The Effect of Restoration Treatments on the Spatial Variability of Soil Processes under Longleaf Pine Trees

Abstract
:1. Introduction
2. Experimental Section
2.1. Study Sites
2.2. Sampling Design
2.3. Soil Analysis
2.4. Statistical Analysis
3. Results
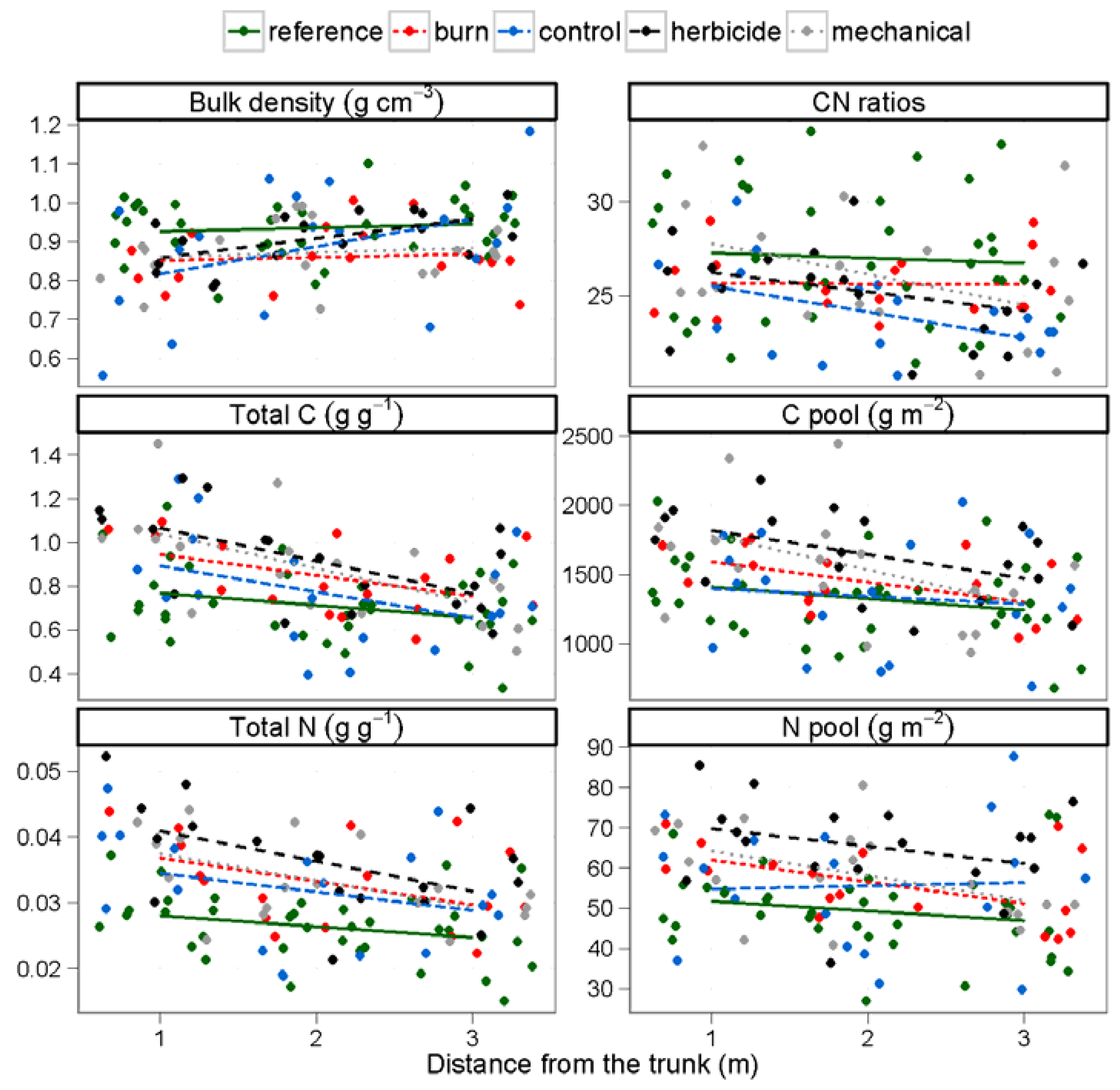
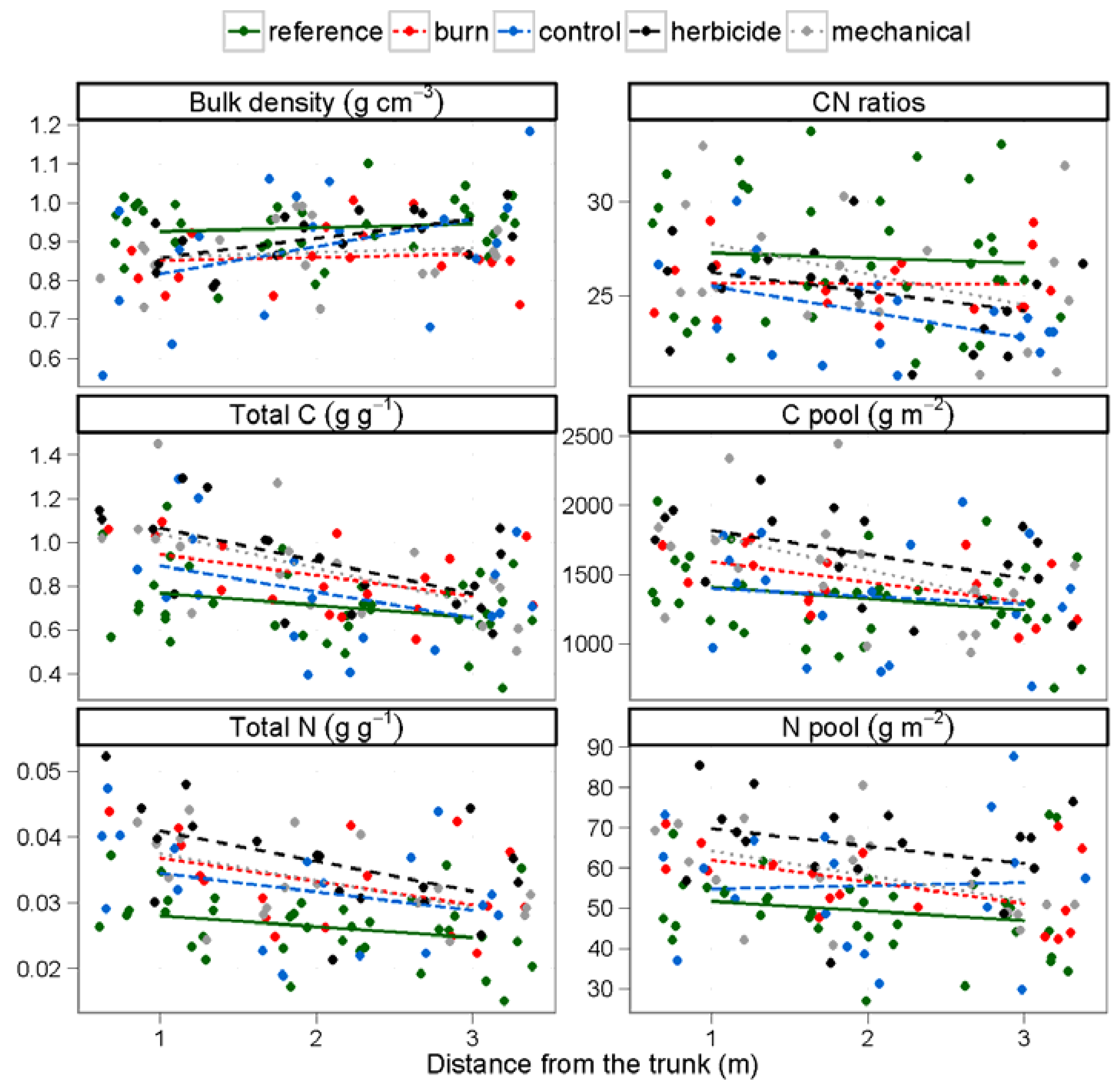
3.1. Soil Nutrients

{kind=link}
{kind=link}
{kind=link}
| Responses variables | Distance | Treatment | D × T | |||||
|---|---|---|---|---|---|---|---|---|
| F-value | p-value | F-value | p-value | F-value | p-value | |||
| Soil characteristics | Bulk Density (g cm−3) | 5.7 | 0.019 | 2.7 | 0.034 | 1.2 | 0.285 | |
| Moisture Content (%) | 0.8 | 0.387 | 1.1 | 0.379 | 0.1 | 0.988 | ||
| C (%) | 22.8 | < 0.001 | 4.9 | 0.001 | 0.9 | 0.492 | ||
| N (%) | 16.4 | < 0.001 | 9.2 | < 0.001 | 0.6 | 0.692 | ||
| CN ratio | 5.2 | 0.0244 | 3.3 | 0.014 | 0.8 | 0.545 | ||
| C pool (g m−2) | 12.4 | < 0.001 | 3.9 | 0.005 | 0.8 | 0.538 | ||
| Soil characteristics | N pool (g m−2) | 5.9 | 0.017 | 6.4 | < 0.001 | 0.8 | 0.571 | |
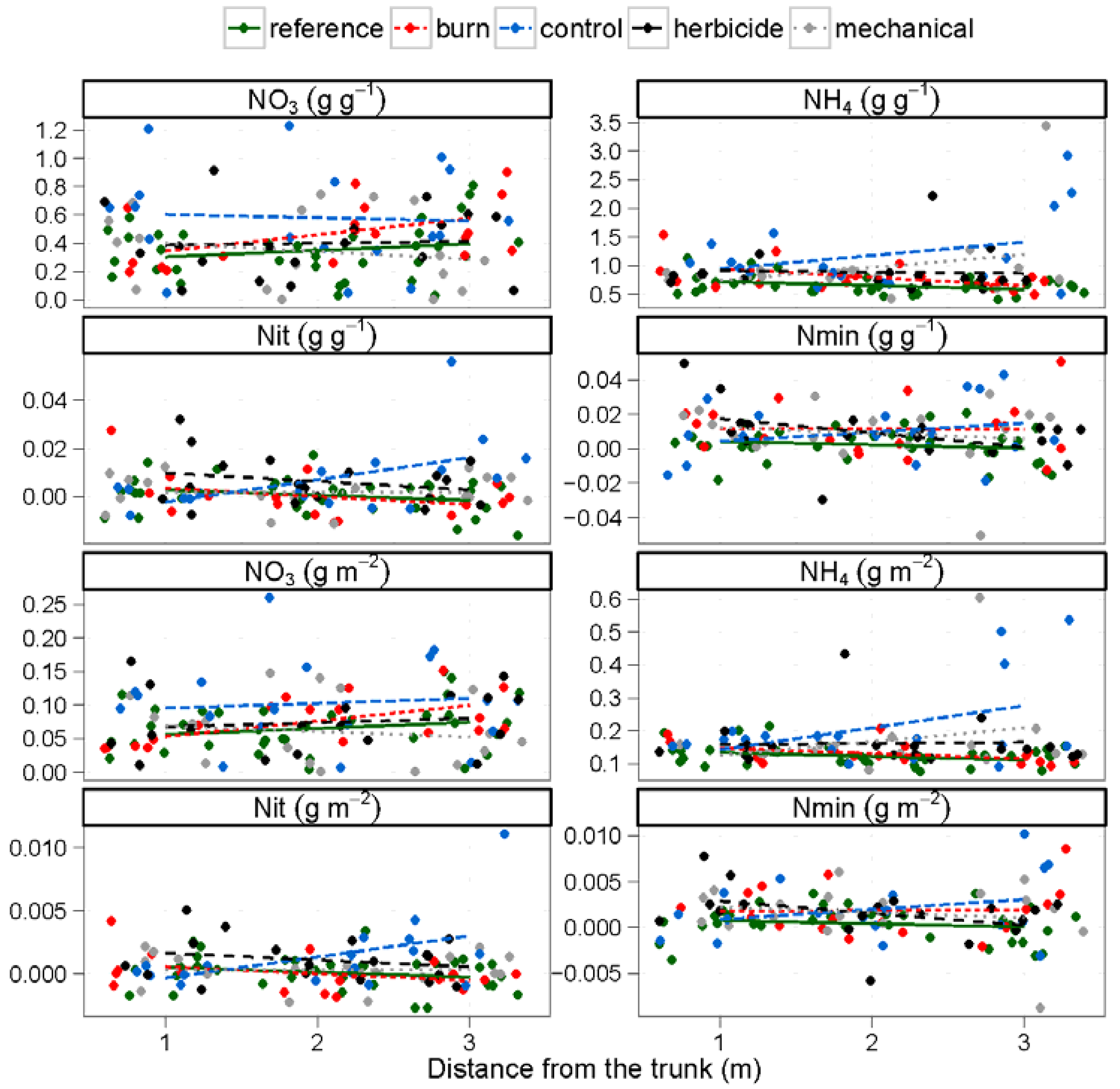
| NH4 (g N gdw−1) | 0.4 | 0.562 | 1.2 | 0.324 | 2.0 | 0.105 | ||
| NO3 (g N gdw−1) | 0.5 | 0.463 | 3.0 | 0.022 | 0.7 | 0.571 | ||
| NH4 (g m−2) | 0.1 | 0.906 | 1.0 | 0.425 | 2.4 | 0.059 | ||
| NO3 (g m−2) | 1.9 | 0.172 | 2.4 | 0.054 | 0.7 | 0.572 | ||
| Nitrification (g N gdw−1 d−1) | 0.1 | 0.719 | 2.6 | 0.041 | 4.3 | 0.003 | ||
| Mineralization (g N gdw−1 d−1) | 0.1 | 0.858 | 2.1 | 0.087 | 1.1 | 0.356 | ||
| Nitrification (g N m−2 d−1) | 0.1 | 0.834 | 2.6 | 0.038 | 4.1 | 0.004 | ||
| Mineralization (g N m−2 d−1) | 0.0 | 0.959 | 2.1 | 0.098 | 1.1 | 0.356 | ||
| Initial C flux rate (µg C gdw−1 h−1) | 0.8 | 0.361 | 0.6 | 0.641 | 0.3 | 0.897 | ||
| 6-week C flux rate (µg C gdw−1 h−1) | 0.9 | 0.352 | 1.1 | 0.353 | 1.3 | 0.268 | ||
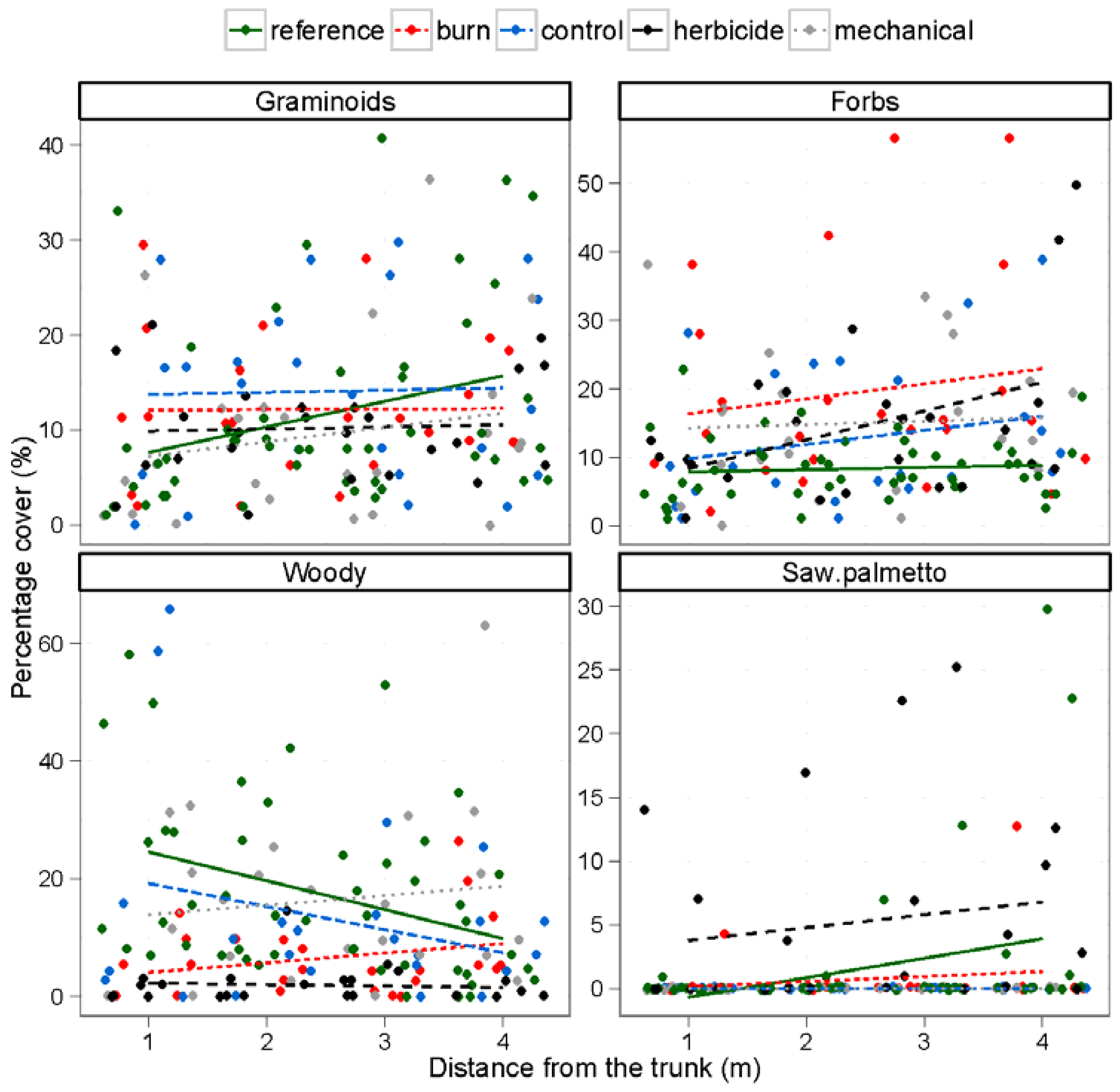
| Plant functional groups | Graminoids (%) | 3.5 | 0.062 | 1.0 | 0.417 | 0.7 | 0.561 | |
| Forbs (%) | 4.5 | 0.035 | 5.5 | < 0.001 | 0.7 | 0.571 | ||
| Woody species (%) | 3.7 | 0.057 | 8.0 | < 0.001 | 2.4 | 0.054 | ||
| Saw palmetto (%) | 4.7 | 0.003 | 5.5 | < 0.001 | 0.9 | 0.468 | ||
3.2. Nitrogen and Carbon Mineralization Rates
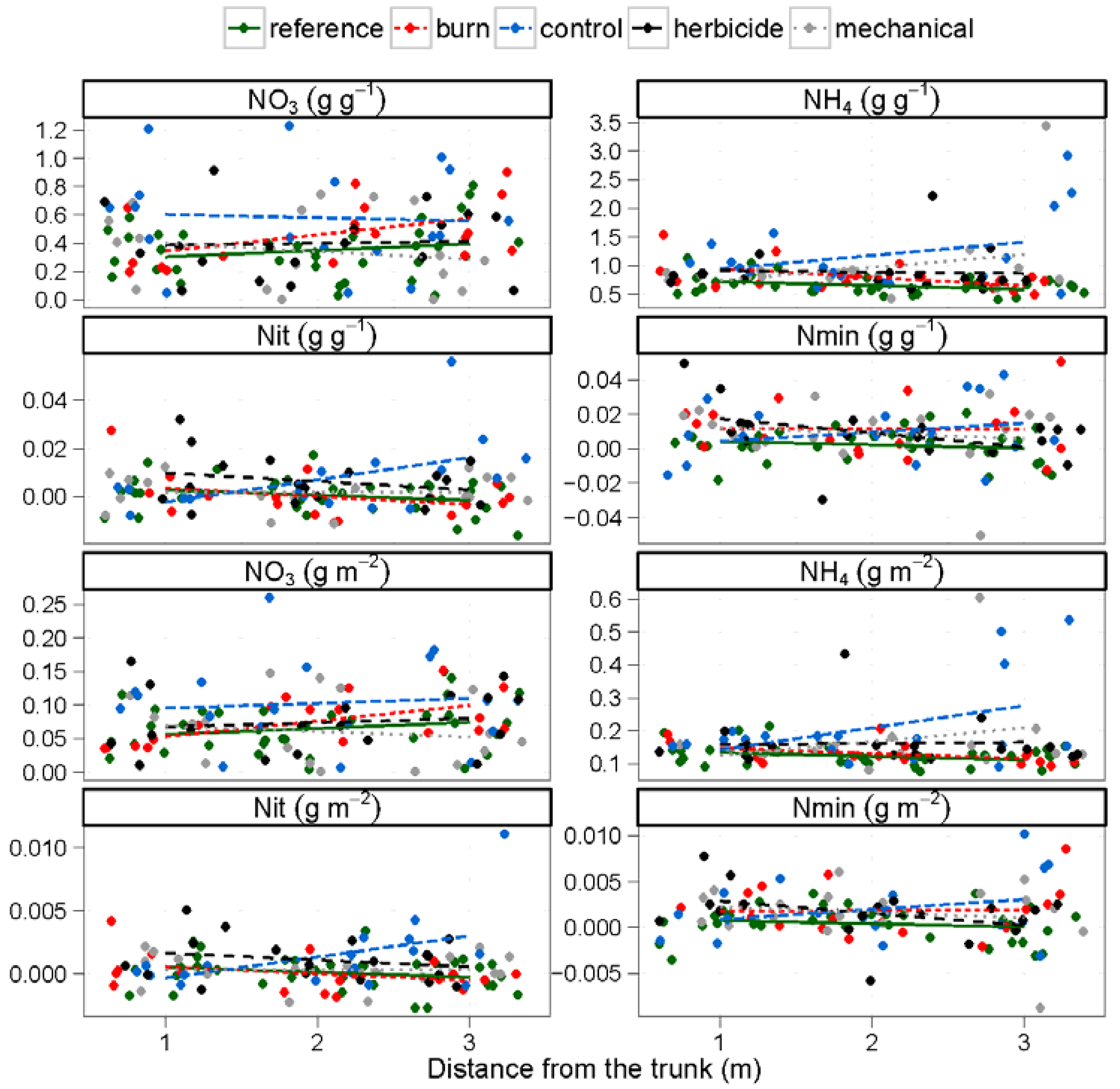
3.3. Understory Vegetation
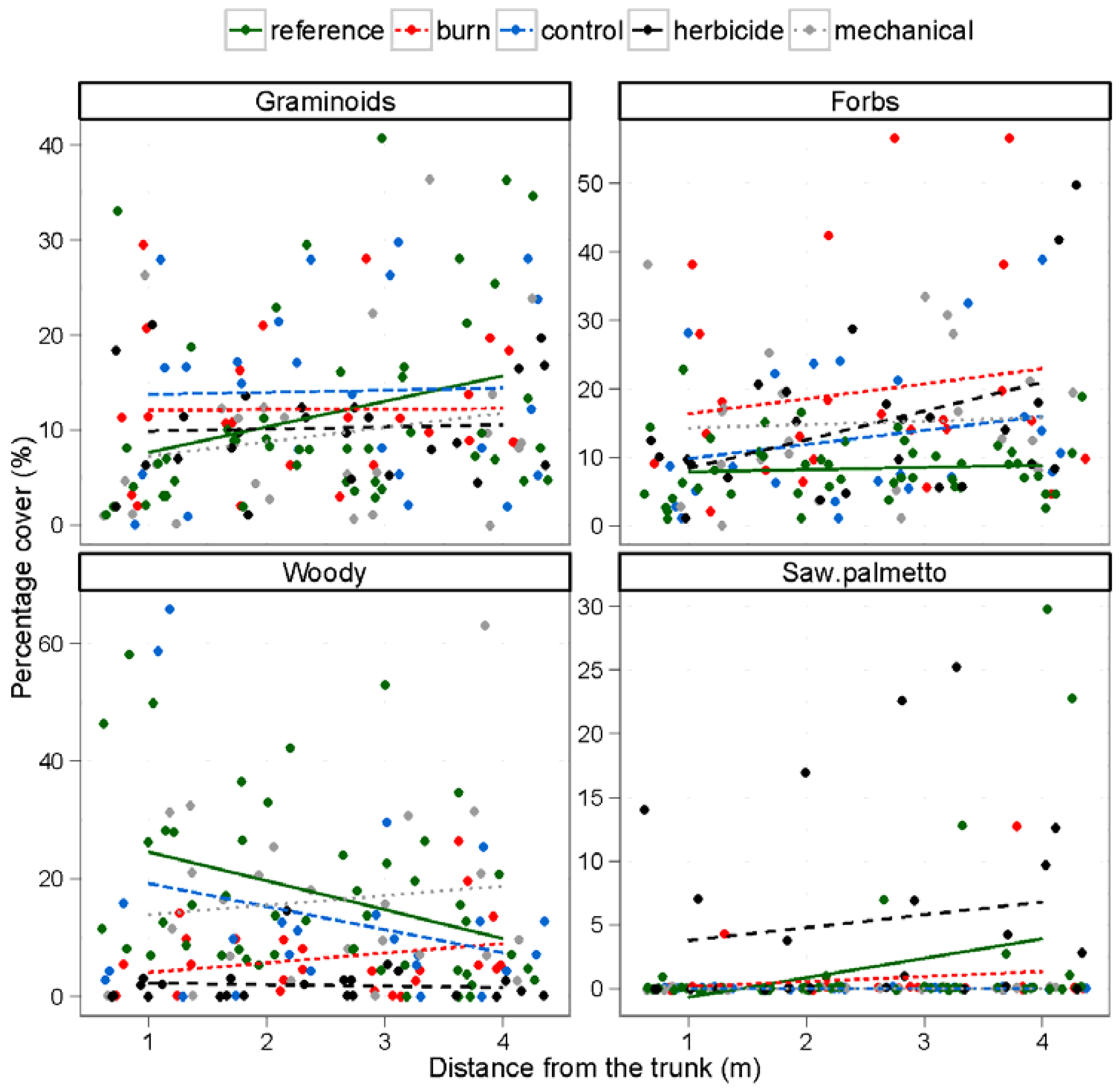
4. Discussion
4.1. Soil Gradient
4.2. Vegetation Gradients
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Zinke, P.J. The pattern of influence of individual forest trees on soil properties. Ecology 1962, 43, 130–133. [Google Scholar] [CrossRef]
- Rhoades, C.C. Single-tree influence on soil properties in agroforestry: Lessons from natural forest and savanna ecosystems. Agrofor. Syst. 1997, 35, 71–94. [Google Scholar] [CrossRef]
- Saestre, P. Spatial patterns of ground vegetation, soil microbial biomass and activity in a mixed spruce-birch stand. Ecography 1999, 22, 183–192. [Google Scholar] [CrossRef]
- Ludvig, F.; de Kroon, H.; Berendse, F.; Prins, H.H.T. The influence of savanna trees on nutrient, water and light availability and the understorey vegetation. Plant Ecol. 2004, 170, 93–105. [Google Scholar] [CrossRef]
- Godefroid, S.; Koedan, N. Tree-induced soil compaction in forest ecosystems: Myth or reality? Eur. J. For. Res. 2010, 129, 209–217. [Google Scholar]
- Weber, P.; Bardgett, R.D. Influence of single trees on spatial and temporal patterns of belowground properties in native pine forest. Soil Biol. Biochem. 2011, 43, 1372–1378. [Google Scholar] [CrossRef]
- Ko, L.J.; Reich, P.B. Oak tree effects on soil and herbaceous vegetation in savannas and pastures in Wisconsin. Am. Midl. Nat. 1993, 130, 31–42. [Google Scholar] [CrossRef]
- Tang, J.; Baldocchi, D.D. Spatial-temporal variation in soil respiration in an oak-grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components. Biogeochemistry 2005, 73, 183–207. [Google Scholar] [CrossRef]
- Mlambo, D.; Nyathi, P.; Mapaure, I. Influence of Colophospermum mopane on surface soil properties and understorey vegetation in a southern African savanna. For. Ecol. Manag. 2005, 212, 394–404. [Google Scholar] [CrossRef]
- Weltzin, J.F.; Coughenour, M.B. Savanna tree influence on understory vegetation and soil nutrients in northewestern Kenya. J. Veg. Sci. 1990, 1, 325–332. [Google Scholar] [CrossRef]
- Amiotti, N.M.; Zalba, P.; Sanchez, L.F.; Peinemann, N. The impact of single trees on properties of loess-derived grassland soils in Argentina. Ecology 2000, 81, 3283–3290. [Google Scholar]
- Penne, C.; Ahrends, B.; Deurer, M.; Bottcher, J. The impact of the canopy structure on the spatial variability in forest floor carbon stocks. Geoderma 2010, 158, 282–297. [Google Scholar] [CrossRef]
- Lodhi, M.A.K. The influence and comparison of individual forest trees on soil properties and possible inhibition of nitrification due to intact vegetation. Am. J. Bot. 1977, 64, 260–264. [Google Scholar] [CrossRef]
- Ford, E.D.; Deans, J.D. The effects of canopy structure on stemflow, throughfall and interception loss in a young Sitka spruce plantation. J. Appl. For. 1978, 15, 905–917. [Google Scholar]
- Seiler, J.; Matzner, E. Spatial variability of throughfall chemistry and selected soil properties as influenced by stem distance in a mature Norway spruce (Picea abies, Karst.) stand. Plant Soil 1995, 176, 139–147. [Google Scholar] [CrossRef]
- Battaglia, M.A.; Mou, P.; Palik, B.; Mitchell, R.J. The effect of spatially variable overstory on the understory light environment of an open-canopied longleaf pine forest. Can. J. Forest Res. 2002, 32, 1984–1991. [Google Scholar] [CrossRef]
- Palik, B.; Mitchell, R.J.; Pecot, S.; Battaglia, M.; Pu, M. Spatial distribution of overstory retention influences resources and growth of longleaf pine seedlings. Ecol. Appl. 2003, 13, 674–686. [Google Scholar] [CrossRef]
- Pecot, S.D.; Horsley, S.B.; Battaglia, M.A.; Mitchell, R.J. The influence of canopy, sky condition, and soilar angle on light quality in a longleaf pine woodland. Can. J. For. Res. 2005, 35, 1356–1366. [Google Scholar] [CrossRef]
- Brockway, D. Gap-phase regeneration in longleaf pine wiregrass ecosystems. For. Ecol. Manag. 1998, 106, 125–139. [Google Scholar] [CrossRef]
- Noss, R.F.; LaRoe, E.T.; Scott, J.M. Endangered Ecosystems of the United States: A Preliminary Assessment of Loss and Degradation; Biological Report 28; United States Department of Interiors: Washington, DC, USA, 1995. [Google Scholar]
- Frost, C.C. History and future of the longleaf pine ecosystem. In The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration; Jose, J., Jokela, E.J., Miller, D.L., Eds.; Springer: Berlin, Germany, 2006; pp. 9–48. [Google Scholar]
- Varner, J.M., III.; Gordon, D.R.; Putz, F.E.; Hiers, J.K. Restoring fire to long-unburned Pinus palustris ecosystems: Novel fire effects and consequences for long-unburned ecosystems. Restor. Ecol. 2005, 13, 536–544. [Google Scholar] [CrossRef]
- Provencher, L.; Herring, B.J.; Gordon, D.R.; Rodgers, H.L.; Galley, K.E.M.; Tanner, G.W.; Hardesty, J.L.; Brennan, L.A. Effects of hardwood reduction techniques on longleaf pine sandhill vegetation in northwest Florida. Restor. Ecol. 2001, 9, 13–27. [Google Scholar] [CrossRef]
- Peet, R.K. Ecological classification of longleaf pine woodlands. In The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration; Jose, J., Jokela, E.J., Miller, D.L., Eds.; Springer: Berlin, Germany, 2006; pp. 51–93. [Google Scholar]
- Moser, W.K.; Wade, D.D. Fire exclusion as a disturbance in the temperate forests of the USA: Examples from longleaf pine forests. Scand. J. For. Res. 2005, 20, 17–26. [Google Scholar] [CrossRef]
- Brockway, D.G.; Outcalt, K.W.; Wilkins, R.N. Restoring longleaf pine wiregrass ecosystems: Plant cover, diversity and biomass following low-rate hexazinone application on Florida sandhills. For. Ecol. Manag. 1998, 103, 159–175. [Google Scholar] [CrossRef]
- McGuire, J.P.; Mitchell, R.J.; Moser, E.B.; Pecot, S.D.; Gjerstad, D.H.; Hedman, C.W. Gaps in a gappy forest: Plant resources response to tree removal in longleaf pine savannas. Can. J. For. Res. 2001, 31, 765–778. [Google Scholar]
- Provencher, L.; Herring, B.J.; Gordon, D.R.; Rodgers, H.L.; Tanner, G.W.; Hardesty, J.L.; Brennan, L.A.; Litt, A.R. Longleaf pine and oak responses to hardwood reduction techniques in fire-suppressed sandhills in northwest Florida. For. Ecol. Manag. 2001, 148, 63–77. [Google Scholar] [CrossRef]
- Brudvig, L.A.; Asbjornsen, H. The removal of woody encroachment restores biophysical gradients in Midwestern oak savannas. J. Appl. Ecol. 2009, 46, 231–240. [Google Scholar]
- Blair, B.C. Fire effects on the spatial patterns of soil resources in a Nicaraguan wet tropical forest. J. Trop. Ecol. 2005, 21, 435–444. [Google Scholar] [CrossRef]
- Rodriguez, A.; Duran, J.; Fernandez-Palacios, J.M.; Gallardo, A. Spatial variability of soil properties under Pinus canariensis canopy in two contrasting soil textures. Plant Soil 2009, 322, 139–150. [Google Scholar] [CrossRef]
- Lavoie, M.; Mack, M.C. Spatial heterogeneity of understory vegetation and soil in an Alaskan upland boreal forest fire chronosequence. Biogeochemistry 2012, 107, 227–239. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Wade, D.D.; Waldrop, T.A.; Kennard, D.K.; Achtemeir, G.L. Fire in Southern Forest Landscapes. In Southern Forest Resource Assessment: Summary Report; General Technical Report SRS-54; Wear, D.N., Greis, J.G., Eds.; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2002; pp. 607–630. [Google Scholar]
- Gagnon, J.L.; Jack, S.B. A Comparison of the Ecological Effects of Herbicide and Prescribed Fire in a Mature Longleaf Pine Forest: Response of Juvenile and Overstory Pine; General Technical Report SRS-71; US Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 2004; pp. 304–308. [Google Scholar]
- Menges, E.S.; Gordon, D.R. Should mechanical treatments and herbicides be used as fire surrogates to manage Florida’s upland? A review. Florida Sci. 2010, 73, 147–174. [Google Scholar]
- Stokes, T.A.; Samuelson, L.J.; Kush, J.S.; Farris, M.G.; Gilbert, J.C. Structure and diversity of longleaf pine (Pinus palustris Mill.) forest communities in the mountain longleaf national wildlife refuge, Northeastern Alabama. Nat. Area J. 2010, 30, 211–225. [Google Scholar] [CrossRef]
- Provencher, L.; Galley, K.E.M.; Herring, B.J.; Sheehan, J.; Gobris, N.M.; Gordon, D.R.; Tanner, G.W.; Hardesty, J.L.; Rodgers, H.L.; McAdoo, J.P.; Northrup, M.M.; McAdoo, S.J. Post-Treatment Analysis of Restoration Effects on Soils, Plants, Arthropods, and Birds in Sandhill Systems at Eglin Air Force Base, Florida; Annual Report to Natural Resources Division, Eglin Air Force Base, Niceville; FL; Public Lands Program, The Nature Conservancy: Gainesville, FL, USA, 1998. [Google Scholar]
- Overing, J.D.; Weeks, H.H.; Wilson, J.P.; Sullivan, J.; Ford, R.D. Soil Survey of Okaloosa Country, Florida; US Department of Agriculture, Natural Resource Conservation Service: Washington, DC, USA, 1995. [Google Scholar]
- Myers, R.L. Schrub and high pine. In Ecosystem of Florida; Myers, R.L., Ewel, J., Eds.; University of Florida Presses: Gainesville, FL, USA, 1990; pp. 174–193. [Google Scholar]
- Landers, J.; Boyer, W.D. An Old-Growth Definition for Upland Longleaf and South Florida Slash Pine Forests, Woodlands, and Savannas; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 1999; pp. 1–15. [Google Scholar]
- Jones, R.H.; Mitchell, R.J.; Stevens, G.N. Controls of fine root dynamics across a gradient of gap sizes in a pine woodland. Oecologia 2003, 134, 132–143. [Google Scholar] [CrossRef]
- Hendricks, J.J.; Hendricks, R.L.; Wilson, C.A.; Mitchell, R.J.; Pecot, S.D.; Guo, D. Assessing the patterns and controls of fine root dynamics: An empirical test and methodological review. J. Ecol. 2006, 94, 40–57. [Google Scholar] [CrossRef]
- Canfield, R.H. Application of the lime interception method in sampling range vegetation. J. For. 1941, 39, 388–394. [Google Scholar]
- Bauer, H.L. The statistical analysis of chaparral and other plant communities by means of transect samples. Ecology 1943, 24, 45–60. [Google Scholar] [CrossRef]
- Keeney, D.R.; Nelson, D.W. Nitrogen-inorganic forms. In Methods of Soil Analysis Part 2—Chemicaland Microbiological Properties, 2nd; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Soil Science Society of America: Madison, WI, USA, 1982; pp. 643–698. [Google Scholar]
- Amiotti, N.M.; Bravo, O.; Zalba, P.; Peinemann, N. Effect of landscape position on the acidification of loess-derived soils under Pinus radiata. Austral Ecol. 2007, 32, 534–540. [Google Scholar] [CrossRef]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveiliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Crawley, M.J. The R Book; John Wiley & Sons Ltd.: Chichester ,UK, 2007. [Google Scholar]
- R Development Core Team, R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012.
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nlme: Linear and Nonlinear Mixed Effects Models, version 3.1-104; R Development Core Team: Vienna, Austria, 2012. [Google Scholar]
- Avery, C.R.; Cohen, S.; Parker, K.C.; Kush, J.S. Spatial pattern of longleaf pine (Pinus palustris) seedling establishment on the croatan national forest, North Carolina. J. North Carol. Acad. Sci. 2004, 120, 131–142. [Google Scholar]
- Boring, L.R.; Hendricks, J.J.; Wilson, C.A.; Mitchell, R.J. Seasonal of burn and nutrient losses in a longleaf pine ecosystem. Int. J. Wildland Fire 2004, 13, 443–453. [Google Scholar] [CrossRef]
- Noble, M.M.; Dillon, W.; Mhila, M. Initial response of soil nutrient pools to prescribed burning and thinning in a managed forest ecosystems of northern Alabama. Soil Sci. Soc. Am. J. 2009, 73, 285–292. [Google Scholar]
- Rodgers, H.L.; Provencher, L. Analysis of longleaf pine sandhill vegetation in northwestern Florida. Castanea 1999, 64, 138–162. [Google Scholar]
- Wilson, C.A.; Mitchell, R.J.; Hendricks, J.J.; Boring, L.R. Patterns and control of ecosystem function in longleaf pine-wiregrass. II. Nitrogen dynamics. Can. J. For. Res. 1999, 29, 752–760. [Google Scholar] [CrossRef]
- Platt, W.J.; Carr, S.M.; Reilly, M.; Fahr, J. Pine savanna overstorey influences on ground-cover biodiversity. Appl. Veg. Sci. 2006, 9, 37–50. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lavoie, M.; Mack, M.C.; Hiers, J.K.; Pokswinski, S. The Effect of Restoration Treatments on the Spatial Variability of Soil Processes under Longleaf Pine Trees. Forests 2012, 3, 591-604. https://doi.org/10.3390/f3030591
Lavoie M, Mack MC, Hiers JK, Pokswinski S. The Effect of Restoration Treatments on the Spatial Variability of Soil Processes under Longleaf Pine Trees. Forests. 2012; 3(3):591-604. https://doi.org/10.3390/f3030591
Chicago/Turabian StyleLavoie, Martin, Michelle C. Mack, John K. Hiers, and Scott Pokswinski. 2012. "The Effect of Restoration Treatments on the Spatial Variability of Soil Processes under Longleaf Pine Trees" Forests 3, no. 3: 591-604. https://doi.org/10.3390/f3030591