Tree Compatibility, Incomplete Directed Perfect Phylogeny, and Dynamic Graph Connectivity: An Experimental Study

Department of Computer Science, Iowa State University, Ames, IA 50011, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Algorithms 2019, 12(3), 53; https://doi.org/10.3390/a12030053
Submission received: 28 December 2018
/
Revised: 20 February 2019
/
Accepted: 22 February 2019
/
Published: 28 February 2019
Abstract
:We study two problems in computational phylogenetics. The first is tree compatibility. The input is a collection of phylogenetic trees over different partially-overlapping sets of species. The goal is to find a single phylogenetic tree that displays all the evolutionary relationships implied by . The second problem is incomplete directed perfect phylogeny (IDPP). The input is a data matrix describing a collection of species by a set of characters, where some of the information is missing. The question is whether there exists a way to fill in the missing information so that the resulting matrix can be explained by a phylogenetic tree satisfying certain conditions. We explain the connection between tree compatibility and IDPP and show that a recent tree compatibility algorithm is effectively a generalization of an earlier IDPP algorithm. Both algorithms rely heavily on maintaining the connected components of a graph under a sequence of edge and vertex deletions, for which they use the dynamic connectivity data structure of Holm et al., known as HDT. We present a computational study of algorithms for tree compatibility and IDPP. We show experimentally that substituting HDT by a much simpler data structure—essentially, a single-level version of HDT—improves the performance of both of these algorithm in practice. We give partial empirical and theoretical justifications for this observation.
1. Introduction
A phylogenetic tree is a graphical depiction of the evolutionary history of a collection of taxa (typically species or genes). The leaves are in one-to-one correspondence with the taxa, the internal nodes correspond to hypothetical ancestral taxa, while edges represent ancestor-descendant relationships. Here, we consider two problems in computational phylogenetics.
- The input to the tree compatibility problem is a collection of rooted phylogenetic trees with partially-overlapping taxon sets. and the trees within it are called, respectively, a profile and the input trees. The problem is to find a tree whose taxon set is the union of the taxon sets of the input trees, such that each input tree can be obtained from the restriction of to the leaf set of through edge contraction. If such a tree exists, then is said to be compatible; otherwise, is incompatible.
- The input to the incomplete directed perfect phylogeny problem (IDPP) is an character matrix , where each row corresponds to a taxon and each column to a character. The state of taxon i on character j is zero, one, or ?, depending on whether, for species i, character j is absent, present, or the state is unknown. A completion of A is a matrix obtained from A by replacing each ? by either zero or one. IDPP asks if A has a completion B with the following property. There exists a phylogenetic tree that explains the evolution of the taxa described by B with at most one state transition on each character.
It is well known that testing the compatibility of a collection of unrooted trees—an NP-complete problem [1]—is equivalent to the undirected version of IDPP, namely the problem of testing the compatibility of a collection of “partial binary characters” (bipartitions of a subset of a set of species) [2]. Since a profile of rooted trees is effectively a collection of unrooted trees that have a common root taxon, the preceding observation establishes the connection between rooted tree compatibility and IDPP. We should also note that a reduction from rooted tree compatibility to IDPP is implicit in the work of Chimani et al. [3].
Here, for completeness, we provide a short and direct proof of the equivalence of tree compatibility and IDPP. We go further and argue that a previous algorithm for IDPP is effectively a variation of a recent tree compatibility algorithm. We then present a computational study of algorithms for these problems. This study provides an opportunity to analyze the engineering issues that arise when applying dynamic graph connectivity data structures in a highly-specific context. Our empirical results show that, in this setting, simple data structures perform better than more sophisticated ones with better asymptotic bounds.
1.1. Background
Tree compatibility testing arises in supertree construction [4,5,6,7], where the goal is to assemble a comprehensive phylogenetic tree out of smaller trees for restricted sets of taxa. The tree compatibility problem, however, has wider uses. In fact, the first polynomial-time algorithm for testing tree compatibility, the Build algorithm of Aho et al. [8], was designed to solve a problem in relational databases, not phylogenetics. The connection with phylogenetics was noted later [1].
A recent algorithm, called [9,10], solves the tree compatibility problem for an arbitrary profile in time, where denotes the total size of the trees in . is closely related to Semple and Steel’s version of Build [2] (the suffix “NT” refers to the fact that the algorithm can be extended to profiles of trees with “nested taxa”; i.e., where internal nodes are labeled with higher order species [10]). There is one important difference between the two algorithms. Build relies on the triple graph, whose nodes are the species and where there is an edge between two species a and b if they are involved in a rooted triple in some input tree; that is, if there is a third species c such that, for some input tree, the lowest common ancestor of a and b is a descendant of the lowest common ancestor of a, b, and c. In contrast, relies on the display graph of the profile, a graph that was first studied in the context of testing the compatibility of unrooted trees [11]. The display graph is closely related to the intersection graph of certain sets of clusters that appear in the input trees (a cluster is a set of species that descend from the same node). As explained in [9], this cluster-based view provides a link to Build’s triplet-based approach. It also leads to improved performance for high-degree trees.
, and essentially every other known algorithm for tree compatibility, involves maintaining the connected components of a graph under a series of edge and vertex deletions [9,12,13]. Thus, the efficiency of these algorithms depends heavily on the dynamic graph connectivity data structure used. Conversely, tree compatibility has been cited as a motivation for developing efficient dynamic graph connectivity data structures [12,14,15].
IDPP arises when building phylogenetic trees based on rare genomic changes, such as the insertion of short interspersed nuclear elements (SINEs) [13,16]. IDPP is also useful for resolving genotypes into haplotypes [17]. The algorithm of Pe’er et al. [13] solves IDPP for an matrix A in time, where m denotes the number of ones in A. This algorithm also relies on dynamic graph connectivity.
Several dynamic graph connectivity data structures have been proposed [15,18,19,20,21]. Notable among them is the data structure of Holm et al., known as HDT [15,20]. HDT allows one to maintain a graph under a sequence of vertex and edge deletions and insertions in polylogarithmic amortized time per operation. The above-mentioned time bounds for tree compatibility and IDPP are based on using HDT.
Like other dynamic connectivity data structures, HDT maintains a spanning forest of the given graph, where there is one spanning tree for each connected component. Edges in the forest are called tree edges. The deletion of a tree edge breaks a tree in two and triggers the search for an edge, called a replacement edge, to re-link the trees. To ensure polylogarithmic amortized time per update, HDT stores edges in a multi-level structure, where an edge can appear in multiple levels (see Section 2.2).
To our knowledge, there is no previously-published computational study of any tree compatibility algorithm. There is, however, a previous experimental study of HDT [22]. That paper offers insights into the implementation details and the factors that affect HDT’s performance in practice. The focus is on assessing how well HDT’s amortized bounds for updates (edge/vertex insertion/deletion) are realized in practice on non-problem-specific graphs.
1.2. Contributions
As stated earlier, one of our contributions is to elucidate the connection between tree compatibility and IDPP. Our computational study investigates the performance of the tree compatibility algorithm of [9] and the IDPP algorithm of [13] over a wide range of real and simulated input profiles. Our primary goal is to determine the impact of the underlying dynamic graph connectivity data structure, in this case HDT. In contrast to the experimental work on HDT of Iyer et al. [22], our focus is on aggregate performance over an entire sequence of edge deletions. A secondary goal is to compare the performance of the specialized IDPP algorithm against the more general tree compatibility algorithm in the context of IDPP.
Our experiments suggest that, in the specific setting of compatibility testing and IDPP, we can dispense with much of the complexity of HDT—indeed, we can go from a multi-level structure to a single-level data structure, considerably simplifying the code—and actually accelerate the compatibility testing algorithm while reducing its memory footprint.
1.3. Contents
Section 2 reviews graph and tree notation, phylogenetic trees, and the HDT data structure. Section 3 defines tree compatibility and IDPP formally and explains the relationship between the two problems. Section 4 reviews the tree compatibility algorithm of [10] and the IDPP algorithm of [13] and explains the connections between them. Section 5 and Section 6 present the results of our experiments with tree compatibility and IDPP, respectively. Section 7 delves deeper into the reasons behind the observed performance of the algorithms for these problems, focusing on the impact of dynamic connectivity testing. Section 8 gives some concluding remarks.
2. Preliminaries
For each positive integer r, denotes the set . Throughout the paper, X denotes a set of taxa.
2.1. Graphs and Phylogenetic Trees
Let G be a graph. and denote the node and edge sets of G. A tree is an acyclic connected graph. In this paper, all trees are assumed to be rooted. For a tree T, denotes the root of T. Suppose . Then, u is an ancestor of v in T, and v is a descendant of u, if u lies on the path from v to in T. If u is an ancestor of v and , then u is the parent of v and v is a child of u. The degree of a node is the number of children of u. T is binary if every non-leaf node has degree two. For , we write to denote the subtree of T rooted at u.
A phylogenetic X-tree is a pair where T is a tree in which every internal node has at least two children and is a bijection from the leaf set of T into X. For each leaf v of T, is the label of v. A rooted triple is a binary phylogenetic tree on three leaves.
Let be a phylogenetic X-tree. For each , the cluster at u, denoted by , is the set of all taxa in . denotes the set of all clusters of . The cluster and the clusters such that u is a leaf of are called trivial; all other clusters in are non-trivial. A phylogenetic X-tree is completely determined by ([2], Theorem 3.5.2). That is, if for some other phylogenetic X-tree , then and are isomorphic.
Suppose . The restriction of to Y, denoted , is the phylogenetic Y-tree whose cluster set is Equivalently, is obtained from the minimal rooted subtree of T that connects the leaves in by suppressing all non-root internal vertices of degree one.
Let be a phylogenetic X-tree and be a phylogenetic Y-tree such that . displays if .
2.2. Dynamic Graph Connectivity
2.2.1. Spanning Forests and Euler Tour Trees
Like other dynamic graph connectivity data structures [19,21], HDT maintains a spanning forest F of the given graph throughout the lifetime of this graph. Edges of F are called tree edges; all other edges are non-tree edges. Each tree T in F is represented using a Euler tour tree (ET tree) [19], a balanced binary tree over a Euler tour of T. A Euler tour of an n-node tree has nodes, and a single node of the tree may appear multiple times in the tour. ET trees support the following operations in logarithmic time per operation: determining the size of the tree containing a given node, testing if two nodes are in the same tree, linking two trees with an edge, and deleting an edge from a tree.
2.2.2. Edge Deletion in HDT
We now review how HDT handles edge deletions, focusing on the aspects that are most relevant for compatibility testing and IDPP. For further details, we refer the reader to [15].
Two cases arise when deleting an edge e. If e is a non-tree edge, the graph remains connected; no further action is needed. Handling this case takes constant time. If e is a tree edge, the tree T in F containing e is split into two trees and . Since the vertices of T may still be connected by a non-tree edge in the original graph, HDT searches for a replacement edge f to re-link and . Next, we explain how a replacement edge is found.
HDT associates with each tree or non-tree edge e an integer level . Initially, . Promoting e means increasing by one. Let denote the sub-forest of F induced by the edges with level . Thus, . HDT maintains the following invariants: (i) if we interpret the levels of the edges as their weights, then the edges of F constitute a maximum spanning forest of the graph; (ii) the number of nodes in any tree in is at most , where n is the number of nodes in the graph. Thus, (a) if is a non-tree edge, then u and v are connected in , and (b) .
Let be the tree edge to be deleted. Since F was a maximum spanning forest, e’s replacement must have level at most . We set and look for a replacement at level i as follows. Let and be the trees in that contain u and v, respectively. Assume . Before deleting , was a tree of such that . By Invariant (ii), . Thus, . For each tree edge f of , we promote f, making a tree in . Next, we scan the level-i non-tree edges incident to until we either find a replacement edge or all non-tree edges incident to have been examined. If a visited edge f reconnects and , then f is the replacement edge, and the scan stops. Otherwise, we promote f. If no replacement is found at level i, we decrease i by one and repeat the search. We stop if the search succeeds or i drops below zero (which means that no replacement edge exists).
HDT maintains each forest as a collection of ET trees. Thus, deleting an edge requires cutting it from at most ET trees, and if a replacement edge is found, this edge is used to link at most ET trees. The worst-case time for cutting and linking is therefore . The amortized cost of the edge scans can be shown to also be .
2.2.3. Level Truncation
Maintaining HDT’s multi-level structure may require several expensive dynamic memory allocation operations. Although this expense could be reduced by allocating the space for all levels in advance, this would be wasteful, since the higher levels tend to be sparsely populated.
Iyer et al. [22] showed that level truncation—i.e., putting a limit on the number of levels in the data structure—improves HDT’s performance in practice. An extreme version of this idea is to disable edge promotion entirely and use a single level: Level 0. That is, we maintain a single spanning forest F of the graph, where each tree in F is represented using an ET tree. Deleting non-tree edges is, again, trivial. Deleting a tree edge splits the tree T in F containing e into two trees and . Suppose is the smaller tree. For each node v in , we scan the edges incident on v to determine if any of them is a replacement edge. This requires two queries to the ET trees containing the endpoints of each edge. If a replacement edge f is found, we re-link and using f and stop.
In the rest of the paper, we refer to the version of HDT where edge promotion is disabled as HDT. The following observation is easy to verify.
Proposition 1.
Let G be an n-node graph. Then, HDT(0) handles any sequence of q edge deletions in G in time, where s is the total number of edge scans performed over the entire sequence.
Note that Proposition 1 holds regardless of whether one scans the edges incident on the smaller component or those incident on the larger component. We shall nevertheless assume that the smaller component is the one whose edges are scanned, since in practice, that component tends to have fewer incident non-tree edges.
3. Tree Compatibility and Incomplete Directed Perfect Phylogeny
3.1. Tree Compatibility
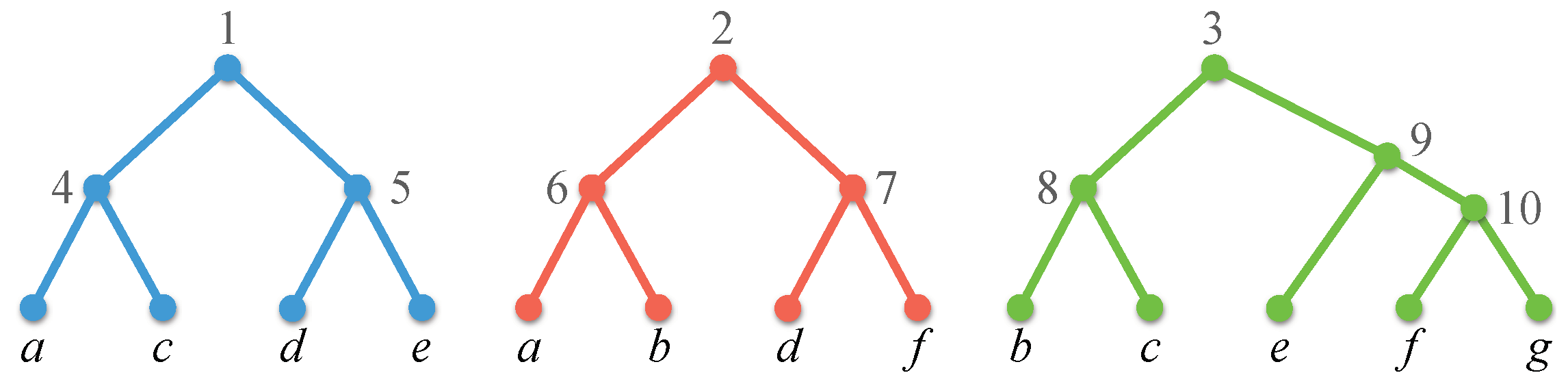
A profile on X is a set where, for each , is a phylogenetic -tree on some set , and (Figure 1). denotes , and denotes . The size of is .
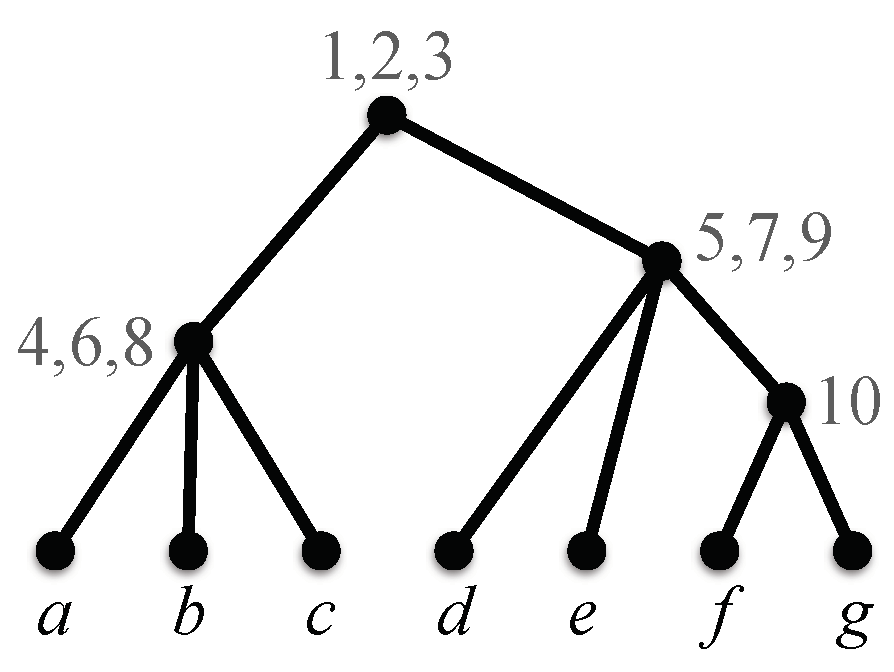
is compatible if there exists a phylogenetic X-tree such that displays , for each . Such a tree , if it exists, is a compatible supertree for (Figure 2). The problem of finding a compatible supertree for a profile, or reporting that no such supertree exists, is called the tree compatibility problem.
Profiles consisting of binary trees (and, in particular, rooted triples) are common in practice. As Figure 1 and Figure 2 show, a compatible supertree for a profile of binary trees need not itself be binary.
Suppose is a compatible supertree for profile . It follows from the definition of the notion of “displays” that, for any cluster present in some tree in , there is a corresponding cluster in . More formally, for each , there exists a mapping from to with the following property. For each , (here, we use the cluster notation of Section 2.1). We say that v maps to ; see Figure 2. Note that the mapping need not be unique.
3.2. The Display Graph
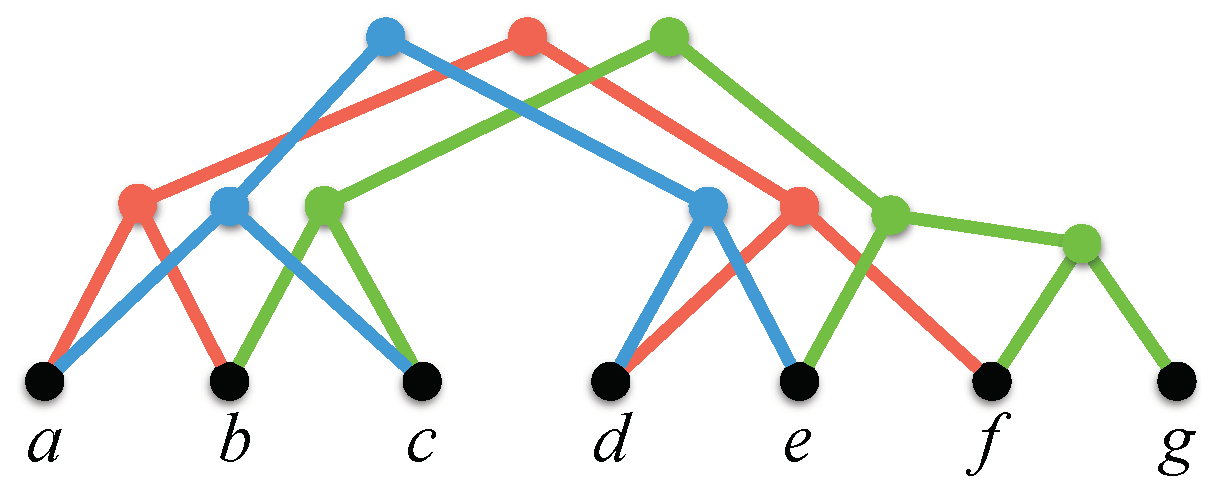
The display graph of a profile , denoted by , is the graph obtained from the disjoint union of the underlying trees by identifying leaves that have a common label (Figure 3). has nodes and edges and can be constructed in time.
We refer to a node of that results from identifying multiple leaves as a leaf of . Let v be any leaf of . The label of v is the common label of the leaves of that were identified to create v. We refer to a node v of that is not a leaf as an internal node. A node u in is a child of an internal node v if u is the child of v in some tree in .
3.3. Incomplete Directed Perfect Phylogeny
Assume , and let be a set of characters. A character matrix is an matrix , where . Entry is called the state of taxon on character . For each and each , the s-set of character is the set of taxa .
A completion of a -matrix A is a -matrix B obtained by replacing all the ?s in A by zeroes and ones (thus, for all ). A perfect phylogeny for a completion B of A is a phylogenetic X-tree such that for every . If such a tree exists, we call it a perfect phylogeny for A as well.
The input to the incomplete directed perfect phylogeny problem (IDPP) is an -matrix A. The problem is to find a perfect phylogeny for A or report that no perfect phylogeny exists.
For each , we say that column j of A is trivial if or . Let be the matrix obtained from A by striking out all trivial columns. It is straightforward to show that A has a perfect phylogeny if and only if A does. Thus, in the the rest of the paper, we assume that A contains no trivial columns.
3.4. The Relationship between Tree Compatibility and IDPP
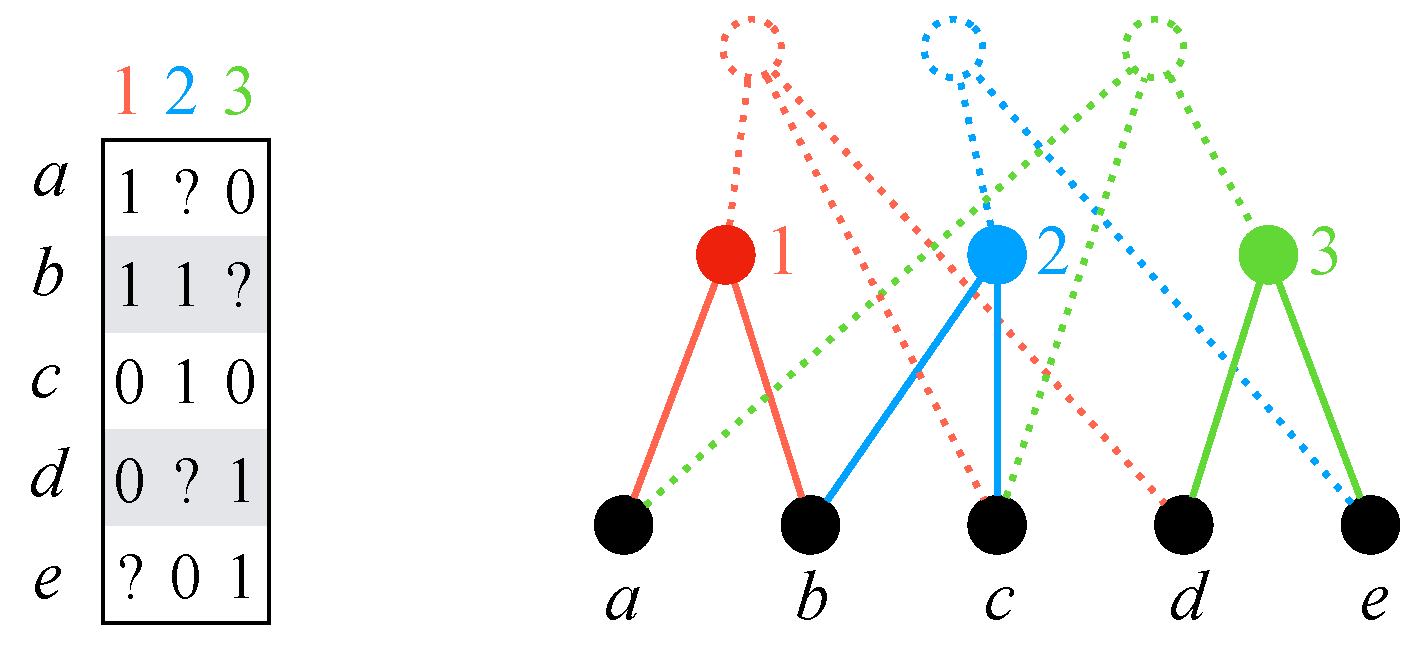
Given an instance A of IDPP, let us define a profile as follows. For each , is the phylogenetic -tree where and where contains only one non-trivial cluster: (Figure 4).
Lemma 1.
An instance A of IDPP has a perfect phylogeny if and only if is a compatible profile.
Proof.
We claim that a phylogenetic X-tree is a perfect phylogeny for A if and only if displays for each ; that is, if and only if is a compatible supertree of .
Indeed, is a perfect phylogeny for A if and only if there is a completion B of A such that for every . This holds if and only if, for each , , where . Since is the only non-trivial cluster in , this is equivalent to saying that displays . □
4. Algorithms for Tree Compatibility and Incomplete Directed Perfect Phylogeny
4.1. Tree Compatibility
(Algorithm 1) builds a compatible supertree for a profile by traversing the display graph top-down, starting from the roots of the input trees, successively decomposing into subgraphs that correspond to subtrees of the compatible supertree [10]. If it is impossible to decompose , the algorithm reports that is incompatible. To explain in more detail, we need some definitions and notation. Figure 5 illustrates several of these notions.
A position in is a vector , where , for each . For each , let , and let . Note that, since labels may be shared among trees and is obtained by identifying leaves with the same label, we may have , for with .
A position U is valid if the following holds for each .
- If , then the elements of are siblings in and
- .
For any valid position U, let denote the subgraph of induced by . Then, is the subgraph of obtained by deleting all nodes in , along with all incident edges [10].
Let U be a valid position, and let v be a vertex in U. Then, v is semi-universal in U if , for every such that . Let denote the set of semi-universal labels in U. The successor of U is the position such that, for each , if , for some , then , where denotes the set of children of v in ; otherwise, .
The root position is the position where, for each , is a singleton containing . It is obvious that is valid, that , that , and that every vertex in is semi-universal.
The key idea behind is that if is connected, then all semi-universal nodes in U can map to the same node in a compatible supertree for . The set of labels in the cluster at is precisely the set of labels that appear in . Further, each connected component of corresponds to a distinct subtree of in .
uses a first-in first-out queue Q to store pairs , where U is a valid position in and is a reference to the parent of the node corresponding to U in the supertree built so far. initializes Q to contain the starting position, , with a null parent. Each iteration of the while loop of Lines 3–15 starts by de-queuing a pair . Line 5 computes the set S of semi-universal labels in U. It can be shown that if S is empty, then is incompatible [10]. This case is handled in Lines 6–7. If S is not empty, the algorithm creates a tentative root labeled by S for the tree for and links to its parent (Line 8). If S consists of exactly one element that is a leaf in , then is a potential leaf in the output tree . We set the label of appropriately, skip the rest of the current iteration of the while loop, and continue to the next iteration (Lines 9–11). Line 12 replaces U by its successor with respect to S. Lines 14–15 enqueue each of —where denotes the position —along with , for processing in subsequent iterations. If the while loop terminates without detecting incompatibility, returns the phylogenetic X-tree , where T is the tree with root and is the labeling function constructed in Line 10.
| Algorithm 1: |
 |
There are two main contributors to the running time of . The first is the time to compute the successor of U along with the connected components of in Lines 12–13. Let be the successor of U with respect to . Then, can be obtained from by doing the following for each : (1) for each such that , delete all edges between v and ; (2) delete v. Using HDT (Section 2.2.2), each deletion takes amortized time. Since the total number of edge and node deletions is , the total work done in these lines is . If instead of HDT, we use its single-level version, HDT(0) (Section 2.2.3), then, by Proposition 1, the running time is , where s is the number of edge scans performed by over its entire execution.
The other main contributor to the running time of is maintaining the semi-universal nodes of the various components that result from edge deletions, so that the labels can be quickly retrieved in Line 5. Suppose this information is known for the connected component being processed in the current iteration of ’s while loop. When an edge deletion splits a connected component of in two, the algorithm traverses the smaller component to update the set of semi-universal vertices in that component. It can be shown that, over the entire execution of , any given vertex is visited times, spending time per visit. Thus, the total time spent to update semi-universal vertex information throughout the entire execution of the algorithm is .
The following result is adapted from [10].
Theorem 1.
Let be a profile. If is compatible, then returns a tree T that displays ; otherwise, returns . When implemented using HDT, the running time of is . When implemented using HDT, the running time is , where s is the total number of edge scans performed over the entire execution of .
4.2. IDPP
By Lemma 1 (Section 3.4), we can solve any instance A of IDPP by converting it to a profile and then using to determine if there exists a tree that displays . If exists, then it must be a perfect phylogeny for A. Otherwise, no perfect phylogeny for A exists.
In [13], Pe’er et al. gave a specialized algorithm for IDPP; we refer to their algorithm as . can be viewed as a variation of , with two key differences, which we explain next. Let A be an instance of IDPP, and let .
- works with instead of . This is correct, since every vertex in is semi-universal. Let m denote the number of ones in A. Then, has nodes and m edges (see Figure 4). Note that m can be considerably smaller than , since contains edges for both the zeroes and the ones of A. Since the number of edge deletions is , the total work to maintain the connected components throughout the entire execution of is , if we use HDT, and , if we use HDT(0).
- updates the set of semi-universal nodes after it computes a successor position. It does so by traversing each of the resulting connected components. Each such traversal takes time, assuming the connected components are represented by a spanning forest. The number of times a successor position is computed is bounded by the number of edges in the final phylogeny, which is . Thus, the total work performed by in updating semi-universal nodes is . In contrast, updates the set of semi-universal nodes while computing a successor position; i.e., after each tree edge deletion.
The running time of is therefore , if is implemented using HDT, and , if we use HDT(0) (we note that a somewhat better running time can be achieved if is extremely dense [13]).
5. Experiments with Tree Compatibility
We implemented using treaps [23] to represent ET trees, as done by Iyer et al. [22]. We refer to our program, written in C++, as FCT (https://zenodo.org/record/2114273#.XA7iIy2ZPOQ). FCT implements level truncation (Section 2.2.3), allowing us to specify the maximum level to which an edge can be promoted in HDT. Two extreme cases are of special interest. One permits HDT to promote edges up to the maximum allowed level . We refer to the version of FCT that implements this strategy as FCT(1). The other extreme is to disallow edge promotions entirely; i.e., we use HDT(0). We refer to this version of FCT as FCT.
We ran all experiments on a device with a 2.7-GHz dual core-Intel Core i5 processor and 8-G 1866-MHz LPDDR3 memory. The times reported here do not account for the initialization of the data structures.
5.1. Real Datasets
Table 1 shows the running times, in seconds, of FCT(0) and FCT(1) on three well-known datasets: Legumes (471 taxa, 22 trees) [24], Seabirds (121 taxa; 7 trees) [25], and Placental Mammals (116 taxa; 726 trees) [26]. In all three cases, FCT(0) and FCT(1) terminated quickly and correctly reported incompatibility, but FCT(0) was always considerably faster.
5.2. Generating Simulated Data
An inherent limitation of testing FCT on real datasets, such as the three considered in the previous section, is that they are often incompatible. Incompatible inputs do not exercise FCT as thoroughly as we would like, since the program is likely to terminate early, leaving large parts of unexamined. In order to conduct more extensive tests, we implemented a generator of compatible input profiles.
Our generator begins by producing a random phylogenetic X-tree on n leaves whose internal nodes have a user-specified degree , except possibly for the root, which may have degree less than D. The generator produces a compatible profile , by restricting to different subsets of X. We focus on two types of profiles.
- Profiles of rooted triples. We start from a binary phylogenetic X-tree . For each , we obtain by restricting to a distinct three-element subset of X. We have . If , we choose to be a set of rooted triples that defines . That is, is the only compatible supertree for (the existence of such sets of triples is a folklore theorem in phylogenetics). If , consists of every rooted triple that can be obtained by restricting to a three-element subset of X. In the latter case, we say that is a complete set of rooted triples.
- Profiles of phylogenetic trees of specified degree. We start from a phylogenetic X-tree whose nodes have degree D. We obtain by restricting to k randomly-chosen subsets of X; each label is chosen to be in a set with probability .
We conducted a series of tests on simulated datasets. Each reported data point is the average execution time, in seconds, over 30 trials.
5.3. Impact of Level Truncation
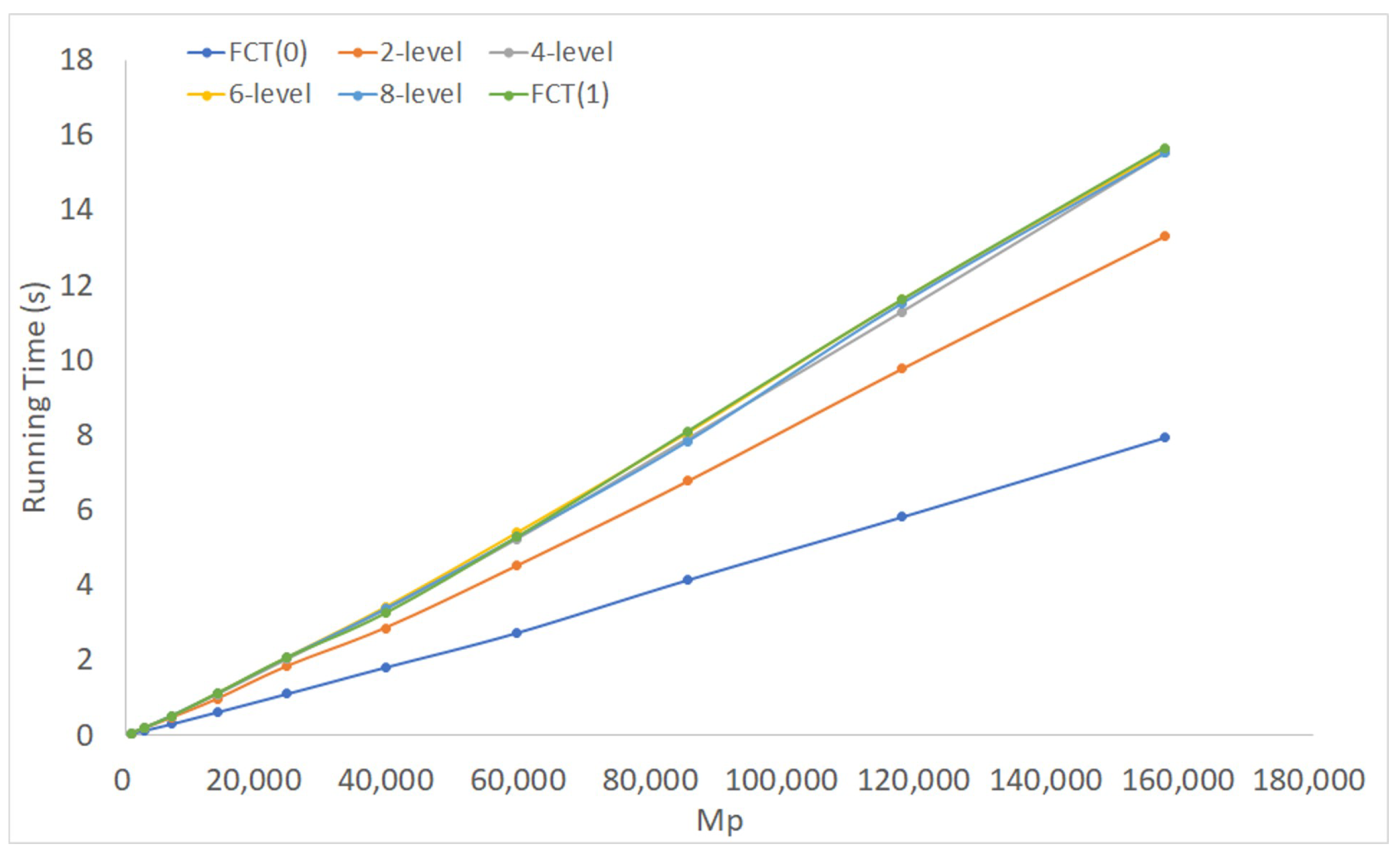
Figure 6 shows the running time of FCT on complete sets of triples, with maximum truncation levels set to 1 (i.e., FCT(0)), 2, 4, 6, 8, and (i.e., FCT(1)). The number of taxa, n, ranged from 10–55 with increments of 5. Thus, .
Observe that going beyond 4 levels made little difference. Indeed, beyond Level 1, the difference is small. This observation is consistent with the intuition that the higher levels of HDT tend to be sparsely populated and are rarely used.
FCT(0) is the clear winner in these tests. The same was true for every dataset we considered, whether they were profiles of rooted triples or profiles of more general phylogenetic trees (Figures S1–S7 in the Supplementary Materials). Thus, in the rest of this section, we focus our attention on HDT(0).
5.4. Worst-Case Time versus Empirically-Observed Time
By Theorem 1, the worst-case time of FCT(0) depends on the number of edge scans performed when searching for replacement edges. A naive estimate yields a worst-case bound of for this number: edge deletions, each requiring scans. This implies a time bound of for the entire execution of FCT(0). In contrast, we now present evidence that FCT(0)’s performance in practice may be closer to than to its worst-case bound.
Performance on Rooted Triples
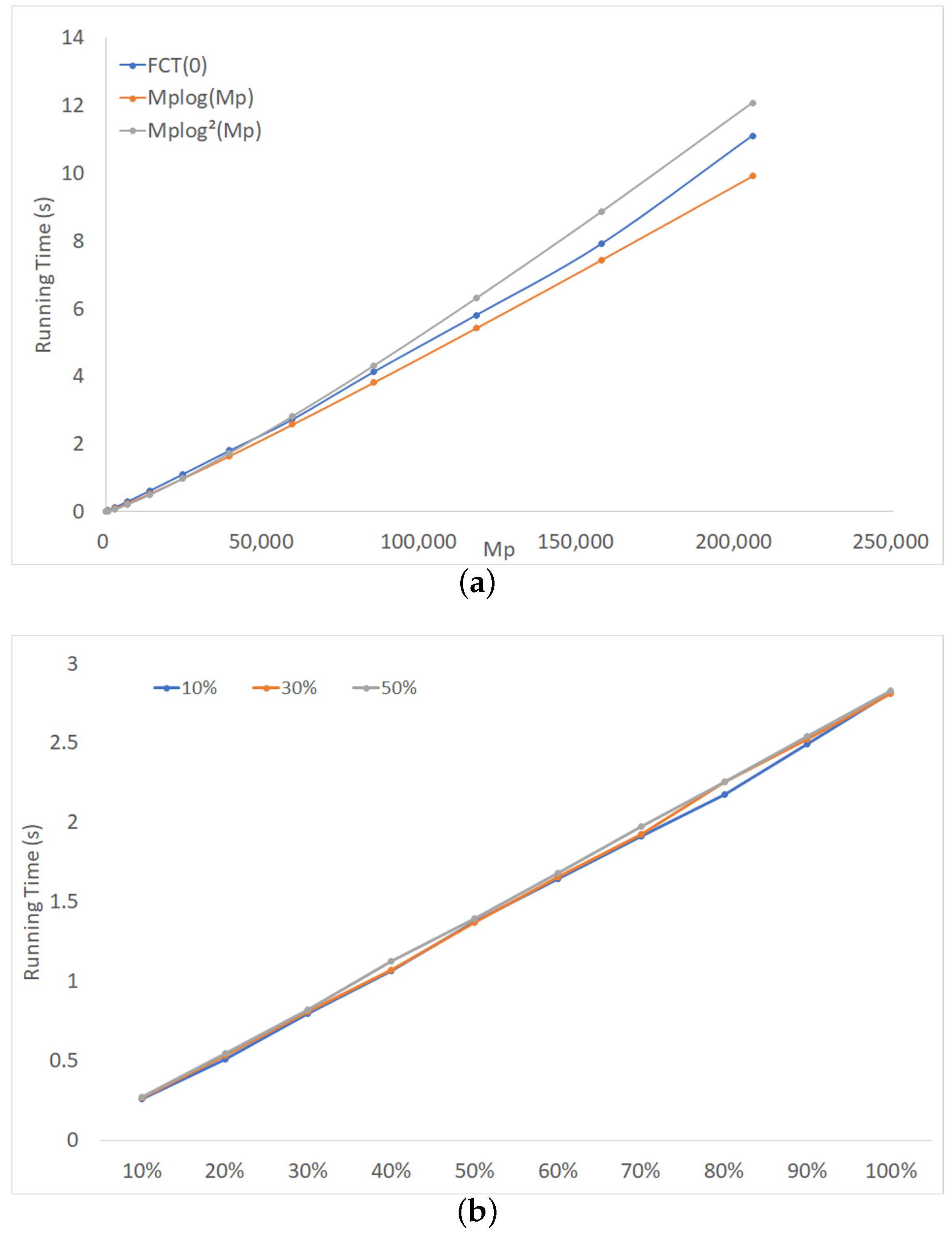
Figure 7a shows the running time of FCT(0) on complete sets of triples. The number of taxa, n, varied from 5–60 with increments of 5, and varied from 65–205,380. Also plotted in that figure are the functions and , where , , , and are appropriate constants. The plot suggests that, in practice, the running time of FCT(0) was far better than the above-mentioned worst case and may be close to .
We also explored the effect of altering the balance factor of the binary phylogenetic X-tree from which the triples wee derived (the balance factor is the ratio of the size of the smaller subtree to that of the larger subtree: a binary phylogenetic tree is balanced when ). Figure 7b shows the results of running FCT(0) on the profile of triples on labels for three balance factors: 10%, 30%, and 50%. The x-axis indicates the percentage of the maximum possible number of triples, , included in a profile. The percentage varied from 10%–100%, with increments of 10%. The running time of FCT(0) appeared to be close to linear in the number of triples; the balance factor has a negligible impact. Thus, in the rest of this section, we use starting trees that are, on average, balanced.
5.5. Performance on Profiles of More General Phylogenetic Trees
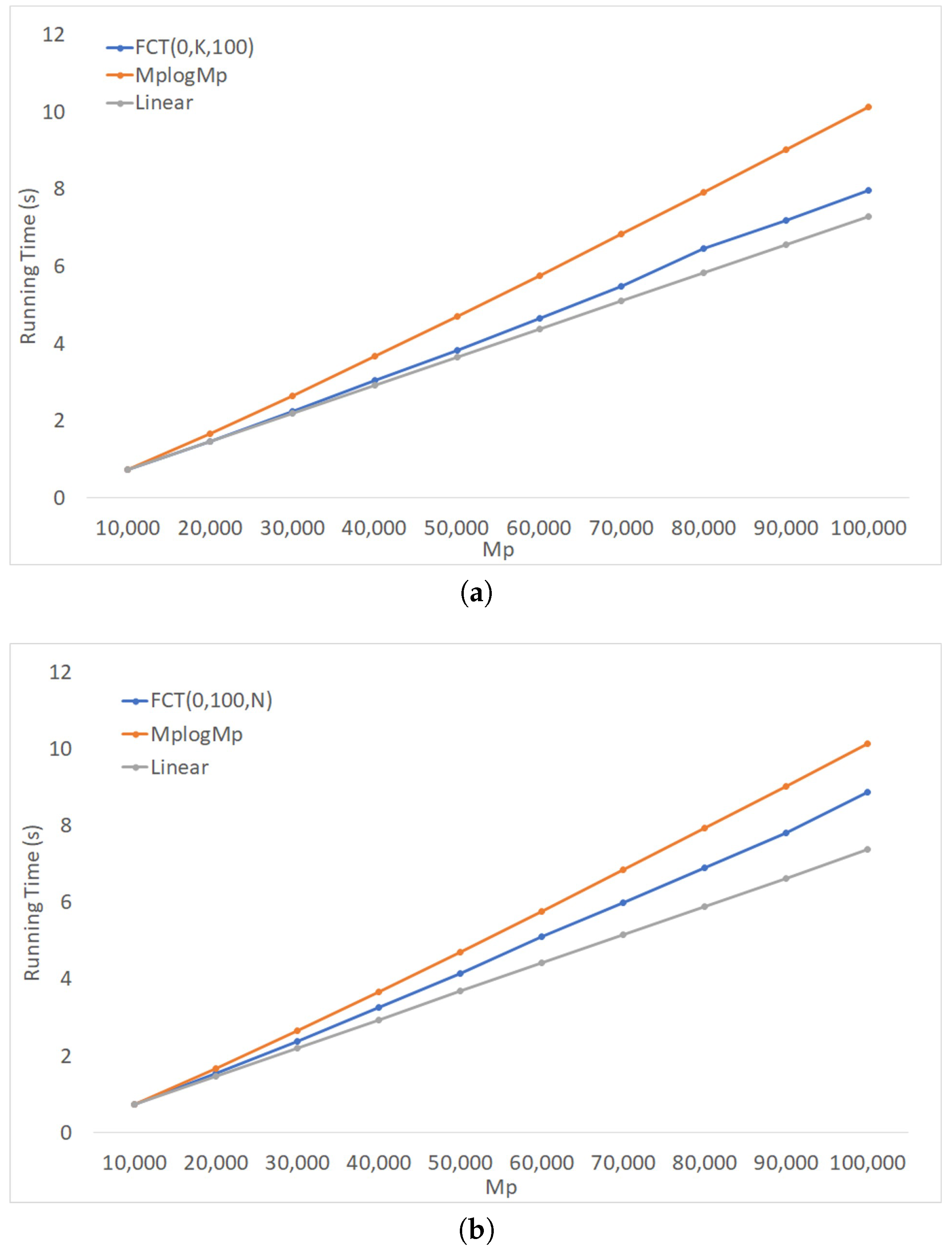
Figure 8a shows the running time of FCT(0) on profiles of binary trees when the number, k, of trees was fixed at 100 and the number, n, of taxa varied from 100–1000, with increments of 100. Figure 8b shows the performance of FCT(0) on profiles of binary trees when the number of taxa was fixed at 100, while k varied from 100–1000 with increments of 100. Both figures compare the empirical performance against and curves, using the product as a proxy for . Although the running time of FCT(0) grew non-linearly, its behavior appeared to be close to .
We also studied the performance of FCT(0) on profiles of trees of degrees 4 and 7. The results, which appear in Figures S11 and S12 of the Supplementary Materials, were qualitatively similar to those for binary trees. Note that a greater number of higher degree internal nodes can be beneficial for dynamic connectivity data structures based on spanning forests: an abundance of such nodes increased the number of non-tree edges in , which were trivial to delete (see Section 2.2). We return to this issue in Section 7.
5.6. Connectivity Testing versus Maintaining Semi-Universal Labels
Throughout all of our experiments, connectivity testing took on average slightly more than 50% of FCT(0)’s running time. FCT(0) spent most of its rest time maintaining semi-universal nodes. As noted in Section 4, the time to do the latter was asymptotically . As one might expect, there was less variability in this aspect than in dynamic connectivity testing.
6. Experiments with IDPP
We implemented , the IDPP algorithm of Pe’er et al. described in Section 4.2; we refer to our implementation as FPP (https://zenodo.org/record/2115972#.XA7mey2ZPOQ). Like FCT, FPP was implemented in C++ and used treaps to represent ET trees. There were two versions of FPP: FPP(1) used the full implementation of HDT, which allowed promotion of edges up to level ; FPP(0) used HDT(0). We ran our tests on the same machine used to obtain the results reported in Section 5.
6.1. Simulated Datasets
To generate a random instance of IDPP, our generator proceeded as follows. We started from a Prüfer code of length of , which defines a unique tree with n leaves (taxa) and n internal nodes (characters) [27,28]. We rooted at a randomly-chosen node and then translated the tree into an zero-one matrix C that encoded the clusters in (each column of C corresponds to a cluster; a one indicates that a taxon is present in the cluster, a zero that it is absent). We obtained a seed matrix B by duplicating columns in C. We built different instances of IDPP from a seed matrix by converting a randomly-chosen set of zero- and/or one-entries to question marks.
The generator can build seed matrices of different densities, where the density of a matrix is the ratio of its number of ones to its total number of entries. Let us call a column of C trivial if all its entries are one. We built a seed matrix B by duplicating non-trivial columns where the percentage of ones was above a specified threshold. If the threshold is 25%, we say that the matrix has medium density; if the threshold is 50%, we say that the matrix has high density. We also consider matrices generated without a mandatory threshold, allowing any columns to be duplicated; we refer to such instances as low-density matrices. There is a caveat: The process that we use to generate the starting tree may lead to a matrix C where no non-trivial column has the mandatory threshold. If this was the case, we reduced the threshold and tried again. For medium-density matrices, if 25% failed, we successively tried 20% and 10%. For high-density matrices, if 50% failed, we tried 40%, 30%, 20%, and 10%.
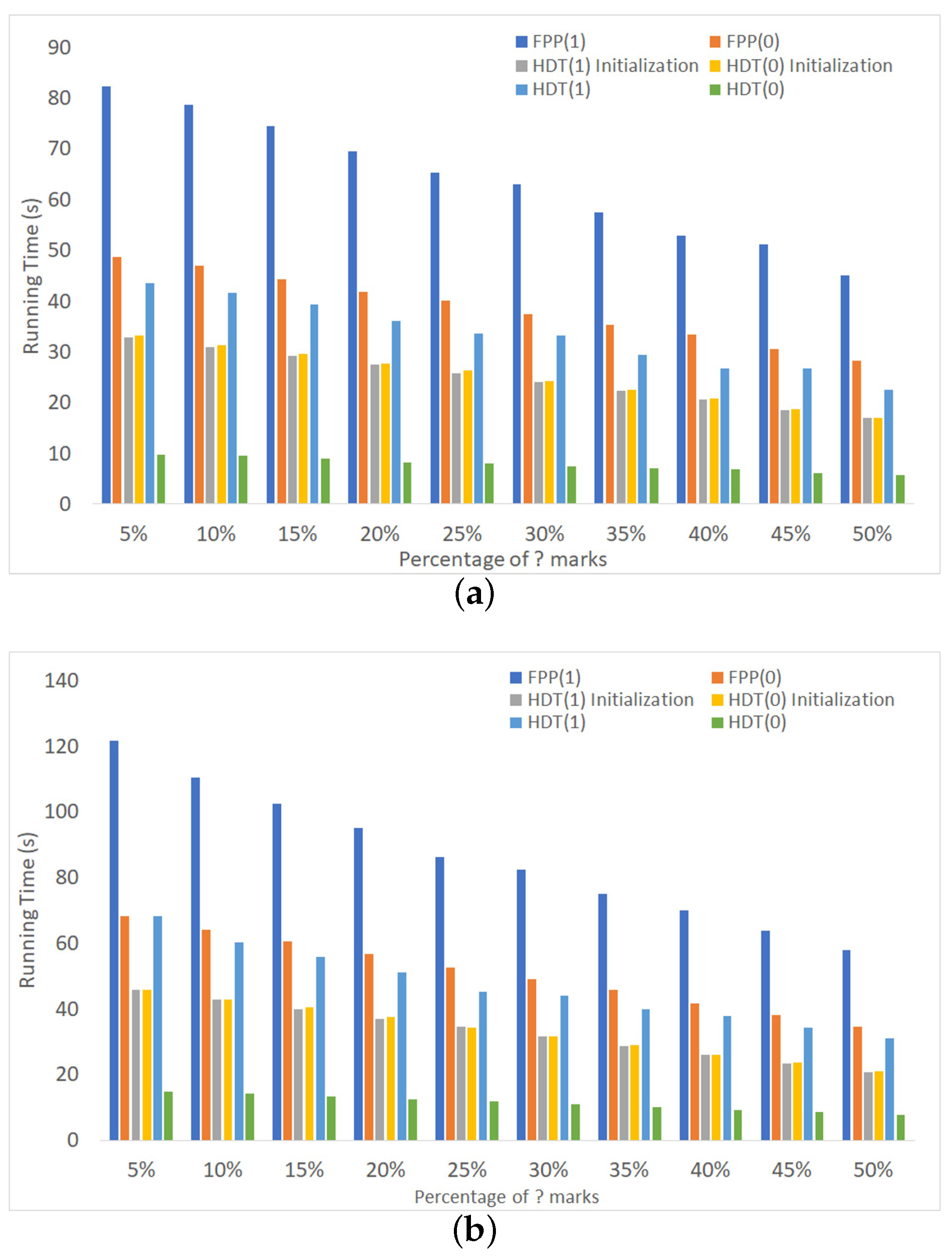
Figure 9 shows the execution time of FPP(1) and FPP(0) on medium- and high-density matrices. In both cases, n and k are fixed. For each data point reported, we started with a particular (random) seed matrix and then generated multiple IDPP instances by replacing a certain percentage of ones in the seed matrix by question marks. To be consistent, we also converted the same percentage of zeroes to question marks. The percentage ranged from 5%–50% with increments of 5%. The figures also show the time needed to initialize the connectivity data structures and the total time spent on manipulating these structures.
Without exception, the experiments showed that there was no benefit to enabling edge promotion in HDT; a single level suffices. A more extensive set of results, leading to the same conclusion, is reported in the Supplementary Materials (Figure S13).
6.2. Solving IDPP via Tree Compatibility
As explained in Section 3.4 and Section 4.2, any instance A of IDPP can be solved by transforming it into a profile and then checking if is compatible. Table 2 compares the running time of FPP(0) against that of FCT(0) applied to the corresponding instance of tree compatibility, on high-density input matrices of order . As in Figure 9, we varied the percentage of zeroes and ones converted to question marks from 5%–50% with increments of 5%. The table also shows the time the programs spent on maintaining connectivity information. FPP(0) was uniformly faster than FCT(0). Much of this speedup can be attributed to the smaller amount of time FPP(0) spent in connectivity-related work compared to FCT(0). This reflects the fact that the former operates on a graph that contains edges only for the one-entries of A, whereas the latter works with the entire display graph of , which contains edges for every one and zero-entry of A. We found similar results for low-density matrices (Supplementary Materials, Table S2).
7. Analysis
In our experiments, the most striking and consistent observation was the effectiveness of HDT(0) for both compatibility testing and IDPP, across the entire range of inputs considered. A partial explanation is that HDT(0) incurs far less overhead than HDT. This, however, does not fully explain why the running time of FCT(0) was close to in practice. Theorem 1 indicates that this performance may be tied to the small number of edge scans performed throughout the execution of the algorithms. Here, we explore this issue in more detail. First, we break down in more detail the factors that affect the performance of HDT(0) in and .
For each vertex , let denote the number of children of v. At any stage of its execution, maintains a graph H obtained from through edge and node deletions. Because these deletions are performed top-down, the number of edges incident on any node v in H is at most .
Lemma 2.
Let be a tree edge in the current spanning forest F maintained by HDT(0) during the execution of (or ), and let and be the two trees that result from deleting e from the tree in F that contains e. Assume, without loss of generality, that . Then, searching for a replacement edge for e requires scanning at most non-tree edges.
Proof.
We claim that the number of non-tree edges scanned is at most one more than the number of edges with both endpoints in . To see why, note that the scan for a replacement edge stops as soon as either (i) we encounter an edge that has one endpoint, x, in and the other, y, outside (and, thus, is a replacement edge) or (ii) we find that all non-tree edges incident on have both of their endpoints in (and, thus, no replacement edge exists). To complete the proof, we argue that the number of non-tree edges with both endpoints in is at most .
Let be the total number of tree and non-tree edges incident on . Then, . The number of tree edges with both endpoints in is . The number of non-tree edges is thus , as claimed. □
Proposition 2.
Let F be the current spanning forest maintained by HDT(0) during the execution of (or ); let v be the semi-universal node being processed during the computation of the next successor position; and let e be the next edge incident on v to be deleted. Then, the following hold.
- 1.
- Deleting e takes:
- (a)
- time if e is a non-tree edge,
- (b)
- time if e is the only tree edge incident on v, and
- (c)
- time otherwise, where T is the smaller of the two trees created by deleting e from F. In particular, if the input profile consists of binary trees, the time is .
- 2.
- Suppose and that e is the first edge incident on v that is deleted. Let be the other edge incident on v. Then, at the time of deletion, is a tree edge, and deleting it takes time.
Proof.
(1) Part (a) was noted in Section 2.2.2. For Part (b) observe that if e is the only tree edge incident on v, then deleting e leaves v as an isolated node in the spanning forest; i.e, as a component of size one, and, hence, as the smaller component. Any non-tree edge incident on v must be a replacement edge, and, if such an edge is found, it takes time to re-link v to the rest of the forest. Part (c) follows from Lemma 2 and the fact that there are at most edges with both endpoints incident in T. The claim for profiles of binary trees follows by noting that for every node w.
(2) There are two cases. If was a tree edge before deleting e, then remains a tree edge after the deletion of e. If is a non-tree edge before deleting e, then must be used as a replacement edge during the deletion of e, so becomes a tree edge.
HDT(0) deletes by first splitting, in time, the ET-tree that contains both endpoints of . This leaves vertex v as the sole element in the smaller component, after which the vertex v is simply discarded. □
Proposition 2 points to three key issues that affect the performance of HDT(0) when used in either or : the number of non-tree edge deletions, the number of tree edge deletions, and the size of the smaller components resulting from tree edge deletions.
For profiles consisting of binary phylogenetic trees (including profiles of triples), Proposition 2(2) implies that at least half of HDT(0)’s tree-edge deletions take time in the worst case. This is faster than the amortized time they take when performed by HDT. Proposition 2(1) notes the downside: the other half of the deletions could be expensive. These observations hold to some extent for profiles consisting of trees of small degree, although the ratio of inexpensive to expensive tree-edge deletions goes down. On the other hand, for larger degree trees, and in particular for the denser inputs generated by IDPP, non-tree edges are relatively abundant. As indicated in Proposition 2(1a), such edges are trivial to delete.
Proposition 2(1c) implies that if the majority of the smaller components resulting from tree edge deletions are very small, then the time that HDT(0) spends on maintaining connectivity information throughout the execution of (or ) will also be small. More precisely, let be the successive tree edges deleted by (or ), and let , where is the smaller of the two trees of HDT(0)’s spanning forest created by deleting . Then, the total time spent in all tree edge deletions is . If, for instance, , then the total time to maintain connectivity information (including the total time for non-tree edge deletions, which take time each) would be , matching the behavior we observed in Section 5.
In the next sections, we examine separately the three key factors affecting the performance of HDT(0) on and . Section 7.1 studies the impact of deletion of non-tree edges on overall performance. As one would expect, the number of such deletions increases with the degree of the input trees. Section 7.2 examines the impact of tree-edge deletion. Here, the focus is on the number of edges scanned in searching for a replacement edge. As we shall see, the total number of such edges grows at a rate that seems only slightly super-linear in . Section 7.3 studies the size of the smaller components encountered during tree edge deletions. Surprisingly, we find that components of size at most two constitute the overwhelming majority across a wide range of profiles.
7.1. The Impact of Deleting Non-Tree Edges
We first examine the prevalence of non-tree edge deletions and their total contribution to the execution time. Recall that each such deletion takes time (Proposition 2(1a)).
Table 3 shows HDT(0)’s performance on complete sets of triples. The first row of the table shows the ratio of the number of actually deleted non-tree edges to the total number of edges in . The second row shows the percentage of time that HDT(0) spends on deleting non-tree edges. Both numbers are small, indicating that the work is dominated by processing tree edges.
Table 4 and Table 5 show the running time of FCT(0) on profiles of binary phylogenetic trees. Similar to the situation for triples, the number of non-tree edge deletions and the amount of time performing them is relatively small.
Table 6 and Table 7 show the running time of FCT(0) on phylogenetic trees where internal node have degree seven. As expected, by increasing the degree of internal nodes, we increased the number of non-tree edges. On the other hand, the contribution of these edges to the total running time did not increase markedly.
Table 3, Table 4, Table 5, Table 6 and Table 7 indicate that to get a better understanding of the behavior of FCT(0) on profiles of low-degree trees, it is necessary to focus on tree edge deletions and, more specifically, on the time spent scanning for replacement edges. We study this issue in the next section. Before doing so, we consider the case of high-degree vertices, which is encountered in IDPP.
Table 8, Table 9 and Table 10 show the performance of FPP(0) on low-, medium-, and high-density inputs. The results show that deletions were mostly done on the non-tree edges, which makes sense due to their relative abundance. Further, the program spent a large fraction of its time (in some cases, upwards of 50%) on such edges. This is significant, since it suggests the potentially more expensive deletions of tree edges are not as expensive as the worst-case bound would indicate. As we shall see in the next section, this appears to be due to the fact that the total number of edges scanned in search of replacement edges is relatively small.
7.2. The Number of Edges Scanned
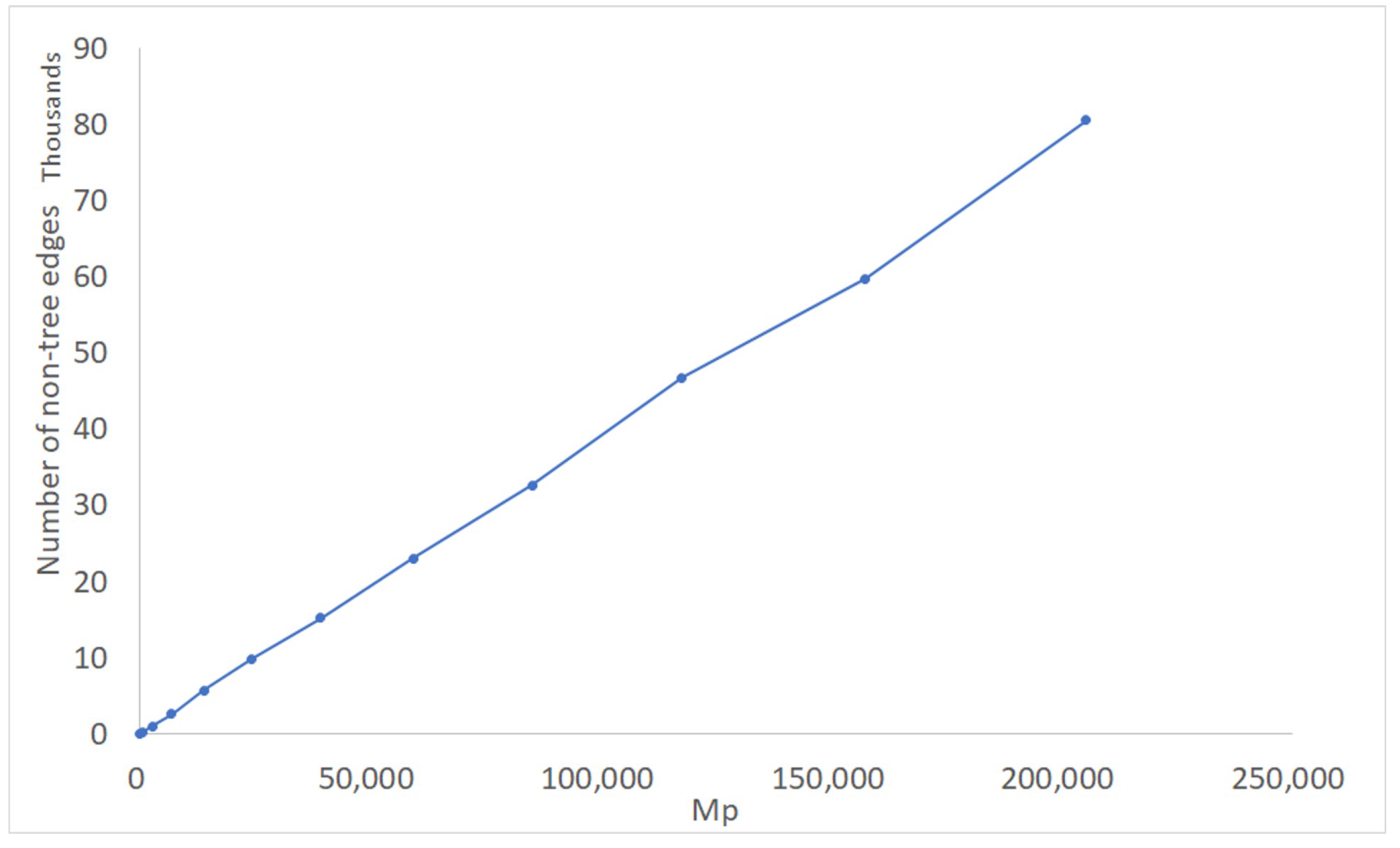
We now examine the impact of edge scans more closely. Figure 10 shows the average number of non-tree edges, as a function of , actually scanned by FCT(0) when operating on complete sets of rooted triples. Observe that not only does the number of edge scans increase at an only slightly super-linear rate, but also the average number of non-tree edges scanned is considerably smaller than . Together with Theorem 1, this explains the near- behavior seen in Figure 7.
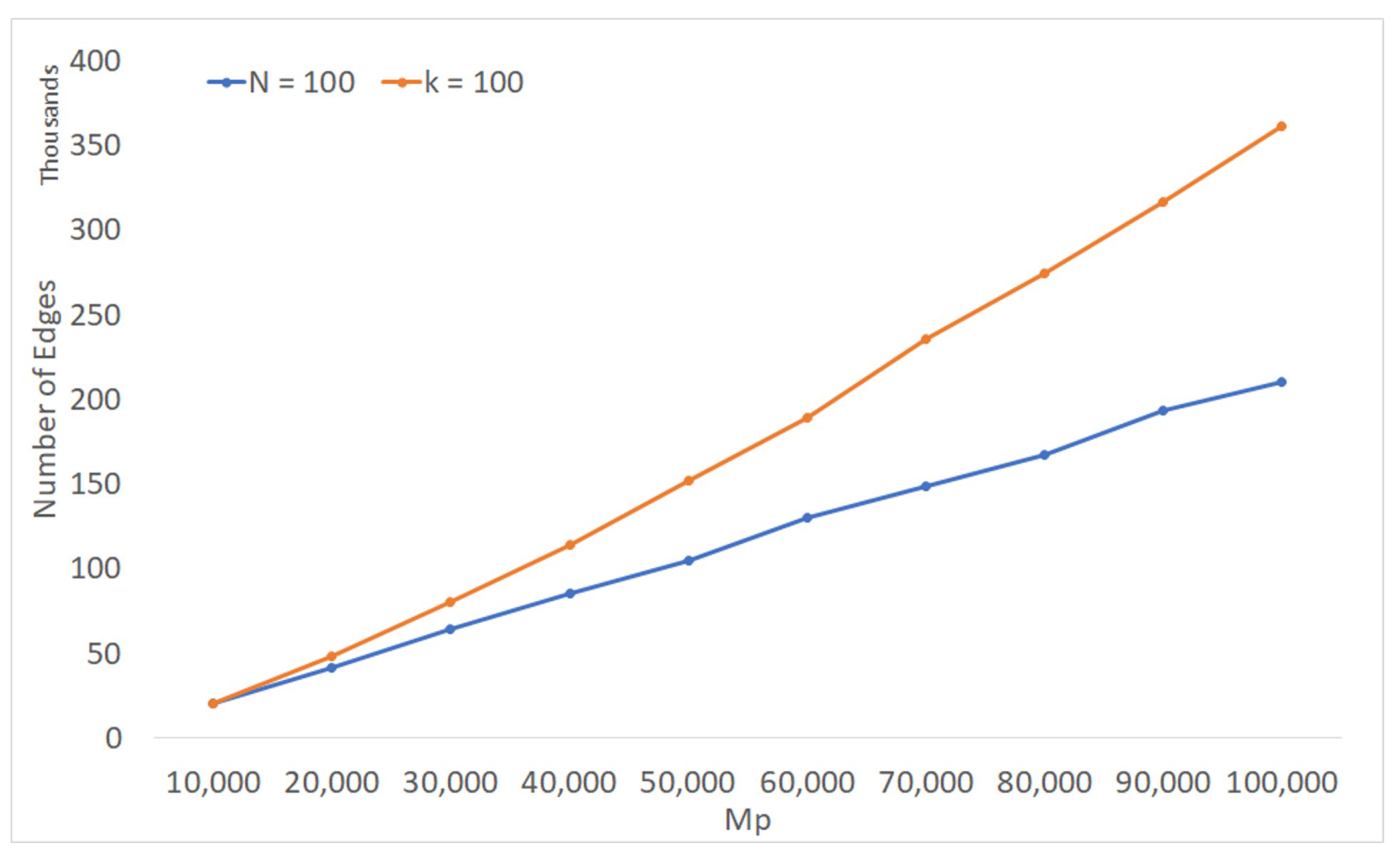
Figure 11 shows the average number of non-tree edges actually scanned by FCT(0) on profiles of binary phylogenies. The input profiles were generated as described in Section 5.4. As in that section, we varied one of n, the number of labels, or k, the number of trees, while keeping the other quantity fixed, and we took the product as a proxy for .
When n was fixed, the number of non-tree edges scanned was roughly proportional to , suggesting that the running time of FCT(0) was . This is reflected in Figure 8a. On the other hand, when k was fixed, the number of non-tree edges scanned appeared to grow in a slightly super-linear manner. The explanation appears to be that increasing k increased the amount of overlap among the trees, and consequently also the number of non-tree edges. Nevertheless, the overall running time of FCT(0) in this case appeared close to ; see Figure 8b. This may be because the effect of increasing the number of non-tree edges was mitigated by another factor: the sizes of the smaller of two subtrees resulting from edge deletion.
Table 11 reports the number of non-tree edges scanned throughout the execution of FPP(0). Overall, the number of scanned non-tree edges was smaller than the total number of edges. This again illustrates that searching for replacement edges did not significantly contribute to the running time. Surprisingly, the largest number of edge scans occurred for medium-density matrices. We have not found a satisfactory explanation for this observation.
7.3. The Size of the Smaller Component
In the previous section, we saw that HDT(0) scanned relatively few non-tree edges when used in either FCT(0) or FPP(0). Table 12, Table 13, Table 14, Table 15 and Table 16 show one reason for this: the smaller component was often very small. In fact, the small component often contained just one or two nodes. The tables show the number of times the smaller subtree resulting from an edge deletion had at most two nodes (“Num. ”) and more than two nodes (“Num. ”) for different types of input profiles. In all cases, subtrees of a size at most two outnumbered the rest by a wide margin.
Subtrees with at most two nodes (and, thus, one edge) were easily handled in time: the search for a replacement edge terminated immediately after encountering the first non-tree edge incident on one of the at most two nodes in the subtree, assuming such an edge exists.
Table 17 examines the distribution of the size of the smaller component in greater detail for profiles of binary trees. The number of small components of a given size declined rapidly as the size increased. This pattern was evident even for profiles of trees of larger degrees, including for instances of IDPP, although it was somewhat less marked; see Tables S34–S46 of the Supplementary Materials. Invariably in our experiments, the overwhelming majority of the smaller subtrees had just one node. This means that, prior to deletion, the single node in that small component was a semi-universal node with just one incident tree edge.
8. Discussion
Our experimental results show that both the tree compatibility testing algorithm of [10] and the IDPP algorithm of [9] performed at least as well as their theoretical worst-case bounds, and often better, if implemented using HDT(0), a much-simplified version of the HDT dynamic connectivity data structure.
The results of Section 7 indicate that the main reason for the observed performance of HDT(0) is that the components that were scanned in search for replacement edges tended to be extremely small. Since we observed this phenomenon for a wide range of input profiles, we suspect that it is not an artifact of our experimental setup, but instead reflects a basic property of the graphs with which we are dealing. Indeed, and use very special kinds of graphs—constructed from profiles of phylogenetic trees by gluing leaves with the same label—and perform deletions top-down. It is an open question whether there is a way to bound the total sizes of the smaller trees encountered during the execution of those algorithms, either asymptotically or in expectation.
Profiles consisting of binary trees—and, in particular, triples—are quite common in practice. Proposition 2(1b) of Section 7 implies that for such profiles, certain spanning forests are better than others for maintaining connectivity information. A “good” spanning forest is one where the number of semi-universal nodes incident to only one tree edge is large, relative to the total number of semi-universal nodes. The results reported in Section 7.3 (in particular, Table 17) indicate that the spanning forests our programs generated were good in this sense, despite the fact that the programs took no special measures to ensure this. It is an open question whether there is an analytical explanation for this phenomenon. Another open question is whether good spanning forests always exist and, if so, whether they can be constructed and maintained efficiently.
In our experiments, we never encountered a setting where HDT outperformed HDT(0). Of course, this does not mean that the latter is always to be preferred over the former. It would be interesting to find an application of dynamic graph connectivity where HDT is preferable to HDT(0) in practice.
Supplementary Materials
Additional figures and tables are available online at https://www.mdpi.com/1999-4893/12/3/53/s1. Figures S1–S17, Tables S1–S46.
Author Contributions
Conceptualization, D.F.-B. and L.L.; methodology, D.F.-B. and L.L.; software, L.L.; validation, D.F.-B. and L.L.; formal analysis, D.F.-B. and L.L.; investigation, L.L.; writing, original draft preparation, D.F.-B. and L.L.; writing, review and editing, D.F.-B. and L.L.; visualization, L.L.; supervision, D.F.-B.; project administration, D.F.-B.; funding acquisition, D.F.-B.
Funding
This research was funded by the National Science Foundation Grants IOS-1444806 and CCF-1422134.
Acknowledgments
The authors thank the reviewers for their comments, which helped to improve the presentation of the paper.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| IDPP | Incomplete directed perfect phylogeny |
| HDT | The dynamic graph connectivity data structure of Holm, de Lichtenberg, and Thorup [15] |
| HDT(0) | HDT with edge promotion disallowed |
| The tree compatibility algorithm of [9] | |
| FCT | The authors’ implementation of |
| FCT(1) | Version of FCT that uses HDT |
| FCT(0) | Version of FCT that uses HDT(0) |
| The IDPP algorithm of [13] | |
| FPP | The authors’ implementation of PPSS |
| FPP(1) | Version of FPP that uses HDT |
| FPP(0) | Version of FPP that uses HDT(0) |
| NTE | Non-tree edge |
References
- Steel, M.A. The complexity of reconstructing trees from qualitative characters and subtrees. J. Classif. 1992, 9, 91–116. [Google Scholar] [CrossRef]
- Semple, C.; Steel, M. Phylogenetics; Oxford Lecture Series in Mathematics; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Chimani, M.; Rahmann, S.; Böcker, S. Exact ILP solutions for phylogenetic minimum flip problems. In Proceedings of the First ACM International Conference on Bioinformatics and Computational Biology, Niagara Falls, NY, USA, 2–4 August 2010; ACM: New York, NY, USA, 2010; pp. 147–153. [Google Scholar]
- Bininda-Emonds, O.R.P. (Ed.) Phylogenetic Supertrees: Combining Information to Reveal the Tree of Life; Series on Computational Biology; Springer: Berlin, Germany, 2004; Volume 4. [Google Scholar]
- Warnow, T. Supertree Construction: Opportunities and Challenges. arXiv, 2018; arXiv:1805.03530. [Google Scholar]
- Hinchliff, C.E.; Smith, S.A.; Allman, J.F.; Burleigh, J.G.; Chaudhary, R.; Coghill, L.M.; Crandall, K.A.; Deng, J.; Drew, B.T.; Gazis, R.; et al. Synthesis of phylogeny and taxonomy into a comprehensive tree of life. Proc. Natl. Acad. Sci. USA 2015, 112, 12764–12769. [Google Scholar] [CrossRef] [PubMed]
- Redelings, B.D.; Holder, M.T. A supertree pipeline for summarizing phylogenetic and taxonomic information for millions of species. PeerJ 2017, 5, e3058. [Google Scholar] [CrossRef] [PubMed]
- Aho, A.V.; Sagiv, Y.; Szymanski, T.G.; Ullman, J.D. Inferring a tree from lowest common ancestors with an application to the optimization of relational expressions. SIAM J. Comput. 1981, 10, 405–421. [Google Scholar] [CrossRef]
- Deng, Y.; Fernández-Baca, D. Fast compatibility testing for rooted phylogenetic trees. Algorithmica 2018, 80, 2453–2477. [Google Scholar] [CrossRef]
- Deng, Y.; Fernández-Baca, D. An efficient algorithm for testing the compatibility of phylogenies with nested taxa. Algorithms Mol. Biol. 2017, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Bryant, D.; Lagergren, J. Compatibility of unrooted phylogenetic trees is FPT. Theor. Comput. Sci. 2006, 351, 296–302. [Google Scholar] [CrossRef]
- Henzinger, M.R.; King, V.; Warnow, T. Constructing a tree from homeomorphic subtrees, with applications to computational evolutionary biology. Algorithmica 1999, 24, 1–13. [Google Scholar] [CrossRef]
- Pe’er, I.; Pupko, T.; Shamir, R.; Sharan, R. Incomplete directed perfect phylogeny. SIAM J. Comput. 2004, 33, 590–607. [Google Scholar] [CrossRef]
- Thorup, M. Decremental dynamic connectivity. J. Algorithms 1999, 33, 229–243. [Google Scholar] [CrossRef]
- Holm, J.; de Lichtenberg, K.; Thorup, M. Poly-logarithmic deterministic fully-dynamic algorithms for connectivity, minimum spanning tree, 2-edge, and biconnectivity. J. ACM 2001, 48, 723–760. [Google Scholar] [CrossRef]
- Nikaido, M.; Rooney, A.P.; Okada, N. Phylogenetic relationships among cetartiodactyls based on insertions of short and long interpersed elements: hippopotamuses are the closest extant relatives of whales. Proc. Natl. Acad. Sci. USA 1999, 96, 10261–10266. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, G.; Shamir, R. The incomplete perfect phylogeny haplotype problem. J. Bioinform. Comput. Biol. 2005, 3, 359–384. [Google Scholar] [CrossRef] [PubMed]
- Even, S.; Shiloach, Y. An On-Line Edge-Deletion Problem. J. ACM 1981, 28, 1–4. [Google Scholar] [CrossRef]
- Henzinger, M.R.; King, V. Randomized fully dynamic graph algorithms with polylogarithmic time per operation. J. ACM 1999, 46, 502–516. [Google Scholar] [CrossRef]
- Thorup, M. Near-optimal fully-dynamic graph connectivity. In Proceedings of the 32nd Annual ACM Symposium on Theory of Computing, Portland, OR, USA, 21–23 May 2000; ACM: New York, NY, USA, 2000; pp. 343–350. [Google Scholar]
- Kapron, B.M.; King, V.; Mountjoy, B. Dynamic graph connectivity in polylogarithmic worst case time. In Proceedings of the Twenty-Fourth Annual ACM-SIAM Symposium on Discrete Algorithms, Society for Industrial and Applied Mathematics, New Orleans, LA, USA, 6–8 January 2013; pp. 1131–1142. [Google Scholar]
- Iyer, R.; Karger, D.; Rahul, H.; Thorup, M. An Experimental Study of Polylogarithmic, Fully Dynamic, Connectivity Algorithms. J. Exp. Algorithmics 2001, 6, 4. [Google Scholar] [CrossRef]
- Seidel, R.; Aragon, C.R. Randomized search trees. Algorithmica 1996, 16, 464–497. [Google Scholar] [CrossRef]
- Wojciechowski, M.; Sanderson, M.; Steele, K.; Liston, A. Molecular phylogeny of the “Temperate Herbaceous Tribes” of Papilionoid legumes: A supertree approach. In Advances in Legume Systematics; Herendeen, P., Bruneau, A., Eds.; Royal Botanic Gardens: Kew, UK, 2000; Volume 9, pp. 277–298. [Google Scholar]
- Kennedy, M.; Page, R.D.M. Seabird supertrees: Combining partial estimates of procellariiform phylogeny. Auk 2002, 119, 88–108. [Google Scholar] [CrossRef]
- Beck, R.M.D.; Bininda-Emonds, O.R.P.; Cardillo, M.; Liu, F.G.R.; Purvis, A. A higher-level MRP supertree of placental mammals. BMC Evol. Biol. 2006, 6, 93. [Google Scholar] [CrossRef] [PubMed]
- Prüfer, H. Neuer Beweis eines Satzes über Permutationen. Arch. Math. Phys. 1918, 27, 742–744. [Google Scholar]
- Pemmaraju, S.; Skiena, S. Computational Discrete Mathematics: Combinatorics and Graph Theory with Mathematica®; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
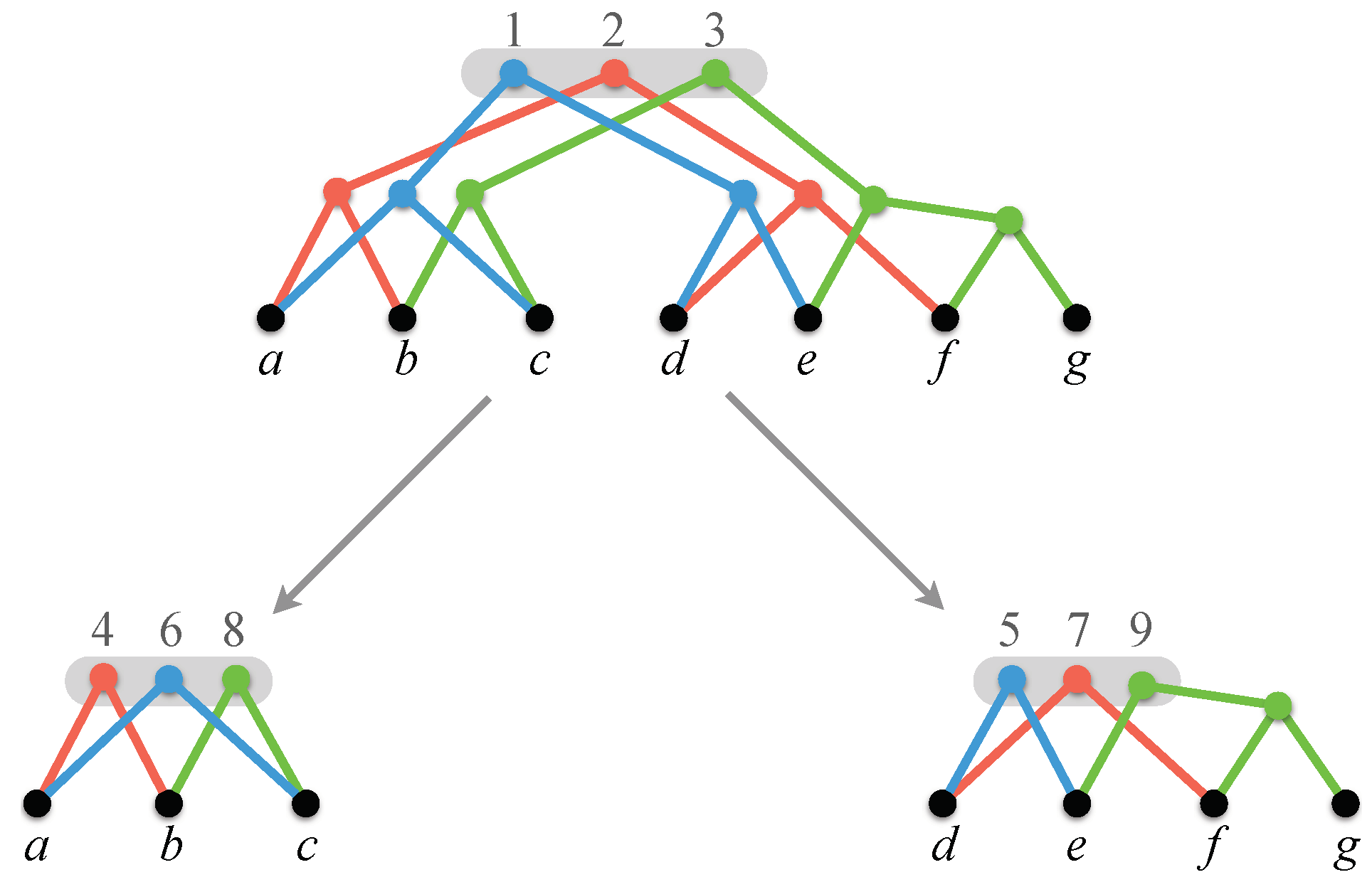
Figure 1.
A profile. Leaves are labeled with species; internal nodes are numbered for later reference.
Figure 1.
A profile. Leaves are labeled with species; internal nodes are numbered for later reference.

Figure 2.
A compatible supertree for the profile of Figure 1. Each internal node is labeled with the set of nodes that are mapped to it by the tree compatibility algorithm described in Section 4.1.
Figure 2.
A compatible supertree for the profile of Figure 1. Each internal node is labeled with the set of nodes that are mapped to it by the tree compatibility algorithm described in Section 4.1.

Figure 3.
The display graph for the profile of Figure 1.
Figure 3.
The display graph for the profile of Figure 1.

Figure 4.
(Left) A character matrix A. (Right) The display graph for . The solid nodes and edges are the only part of the display graph that algorithm (Section 4.2) uses.
Figure 4.
(Left) A character matrix A. (Right) The display graph for . The solid nodes and edges are the only part of the display graph that algorithm (Section 4.2) uses.

Figure 5.
The root position and its successor for the profile of Figure 1. Sets of nodes inside shaded boxes are positions. As a result of computing the successor, graph breaks down into two components. In this example, all nodes in the positions shown are semi-universal. Note, however, that, in general, not all nodes in a position are semi-universal (see [9,10]).
Figure 5.
The root position and its successor for the profile of Figure 1. Sets of nodes inside shaded boxes are positions. As a result of computing the successor, graph breaks down into two components. In this example, all nodes in the positions shown are semi-universal. Note, however, that, in general, not all nodes in a position are semi-universal (see [9,10]).

Figure 6.
Performance of FCT for varying degrees of level truncation on complete sets of triples.

Figure 7.
FCT(0) on profiles of rooted triples. (a) Complete sets of triples with varying number of taxa. (b) Rooted triples on 40 labels for different percentages of the maximum number of triples.
Figure 7.
FCT(0) on profiles of rooted triples. (a) Complete sets of triples with varying number of taxa. (b) Rooted triples on 40 labels for different percentages of the maximum number of triples.

Figure 8.
FCT(0) on profiles of binary trees. (a) Running time for 100 trees and varying number of taxa. (b) Running time for 100 taxa and varying number of trees.
Figure 8.
FCT(0) on profiles of binary trees. (a) Running time for 100 trees and varying number of taxa. (b) Running time for 100 taxa and varying number of trees.

Figure 9.
Running time of FPP(0) and FPP(1) on matrices of order for (a) medium- and (b) high-density inputs.
Figure 9.
Running time of FPP(0) and FPP(1) on matrices of order for (a) medium- and (b) high-density inputs.

Figure 10.
Number of non-tree edges scanned by FCT(0) for complete sets of rooted triples.

Figure 11.
Number of non-tree edge FCT scans for profiles of binary phylogenetic trees. The blue curve corresponds to varying k, while keeping n fixed at 100. The orange curve corresponds to varying n while keeping k fixed at 100.
Figure 11.
Number of non-tree edge FCT scans for profiles of binary phylogenetic trees. The blue curve corresponds to varying k, while keeping n fixed at 100. The orange curve corresponds to varying n while keeping k fixed at 100.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Runtime on real datasets.
| Legumes | Seabirds | Mammals | |
|---|---|---|---|
| FCT(0) | 0.0277 s | 0.0051 s | 0.1327 s |
| FCT(1) | 0.0926 s | 0.0192 s | 0.3623 s |
Table 2.
Comparison between execution time (in seconds) of the tree compatibility algorithm on transformed IDPP and original IDPP on high-density matrices of order .
Table 2.
Comparison between execution time (in seconds) of the tree compatibility algorithm on transformed IDPP and original IDPP on high-density matrices of order .
| 5% | 10% | 15% | 20% | 25% | 30% | 35% | 40% | 45% | 50% | |
|---|---|---|---|---|---|---|---|---|---|---|
| FCT(0) | 10.07 | 10.00 | 9.87 | 9.61 | 9.52 | 9.41 | 9.36 | 9.07 | 9.05 | 8.80 |
| FPP(0) | 6.43 | 6.48 | 6.38 | 6.32 | 6.43 | 5.68 | 6.30 | 6.55 | 6.44 | 6.12 |
| Connectivity (FCT) | 3.37 | 3.37 | 3.32 | 3.24 | 3.28 | 3.22 | 3.30 | 3.16 | 3.15 | 3.07 |
| Connectivity (FPP) | 1.38 | 1.36 | 1.33 | 1.28 | 1.36 | 1.26 | 1.35 | 1.42 | 1.38 | 1.30 |
Table 3.
FCT(0) on complete sets of triples: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
Table 3.
FCT(0) on complete sets of triples: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
| 65 | 730 | 2745 | 6860 | 13,825 | 24,390 | 39,305 | 59,320 | 85,185 | 117,650 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 9.25% | 19.51% | 21.88% | 21.08% | 22.49% | 23.00% | 23.20% | 22.49% | 23.05% | 23.13% | |
| 7.48% | 9.49% | 10.04% | 9.86% | 10.09% | 10.28% | 10.22% | 9.99% | 9.99% | 9.83% |
Table 4.
FCT(0) on profiles of binary phylogenetic trees for and varying n: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
Table 4.
FCT(0) on profiles of binary phylogenetic trees for and varying n: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
| 10,000 | 20,000 | 30,000 | 40,000 | 50,000 | 60,000 | 70,000 | 80,000 | 90,000 | 100,000 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 17.13% | 17.32% | 17.25% | 17.27% | 17.25% | 17.27% | 17.35% | 17.35% | 17.32% | 17.42% | |
| 8.03% | 7.97% | 7.64% | 7.82% | 7.81% | 7.58% | 7.69% | 7.58% | 7.62% | 7.63% |
Table 5.
FCT(0) on profiles of binary phylogenetic trees for and varying k: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
Table 5.
FCT(0) on profiles of binary phylogenetic trees for and varying k: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
| 10,000 | 20,000 | 30,000 | 40,000 | 50,000 | 60,000 | 70,000 | 80,000 | 90,000 | 100,000 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 17.87% | 18.76% | 18.76% | 18.79% | 18.76% | 19.17% | 19.33% | 19.13% | 19.11% | 19.35% | |
| 8.21% | 8.30% | 8.29% | 8.35% | 8.47% | 8.22% | 8.30% | 8.38% | 8.29% | 8.39% |
Table 6.
FCT(0) on profiles of phylogenetic trees of degree 7 with and varying n: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
Table 6.
FCT(0) on profiles of phylogenetic trees of degree 7 with and varying n: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
| 10,000 | 20,000 | 30,000 | 40,000 | 50,000 | 60,000 | 70,000 | 80,000 | 90,000 | 100,000 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 42.23% | 42.72% | 42.69% | 42.16% | 42.27% | 42.13% | 42.11% | 42.47% | 42.14% | 42.23% | |
| 14.07% | 13.85% | 13.39% | 13.20% | 13.32% | 12.88% | 12.65% | 13.02% | 12.92% | 12.99% |
Table 7.
FCT(0) on profiles of phylogenetic trees of degree 7 with and varying k: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
Table 7.
FCT(0) on profiles of phylogenetic trees of degree 7 with and varying k: Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
| 10,000 | 20,000 | 30,000 | 40,000 | 50,000 | 60,000 | 70,000 | 80,000 | 90,000 | 100,000 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 42.20% | 42.59% | 44.90% | 45.04% | 44.88% | 44.92% | 46.22% | 46.44% | 44.79% | 44.97% | |
| 14.00% | 13.94% | 15.00% | 14.93% | 15.15% | 15.13% | 15.34% | 15.40% | 14.96% | 14.94% |
Table 8.
FPP(0) on low-density matrices of order : Ratio of the number of deleted non-tree edges to the total number of edges and the ratio of time spent on deleting non-tree edges to total HDT execution time.
Table 8.
FPP(0) on low-density matrices of order : Ratio of the number of deleted non-tree edges to the total number of edges and the ratio of time spent on deleting non-tree edges to total HDT execution time.
| 5% | 10% | 15% | 20% | 25% | 30% | 35% | 40% | 45% | 50% | |
|---|---|---|---|---|---|---|---|---|---|---|
| 88.53% | 87.95% | 87.42% | 87.12% | 86.54% | 85.94% | 85.36% | 84.62% | 83.73% | 82.70% | |
| 41.77% | 41.01% | 40.18% | 39.86% | 38.96% | 38.07% | 37.83% | 36.89% | 35.07% | 34.10% |
Table 9.
FPP(0) on medium-density matrices of order : Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
Table 9.
FPP(0) on medium-density matrices of order : Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
| 5% | 10% | 15% | 20% | 25% | 30% | 35% | 40% | 45% | 50% | |
|---|---|---|---|---|---|---|---|---|---|---|
| 88.78% | 88.28% | 87.57% | 87.03% | 86.32% | 85.00% | 83.87% | 82.61% | 82.12% | 79.89% | |
| 56.36% | 54.65% | 54.61% | 55.24% | 53.33% | 52.79% | 52.84% | 51.24% | 50.24% | 49.78% |
Table 10.
FPP(0) on high-density matrices of order : Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
Table 10.
FPP(0) on high-density matrices of order : Ratio of number of deleted non-tree edges to total number of edges and ratio of time spent on deleting non-tree edges to total HDT execution time.
| 5% | 10% | 15% | 20% | 25% | 30% | 35% | 40% | 45% | 50% | |
|---|---|---|---|---|---|---|---|---|---|---|
| 97.80% | 97.69% | 97.58% | 97.37% | 97.24% | 97.11% | 96.88% | 96.62% | 96.39% | 96.07% | |
| 68.38% | 67.74% | 67.17% | 66.15% | 65.99% | 65.05% | 64.53% | 63.25% | 62.74% | 61.73% |
Table 11.
Number of non-tree edges scanned by FPP(0) for matrices of order of 300 × 6000 with different density levels.
Table 11.
Number of non-tree edges scanned by FPP(0) for matrices of order of 300 × 6000 with different density levels.
| 5% | 10% | 15% | 20% | 25% | 30% | 35% | 40% | 45% | 50% | |
|---|---|---|---|---|---|---|---|---|---|---|
| Low | 69,133 | 65,415 | 61,975 | 57,305 | 53,638 | 47,722 | 44,590 | 40,668 | 35,545 | 31,668 |
| Medium | 461,875 | 420,106 | 372,561 | 400,387 | 387,020 | 353,904 | 334,324 | 292,983 | 250,507 | 251,315 |
| High | 42,210 | 32,766 | 41,489 | 36,797 | 40,709 | 29,770 | 29,068 | 29,206 | 26,317 | 26,811 |
Table 12.
FCT(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for complete sets of triples.
Table 12.
FCT(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for complete sets of triples.
| 65 | 730 | 2745 | 6860 | 13,825 | 24,390 | 39,305 | 59,320 | 85,185 | 117,650 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Num. | 29 | 355 | 1385 | 3498 | 7130 | 12,460 | 20,110 | 30,433 | 43,911 | 61,348 |
| Num. | 2 | 23 | 50 | 81 | 113 | 151 | 191 | 233 | 277 | 318 |
Table 13.
FCT(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for profiles of binary trees with varying number of trees and 100 taxa.
Table 13.
FCT(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for profiles of binary trees with varying number of trees and 100 taxa.
| 10,000 | 20,000 | 30,000 | 40,000 | 50,000 | 60,000 | 70,000 | 80,000 | 90,000 | 100,000 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Num. | 7453 | 14,985 | 22,566 | 29,957 | 37,532 | 44,959 | 52,500 | 59,630 | 66,984 | 74,780 |
| Num. | 667 | 1156 | 1648 | 2104 | 2555 | 3201 | 3460 | 3966 | 4643 | 4975 |
Table 14.
FCT(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for profiles of binary trees with varying number of taxa and 100 trees.
Table 14.
FCT(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for profiles of binary trees with varying number of taxa and 100 trees.
| 10,000 | 20,000 | 30,000 | 40,000 | 50,000 | 60,000 | 70,000 | 80,000 | 90,000 | 100,000 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Num. | 7473 | 15,098 | 22,711 | 30,403 | 37,903 | 45,636 | 53,083 | 60,638 | 68,336 | 75,946 |
| Num. | 683 | 1340 | 2037 | 2722 | 3387 | 4047 | 4759 | 5394 | 6017 | 6743 |
Table 15.
FPP(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for medium-density matrices of order .
Table 15.
FPP(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for medium-density matrices of order .
| 5% | 10% | 15% | 20% | 25% | 30% | 35% | 40% | 45% | 50% | |
|---|---|---|---|---|---|---|---|---|---|---|
| Num. | 27,592 | 27,793 | 26,951 | 27,358 | 27,358 | 27,126 | 26,711 | 26,839 | 26,590 | 26,003 |
| Num. | 924 | 926 | 940 | 972 | 969 | 983 | 983 | 976 | 976 | 979 |
Table 16.
FPP(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for high-density matrices of order .
Table 16.
FPP(0): Number of subtrees of a size at most two versus number of subtrees of size greater than two for high-density matrices of order .
| 5% | 10% | 15% | 20% | 25% | 30% | 35% | 40% | 45% | 50% | |
|---|---|---|---|---|---|---|---|---|---|---|
| Num. | 31,130 | 30,709 | 31,054 | 30,443 | 30,194 | 29,359 | 29,393 | 28,839 | 27,603 | 26,696 |
| Num. | 882 | 908 | 905 | 907 | 901 | 908 | 892 | 912 | 900 | 879 |
Table 17.
FCT(0): Number of subtrees of sizes and greater than 8 for profiles of binary trees with and varying n.
Table 17.
FCT(0): Number of subtrees of sizes and greater than 8 for profiles of binary trees with and varying n.
| 10,000 | 20,000 | 30,000 | 40,000 | 50,000 | 60,000 | 70,000 | 80,000 | 90,000 | 100,000 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 7012 | 14,188 | 21,324 | 28,550 | 35,603 | 42,883 | 49,848 | 56,924 | 64,181 | 71,334 |
| 2 | 461 | 910 | 1387 | 1853 | 2300 | 2753 | 3235 | 3714 | 4155 | 4612 |
| 3 | 158 | 287 | 434 | 586 | 721 | 862 | 1016 | 1154 | 1288 | 1448 |
| 4 | 88 | 157 | 239 | 315 | 393 | 472 | 560 | 615 | 689 | 794 |
| 5 | 56 | 101 | 149 | 197 | 242 | 290 | 350 | 392 | 431 | 495 |
| 6 | 38 | 70 | 102 | 137 | 166 | 197 | 238 | 266 | 294 | 348 |
| 7 | 27 | 48 | 71 | 101 | 125 | 143 | 174 | 190 | 218 | 245 |
| 8 | 20 | 37 | 54 | 78 | 95 | 113 | 132 | 151 | 167 | 182 |
| >8 | 296 | 640 | 988 | 1308 | 1645 | 1970 | 2289 | 2626 | 2930 | 3231 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fernández-Baca, D.; Liu, L. Tree Compatibility, Incomplete Directed Perfect Phylogeny, and Dynamic Graph Connectivity: An Experimental Study. Algorithms 2019, 12, 53. https://doi.org/10.3390/a12030053
AMA Style
Fernández-Baca D, Liu L. Tree Compatibility, Incomplete Directed Perfect Phylogeny, and Dynamic Graph Connectivity: An Experimental Study. Algorithms. 2019; 12(3):53. https://doi.org/10.3390/a12030053
Chicago/Turabian StyleFernández-Baca, David, and Lei Liu. 2019. "Tree Compatibility, Incomplete Directed Perfect Phylogeny, and Dynamic Graph Connectivity: An Experimental Study" Algorithms 12, no. 3: 53. https://doi.org/10.3390/a12030053
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.