.C.; Tíscar, P.A. Climate change impacts and vulnerability of the southern populations of Pinus nigra subsp. salzmannii. Tree Physiol. 2010, 30, 795–806. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Fonseca, T.; Parresol, B.R.; Silva-Santos, P.; García-Morote, F.A.; Tíscar, P. Modelling Spanish black pine seedling emergence: Establishing management strategies for endangered forest areas. For. Ecol. Manag. 2011, 262, 195–202. [Google Scholar] [CrossRef]
- Tíscar, P.A.; Lucas-Borja, M.E.; Candel-Pérez, D. Changes in the structure and composition of two Pinus nigra subsp. salzmannii forests over a century of different silvicultural treatments. For. Syst. 2011, 3, 525–535. [Google Scholar]
- Coll, M.; Peñuelas, J.; Ninyerola, M.; Pons, X.; Carnicer, J. Multivariate effect gradients driving forest demographic responses in the Iberian Peninsula. For. Ecol. Manag. 2013, 303, 195–209. [Google Scholar]
- Cerro, A.; Lucas-Borja, M.E.; Martínez, E.; López, F.R.; Andrés, M.; García, F.A.; Navarro, R. Influence of stand density and soil treatment on the Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii) regeneration in Spain. Investi. Agro. Sistemas y Recursos Forest. 2009, 18, 167–180. [Google Scholar]
- Tíscar, P.A. Conditions and limits to the natural regeneration of a mountain pine forest (Pinus nigra subsp. salzmannii). For. Syst. 2003, 12, 55–64. [Google Scholar]
- Jennings, S.B.; Brown, N.D.; Sheil, D. Assessing forest canopies and understory illumination: Canopy closure, canopy cover and other measures. Forestry 1999, 72, 59–73. [Google Scholar]
- Marchi, A.; Paletto, A. Relationship between forest canopy and natural regeneration in the subalpine spruce-larch forest (north-east Italy). Folia For. Pol. 2010, 52, 3–12. [Google Scholar]
- Castedo-Dorado, F.; Gómez-Vázquez, I.; Fernandes, P.M.; Crecente-Campo, F. Shrub fuel characteristics estimated from overstory variables in NW Spain pine stands. For. Ecol. Manag. 2012, 275, 130–141. [Google Scholar]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- United Nations Environment Programme (UNEP). World Atlas of Desertification, 2nd ed.; United Nations Environment Programme: London, UK, 1997. [Google Scholar]
- Gonzalo, J. Diagnosis Fitoclimática de la España Peninsular.CD con Bases de Datos Climáticas y Fitoclimaticos; Ministerio de Medio Ambiente, Rural y Marino.: Madrid, Spain, 2010. [Google Scholar]
- Ninyerola, M.; Pons, X.; Roure, J.M. Atlas Climático Digital de la Península Ibérica. Metodología y Aplicaciones en Bioclimatología y Geobotánica. Available online: http://opengis.uab.es/wms/iberia/index.htm (accessed on 1 July 2011).
- Tíscar, P.A. Estructura, Regeneración y Crecimiento de Pinus Nigra en el Área de Reserva Navahondona-Guadahornillos (Sierra de Cazorla, Jaén). Ph.D. Thesis, Polytechnic University of Madrid, Madrid, Spain, 2004. [Google Scholar]
- Shaw, R.G.; Mitchell-Olds, T. Anova for unbalanced data: An overview. Ecology 1993, 74, 1638–1645. [Google Scholar]
- Bolker, B. Ecological Models and Data in R; Princenton University Press: Princenton, NJ, USA, 2008. [Google Scholar]
- Ruiz-Benito, P.; Gómez-Aparicio, L.; Zavala, M.A. Large-scale assessment of regeneration and diversity in Mediterranean planted pine forests along ecological gradients. Divers. Distribut. 2012, 18, 1092–1106. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Aust. Ecol. 2001, 26, 32–46. [Google Scholar]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology. Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997; p. 522. [Google Scholar]
- Anderson, M.J.; Robinson, J. Generalised discriminant analysis based on distances. Aust. N. Zeal. J. Stat. 2003, 45, 301–318. [Google Scholar]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: A useful method of constrained ordination for ecology. Ecology 2003, 84, 511–552. [Google Scholar] [CrossRef]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–871. [Google Scholar] [CrossRef]
- Linares, J.C. Shifting limiting factors for population dynamics and conservation status of the endangered English yew (Taxus baccata L., Taxaceae). For. Ecol. Manag. 2013, 291, 119–127. [Google Scholar] [CrossRef]
- Alejano, R.; González, J.M.; Serrada, R. Selvicultura de Pinus nigra Arn. subsp. salzmannii (Dunal) Franco. In Compendio de Selvicultura Aplicada en España; Serrada, R., Montero, G., Reque, J.A., Eds.; INIA: Madrid, Spain, 2008; pp. 313–356. [Google Scholar]
- Lucas-Borja, M.E.; Fonseca, T.; Linares, J.C.; García-Morote, F.A.; López-Serrano, F.R. Does the recruitment pattern of Spanish black pine (Pinus nigra Arn ssp. salzmannii) change the regeneration niche over the early life cycle of individuals? For. Ecol. Manag. 2012, 284, 93–99. [Google Scholar] [CrossRef]
- González, J.M. Modelos de transformación de masa regular a irregular. For. Syst. 2000, 9, 237–252. [Google Scholar]
- Gómez-Aparicio, L.; Valladares, F.; Zamora, R. Differential light responses of Mediterranean tree saplings: Linking ecophysiology with regeneration niche in four co-occurring species. Tree Physiol. 2006, 26, 947–958. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Zavala, M.A.; Bonet, F.J.; Zamora., R. Are Pine plantations valid tools for restoring Mediterranean forests? An assessment along abiotic and biotic gradients. Ecol. Appl. 2009, 19, 2124–2141. [Google Scholar] [CrossRef] [Green Version]
- Tíscar, P.A.; Lucas-Borja, M.E. Seed mass variation, germination time and seedling performance in a population of Pinus nigra subsp. salzmannii. For. Syst. 2010, 19, 344–353. [Google Scholar]
- Michalet, R.; Bagousse-Pinguet, Y.L.; Maalouf, J.P.; Lortie, C.J. Two alternatives to the stress-gradient hypothesis at the edge of life: The collapse of facilitation and the switch from facilitation to competition. J. Veg. Sci. 2013. [Google Scholar] [CrossRef]
- Plieninger, T.; Rolo, V.; Moreno, G. Large-scale patterns of Quercus ilex, Quercus Suber, and Quercus pyrenaica regeneration in central-western Spain. Ecosystems 2010, 13, 644–660. [Google Scholar] [CrossRef]
- Siles, G.; Rey, P.J.; Alcántara, J.M.; Ramírez, J.M. Assessing the long-term contribution of nurse plants to restoration of Mediterranean forests through Markovian models. J. Appl. Ecol. 2008, 45, 1790–1798. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tíscar, P.A.; Linares, J.C. Large-Scale Regeneration Patterns of Pinus nigra Subsp. salzmannii: Poor Evidence of Increasing Facilitation Across a Drought Gradient. Forests 2014, 5, 1-20. https://doi.org/10.3390/f5010001
Tíscar PA, Linares JC. Large-Scale Regeneration Patterns of Pinus nigra Subsp. salzmannii: Poor Evidence of Increasing Facilitation Across a Drought Gradient. Forests. 2014; 5(1):1-20. https://doi.org/10.3390/f5010001
Chicago/Turabian StyleTíscar, Pedro Antonio, and Juan Carlos Linares. 2014. "Large-Scale Regeneration Patterns of Pinus nigra Subsp. salzmannii: Poor Evidence of Increasing Facilitation Across a Drought Gradient" Forests 5, no. 1: 1-20. https://doi.org/10.3390/f5010001