1. Introduction
Cooling water systems remove heat from components and industrial equipment. Water cooling is typically used for cooling large industrial facilities, such as power plants, chemical factories and petroleum refineries. Cooling systems operate by transferring heat from a heat source, such as a power plant or industrial equipment, to a heat sink, typically water. The most common cooling water systems are once-through systems and open recirculating cooling towers. The former is a common choice, where available water resources are abundant, and thus, they usually use natural water from a lake, river or sea. However, materials are susceptible to fouling in natural waters. Fouling consists of biofouling, corrosion products and precipitation. When seawater is the cooling fluid, the phenomenon is accentuated fundamentally due to the strong corrosive nature of salt water and to its elevated biological activity. Biofouling, inorganic fouling and scaling can reduce heat transfer and enhance corrosion. Corrosion reduces the lifetime of the systems and also further increases scaling and fouling by providing uneven surfaces more favorable for the micro-organisms to attach. Fouling on the other hand reduces the heat exchanging capacity of the system.
Natural water sources contain diverse species of micro-organisms: bacteria, algae and fungi. The interaction between the micro-organisms and the cooling system material surfaces challenges the operational performance of the system by such phenomena as microbial-influenced corrosion (MIC) and microfouling. Microbes can generate conditions that increase corrosion through the alteration of pH and redox potential, the excretion of corrosion-inducing metabolites, direct or indirect enzymatic reduction or oxidation of corrosion products and the formation of biofilms that create corrosive microenvironments [
1]. Scaling, where insoluble chemical compounds precipitate on the surfaces, is also possible, although not directly connected with the presence of micro-organisms. Nevertheless, all of these surface processes impair the transfer of heat in the system, thus decreasing the cooling efficiency. Microfouling and inorganic scaling precedes the attachment and growth of macrofouling organisms, such as mussels and macroalgae. Macro-organisms are known to favor surfaces where biofilm is already formed [
2,
3]. Macrofouling reduces the flow rate of cooling water systems and may reduce also the cooling capacity.
Micro-organisms are typically combated by the use of biocides; in the seawater environment, the common approach is chlorination [
4] by, e.g., the addition of hypochlorite compounds [
5,
6]. However, the proper dosing and timing is a balance between the efficiency of the treatment and the avoidance of the corrosive effect and environmental stress caused by the toxic effect of chlorination treatment. Corrosion may be induced by hypochlorite compounds because they are strong oxidizing agents that easily accelerate the anodic oxidizing reaction and induce the localized attack of passivating alloys, such as stainless steels.
Besides using chemicals, microfouling may be combated by the application of coatings. Special antifouling coatings have been developed [
7], with the focus on either surface characteristics, such as hydrophobicity or superhydrophobicity [
8], that aim to reduce the contact between the material and its environment, or antimicrobial tendency,
i.e., the inherent property to influence the microbial counts through interaction between the surface and the micro-organisms.
Seawater is a challenging environment for many metallic materials because of saline content and biological activity. Titanium alloys are commonly-used materials especially in heat exchangers because of their high corrosion resistance, strength and light weight [
9]. Steel or coated steel is often used as a pipe material. In this research, we examine biofouling and materials’ degradation in a brackish seawater environment using a range of test materials, both uncoated and coated. Here, the fouling and corrosion resistance of titanium alloy (Ti-6Al-4V), super austenitic stainless steel (254SMO) and epoxy-coated carbon steel (Intershield Inerta160) were studied in the absence and presence of hypochlorite.
This is one of the very few contributions that involves brackish seawater as the studied environment and, thus, improves the understanding of the interactions of micro-organisms typical for such a low-salt water with cooling system material surfaces.
3. Discussion
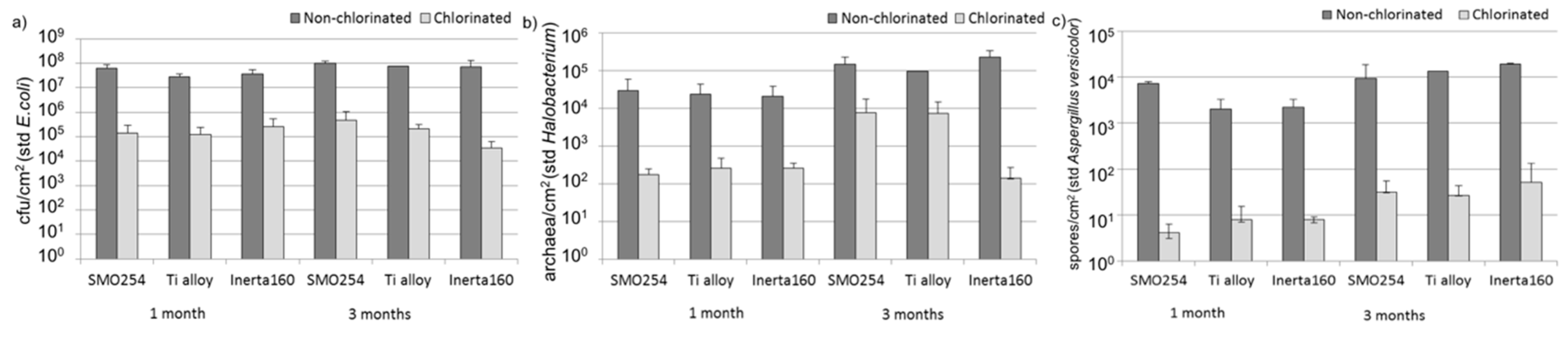
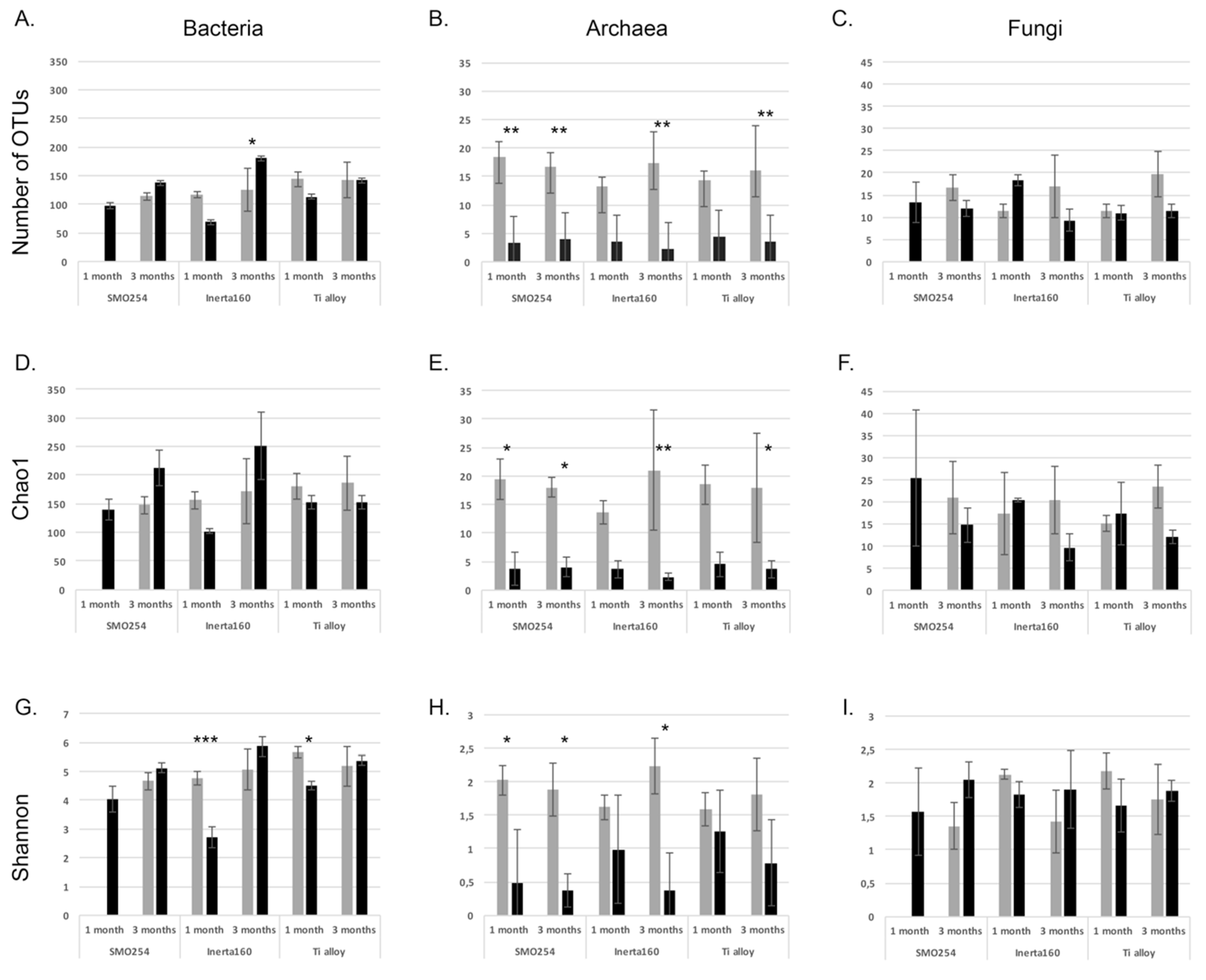
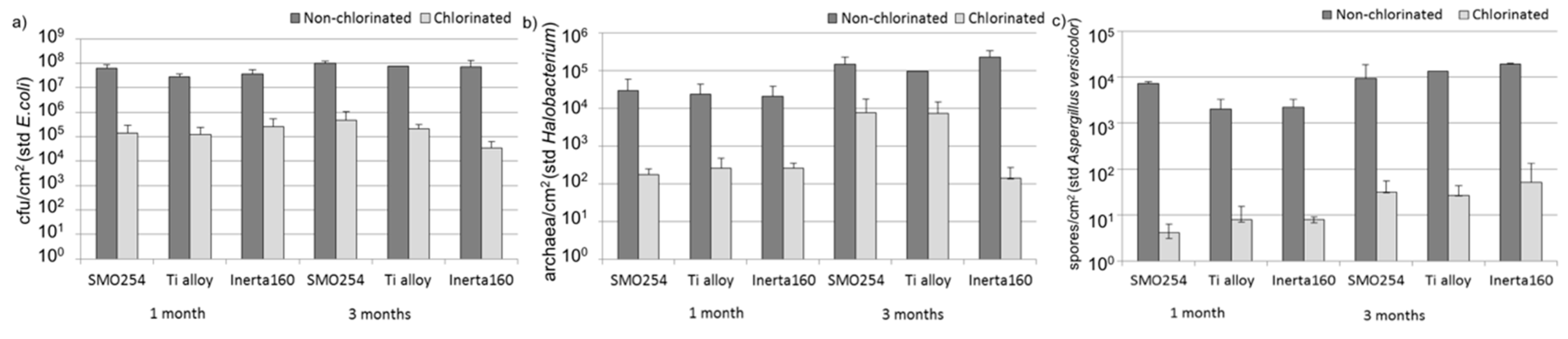
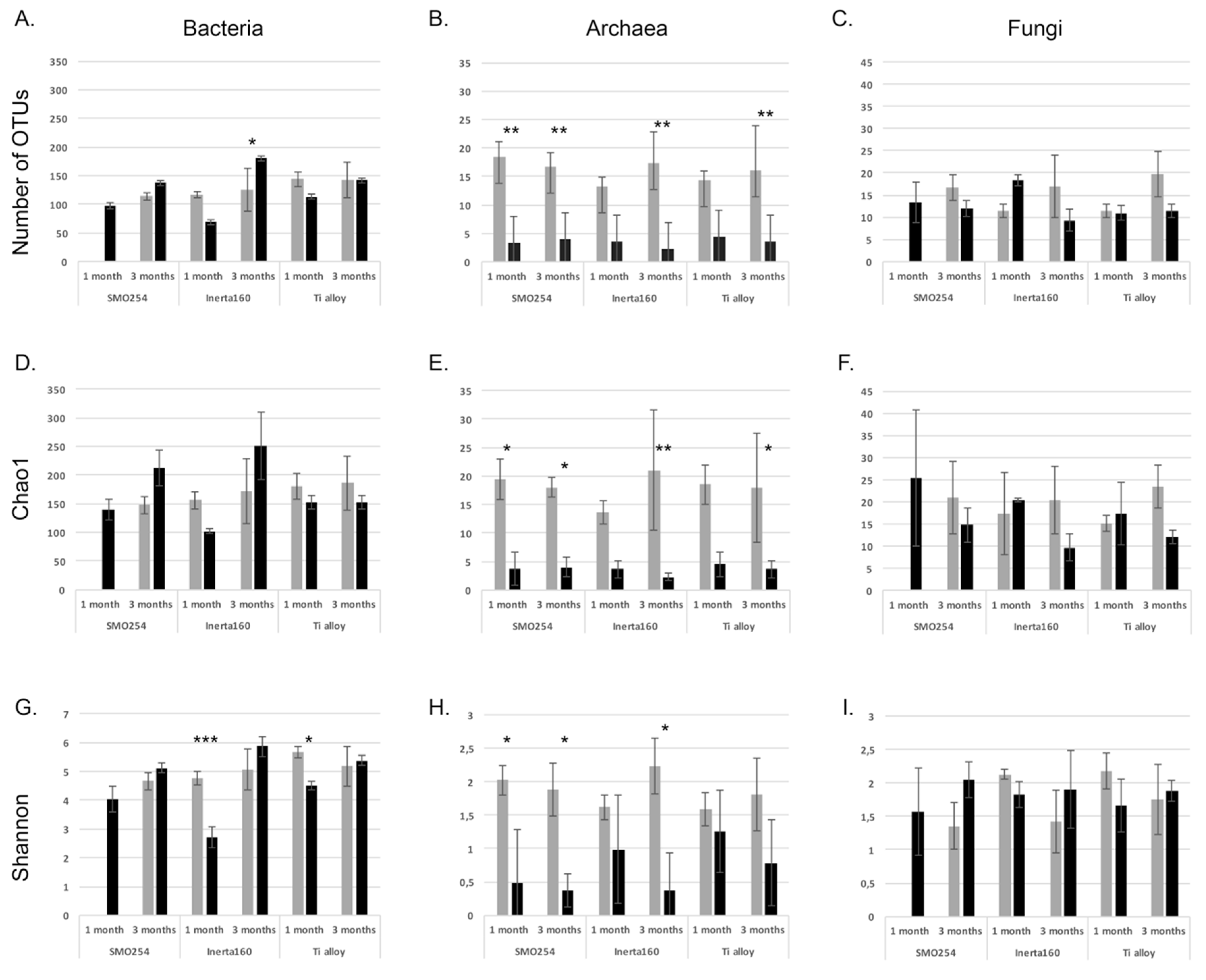
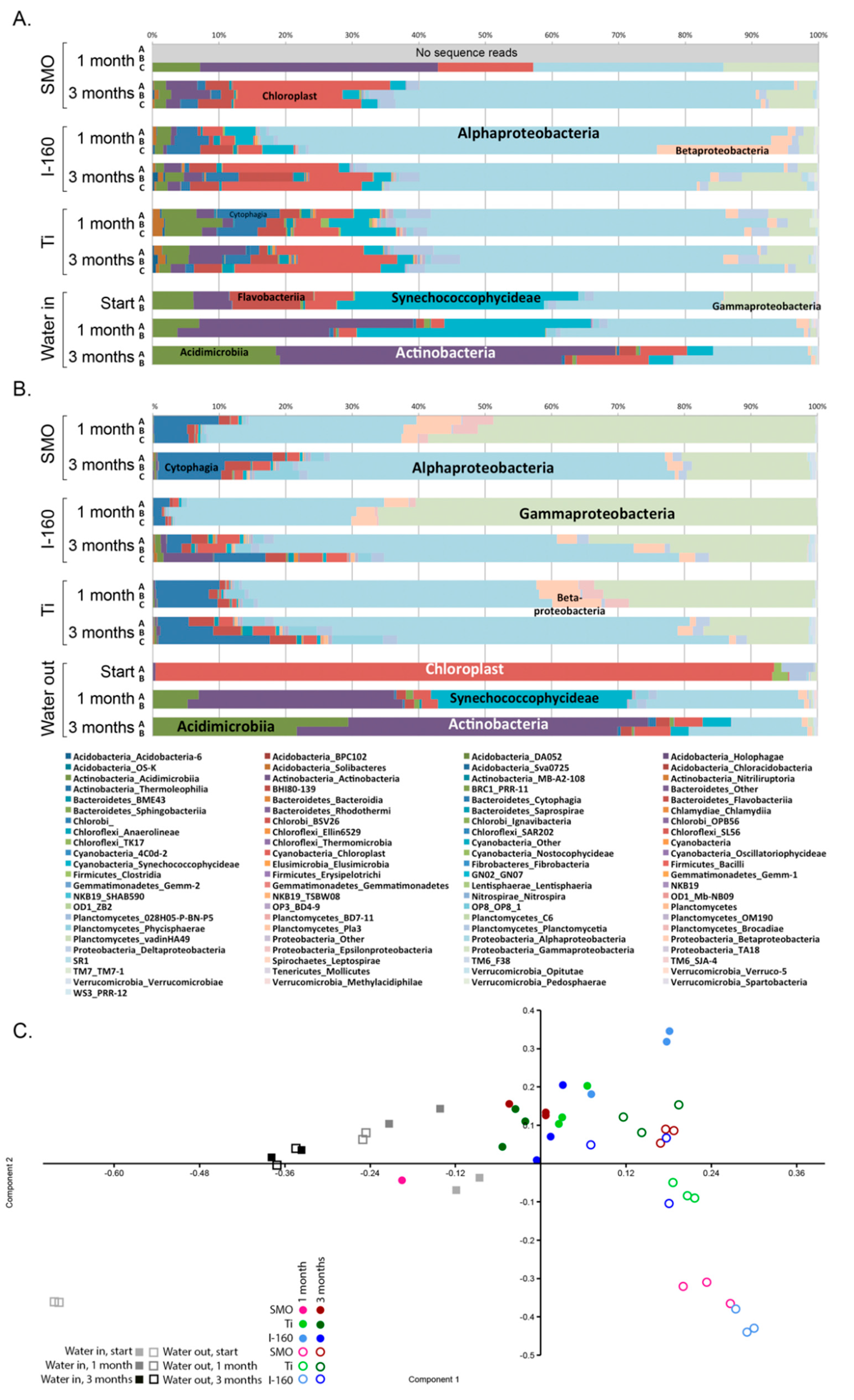
Cooling water systems utilizing natural waters are very demanding applications with respect to the reliable long-term performance of materials and coatings. The results obtained here demonstrated that biological fouling is intensive in cooling systems using brackish seawater in sub-arctic areas during the summer period. The microfouling comprised a vast diversity of bacteria, archaea, fungi, algae and protozoa. Our results demonstrate that chlorination was effective against biological fouling; up to a 10–1000-fold decrease in bacterial and archaeal number was detected. When interpreting the results from the quantitative PCR assay, it has to be borne in mind that the gene copy numbers vary in different microbial species. Thus, the results do not give absolute numbers, but still give an overview estimate of the trend and can be well used for comparing different samples. The coating, Inerta160, combined with chlorination further improved the antifouling effect. However, in non-chlorinated environments, the material selection did not have any significant effect on biofouling.
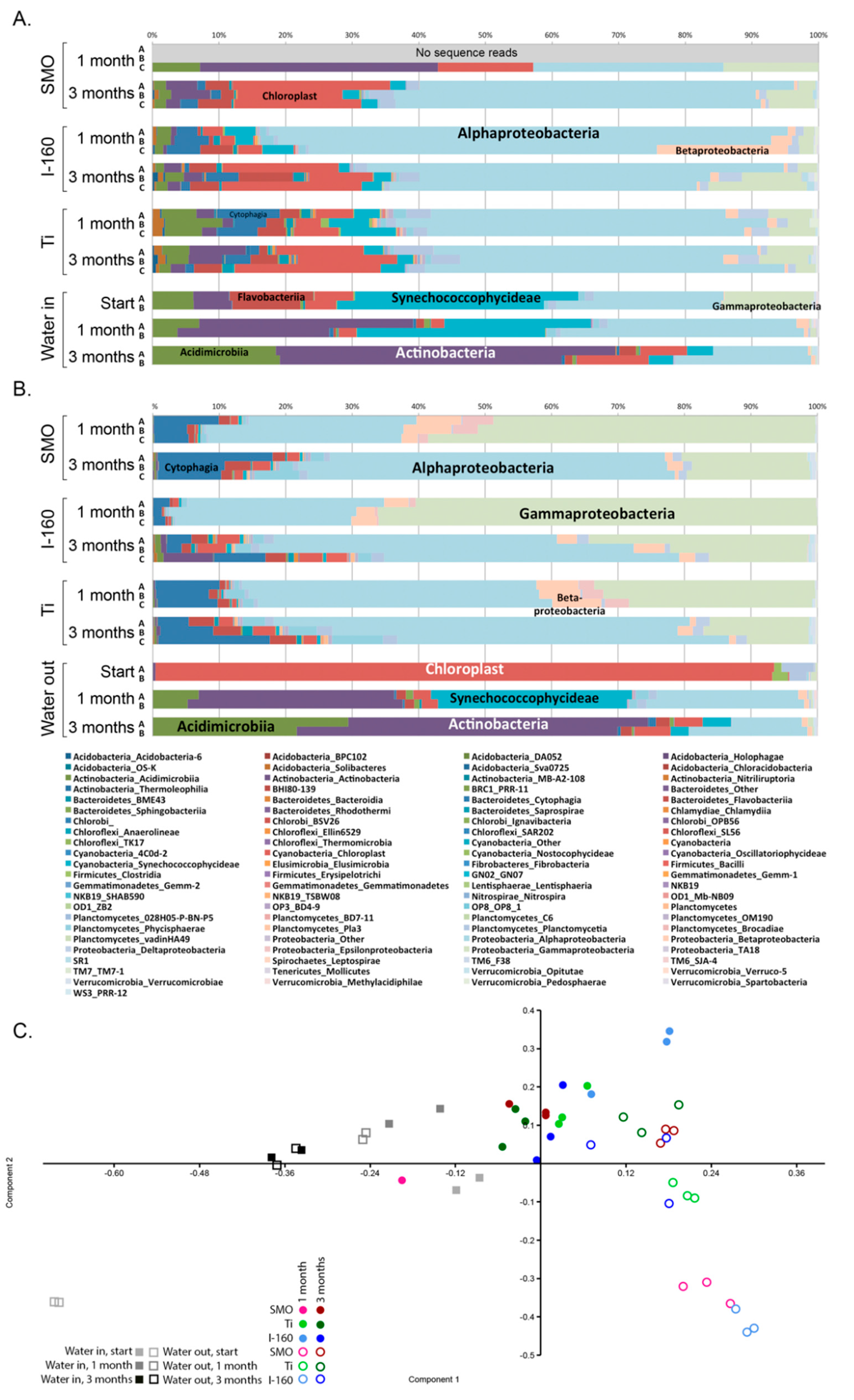
In addition to reducing the number of micro-organisms, the chlorination also had an effect on species diversity attached to surfaces compared to the non-chlorinated environment. Principal component analysis (PCA) was utilized to visualize the difference in sample clustering according to the species detected. In the PCA analysis, there was a clear division of biofouling species between the chlorinated and non-chlorinated environment. Alphaproteobacteria dominated in the biofilm in the non-chlorinated environment regardless of the material, and the Rhodobacteraceae family was abundant on the surfaces of non-chlorinated samples. The relative abundance of alphaproteobacteria increased with longer exposure time, and the biofilm became thicker over time. In chlorinated samples, the majority of the alphaproteobacteria belonged to the families Kordiimonadaceae, Rhodobacteraceae and Erythrobacteraceae. Many bacteria belonging to Rhodobacteraceae are phototrophic iron oxidizing [
10], whereas the Kordiimonadaceae family is often detected from marine environments where they degrade hydrocarbons [
11]. Members of the Erythrobacteraceae family are also commonly detected from marine systems and may oxidize organic carbon compounds [
12]. Erythrobacteraceae may also oxidize manganese, a process that has been linked to localized corrosion of stainless steels [
13]. Bacteria belonging to both Rhodobacteraceae and Erythrobacteraceae have previously been identified as forming biofilms on copper-based antifouling coating in marine systems [
14]. In fact, Erythrobacteraceae bacteria had 10–100-times higher relative abundances in the biofilms of the chlorinated systems compared to the non-chlorinated ones (
Table S2).
Gammaproteobacterial
Pseudoalteromonas species were tolerant to chlorination and formed the majority of the biofilm in the chlorinated environment. Some
Pseudoalteromonas species produce compounds that have antibacterial, algicidal, antifungal or antiviral activity and may thus have a beneficial position when competing for space in biofilm [
15]. On the other hand, certain
Pseudoalteromonas species attract other organisms to surfaces [
16].
Pseudoalteromonas species originating from marine water have been shown to cause corrosion of duplex stainless steel where the bacteria induced local corrosion by decreasing the chromium content under the biofilm and accumulated chloride ions on the metal surfaces [
17]. The accumulation of chloride, as well as the apparent enrichment of
Pseudoalteromonas in the biofilms of the 254SMO and Ti alloy coupons of our chlorinated systems indicate that this bacterium may not be susceptible to the chlorination treatment. The relative abundance of Bacteroidetes in biofilm in a chlorinated environment was also higher compared to the non-chlorinated system and the majority of these resembled hetero-organotrophic
Roseivigra species. After prolonged exposure time, the composition of the biofilms became more uniform on the surface of different materials.
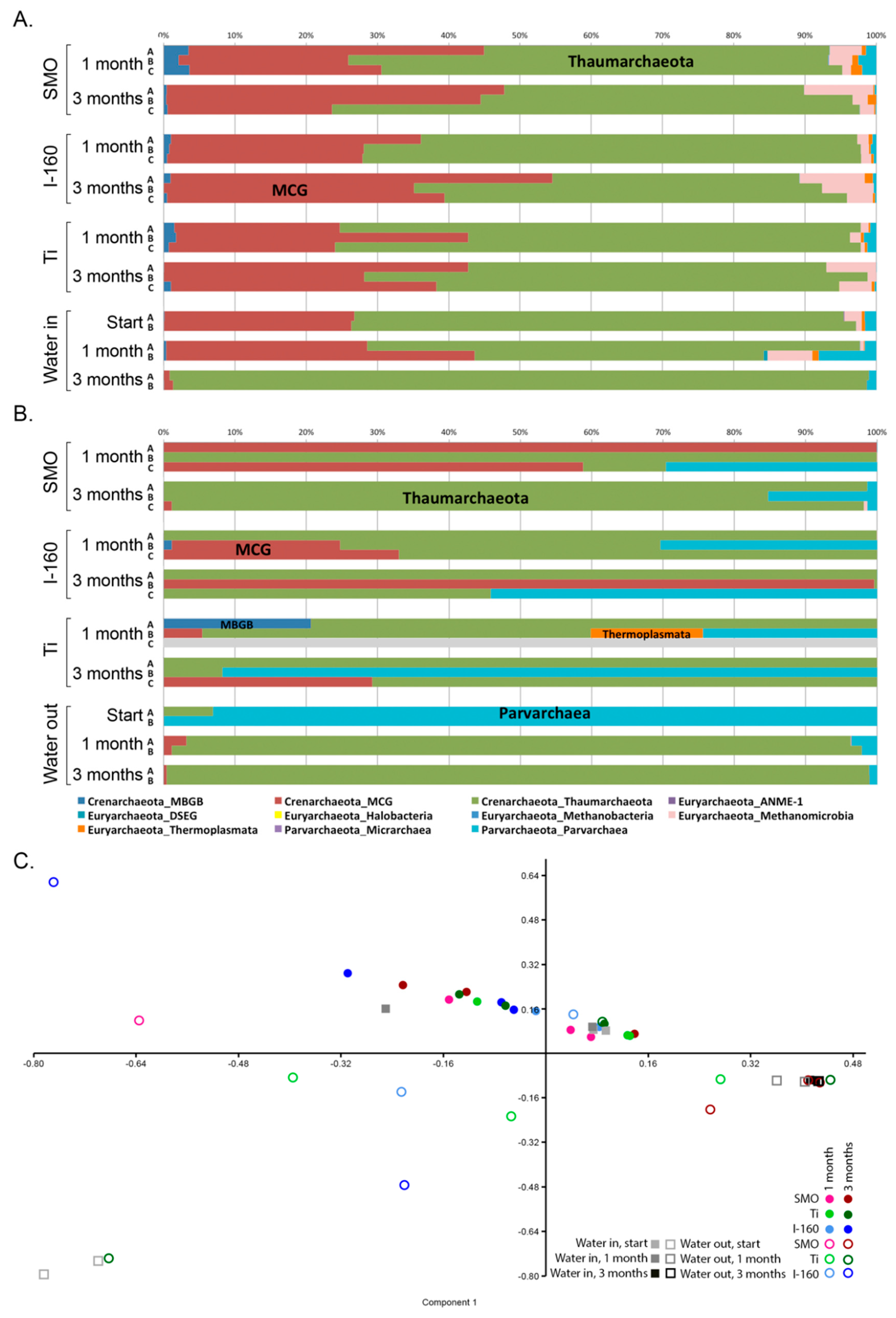
Most studies concentrate mainly on the bacterial fraction of biofouling, and thus, the role of the archaeal community in biofouling is not well known. Archaeal diversity on surfaces was significantly affected by chlorination, but although a small part of the archaeal community consisted of hydrogenotrophic methanogenic archaea, in both the chlorinated and non-chlorinated treatments, Crenarchaeotal lineages (Thaumarchaeota and MCG) dominated. Our results agree with previous studies on the MIC of oil pipes [
18,
19] or carbon steel in marine environments [
20], where the archaeal counterparts causing corrosion have belonged to different hydrogenotrophic methanogenic species. However, to the best of our knowledge, Crenarchaeotal (Thaumarchaeota and MCG) or Parvarchaeota species have not been associated with the MIC of stainless steel or titanium, consistent with our observations. It is very possible that these archaea do not primarily cause corrosion, but they may facilitate MIC and biofilm formation by capturing inorganic carbon into the organic form and recycle nitrogen compounds (ammonia) from decomposing senescent biofilms for the benefit of the other microbial community [
21,
22] members that cause MIC. In addition, the MCG archaea have been shown to have genes encoding extra-cellular protein-degrading enzymes [
23], which would assist in the degradation of organic matter, such as old biofilms, releasing ammonia for the Thaumarchaeota, which may then produce nitrite and nitrate for the nitrite- and nitrate-reducing bacteria.
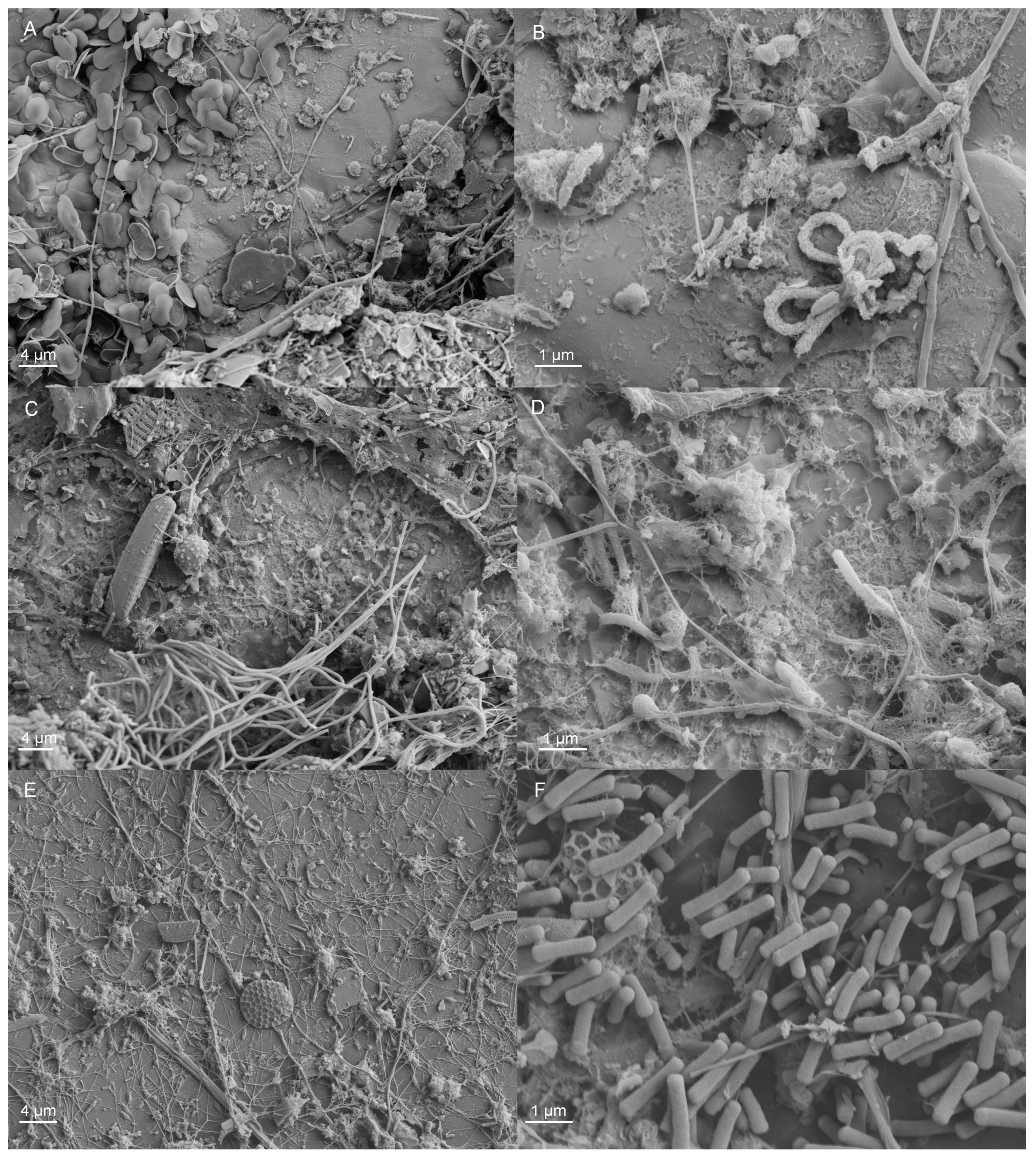
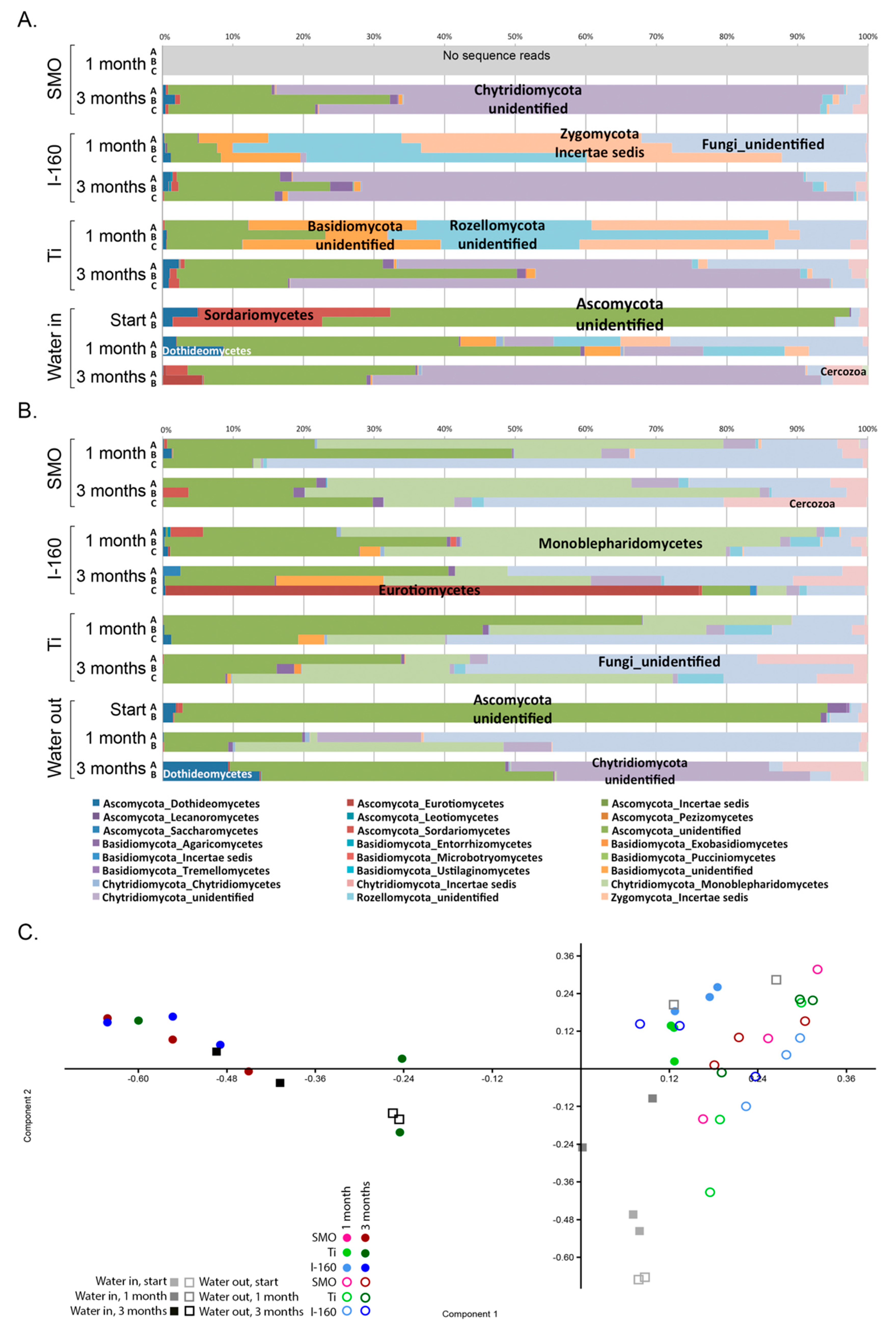
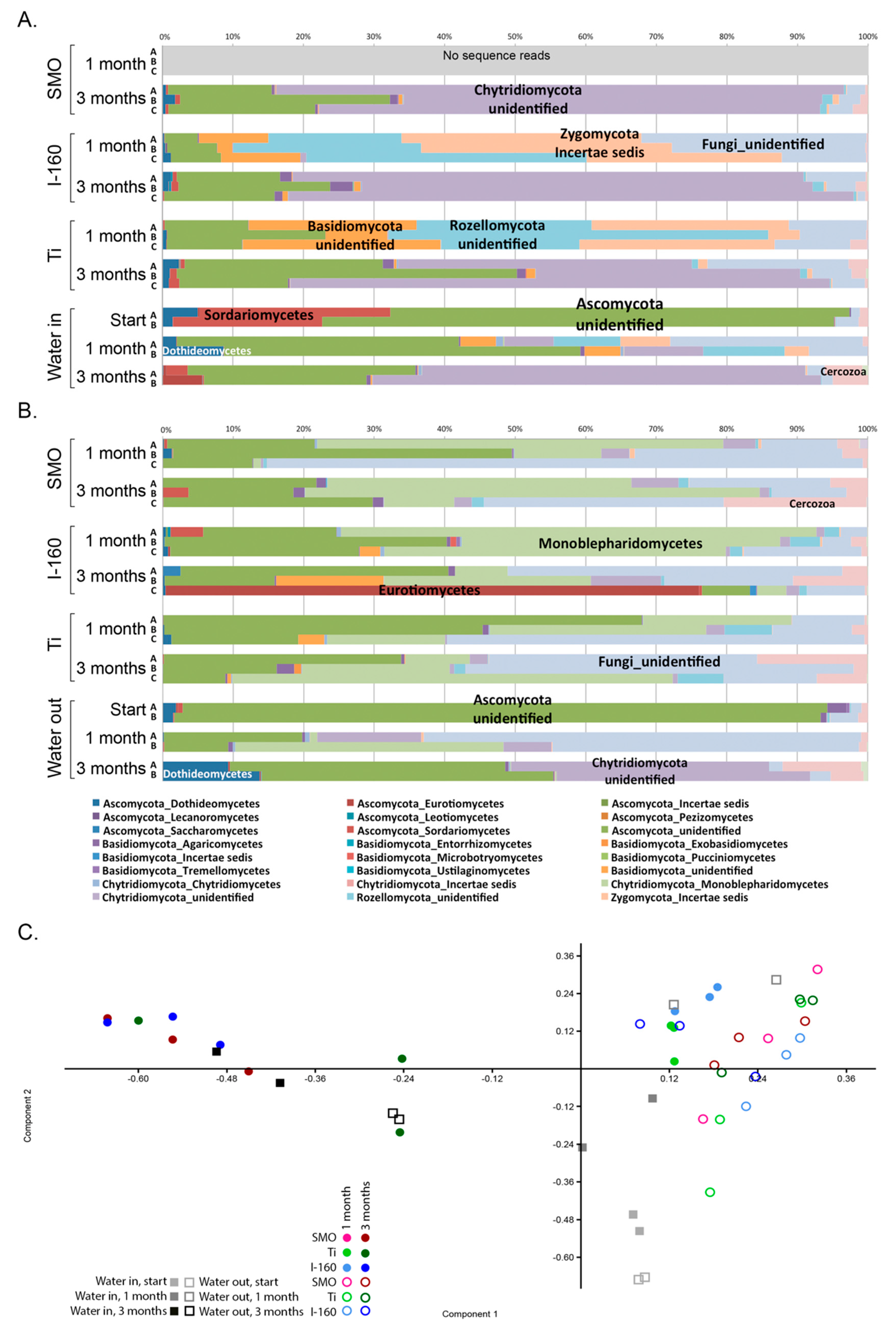
Fungi were generally detected at lower concentrations than the bacteria and archaea, but the fungal communities on the specimen surfaces and in the water were surprisingly diverse. Despite their low abundances, fungi may play a great role in the formation of biofilms, because through their metabolism, the fungi consume oxygen efficiently, thus providing anoxic microhabitats for, e.g., anaerobic sulfate reducers or the methanogenic archaea to operate. Ascomycetes fungi, such as
Aspergillus niger, have been reported to cause corrosion on magnesium alloy in artificial seawater [
24]. Diverse fungi also caused severe corrosion damage to storage tanks and transporting pipelines for oil [
25]. Ascomycetes oxidize manganese [
26], and fungi secrete different types of exudates, such as organic acids and enzymes, which may locally lower the pH and induce localized corrosion and cracking [
27]. In agreement with our results, Pereira
et al. [
28] also found a variable effect of chlorination on the inhibition of Ascomycetes fungi, and Ascomycetes were detected in both the chlorinated and non-chlorinated systems. Clearly, species belonging to Chytridiomycota, Rozellomycota and Zygomycota were more susceptible to chlorination than the Ascomycota, while species of the Monoblepharidomycetes were detected only in the chlorinated treatments.
Chlorophyta chloroplasts were relatively abundantly detected in the bacterial 16S rRNA gene sequence data, especially in the non-chlorinated systems, indicating the presence of eukaryotic green algae. Chlorophyta are common inhabitants of fouling biofilms on different surfaces (e.g., [
29,
30]). Hydrogenase-containing Chlorophyta are able to use cathodic hydrogen and may, under acidic conditions, contribute to corrosion events [
31]. Other non-fungal eukaryotes detected were the Cercozoa protists, which were especially abundant in the chlorinated systems. These protozoans are also common in biofilms and may serve as hosts for pathogenic bacteria, such as Legionella [
32].
Besides biological fouling, also the extent and nature of organic and inorganic fouling changed due to chlorination. Because of chlorination, an overall decrease in the extent of fouling was detected. Carbon was detected in all analyses, indicating that the presence of biofilm also contributed to the results, but it is clear that much of the information also originated from inorganic deposits on the surface. The results indicated that in non-chlorinated systems, the biofilm development facilitated inorganic scaling, particularly SiO
2. Indeed, e.g., SiO
2 is a common inorganic constituent in scales developed in seawater environments [
33,
34,
35], with the biofilm formation known to further enhance its deposition on the surfaces [
33]. This is also the case here, because carbon and silicon were systematically detected in the same analyses. In the chlorinated systems, the contribution of “softer” inorganic constituents, such as Ca, Mg and Al, increased, and SiO
2 that forms hard and tenacious deposits correspondingly decreased, thus demonstrating the viability of chlorination in the scale modification. This is an important aspect with respect to long-term performance and maintenance of seawater cooling systems, because the removal of SiO
2 scales may be challenging due to their high hardness [
35].
The composition of the coating was studied prior to exposure and was detected to contain heterogeneous composite structure, having abundant fillers. The most common fillers were oxides of magnesium and silicon with niobium, tantalum, molybdenum and barium. It is possible that barium is included in the coating as barium sulfate, a common anti-fouling agent used due to its favorable price [
36]. In EDS spectra, the peaks of molybdenum and sulfur overlap, hiding the sulfur in the results. Furthermore quartz, SiO
2, is a frequently-used low-price filler that improves the wear resistance of the coating. Another important practical finding is the enhanced cracking of the epoxy coating Inerta160 in the chlorinated system. It is expected that the cracks did not reach the substrate during the three-month exposure period, because no corrosion products of steel were detected in the epoxy coating. However, the operation of epoxy coatings relies on the barrier effect,
i.e., the non-conducting layer physically insulating the substrate (here steel) from the environment, which was challenged by chlorination. Earlier, UV radiation [
37] and thermal oxidation [
38] have been linked with the cracking of epoxy-based coatings.
The overall experiment period was short, three months at the longest, and lasted for only one warm period (summer season) when pulse chlorination is utilized to prevent biofouling. The chlorination was ended for the colder winter season, the same time as the experiment period ended. The rate of biofilm formation is known to correlate with sea water temperature, which changes with the season [
39]. This is especially important in sub-arctic areas, such as the Baltic Sea area, where this study was conducted, where the temperatures change drastically between, e.g., summer and winter periods. To obtain reliable results of the effects of the fouling and chlorination on materials’ performance, experiments lasting for several chlorination periods are needed. Nevertheless, during this three-month experiment period, it was already demonstrated that chlorination has a clear effect on the fouling and species composition of the biofilms developing on the surfaces.
4. Materials and Methods
4.1. Materials
Two of the materials were uncoated: super austenitic stainless steel 254SMO and titanium-based alloy Ti-6Al-4V. The composition of stainless steel 254SMO was 0.01 wt% C, 0.20% N, 20.0% Cr, 18.0% Ni, 6.1% Mo, 0.7% Cu and Fe (balance). For Ti-6Al-4V, the composition was 6.0 wt% Al, 4.0% V, ≤0.25% Fe, ≤0.2% and Ti (balance). Test materials also included epoxy-coated carbon steel. Red epoxy was an abrasion-resistant epoxy Intershield 163 Inerta160 (International Marine Coatings, Gateshead, UK) intended for corrosion protection applications in the underwater areas of ice-going vessels. The coatings were applied as recommended by the manufacturer. In each case, the coupon size was 25 × 75 mm.
Coupons were cleaned with FreeBact-20 (AquaFix, Saltsjöbaden, Sweden) and sterile MQ water and air-dried. Each biofouling cell contained 8 coupons of each material. Three parallel coupons were used for material analyses, three coupons for molecular biological analyses and two for microscopy analyses. The coupons were photographed prior to the experiment.
4.2. Experiment Setup
The test materials were placed in a biofouling cell in the cooling water system of a nuclear power plant located in the western coast of Finland. Two cells were installed in the cooling water cycle before the chlorination point and two cells after the chlorination point. The water flow through the biofouling cell was set at 0.01 m·s−1 using rotameters. The chlorination of the cooling water system was started in July 2015 and lasted 106 days. For chlorination, 15% of Na-hypochlorite was used with a final target concentration 3 mg·L−1. During the first 30 days, chlorination was applied for two hours with 10 h brakes between treatments and, after that, for one hour with 11 h breaks between. The experimental period started a week after chlorination treatment was initiated. One cell before the chlorination point and one cell after it were removed after 1 month (32 days) of exposure. The remaining two cells were removed after 3 months (90 days) of exposure. After the removal of the biofouling cells from the water cycle, the cells were sealed to retain the water inside them. The cells were dismantled aseptically in the laboratory.
4.3. Surface Characterization
Coupons for corrosion studies were immersed in 96% ethanol, air-dried and placed in glass desiccators. All coupons for corrosion analyses were photographed and examined with a stereomicroscope to verify the surface condition. The surface deposits of selected coupons were analyzed with a scanning electron microscopy (SEM) and energy-dispersive spectroscopy (EDS). Furthermore, the as-received Inerta160-coated steel specimens were subjected to investigations by SEM-EDS to disclose the original coating structure in the surface and cross-section. To enable the investigations, a thin layer of gold was sputtered on the coated specimens to make them electrically conductive. Field-emission (FE) SEM Zeiss ULTRAplus equipped with a Noran EDS system was employed for the investigations. Surface characteristics (hydrophobicity/hydrophilicity) of the Inerta160 coating were studied using an unexposed specimen by contact angle measurements.
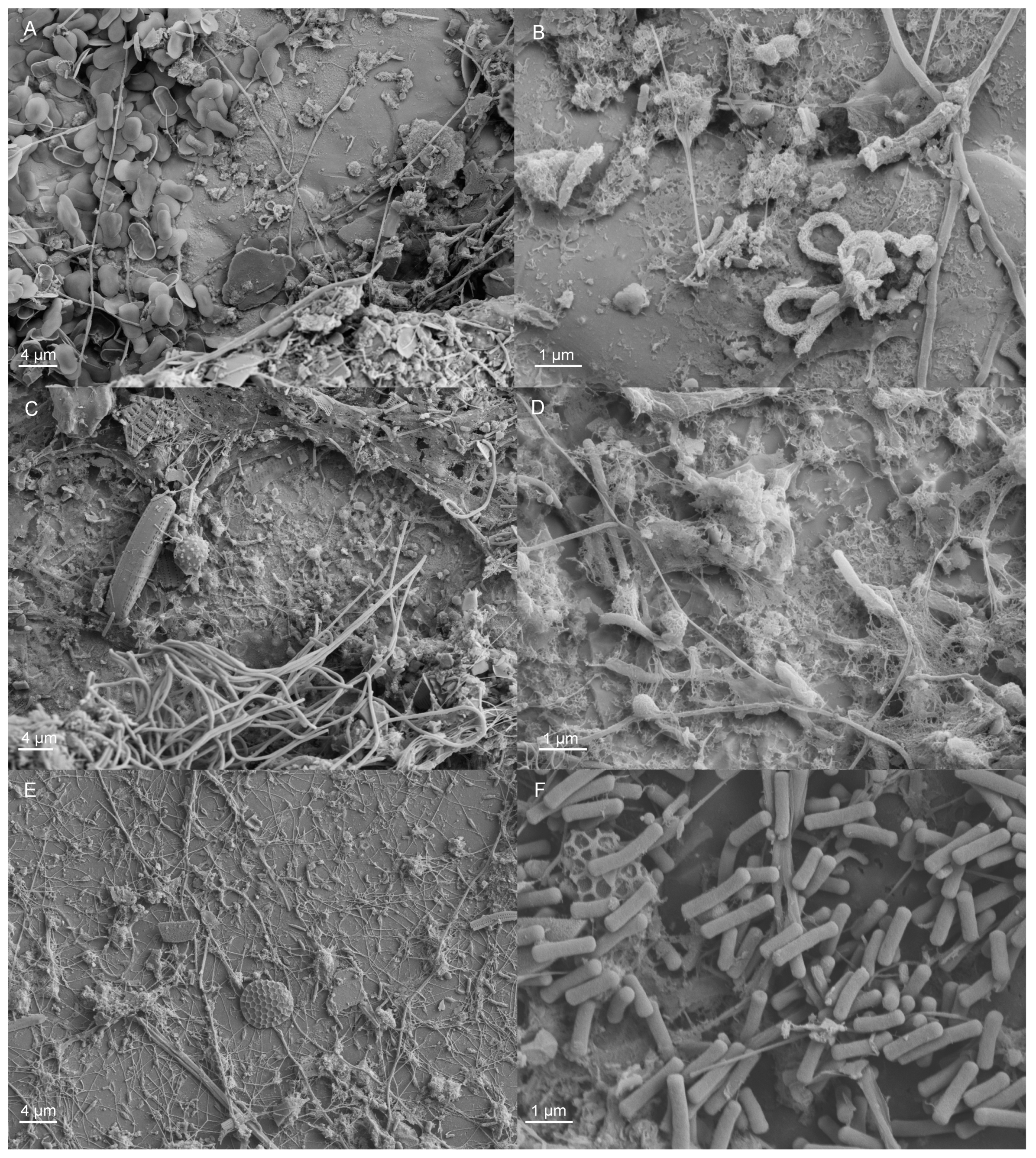
To visualize the biofilm formed on the surface, selected samples were fixed in phosphate (0.1 M, pH 7.2), buffered with 2.5% glutaraldehyde for 2 h and dehydrated with ethanol series followed by final drying in hexamethyldisilazane. The specimens were coated with Au/Pd (10–15 nm) and examined with a Zeiss SIGMA VP field-emission scanning microscope (FESEM; Carl Zeiss SMT GmBH, Oberkochen, Germany).
4.4. DNA Extraction
Coupons for microbiological analyses were subjected to biofilm extraction immediately after they were removed from the biofouling cell. The biomass from the coupon surfaces was removed by sonicating the coupons for 10 min (Bransonic 2210-DTH, Branson Ultrasonics, Danbury, CT, USA, 47 kHz, 70W) and thereafter vortexing for 1 min. The biomass suspensions were filtered into Sterivex-GP 0.22-µm filter units (Millipore, Billerica, MA, USA) and stored frozen (−80 °C) until DNA was extracted. Non-chlorinated and chlorinated water samples were filtered at the beginning of the experiment and at both sampling times as two 500-mL replicates to Corning PES (polyethersulfone) 0.22-µm filter units (Corning, New York, NY, USA), which were cut out with a sterile scalpel and stored frozen (−80 °C). Subsequently, the Sterivex filter units were aseptically broken with a hammer, and the Sterivex and Corning filters were cut into pieces with a sterile scalpel and placed into the lysing tube of the DNA extraction kit with sterile forceps. DNA was extracted using the Fast DNA Spin Kit for Soil (MP Biomedicals, Santa Ana, CA, USA) according to the manufacturer’s instructions, with the modification that the cells were homogenized in a FastPrep-24 instrument (MP Biomedicals, Santa Ana, CA, USA) at 6 m·s−1 for 3 min.
4.5. Quantitative PCR
The bacterial, archaeal and fungal biomass on the surface of the samples and in seawater was evaluated with quantitative PCR (qPCR) using the LightCycler 480 instrument (Roche Diagnostics, Basel, Switzerland). The DNA concentration of all samples was adjusted to ≤10 ng·μL
−1 prior to qPCR in order to avoid PCR inhibition. For bacteria, an approximately 200-bp fragment of the 16S rRNA gene was amplified with primers 358F (5′-CCT ACG GGA GGC AGC AG-3′) and 534R (5′-ATT ACC GCG GCT GCT GG-3′) [
40] and detected with SYBR green-based detection for double-stranded DNA. The amplification was done in a 10-µL reaction volume with KAPA Sybr Fast qPCR Master mix optimized for Roche LightCycler 480 (KAPA Biosystems, Wilmington, MA, USA), 150 nM of each primer and 1 µL of sample DNA. The amplification reaction consisted of initial denaturation at 95 °C for 15 min, 45 cycles with 10 s at 95 °C, 35 s at 57 °C and 30 s at 72 °C, a final elongation of 3 min at 72 °C and a melting curve analysis. As an external standard, a dilution series (100–10
8 cfu/µL) of
Escherichia coli VTT E-90418 genomic DNA was used. For archaea, an approximately 400-bp fragment of the 16S rRNA gene was amplified with primers A344F (5′-ACG GGG TGC AGC AGG CGC GA-3′) [
41] and A744R (5′-CCC GGG TAT CTA ATC C-3’) modified from 744RA [
42] and detected with SYBR green. The amplification was done in a 10-µL reaction volume with KAPA Sybr Fast qPCR Master mix optimized for Roche LightCycler 480 (KAPA Biosystems, Woburn, MA, USA), 300 nM of each primer and 1 µL of template DNA. The amplification reaction consisted of initial denaturation at 95 °C for 15 min, 45 cycles with 10 s at 95 °C, 35 s at 56 °C and 30 s at 72 °C, final elongation of 3 min at 72 °C and a melting curve analysis. As an external standard, a dilution series of
Halobacterium salinarum VTT E-103154
T genomic DNA was used (100–10
7 cells/µL). Fungal biomass was detected with TaqMan probe assay using ITS (internal transcribed spacer) as a target. Primers 5.8F1 (5′-AAC TTT CAA CAA CGG ATC TCT TGG-3′) and 5.8R1 (5′-GCG TTC AAA GAC TCG ATG ATT CAC-3′) and probe 5.8P1 (5′-CAT CGA TGA AGA ACG CAG CGA AAT GC-3’) were used [
43]. The amplification was done in 10 µL reaction volume with KAPA PROBE FAST qPCR Kit (KAPA Biosystems, Woburn, MA, USA), 500 nM of each primer, 200 nM of probe, and 1 µL of template DNA. The amplification reaction consisted of enzyme activation at 95 °C for 3 min and 40 cycles with 10 s at 95 °C (denaturation), 30 s at 62 °C (annealing) and 1 s at 72 °C (elongation). As an external standard, a dilution series of
Aspergillus versicolor VTT D-96667 genomic DNA was used (10–10
5 spores/µL).
4.6. Amplicon Library
The amplification libraries for high throughput sequencing with Ion Torrent PGM were prepared by PCR from the DNA samples. Bacterial 16S genes were amplified with primers S-D-Bact-0341-b-S-17/S-D-Bact-0785-a-A-21 [
44], targeting the variable region V3-V4 of the 16S rDNA gene, S-D-Arch-0349-a-S-17/S-D-Arch-0787-a-A-20 [
44], archaeal 16S genes with primers S-D-Arch-0349-a-S-17/S-D-Arch-0787-a-A-20 [
44], targeting the V4 region of the gene and fungal internal transcribed spacer (ITS) gene markers with primer pair ITS1 and 58A2R targeting the fungal ITS1 region [
45,
46]. PCR amplification was performed in parallel 25-μL reactions for every sample containing 1× MyTaq™ Red Mix (Bioline, London, UK), 20 pmol of each primer, up to 25 μL molecular-biology-grade water (Sigma, St. Louis, MO, USA) and 2 μL of template. The PCR program consisted of an initial denaturation step at 95 °C for 3 min, 35 cycles for bacteria and fungi and 40 cycles for archaea of 15 s at 95 °C, 15 s at 50 °C and 15 s at 72 °C. A final elongation step of 30 s was performed at 72 °C. The PCR products were verified with agarose gel electrophoresis. Amplicons were sent to Ion Torrent sequencing with PGM equipment (Bioser, Oulu, Finland), and amplicons were purified before sequencing at Bioser.
4.7. Sequence Processing and Analysis
The sequence reads obtained from Ion Torrent sequencing were subjected to quality control using the QIIME-software Version 1.8 [
47] using a minimum quality score of 20, minimum and maximum sequence length of 200 bp and 600 bp, respectively, maximum primer mismatch of 2 nucleotides (nt) and maximum homopolymer stretches of 8 nt. Adapters, barcodes and primers were removed from the sequence reads, and chimeric sequence reads were removed from the dataset with the USEARCH-algorithm [
48] by
de novo detection and through similarity searches against the Greengenes reference dataset (Version gg_13_8) [
49] with bacterial and archaeal sequences, as well as the UNITE (User-friendly Nordic ITS Ectomycorrhiza Database) reference dataset (Version sh_taxonomy_qiime_ver7_97_s_31.01.2016) [
50] with fungal sequences.
The sequences were grouped into Operational Taxonomic Units (OTUs), following the open-reference OTU-picking protocol of QIIME-software. First, all reads that failed to hit the Greengenes or UNITE reference database with a minimum of a 60% identity threshold were discarded as sequencing errors. Subsequently, closed-reference OTUs were picked at 97% clustering identity against the Greengenes or UNITE database, and
de novo OTUs were picked from a randomly subsampled sequence subset that failed the closed-reference OTU-picking stage. Next, singleton OTUs,
i.e., OTUs that were represented by a single sequence, were filtered from the dataset. Finally, taxonomy from the domain level to the species level was assigned to OTUs via representative OTU sequences with the Ribosomal Database Project (RDP) classifier algorithm at a minimum of 80% confidence [
51] with bacterial and archaeal sequences. With fungal ITS sequences, taxonomic assignments were made by the Basic Local Alignment Search Tool (BLAST) algorithm with a maximum E-value of 0.001 [
52]. Unassigned and unidentified sequences were filtered from the datasets before alpha diversity estimations. Alpha diversity indices were calculated on randomly subsampled and filtered datasets containing 1243, 1674 and 1624 sequences for ITS, bacteria and archaea, respectively.
The sequences were deposited in the European Nucleotide Archive (ENA,
https://www.ebi.ac.uk/) under accession number PRJEB14388.
4.8. Statistics Analyses
Significant differences of the qPCR means were tested with variance analysis using sampling time, surface material and chlorination as fixed factors and the subsequent Tukey’s post hoc test using IBM SPSS Statistics Version 21 software. The results were log-transformed to fulfil the assumptions of variance analysis.
Differences in the number of OTUs detected from the different biofilm samples, the Chao1 richness estimates and Shannon indices were tested based on normalized sequence data using One-way ANOVA in the PAST3 program [
52]. We tested significant differences of the mean between chlorinated and non-chlorinated samples using Levine’s test for the homogeneity of variance, Kruskal–Wallis and Tukey’s Q test. The difference in community composition between all samples was tested with principal component analysis of 100 bootstrap repeats on the taxonomic relative abundance data using the PAST3 program.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}