Gelatin-Based Hydrogels Promote Chondrogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stem Cells In Vitro

 , ,
, ,
Abstract
: Due to the weak regeneration potential of cartilage, there is a high clinical incidence of articular joint disease, leading to a strong demand for cartilaginous tissue surrogates. The aim of this study was to evaluate a gelatin-based hydrogel for its suitability to support chondrogenic differentiation of human mesenchymal stem cells. Gelatin-based hydrogels are biodegradable, show high biocompatibility, and offer possibilities to introduce functional groups and/or ligands. In order to prove their chondrogenesis-supporting potential, a hydrogel film was developed and compared with standard cell culture polystyrene regarding the differentiation behavior of human mesenchymal stem cells. Cellular basis for this study were human adipose tissue-derived mesenchymal stem cells, which exhibit differentiation potential along the adipogenic, osteogenic and chondrogenic lineage. The results obtained show a promotive effect of gelatin-based hydrogels on chondrogenic differentiation of mesenchymal stem cells in vitro and therefore encourage subsequent in vivo studies.1. Introduction
Articular cartilage has only weak regeneration potential [1]. Following traumatic or degenerative cartilage injury, this leads to a strong clinical demand for cartilaginous tissue surrogates. Since current strategies of autologous chondrocyte implantation suffer from chondrocyte de-differentiation following expansion in vitro [2], stem cells in conjunction with biomaterial scaffolds are quested to both accelerate and improve cartilage tissue repair [3]. The aim of the present study was to evaluate a new approach for cartilage tissue regeneration based on the utilization of mesenchymal stem cells (MSC) in contact to biodegradable gelatin-based hydrogels as supporting matrix.
The main constituent of the hydrogels developed is gelatin, a non-toxic, biodegradable and water-soluble protein derived from collagen, with the latter being a major component of mesenchymal tissue extracellular matrix (ECM). Gelatin retains informational signals including an arginine-glycine-aspartic acid (RGD) sequence, which promotes cell adhesion, proliferation and stem cell differentiation [4]. According to the production process, two types of gelatin with different isoelectric points can be distinguished [5]. Via acidic hydrolysis of collagen, gelatin type A with an isoelectric point of 6 to 8 is obtained. Conversely, an alkaline collagen treatment results in the development of gelatin type B with an isoelectric point of 4.7 to 5.3 [6]. In the present work, gelatin type B was selected as starting material, exhibiting superior biocompatibility compared to gelatin type A due to the different treatment conditions applied [7]. A characteristic property of gelatin is that it exhibits Upper Critical Solution Temperature behavior (UCST). Above a specific temperature threshold of 40 °C, gelatin can be dissolved in water by the formation of flexible, random single coils. Upon cooling, hydrogen bonding and Van der Waals interactions occur, resulting in the formation of triple helices. These collagen-like triple helices act as junction zones and thus trigger the sol-gel transition. However, in the scope of tissue engineering applications, chemical crosslinks still need to be incorporated to ensure stability around body temperature, as the physical crosslinks melt at these temperatures [8]. To date, several crosslinking strategies have already been elaborated. For example, Van Vlierberghe et al.[8] applied thiol functionalities that can be oxidized to disulfides in the presence of a suitable oxidant such as hydrogen peroxide. However, as unprotected thiols are sensitive to crosslinking in the presence of oxygen, hydrogel processing is not very straightforward in an air environment. Alternatively, a conventional carbodiimide crosslinking strategy is often applied, combining 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC) with N-hydroxysuccinimide (NHS) in order to covalently link the amines present in gelatin with the activated carboxylic acids [9–11]. In the latter case however, the processed hydrogel structure typically is incubated in a solution containing EDC and NHS. As a result, the crosslinking kinetics are of utmost importance, as gelatin also swells in an aqueous environment. Furthermore, genipin, a natural crosslinker extracted from gardenia fruit, has already been reported on as well with respect to gelatin crosslinking, as it combines two primary amines functions [12]. Unfortunately, this crosslinker typically requires longer crosslinking times. In the present work, gelatin was functionalized with methacrylamide side groups, enabling the chemical crosslinking upon applying UV-irradiation in the presence of a UV-active photo-initiator. The above-mentioned properties including its unique gel-forming ability render gelatin an interesting biopolymer towards tissue engineering applications.
MSC are progenitor cells resident in specialized niches within mesenchymal tissues in vivo and govern tissue regeneration by their intrinsic capacity for self-renewal and multipotent mesenchymal differentiation [13,14]. MSC were discovered within the bone marrow stroma [15] as adventitial cells on the outer surface of sinusoidal blood vessels [16]. Later on, the perivasculature was recognized to be the characteristic niche of all MSC independent of their tissue origin [17]. In this study, we used adipose tissue-derived MSC (adMSC). adMSC make up 7% of the cells found in liposuction-derived and collagenase-digested adipose tissue [18] and being present at a concentration of roughly 50,000 cells per ml tissue [19], which is 100-fold higher than that of bone marrow-derived MSC (bmMSC) [20]. Human adMSC were shown to directly differentiate into osteoblasts and endothelial cells in a non-union fracture model in SCID mice [20] and in immunodeficient rats [21]. In that manner, adMSC were demonstrated to reconstitute bone physiology in mice [20], rats [21,22], pigs [23] and dogs [24]. In human case studies, human adMSC were already successfully used to treat critical size calvarial [25] and maxilla defects [26] that, due to the limitation in the amount of auto-graftable bone tissue, could otherwise not have been healed. Finally, adMSC were demonstrated to engraft at long-term at the transplantation site and to migrate to and survive within the perivascular MSC niche [20–22,27]. Therefore, adMSC fulfill all MSC characteristics defined by the International Society for Cellular Therapy, the International Federation of Adipose Therapeutics and Sciences, and others [28–30] and thus are a crucial cell type for regenerative medicine.
2. Experimental Section
2.1. Preparation of Chemically Crosslinked Gelatin Hydrogels
Gelatin type B, isolated from bovine skin, was kindly supplied by Rousselot (Ghent, Belgium). Gelatin samples with an approximate isoelectric point of 5 and a Bloom strength of 257 were used. Methacrylic anhydride (MAA) was purchased from Aldrich (Bornem, Belgium) and was applied as received. Dialysis membranes Spectra/Por 4 (MWCO 12,000 – 14,000 Da) were obtained from Polylab (Antwerp, Belgium). 1-[4-(2-Hydroxyethoxy)-phenyl]-2-hydroxy-2-methyl-1-propane-1-one (Irgacure 2959) was a kind gift from Ciba Specialty Chemicals N.V. (Groot-Bijgaarden, Belgium). A LWUV-lamp model VL-400L (Vilber Lourmat, Marne La Vallee, France) with an intensity of 10 mW/cm2 and a wavelength range of 250 – 450 nm was applied in order to cure the hydrogels.
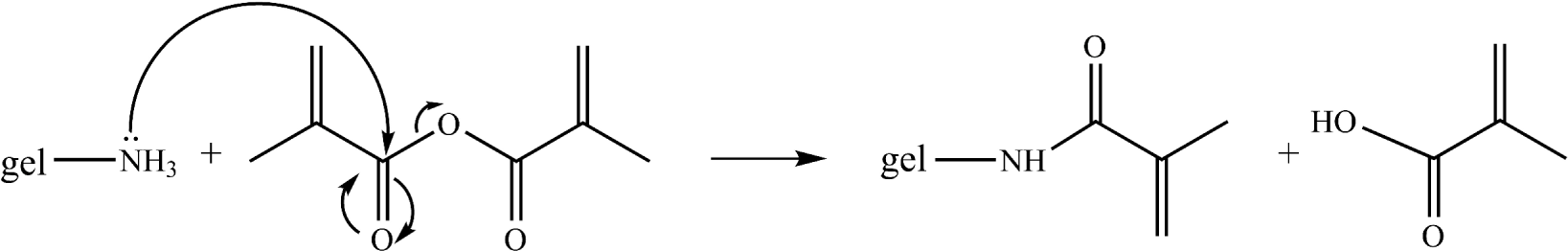
Gelatin was dissolved in PBS buffer (pH 7.3) at 40 °C. While vigorously stirring, 1 equivalent methacrylic anhydride relative to the amine functionalities present in gelatin B was added. After 1 h reaction time, the methacrylamide-modified gelatin (gel-MOD, see reaction scheme below) was purified via dialysis (MWCO: 12,000 – 14,000 Da), followed by freeze-drying. Interestingly, the amount of crosslinkable side groups can be adjusted by varying the amount of methacrylic anhydride added. In the present work, a modification degree of 60% was applied, which was obtained by the addition of one equivalent methacrylic anhydride.
2.2. Characterization of Gelatin-Based Hydrogels
2.2.1. HR-MAS Spectroscopy
High Resolution Magic Angle Spinning NMR spectroscopy (HR-MAS NMR) of the developed hydrogel films was performed on a Bruker Avance II 700 spectrometer (700.13 MHz) using a HR-MAS probe equipped with a 1H, 13C, 119Sn and gradient coil. The spinning rate was adjusted to 6 kHz. HR-MAS samples were obtained by placing a small amount of the freeze-dried hydrogels inside a 4 mm zirconium oxide MAS rotor (50 μL). A few microliter of deuterium oxide (D2O) were added to the rotor, allowing the samples to swell. The samples were homogenized by manual stirring prior to analysis. A teflon-coated cap was used to close the rotor.
2.2.2. X-ray Photoelectron Spectroscopy
X-ray photoelectron spectroscopy (XPS) was applied as a non-destructive, quantitative spectroscopic technique to determine the chemical composition of native and functionalized gelatin. A FISONS S-PROBE with a fine focus Al-Kα source (1486 eV) and a quartz monochromator developed by Fisons Instruments Surface Science (now Sanofi, Paris, France) was used for the measurement performed using a dedicated XPS instrument able to give the ultimate in performance, while providing a high sample throughput. All measurements were done in triplicate in a vacuum of at least 10−9 Pa. Wide and narrow-scan spectra were acquired at pass energies of 158 and 56 eV, respectively. The binding energy was calibrated by the C1s peak at 284.6 eV, and the utilized spot size was 250 μm on 1 mm. Data analysis was performed using S-PROBE software.
2.3. adMSC Isolation and Culture
If not stated otherwise, all plasticware was from Greiner Bio-One (Frickenhausen, Germany), whereas all reagents were from Sigma-Aldrich (Taufkirchen, Germany).
adMSC were isolated from adipose tissue of healthy patients having undergone tumescence-based liposuction. As described previously [31], adipose tissue was digested using Collagenase NB4 (SERVA, Heidelberg, Germany), a mixture of a neutral protease and the collagenases I and II (6 mg/mL in phosphate-buffered saline (PBS); PAA Laboratories, Cölbe, Germany). The cells thus liberated from the digested tissue were incubated over night under standard cell culture conditions (5% CO2 and 37 °C in a humidified atmosphere). CD34+ adventitial cells were isolated from total plastic-adherent cells using an antibody-magnetizable bead-based system (Life Technologies, Darmstadt, Germany) [32]. After three passages, adMSC were seeded into experimentation at 20,000 cells per cm2. Expression of MSC markers CD29, CD44, CD105, and CD166, and absence of CD14+/CD68+ monocytes/macrophages and CD31+ endothelial cells as well as absence of Mycoplasma species was confirmed.
The adMSC cultivation media used were unstimulated control medium (US) and chondrogenic differentiation stimulating medium (CS). US was Dulbecco’s Modified Eagle Medium (DMEM), high glucose, GlutaMAX-I, supplemented with 1% penicillin/streptomycin (100 U/mL and 100 μg/mL, respectively; both Life Technologies), and 10% fetal calf serum (FCS; PAN Biotech, Aidenbach, Germany). CS medium was serum-free US medium supplemented with 100 nM dexamethasone, 50 μg/mL ascorbic acid, 50 ng/mL insulin-like growth factor 1 (R&D Systems, Wiesbaden-Nordenstadt, Germany), 50 ng/mL TGF beta 1 (PeproTech, Hamburg, Germany) and 5 μg/mL insulin, 5 μg/mL transferrin and 5 ng/mL selenious acid (the latter three: BD Biosciences, Heidelberg, Germany). At confluence, culture with distinct differentiation-inducing media began, termed day zero, and took up to 28 days. Renewal of medium was every second to third day.
2.4. Ethical Statement
adMSC were isolated from patient tissue after written informed consent had been obtained that patients are willing to participate in this study. The study was approved by the ethics committee of Rostock University Medical Center ( http://www.ethik.med.uni-rostock.de/) under the registration numbers A17/2007 and A2013-0112, and it complies with the ethical standards defined by the World Medical Association Declaration of Helsinki.
2.5. Analysis of adMSC Chondrogenic Differentiation
2.5.1. Live-Dead Staining of adMSC on Hydrogels
adMSC were seeded onto TCPS and hydrogels and cultured for four weeks in US and CS medium. For live-dead staining, cells were incubated for 10 min under standard culture conditions with US medium supplemented with 3 μM Hoechst 33342, 500 nM propidium iodide and 1 μM calcein acetoxymethyl ester (calcein AM, from Life Technologies, Carlsbad, CA, USA). After medium exchange for US medium, cells were subjected to fluorescence microscopy (Carl Zeiss MicroImaging, Göttingen, Germany) using the blue, green and red emission filters. Emission maxima of the dyes are 460 nm (DAPI), 516 nm (calcein), and 617 nm (propidium iodide). Excitation maxima are 360 nm (DAPI), 496 nm (calcein) and 535 nm (propidium iodide).
2.5.2. Quantification of Chondrogenic Marker Gene Expression
Gene expression analysis was done after 14 and 28 days of adMSC culture on TCPS and hydrogels in US and CS medium, employing real-time reverse transcriptase polymerase chain reaction (real-time RT PCR). Briefly, cells were homogenized by centrifugation on QIAshredder columns (Qiagen, Hilden, Germany), and RNA was isolated from the lysate using the innuPREP RNA Mini Kit (Analytik Jena, Jena, Germany) following the manufacturer’s instructions. Subsequently, RNA was transcribed into cDNA using the QuantiTect Reverse Transcription Kit (Qiagen). Real-time PCR of transcribed samples was then performed on a 7500 Real Time PCR system using the Power SYBR Green PCR Master Mix (both Life Technologies). Expression of chondrogenic marker genes was determined using the primers listed in Table 1:
Both cDNA-specific primers were designed using the Primer3 engine [33]. Gene expression levels were determined by the ∆∆Ct method, normalizing to the expression of actin, beta (ACTB).
2.5.3. Staining of Cartilaginous Matrix Proteins
Acidic polysaccharides present in cartilage, which mostly are glycosaminoglycans, were stained after 28 days of adMSC culture using Alcian Blue, resulting in a greenish-blue staining of the extracellular matrix. Briefly, cells were fixed with 4% paraformaldehyde for 1 h at room temperature, washed 3× in 2% acetic acid and incubated over night at room temperature in Alcian Blue staining solution (1 mg/mL Alcian Blue 8 (pH = 2) mixed isovolumetrically with 80 mg/mL MgCl2, both in 0.9% NaCl). After washing 3× with 2% acetic acid and 3× with double distilled water (ddH2O), cells were subjected to bright-field phase contrast microscopy (Carl Zeiss MicroImaging).
2.6. Data Illustration and Statistics
Data are illustrated as box plots created using R [34]. Since the data obtained were in most cases not normally distributed, testing for significance in the difference between two datasets was done using the non-parametric Mann-Whitney U test. The level of significance was set to a p-value of lower or equal 0.05 (p ≤ 0.05) and determined using R.
3. Results and Discussion
3.1. Hydrogel Physico-Chemical Characterization
Crosslinking of a 10% (w/v) gelatin-methacrylamide (gel-MOD) solution was done via photo-initiation upon applying UV-irradiation in the presence of 2 mol% Irgacure 2959 (with respect to the methacrylamide side chains present). The polymer solutions were injected between two parallel glass plates separated by a 1 mm thick silicone spacer. The materials were allowed to form gels after applying UV-A irradiation.
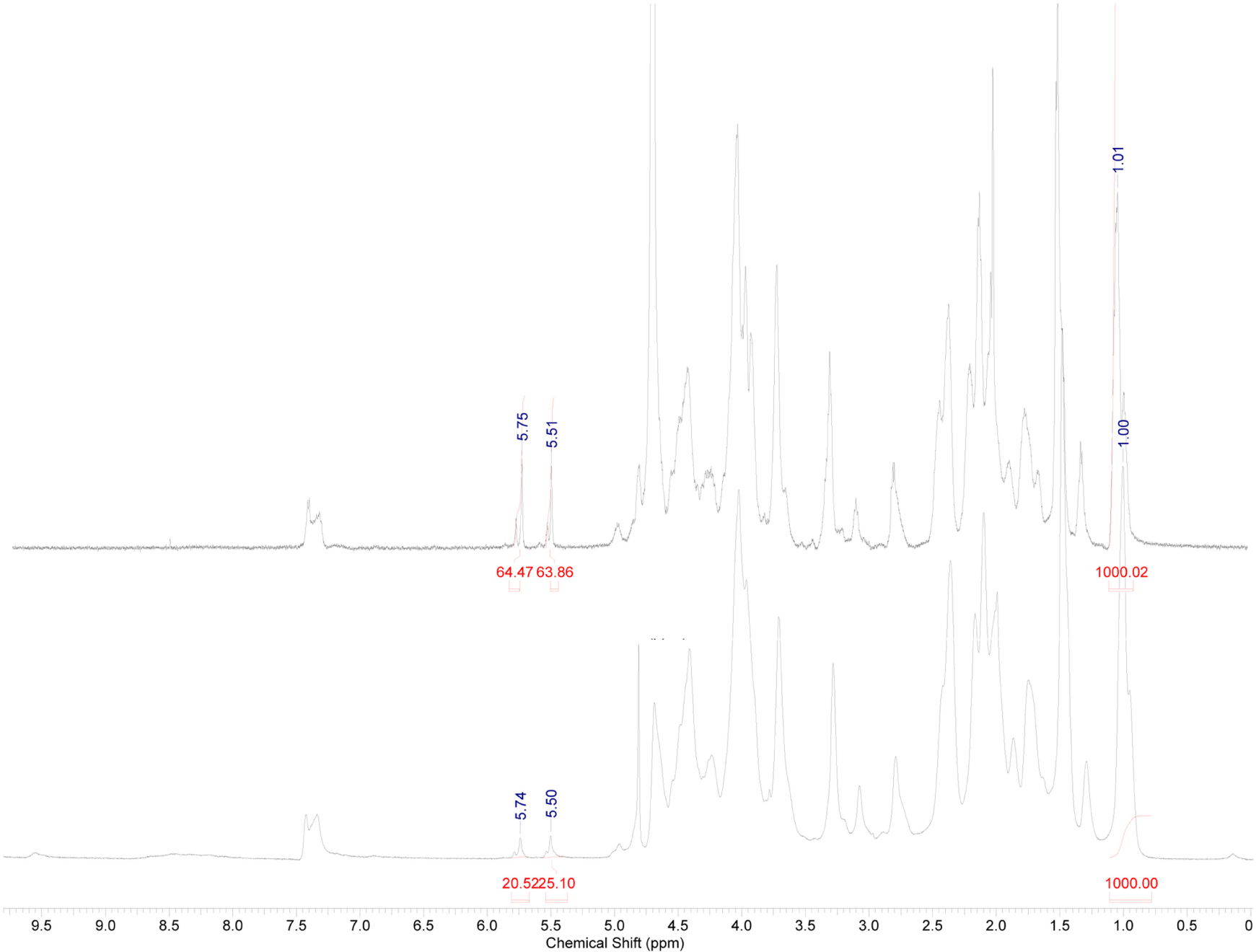
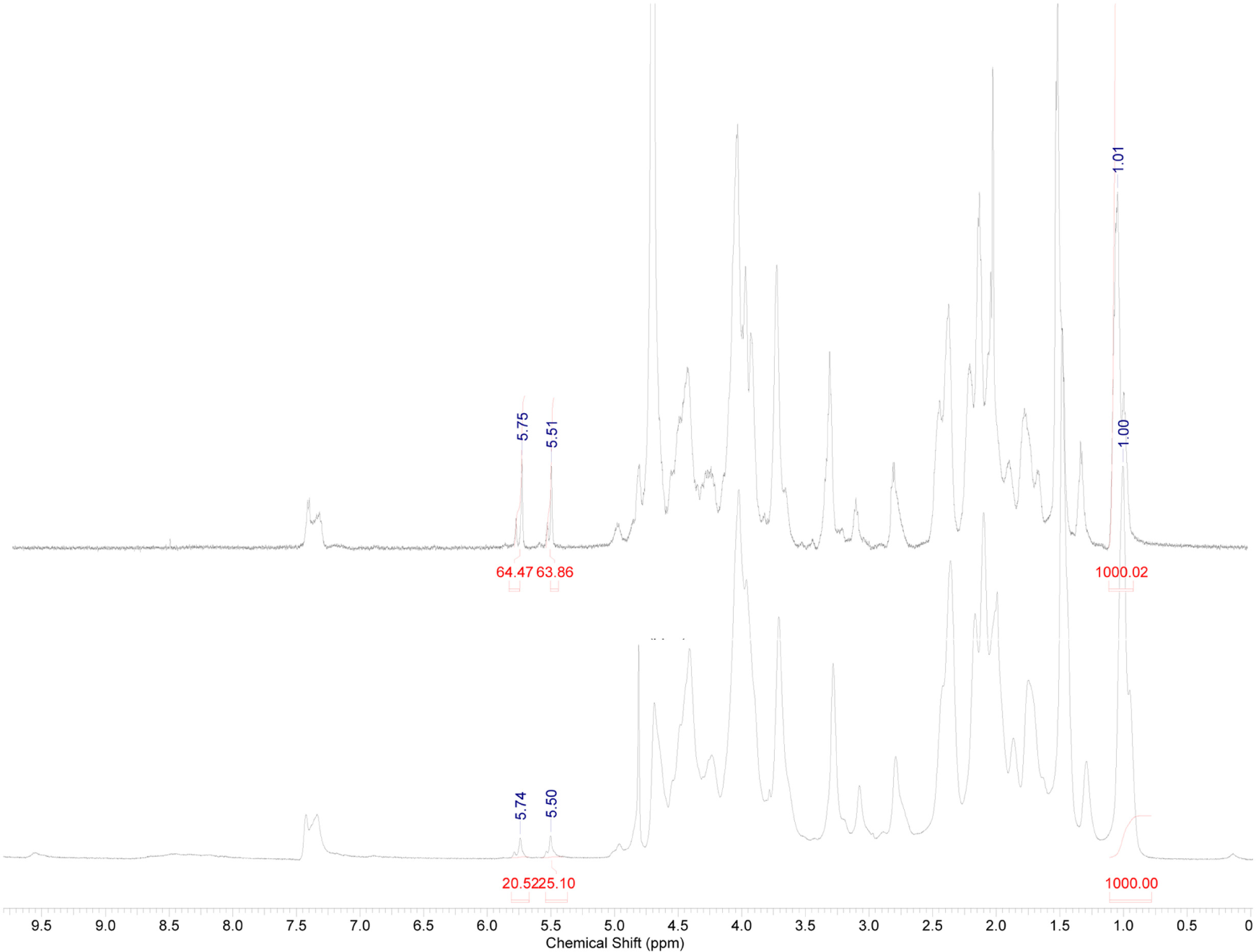
High Resolution Magic Angle Spinning Nuclear Magnetic Resonance (HR-MAS NMR) spectroscopy was applied as a straightforward technique to evaluate the crosslinking efficiency of the UV-cured hydrogels. Conventional 1H-NMR spectroscopy does not enable the characterization of water-insoluble polymer networks due to considerable line broadening resulting from the presence of dipolar couplings and magnetic susceptibility [35,36]. However, this broadening can be circumvented by rapidly rotating (in the order of kilohertz) the sample at a magic angle θ of 54.7° with respect to the static magnetic field [36]. In this way, the contribution of the line broadening effects defined by a (3 cos2 θ − 1)/2 orientation dependence is removed from the spectrum. Further decrease in line width is realized by swelling the hydrogel samples in a suitable deuterated solvent resulting in sufficient, solution-like, rotational mobility of the polymer [37]. As a consequence, highly crosslinked hydrogel materials exhibiting a reduced chain mobility will show broader peaks than less crosslinked materials [38].
The crosslinking efficiency can be determined by comparing the integration of the signal corresponding to the double bonds with the integration of a signal that remains chemically inert during crosslinking. Upon crosslinking, the introduced double bonds are consumed, enabling determination of the crosslinking efficiency (CE). The CE is calculated by comparing the intensity of the signals characterizing the protons of the double bonds, before and after crosslinking. Since different samples have to be compared, normalization is required using the inert signal (i.e., at 1.1 ppm). The crosslinking efficiency for the cured gelatin hydrogels is given by the following formula [37], where the superscript i stands for initial, non-crosslinked and c for crosslinked:
HR-MAS spectroscopy indicated an efficient UV-crosslinking for the gelatin-based hydrogels as a CE of 69% was obtained (see Figure 1).
In a subsequent part of the present work, the effect of the functionalization on the chemical composition of gelatin was determined via XPS measurements. Table 2 shows an overview of the atomic composition of gelatin and gel-MOD. The results indicate that Gel-MOD exhibits lower values compared to gelatin for the N/C ratio (i.e., 0.12 versus 0.19) and the N/O ratio (i.e., 0.56 versus 0.72), as anticipated based on the fact that methacrylamide moieties are introduced on the primary amines of (hydroxy-)lysine upon modification. For each primary amine in the gelatin backbone that is converted into methacrylamide, three additional carbons and one additional oxygen are introduced (see reaction scheme on page 1345). The results therefore indicate that the gelatin modification was successful.
3.2. adMSC Chondrogenic Differentiation on Hydrogels
3.2.1. Live-Dead Staining of adMSC on Hydrogels
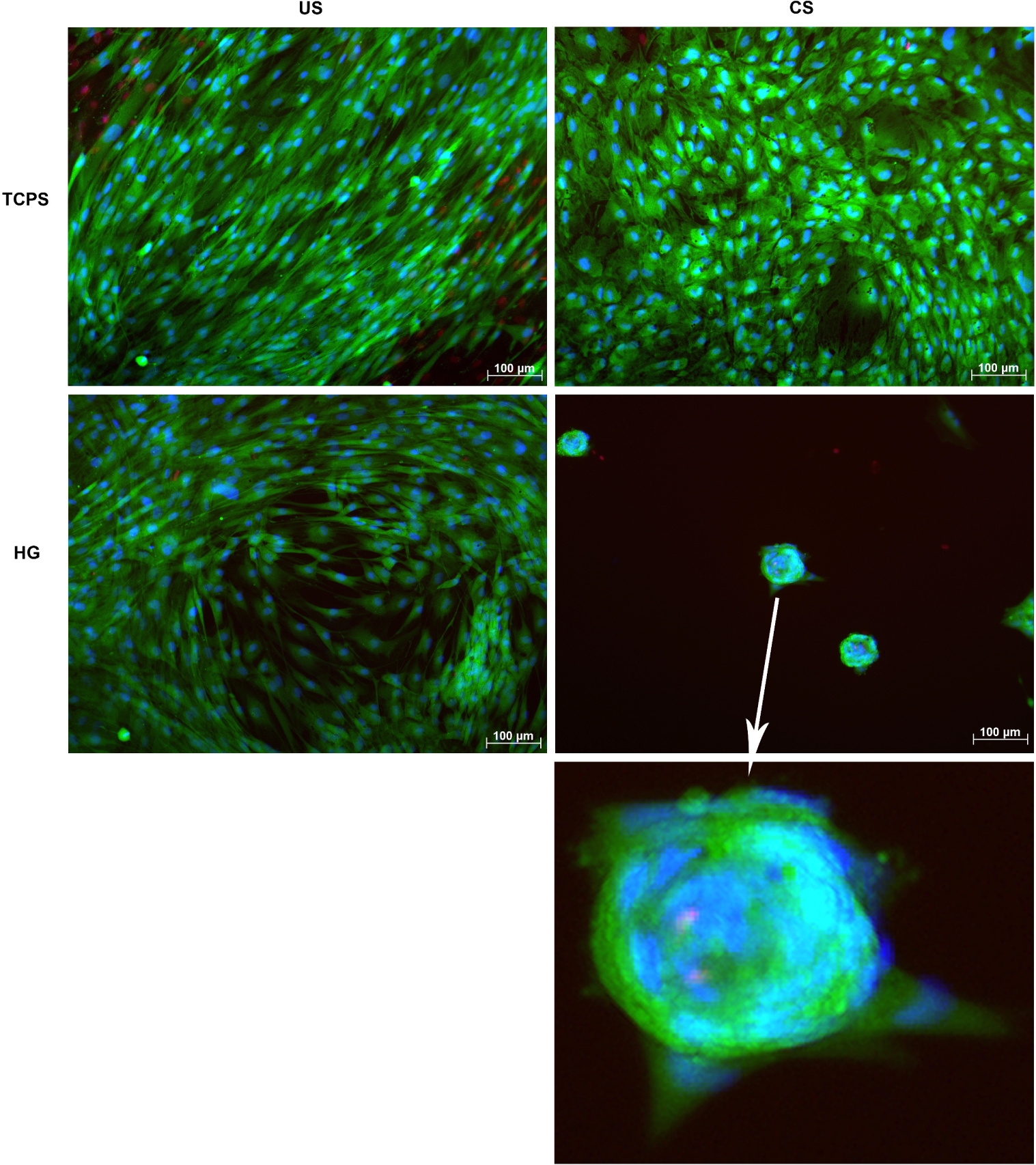
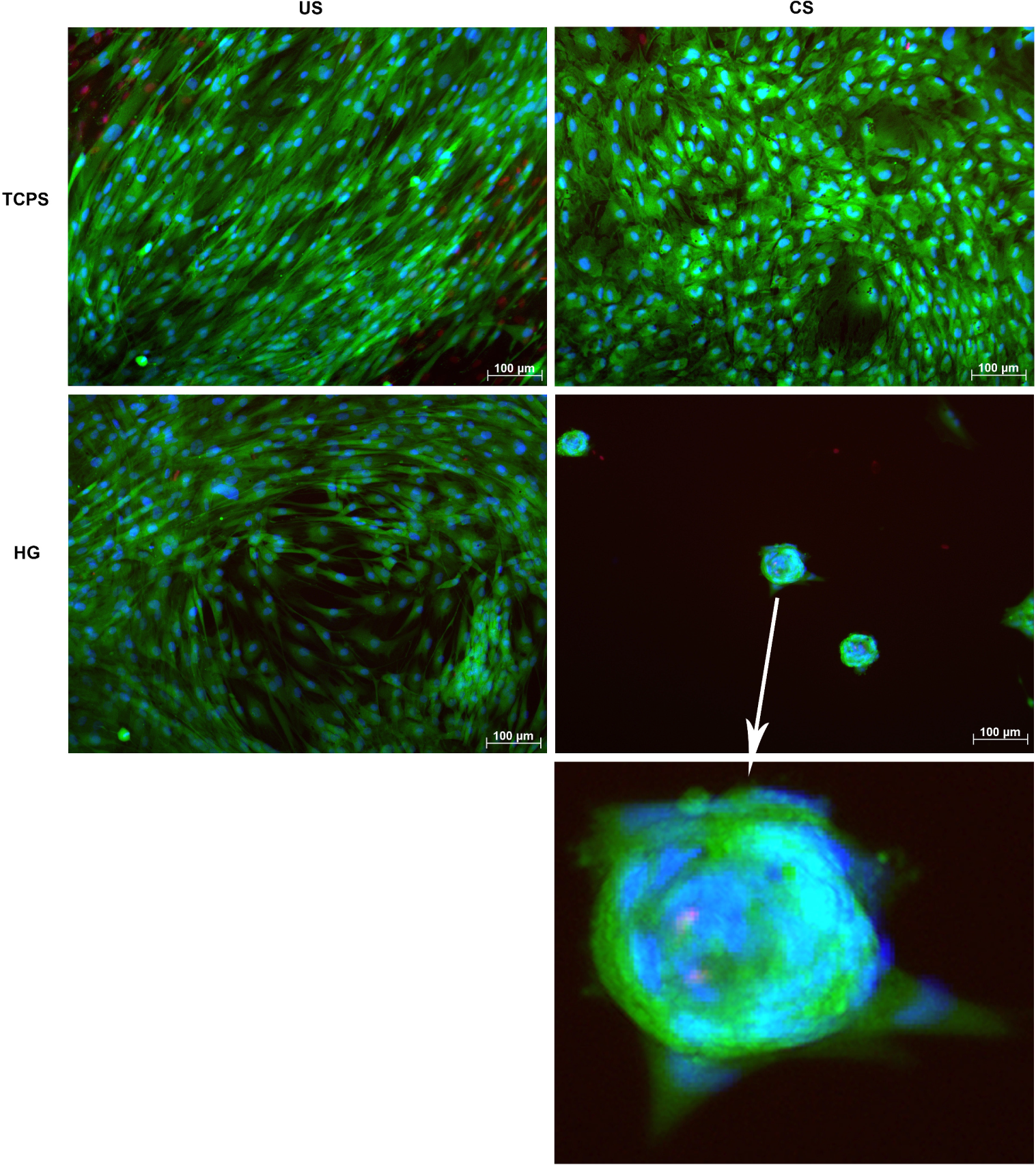
For testing of cytocompatibility and phenotypical changes upon differentiation-inducing conditions, human adMSC were live dead-stained following seeding onto hydrogels pre-incubated for 2 h with cell culture medium. Tissue culture polystyrene (TCPS) was used as a control adhesion surface. After 28 days of chondrogenic stimulation (CS) on TCPS, adMSC showed a more condensed and cobblestone-like phenotype with distinct areas of cell aggregation, which was not the case for the unstimulated (US) control adMSC being evenly spread and showing a spindle-like shape typical for cells of the fibroblastoid lineage (Figure 2). On the hydrogels without chondrogenic stimulation, adMSC tended to aggregate, but still showed a spindle-like phenotype, while chondrogenic stimulation of adMSC on the hydrogels resulted in strong aggregation into spheroid-like structures of approximately 70 μm diameter.
The cellular condensation step observed on the hydrogels after chondrogenic stimulation resembles the process of mesenchymal condensation, i.e., the aggregation of cells into three-dimensional structures that subsequently build up the distinct layers of cell-extracellular matrix composites found in cartilaginous tissue [39,40]. Mesenchymal condensation is a prerequisite for chondrogenic differentiation to occur [41–43] and depends on matrix elasticity. Human bmMSC on flexible silk fibroin matrices with a lateral elasticity modulus of 4.8 GPa were found to maintain a rounded cell phenotype, to promote cell aggregation in chondrogenic medium and to significantly increase cartilaginous type II collagen and glycosaminoglycan accumulation as well as SOX9 expression, while stiffer matrices of 7.8 GPa composed of identical material supported a flattened cell phenotype without cellular aggregation and increased cartilage marker expression [44]. Thus, the gelatin-based hydrogels used here promote adMSC condensation, but require additional chondrogenic stimulation to fully induce this differentiation pathway.
3.2.2. Staining of Cartilaginous Matrix Proteins
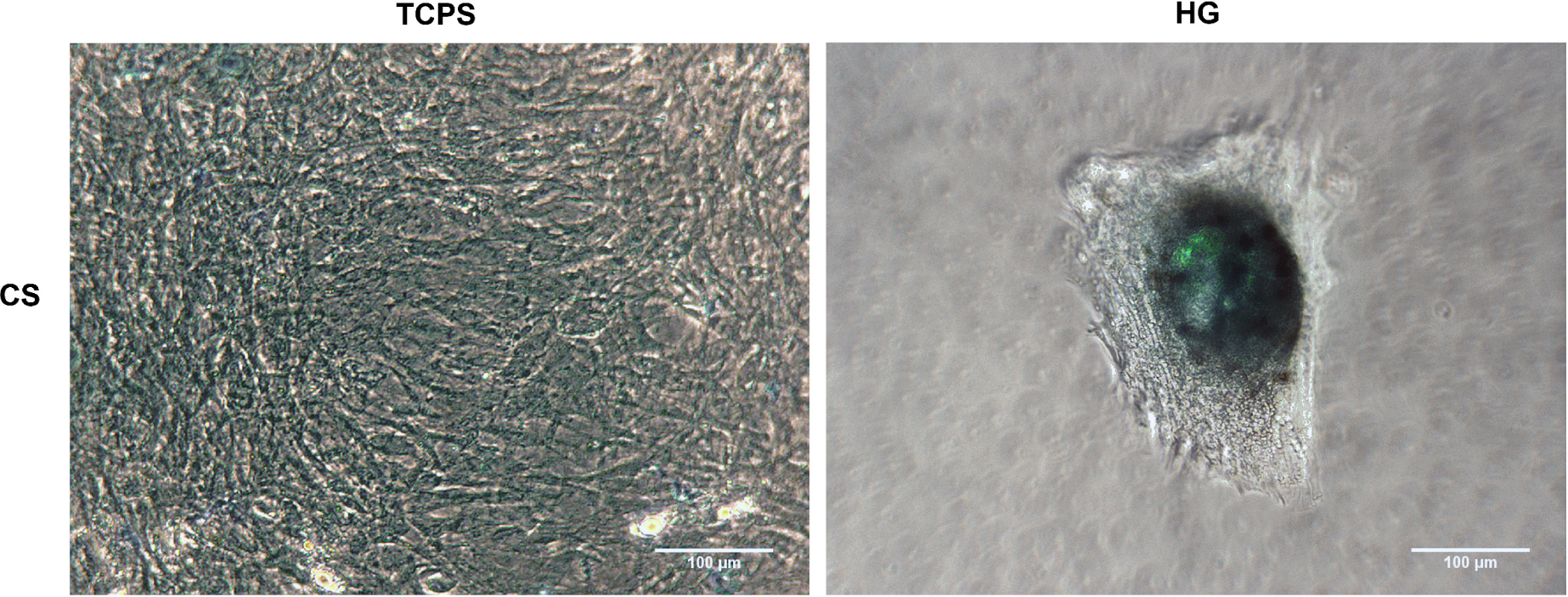
Having noticed that chondrogenically stimulated adMSC aggregated into spheroid-like structures on the hydrogels, we looked for the presence of acidic polysaccharides characteristic for cartilaginous tissue, since cartilage extracellular matrix characteristically consists of proteins glycosylated by glycosaminoglycans, so-called proteoglycans as e.g., aggrecan (ACAN) [2,42]. Staining of acidic glycosaminoglycans on the surface and within the hydrogels was done using Alcian Blue [45,46]. Following Alcian Blue staining, we found a strong accumulation of glycosaminoglycans inside the spheroids formed by chondrogenically stimulated adMSC on the hydrogels, while the even cell layer formed by chondrogenically stimulated adMSC on TCPS showed negligible staining (Figure 3).
Thus, hydrogels, in combination with chondrogenic stimulants, have a strong inductive effect on the deposition of cartilaginous matrix components by adMSC.
3.2.3. Expression of Chondrogenic Marker Genes
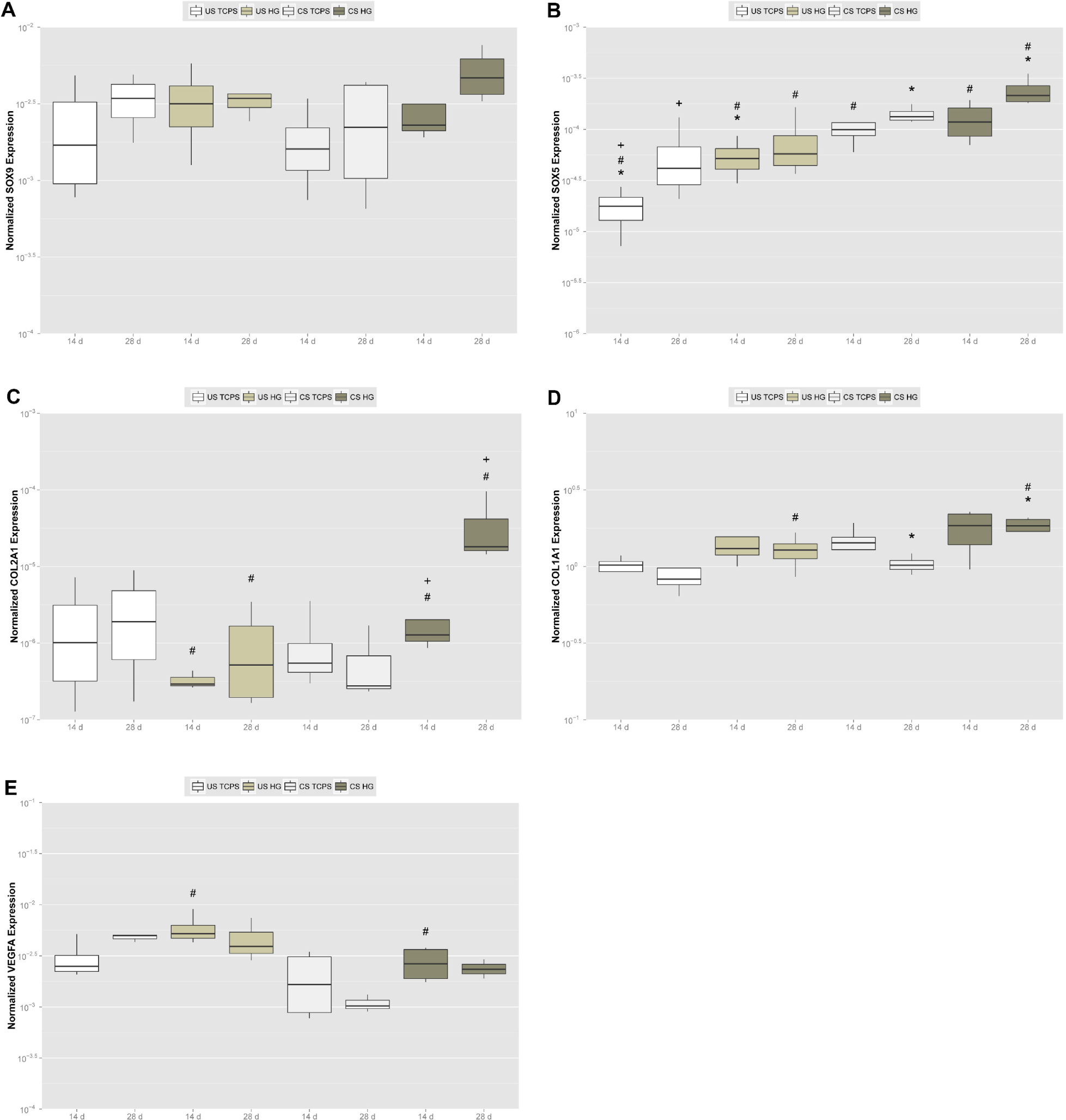
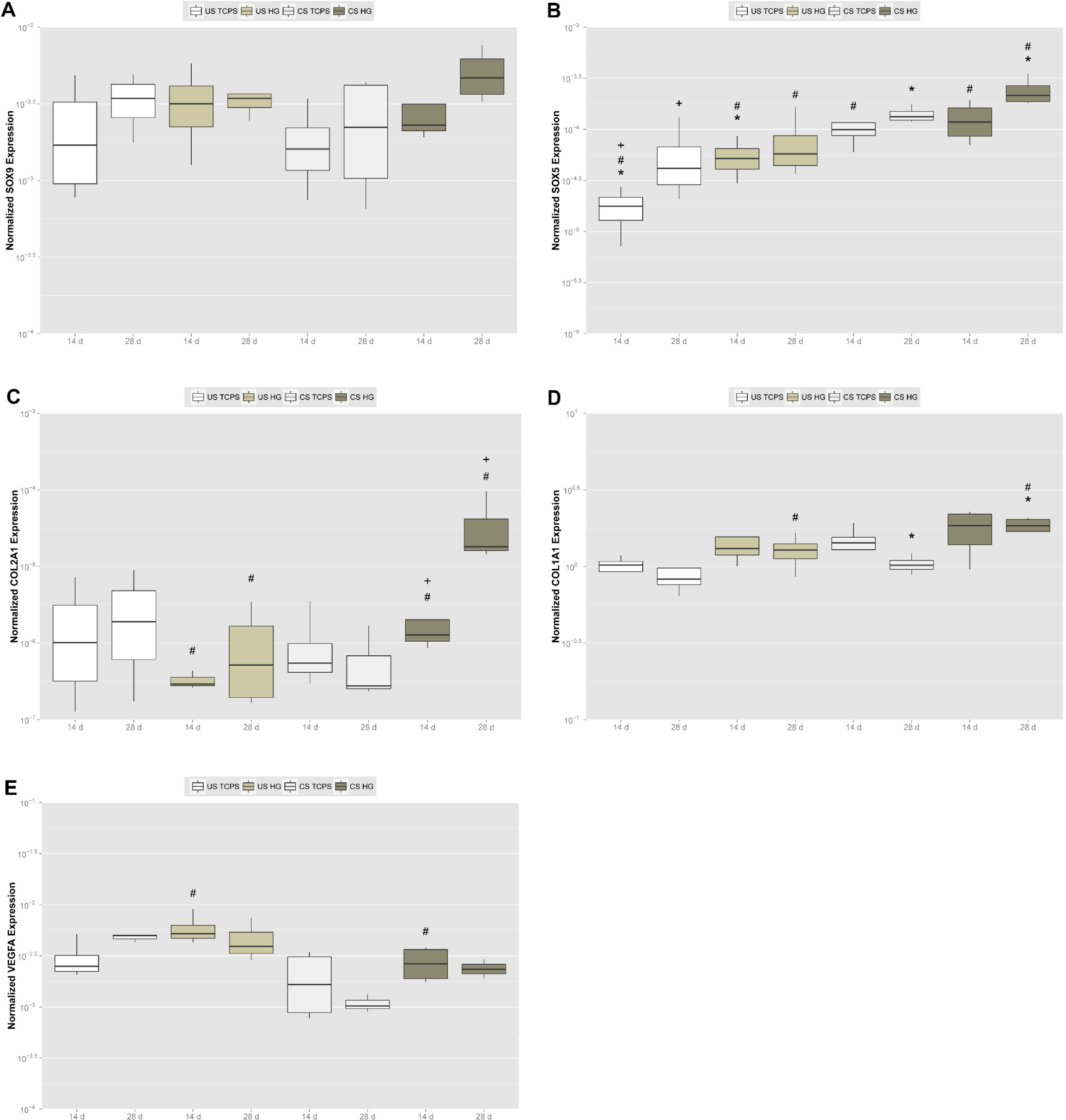
To more closely and quantitatively investigate onset and progression of chondrogenic differentiation of adMSC on the hydrogels, we analyzed expression of genes involved in that process using real-time RT PCR. Expression of SOX9, a gene encoding a member of the SOX (SRY-related HMG-box) family, was upregulated on hydrogels and following chondrogenic stimulation (Figure 4A). The strongest SOX9 expression was found for chondrogenically stimulated adMSC on the hydrogels after 28 days. Expression of the transcription factor SOX5 gene, another member of the HMG-box class DNA-binding proteins, was significantly and time-dependently upregulated on the hydrogels as compared to TCPS (Figure 4B). Upon chondrogenic stimulation of adMSC, expression of this gene significantly increased, an effect that was the strongest on the hydrogels.
The trio of SOX transcription factors 5, 6 and 9 plays a crucial role in chondrogenesis, SOX9 being the mayor player and the single factor containing a transactivation domain, while complexation with SOX5 and 6, which contain only DNA-binding domains, has a promotive effect on cooperative binding of the homodimerized SOX trio to enhancers of chondrocyte-specific genes [47]. Moreover, SOX9 upregulates expression of SOX5 and SOX6 [43] and is required for chondrogenic lineage commitment by MSC condensation, its expression shutting down at the prehypertrophic stage, while the auxiliary SOX5 is expressed in a constitutive manner, being activated in prechondrocytes and highly expressed until the chondroblast stage [41–43]. Chondrogenic stimulation using transforming growth factors beta 1 (TGFβ1) or 3 (TGFβ3) was shown to upregulate SOX9, while osteogenic stimulation via the Wingless Int-1 pathway (WNT) downregulates this factor [43]. Simultaneously, SOX9 was reported to inhibit the osteoblastogenesis key transcription factor runt-related transcription factor 2 (RUNX2) [40]. Therefore, our chondrogenic transcription factor gene expression data underline a promotive impact of the hydrogels on chondrogenic differentiation of adMSC in vitro.
This supportive impact of the hydrogels is sustained by the data obtained for collagen gene expression. Cartilage consists primarily of type II collagen that entraps hydrated proteoglycans and glycoproteins to build a gel-like extracellular matrix, and the SOX trio of transcription factors binds to a type II collagen enhancer to directly drive expression of COL2A1, the gene encoding the alpha-1 chain of type II collagen [41–43]. In our experiments, COL2A1 expression was significantly and time-dependently upregulated for chondrogenically stimulated adMSC on the hydrogels, but downregulated on TCPS (Figure 4C). Expression of COL1A1, the gene encoding the pro-alpha-1 chains of the fibril-forming collagen type I found most connective tissues, was significantly upregulated upon chondrogenic stimulation, an effect that was significantly stronger on the hydrogels (Figure 4D). Since most cartilage in vivo is articular cartilage located in between joint space and bone and type I collagen is the mayor collagen type found in bone [39,48], our gene expression data for type I and II collagen clearly point out a promotive effect of the hydrogels on chondrogenic differentiation of adMSC in vitro.
Finally, we found expression of the VEGFA gene to be strongly upregulated for adMSC on the hydrogels, but to be significantly downregulated when cells were stimulated chondrogenically (see Figure 4E). Vascular endothelial growth factor A (VEGFA) is a prominent player in vasculogenesis whose expression is induced by SOX9 [49] and COL2A1 [50], but most interestingly by the hypoxia-induced transcription factor hypoxia inducible factor 1 alpha (HIF1A), as shown in rat PC12 cells [51], in human pulmonary artery endothelial cells [52] and in human Hep3B cells [53]. Within the avascular and alymphatic cartilaginous tissue [1], HIF1A was shown to be required for endochondral ossification to proceed [39], being a critical survival factor for hypoxic chondrocytes and increasing cartilaginous extracellular matrix accumulation and early chondrogenesis [50]. Thus, the strong upregulation of VEGFA expression for unstimulated adMSC on the hydrogels may reflect the onset of gene expression to counteract hypoxic conditions occurring in the center of the spheroids formed, while additionally stimulating chondrogenically abolishes pro-angiogenic gene expression despite central hypoxia and therefore resembles chondrogenesis in vivo.
4. Conclusions
Hence, our gene and protein expression data argue in favor of a pro-chondrogenic impact of gelatin-based hydrogels on adMSC in vitro. Data on chondrogenic differentiation of MSC on gelatin-based hydrogels are rare in the literature [8]. Commonly used naturally-derived hydrogels in cartilage studies are based on collagen and alginate [48] or on hydroxyapatite [1]. In these studies, stronger cartilaginous extracellular matrix synthesis and a more physiologic compressibility were observed. However, proteoglycan content was only 10% of that present in the nucleus pulposus of intervertebral discs in vivo, which leaves a great need for improvement. Initial studies employing rabbit bmMSC on an osteochondral composite material consisting of a polyethylen glycol and a gelatin microparticles layer promisingly were found to differentiate into osteoblasts or chondrocytes in the respective layer, TGFβ1 supplementation of the gelatin layer and presence of osteoblastic cells in the polyethylen glycol layer both having a supportive effect on chondrogenesis [54]. Moreover, the inductive effect our hydrogels have on spontaneous adMSC spheroid formation are likely to promote chondrogenic differentiation, since rat bmMSC embedded in gelatin hydrogel microspheres releasing TGFβ1 were found to more strongly produce glycosaminoglycans than bmMSC on gelatin sheets [55]. Therefore, the gelatin-based hydrogels we produced are promising candidates for subsequent in vivo studies aiming at disease-state cartilage regeneration.
Acknowledgments
The authors kindly acknowledge the financial support of this work by the European Union and the Federal State of Mecklenburg-Vorpommern (ESF/IV-WM-B34-0006/08). The UGent authors would like to acknowledge Gent University for financial support in the frameworks of the Ugent-GOA project 2010–2015 (BOF10/GOA/005, Biomedical Engineering for Improved Diagnosis and Patient-Tailored Treatment of Aortic Aneurysms and Dissection) and the Ugent Multidisciplinary Research Partnership Nano- and biophotonics (2010–2014). Peter Dubruel would like to thank the Alexander von Humboldt Foundation for the financial support under the form of a granted Research Fellowship. Sandra Van Vlierberghe would like to acknowledge the Research Foundation Flanders for the financial support under the form of a post-doctoral fellowship and a Research Grant (“Development of the ideal tissue engineering scaffold by erging state-of-the-art processing techniques”, FWO Krediet aan Navorsers). Krisztina Fehér and Tim Courtin are acknowledged for their assistance in the recording of the various HR-MAS NMR spectra. The NMR equipment involved in this research is part of the Interuniversity NMR Facility at Gent University and funded by the FFEU-ZWAP initiative of the Flemish Government and the Hercules Foundation.
Author Contributions
Achim Salamon, Sandra van Vlierberghe, Jose C. Martins, Peter Dubruel and Kirsten Peters made substantial contributions to conception and design. Ine van Nieuwenhove, Frank Baudisch, Geert-Jan Graulus, Verena Benecke and Kristin Alberti acquired the data. Achim Salamon, Sandra van Vlierberghe, Ine van Nieuwenhove, Geert-Jan Graulus, Jose C. Martins, Peter Dubruel and Kirsten Peters analyzed and interpreted the data. Achim Salamon, Sandra van Vlierberghe and Ine van Nieuwenhove drafted the article. Achim Salamon, Sandra van Vlierberghe, Ine van Nieuwenhove, Geert-Jan Graulus, Jose C. Martins, Peter Dubruel and Kirsten Peters revised it critically for important intellectual content. All authors gave final approval of the version to be submitted and any revised version. Achim Salamon is corresponding author for the stem cell part of the paper, and Sandra van Vlierberghe is corresponding author for the hydrogel part.
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Kim, I.L.; Mauck, R.L.; Burdick, J.A. Hydrogel design for cartilage tissue engineering: A case study with hyaluronic acid. Biomaterials 2011, 32, 8771–8782. [Google Scholar]
- Tew, S.R.; Murdoch, A.D.; Rauchenberg, R.P.; Hardingham, T.E. Cellular methods in cartilage research: Primary human chondrocytes in culture and chondrogenesis in human bone marrow stem cells. Methods 2008, 45, 2–9. [Google Scholar]
- Noeth, U.; Steinert, A.F.; Tuan, R.S. Technology insight: Adult mesenchymal stem cells for osteoarthritis therapy. Nat. Clin. Pract. Rheumatol 2008, 4, 371–380. [Google Scholar]
- Ratner, B.D.; Hoffman, A.S.; Schoen, F.J.; Lemons, J.E. Biomaterials Science: An Introduction to Materials in Medicine, 3rd ed; Academic Press: San Diego, CA, USA, 2004; pp. 162–164. [Google Scholar]
- Lin, L.H.; Chen, K.M. Preparation and surface activity of gelatin derivative surfactants. Colloids Surf. A Physicochem. Eng. Asp 2006, 272, 8–14. [Google Scholar]
- Johns, P.; Courts, A. Relationship between Collagen and Gelatin. In The Science and Technology of Gelatin; Ward, A.G., Courts, A., Eds.; Academic Press: London, UK, 1977; pp. 137–177. [Google Scholar]
- Sirova, M.; Vlierberghe, S.V.; Matyasova, V.; Rossmann, P.; Schacht, E.; Dubruel, P.; Rihova, B. Immunocompatibility evaluation of hydrogel-coated polyimide implants for applications in regenerative medicine. J. Biomed. Mater. Res. A 2013, 2013. [Google Scholar] [CrossRef]
- Van Vlierberghe, S.; Dubruel, P.; Schacht, E. Biopolymer-based hydrogels as scaffolds for tissue engineering applications: A review. Biomacromolecules 2011, 12, 1387–1408. [Google Scholar]
- Gorgieva, S.; Kokol, V. Preparation, characterization, and in vitro enzymatic degradation of chitosan-gelatine hydrogel scaffolds as potential biomaterials. J. Biomed. Mater. Res. A 2012, 100, 1655–1667. [Google Scholar]
- Lai, J.Y. Influence of solvent composition on the performance of carbodiimide cross-linked gelatin carriers for retinal sheet delivery. J. Mater. Sci. Mater. Med 2013, 24, 2201–2210. [Google Scholar]
- Vorrapakdee, R.; Kanokpanont, S.; Ratanavaraporn, J.; Waikakul, S.; Charoenlap, C.; Damrongsakkul, S. Modification of human cancellous bone using Thai silk fibroin and gelatin for enhanced osteoconductive potential. J. Mater. Sci. Mater. Med 2013, 24, 735–744. [Google Scholar]
- Elzoghby, A.O. Gelatin-based nanoparticles as drug and gene delivery systems: Reviewing three decades of research. J. Control. Release 2013, 172, 1075–1091. [Google Scholar]
- Sacchetti, B.; Funari, A.; Michienzi, S.; di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 2007, 131, 324–336. [Google Scholar]
- Mendez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar]
- Friedenstein, A.J.; Piatetzky-Shapiro, I.I.; Petrakova, K.V. Osteogenesis in transplants of bone marrow cells. J. Embryol. Exp. Morphol 1966, 16, 381–390. [Google Scholar]
- Bianco, P.; Cao, X.; Frenette, P.S.; Mao, J.J.; Robey, P.G.; Simmons, P.J.; Wang, C.Y. The meaning, the sense and the significance: Translating the science of mesenchymal stem cells into medicine. Nat. Med 2013, 19, 35–42. [Google Scholar]
- Nombela-Arrieta, C.; Ritz, J.; Silberstein, L.E. The elusive nature and function of mesenchymal stem cells. Nat. Rev. Mol. Cell Biol 2011, 12, 126–131. [Google Scholar]
- Shi, Y.; Niedzinski, J.R.; Samaniego, A.; Bogdansky, S.; Atkinson, B.L. Adipose-derived stem cells combined with a demineralized cancellous bone substrate for bone regeneration. Tissue Eng. Part A 2012, 18, 1313–1321. [Google Scholar]
- Levi, B.; Longaker, M.T. Concise review: Adipose-derived stromal cells for skeletal regenerative medicine. Stem Cells 2011, 29, 576–582. [Google Scholar]
- James, A.W.; Zara, J.N.; Zhang, X.; Askarinam, A.; Goyal, R.; Chiang, M.; Yuan, W.; Chang, L.; Corselli, M.; Shen, J.; et al. Perivascular stem cells: A prospectively purified mesenchymal stem cell population for bone tissue engineering. Stem Cells Transl. Med 2012, 1, 510–519. [Google Scholar]
- Shoji, T.; Ii, M.; Mifune, Y.; Matsumoto, T.; Kawamoto, A.; Kwon, S.M.; Kuroda, T.; Kuroda, R.; Kurosaka, M.; Asahara, T. Local transplantation of human multipotent adipose-derived stem cells accelerates fracture healing via enhanced osteogenesis and angiogenesis. Lab. Investig 2010, 90, 637–649. [Google Scholar]
- Choi, J.W.; Park, E.J.; Shin, H.S.; Shin, I.S.; Ra, J.C.; Koh, K.S. In vivo differentiation of undifferentiated human adipose tissue-derived mesenchymal stem cells in critical-sized calvarial bone defects. Ann. Plast. Surg 2014, 72, 225–233. [Google Scholar]
- Schubert, T.; Lafont, S.; Beaurin, G.; Grisay, G.; Behets, C.; Gianello, P.; Dufrane, D. Critical size bone defect reconstruction by an autologous 3D osteogenic-like tissue derived from differentiated adipose MSCs. Biomaterials 2013, 34, 4428–4438. [Google Scholar]
- Pourebrahim, N.; Hashemibeni, B.; Shahnaseri, S.; Torabinia, N.; Mousavi, B.; Adibi, S.; Heidari, F.; Alavi, M.J. A comparison of tissue-engineered bone from adipose-derived stem cell with autogenous bone repair in maxillary alveolar cleft model in dogs. Int. J. Oral Maxillofac. Surg 2013, 42, 562–568. [Google Scholar]
- Lendeckel, S.; Joedicke, A.; Christophis, P.; Heidinger, K.; Wolff, J.; Fraser, J.K.; Hedrick, M.H.; Berthold, L.; Howaldt, H.P. Autologous stem cells (adipose) and fibrin glue used to treat widespread traumatic calvarial defects: Case report. J. Craniomaxillofac. Surg 2004, 32, 370–373. [Google Scholar]
- Mesimaki, K.; Lindroos, B.; Tornwall, J.; Mauno, J.; Lindqvist, C.; Kontio, R.; Miettinen, S.; Suuronen, R. Novel maxillary reconstruction with ectopic bone formation by GMP adipose stem cells. Int. J. Oral Maxillofac. Surg 2009, 38, 201–209. [Google Scholar]
- Toupet, K.; Maumus, M.; Peyrafitte, J.A.; Bourin, P.; van Lent, P.L.; Ferreira, R.; Orsetti, B.; Pirot, N.; Casteilla, L.; Jorgensen, C.; et al. Long-term detection of human adipose-derived mesenchymal stem cells after intraarticular injection in SCID mice. Arthritis Rheum 2013, 65, 1786–1794. [Google Scholar]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar]
- Gimble, J.M.; Bunnell, B.A.; Frazier, T.; Rowan, B.; Shah, F.; Thomas-Porch, C.; Wu, X. Adipose-derived stromal/stem cells: A primer. Organogenesis 2013, 9, 3–10. [Google Scholar]
- Salamon, A.; Jonitz-Heincke, A.; Adam, S.; Rychly, J.; Mueller-Hilke, B.; Bader, R.; Lochner, K.; Peters, K. Articular cartilage-derived cells hold a strong osteogenic differentiation potential in comparison to mesenchymal stem cells in vitro. Exp. Cell Res 2013, 319, 2856–2865. [Google Scholar]
- Peters, K.; Salamon, A.; Vlierberghe, S.V.; Rychly, J.; Kreutzer, M.; Neumann, H.G.; Schacht, E.; Dubruel, P. A new approach for adipose tissue regeneration based on human mesenchymal stem cells in contact to hydrogels—An in vitro study. Adv. Eng. Mater 2009, 11, B155–B161. [Google Scholar]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol 2000, 132, 365–386. [Google Scholar]
- R Core Team, R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013.
- Shapiro, M.J.; Chin, J.; Marti, R.E.; Jarosinski, M.A. Enhanced resolution in MAS NMR for combinatorial chemistry. Tetrahedron Lett 1997, 38, 1333–1336. [Google Scholar]
- Ramadhar, T.R.; Amador, F.; Ditty, M.J.T.; Power, W.P. Inverse H-C ex situ HRMAS NMR experiments for solid-phase peptide synthesis. Magn. Reson. Chem 2008, 46, 30–35. [Google Scholar]
- Van Vlierberghe, S.; Fritzinger, B.; Martins, J.C.; Dubruel, P. Hydrogel network formation revised: High-resolution magic angle spinning nuclear magnetic resonance as a powerful tool for measuring absolute hydrogel cross-link efficiencies. Appl. Spectrosc 2010, 64, 1176–1180. [Google Scholar]
- Rueda, J.; Suica, R.; Komber, H.; Voit, B. Synthesis of new polymethyloxazoline hydrogels by the “macroinitiator” method. Macromol. Chem. Phys 2003, 204, 954–960. [Google Scholar]
- Schipani, E. Posttranslational modifications of collagens as targets of hypoxia and Hif-1alpha in endochondral bone development. Ann. N. Y. Acad. Sci 2010, 1192, 317–321. [Google Scholar]
- Studer, D.; Millan, C.; Ozturk, E.; Maniura-Weber, K.; Zenobi-Wong, M. Molecular and biophysical mechanisms regulating hypertrophic differentiation in chondrocytes and mesenchymal stem cells. Eur. Cells Mater 2012, 24, 118–35, discussion 135. [Google Scholar]
- Lefebvre, V.; Smits, P. Transcriptional control of chondrocyte fate and differentiation. Birth Defects Res. C Embryo Today 2005, 75, 200–212. [Google Scholar]
- Ikeda, T.; Kawaguchi, H.; Kamekura, S.; Ogata, N.; Mori, Y.; Nakamura, K.; Ikegawa, S.; Chung, U.I. Distinct roles of Sox5, Sox6, and Sox9 in different stages of chondrogenic differentiation. J. Bone Miner. Metab 2005, 23, 337–340. [Google Scholar]
- Quintana, L. zur Nieden, N.I.; Semino, C.E. Morphogenetic and regulatory mechanisms during developmental chondrogenesis: New paradigms for cartilage tissue engineering. Tissue Eng. Part B Rev 2009, 15, 29–41. [Google Scholar]
- Ghosh, S.; Laha, M.; Mondal, S.; Sengupta, S.; Kaplan, D.L. In vitro model of mesenchymal condensation during chondrogenic development. Biomaterials 2009, 30, 6530–6540. [Google Scholar]
- Scott, J.E.; Dorling, J. Differential staining of acid glycosaminoglycans (mucopolysaccharides) by alcian blue in salt solutions. Histochemie 1965, 5, 221–233. [Google Scholar]
- Goldstein, D.J.; Horobin, R.W. Surface staining of cartilage by Alcian blue, with reference to the role of microscopic dye aggregates in histological staining. Histochem. J 1974, 6, 175–184. [Google Scholar]
- Akiyama, H.; Lefebvre, V. Unraveling the transcriptional regulatory machinery in chondrogenesis. J. Bone Miner. Metab 2011, 29, 390–395. [Google Scholar]
- Tortelli, F.; Cancedda, R. Three-dimensional cultures of osteogenic and chondrogenic cells: A tissue engineering approach to mimic bone and cartilage in vitro. Eur. Cells Mater 2009, 17, 1–14. [Google Scholar]
- Robins, J.C.; Akeno, N.; Mukherjee, A.; Dalal, R.R.; Aronow, B.J.; Koopman, P.; Clemens, T.L. Hypoxia induces chondrocyte-specific gene expression in mesenchymal cells in association with transcriptional activation of Sox9. Bone 2005, 37, 313–322. [Google Scholar]
- Provot, S.; Schipani, E. Fetal growth plate: a developmental model of cellular adaptation to hypoxia. Ann. N. Y. Acad. Sci 2007, 1117, 26–39. [Google Scholar]
- Levy, A.P.; Levy, N.S.; Wegner, S.; Goldberg, M.A. Transcriptional regulation of the rat vascular endothelial growth factor gene by hypoxia. J. Biol. Chem 1995, 270, 13333–13340. [Google Scholar]
- Liu, Y.; Cox, S.R.; Morita, T.; Kourembanas, S. Hypoxia regulates vascular endothelial growth factor gene expression in endothelial cells. Identification of a 5’ enhancer. Circ. Res 1995, 77, 638–643. [Google Scholar]
- Forsythe, J.A.; Jiang, B.H.; Iyer, N.V.; Agani, F.; Leung, S.W.; Koos, R.D.; Semenza, G.L. Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1. Mol. Cell. Biol 1996, 16, 4604–4613. [Google Scholar]
- Guo, X.; Park, H.; Liu, G.; Liu, W.; Cao, Y.; Tabata, Y.; Kasper, F.K.; Mikos, A.G. In vitro generation of an osteochondral construct using injectable hydrogel composites encapsulating rabbit marrow mesenchymal stem cells. Biomaterials 2009, 30, 2741–2752. [Google Scholar]
- Ogawa, T.; Akazawa, T.; Tabata, Y. In vitro proliferation and chondrogenic differentiation of rat bone marrow stem cells cultured with gelatin hydrogel microspheres for TGF-beta1 release. J. Biomater. Sci. Polym. Ed 2010, 21, 609–621. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Forward Primer (5’-3’) | Reverse Primer (5’-3’) |
|---|---|---|---|
| ACTB | NM_001101.2 | cttcctgggcatggagtc | agcactgtgttggcgtacag |
| SOX5 | NM_178010.1 | aagcctttcctgacatgcac | tccagcgagatcccaatatc |
| SOX9 | NM_000346.2 | tgggcaagctctggagac | cgttcttcaccgacttcctc |
| COL1A1 | NM_000088.3 | acgaagacatcccaccaatc | agatcacgtcatcgcacaac |
| COL2A1 | NM_033150.2 | aatggtggcttccatttcag | gtgatgttctgggagccttc |
| VEGFA | NM_001025366.1 | ccttgctgctctacctccac | aatctgcatggtgatgttgg |
| Element | Gelatin (%) | Gel-MOD (%) |
|---|---|---|
| C | 69 | 72 |
| N | 13 | 10 |
| O | 18 | 18 |
| N/C | 0.19 | 0.12 |
| N/O | 0.72 | 0.56 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Salamon, A.; Van Vlierberghe, S.; Van Nieuwenhove, I.; Baudisch, F.; Graulus, G.-J.; Benecke, V.; Alberti, K.; Neumann, H.-G.; Rychly, J.; Martins, J.C.; et al. Gelatin-Based Hydrogels Promote Chondrogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stem Cells In Vitro. Materials 2014, 7, 1342-1359. https://doi.org/10.3390/ma7021342
Salamon A, Van Vlierberghe S, Van Nieuwenhove I, Baudisch F, Graulus G-J, Benecke V, Alberti K, Neumann H-G, Rychly J, Martins JC, et al. Gelatin-Based Hydrogels Promote Chondrogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stem Cells In Vitro. Materials. 2014; 7(2):1342-1359. https://doi.org/10.3390/ma7021342
Chicago/Turabian StyleSalamon, Achim, Sandra Van Vlierberghe, Ine Van Nieuwenhove, Frank Baudisch, Geert-Jan Graulus, Verena Benecke, Kristin Alberti, Hans-Georg Neumann, Joachim Rychly, José C. Martins, and et al. 2014. "Gelatin-Based Hydrogels Promote Chondrogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stem Cells In Vitro" Materials 7, no. 2: 1342-1359. https://doi.org/10.3390/ma7021342