Effects of Light and Temperature on Fatty Acid Production in Nannochloropsis Salina

Abstract
:1. Introduction
2. Results and Discussion
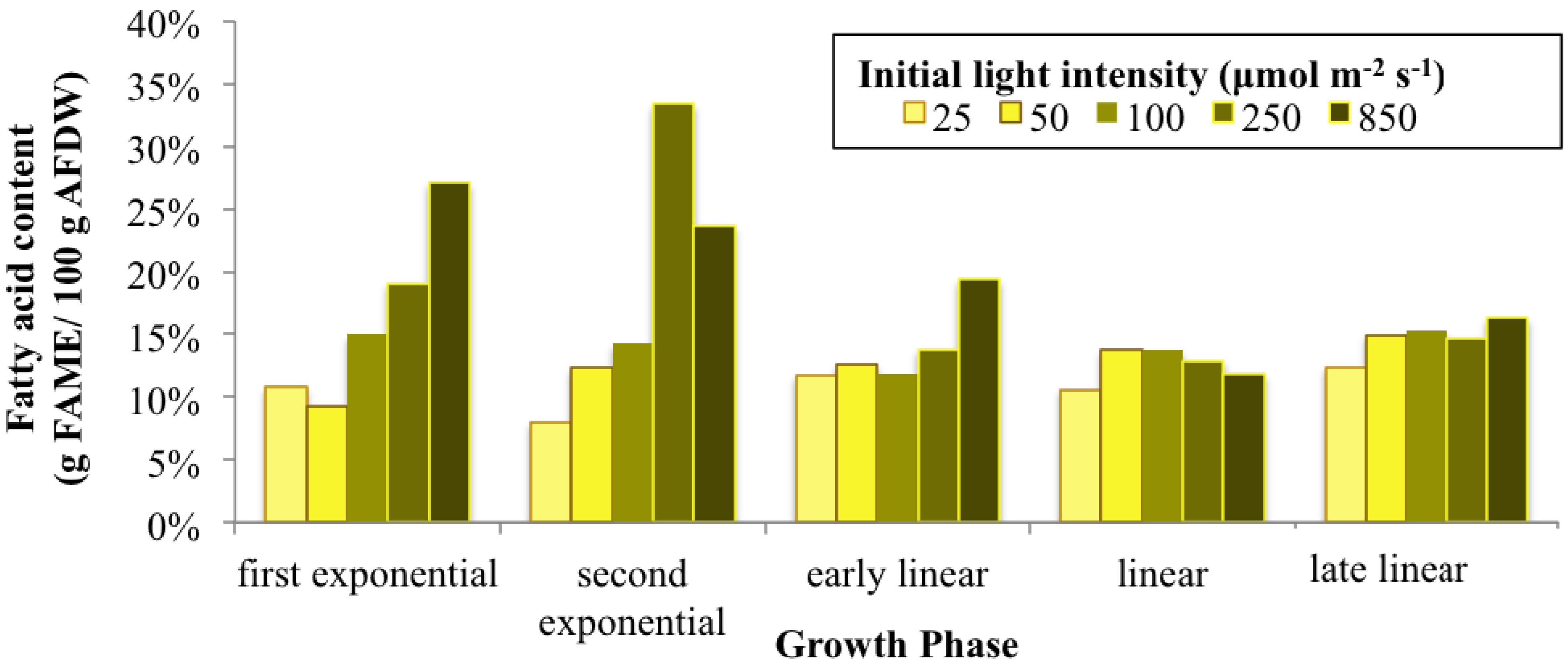
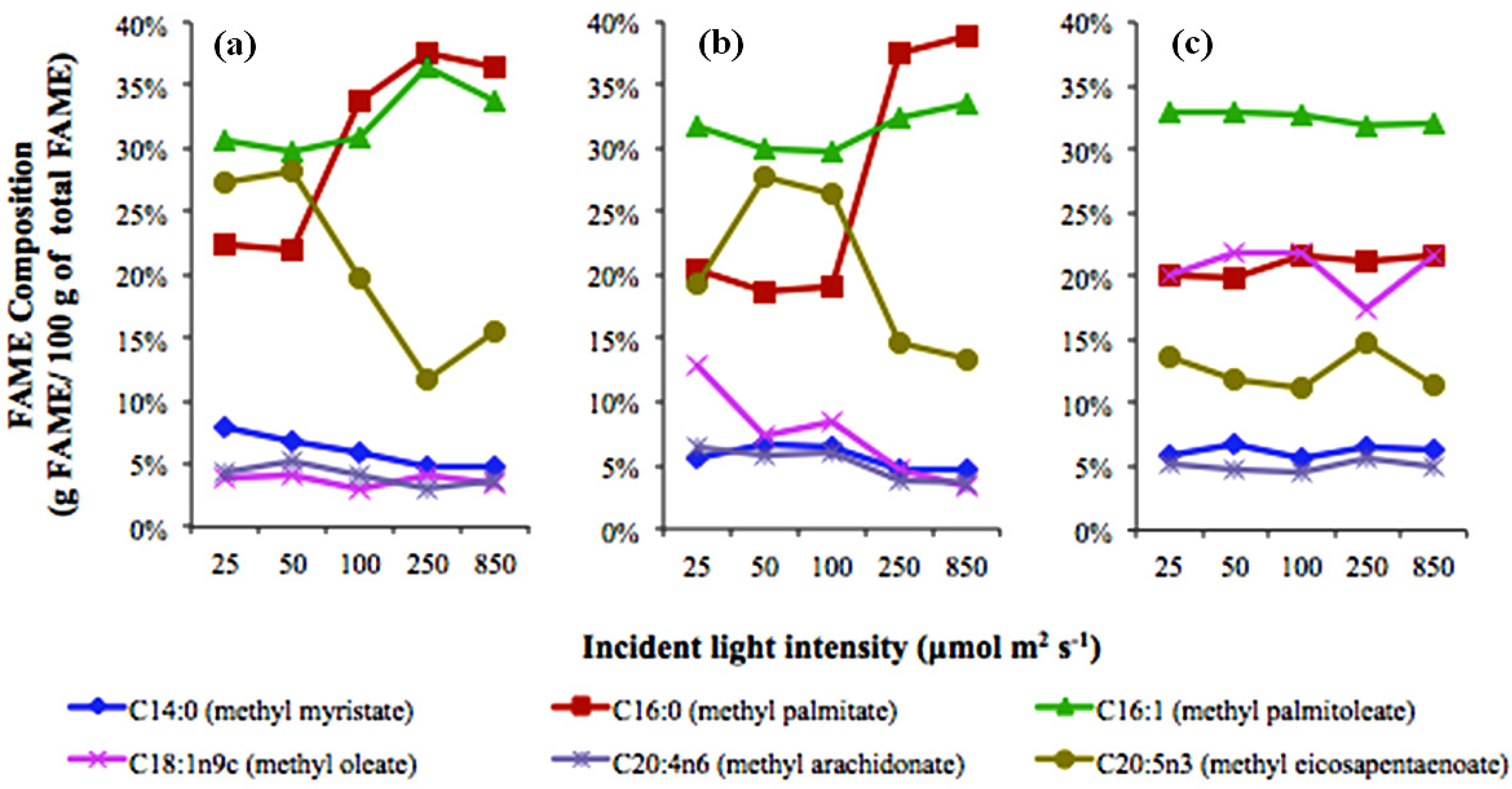
2.1. Light Effects


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PFD ± SD (μmol m−2 s−1) | Volumetric Productivity (g L−1 day−1) | Linear Growth Period a (days) | Maximum Concentration (g L−1) | Biomass Yield vs. Light (g mol−1 Photon Supplied) |
|---|---|---|---|---|
| 5 ± 0.5 | 0.0017 | >35 | 0.079 | 0.18 |
| 26 ± 0.7 | 0.0472 | >56 | 2.5 | 0.96 |
| 55 ± 1.6 | 0.1077 | >59 | 6.2 | 1.03 |
| 96 ± 2 | 0.137 | 48 | 7.0 | 0.75 |
| 234 ± 25 | 0.361 | 32 | 14.2 | 0.81 |
| 236 ± 23 | 0.319 | 22 | 10.4 | 0.71 |
| 833 ± 48 | 0.645 | 23 | 19.4 | 0.41 |
| 827 ± 58 | 0.575 | 28 | 17.2 | 0.37 |

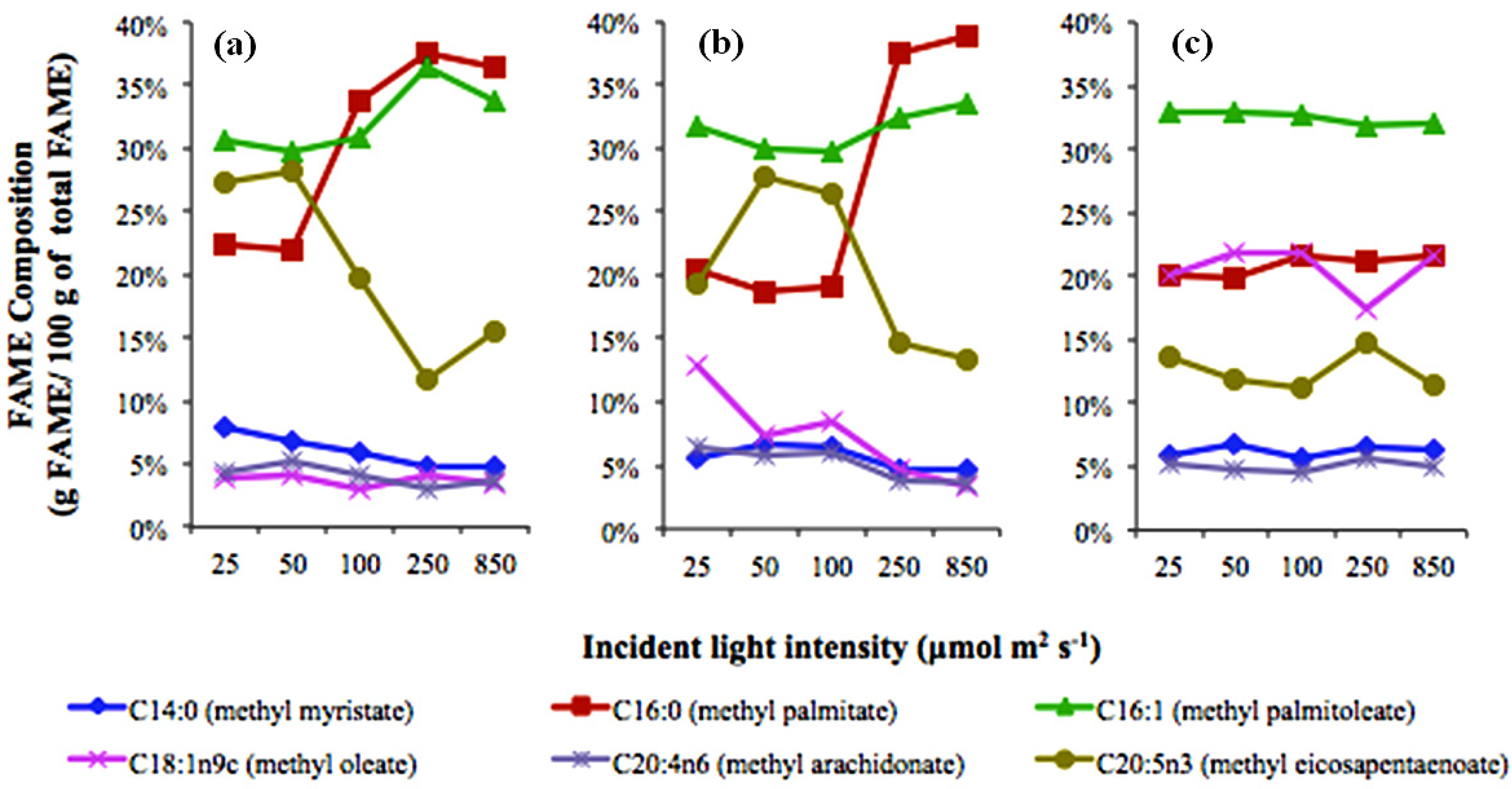
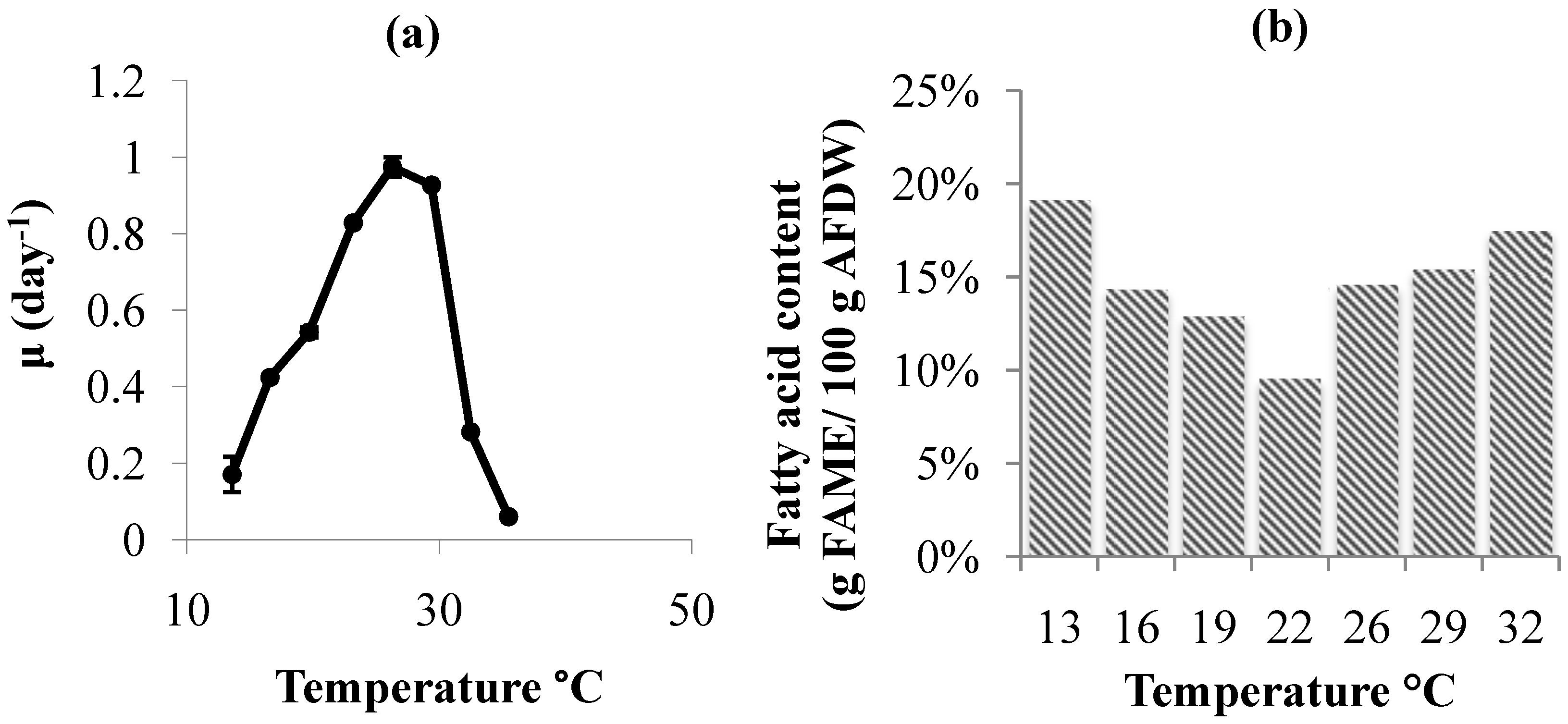
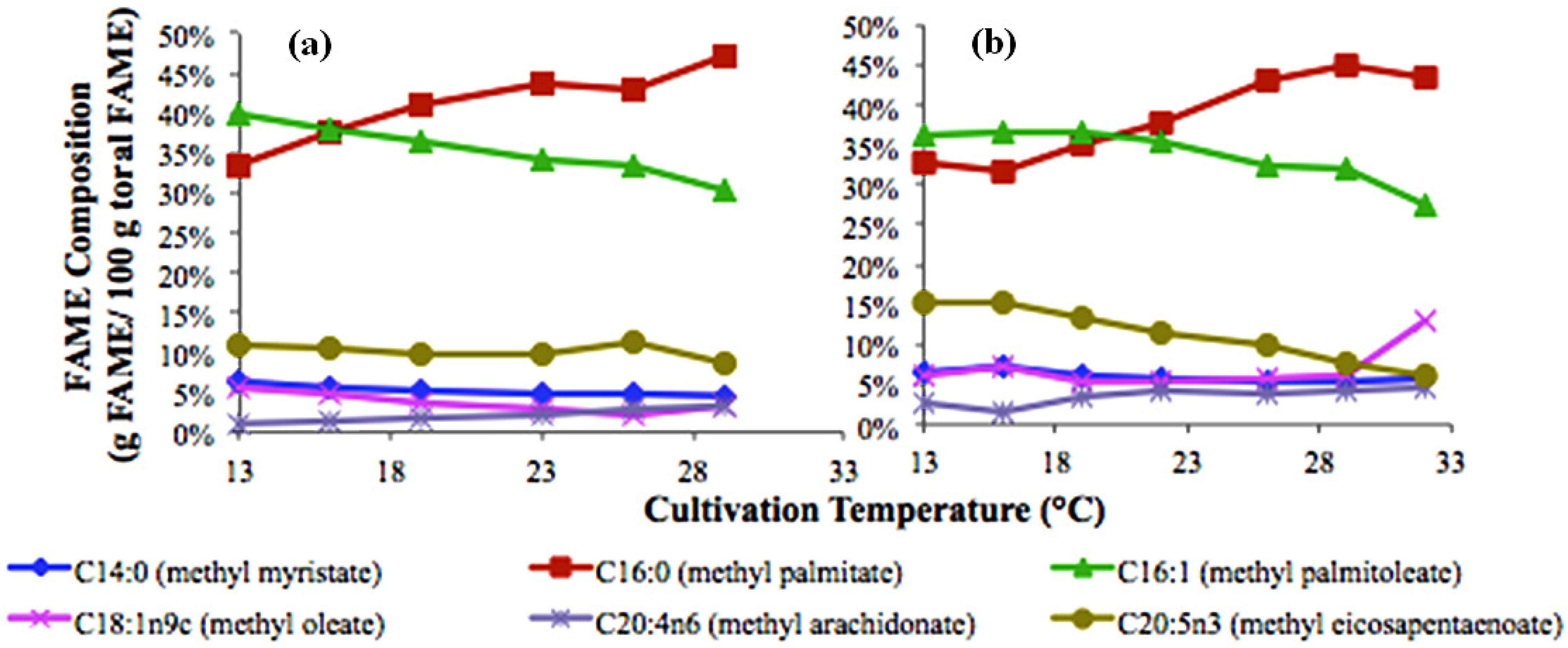
2.2. Temperature Effects
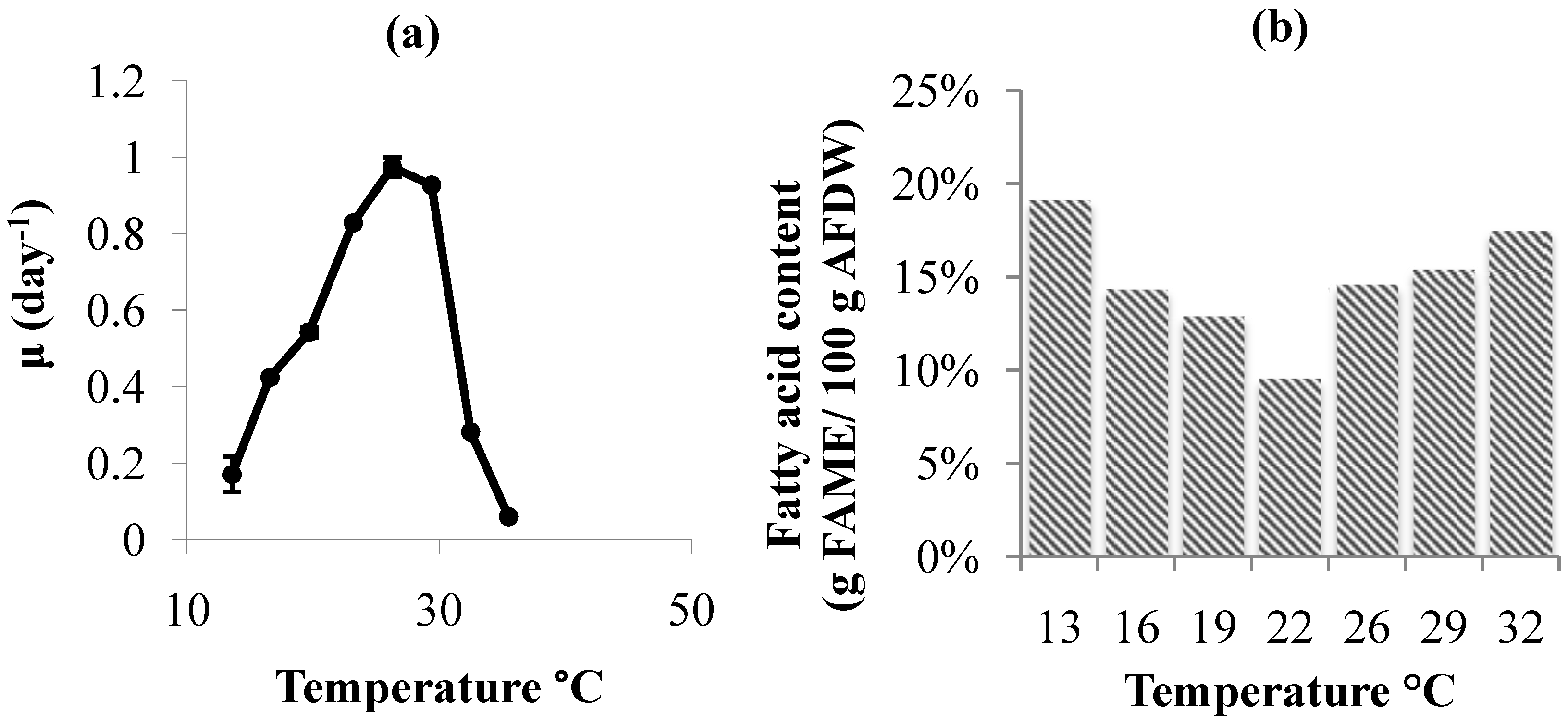
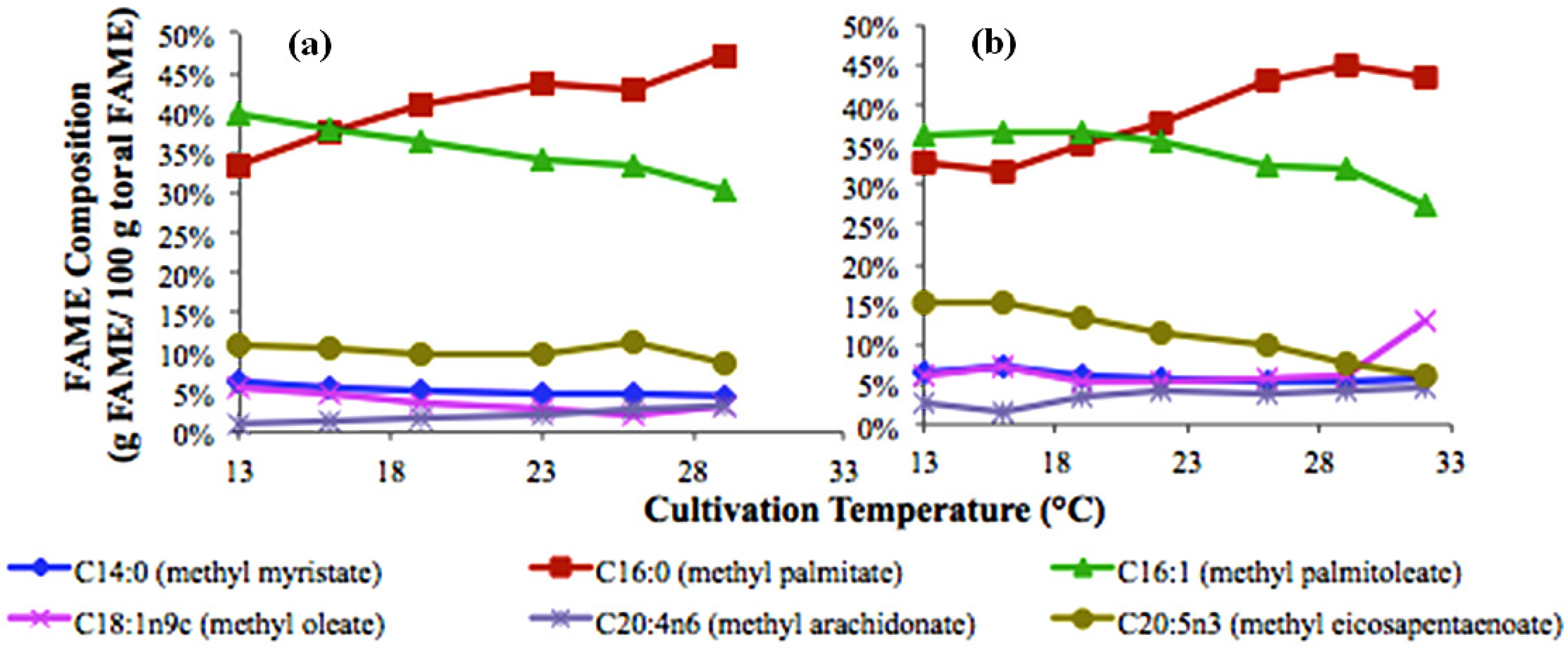
3. Experimental Section
3.1. Maintenance and Analytical Methods
3.2. Thermal Gradient Culture
3.3. Light Effect Determination
3.4. Growth Rate Calculation
3.5. Fatty Acid Quantification
4. Conclusions
Acknowledgments
References
- Sheehan, J. A Look Back at the U.S. Department of Energy’s Aquatic Species Program—Biodiesel from Algae Office of Fuels Development; National Renewable Energy Laboratory Denver: Denver, CO, USA, 1998; Volume 328. [Google Scholar]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Lundquist, T.; Woertz, I.; Quinn, N.; Benemann, J. A Realistic Technology and Engineering Assessment of Algae Biofuel Production. Energy Biosciences Institute: Berkeley, CA, USA, 2010. Available online: http://works.bepress.com/tlundqui/5 (accessed on 7 March 2012).
- Pienkos, P.T.; Darzins, A. The promise and challenges of microalgal-derived biofuels. Renew. Energy 2009, 3, 431–440. [Google Scholar]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. Plant J. Cell Mol. Biol. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Ryan, R.D.; Waller, P.M.; Kacira, M.; Li, P. Aquaculture Raceway Integrated Design. U.S. Patent. 3 February 2011. Available online: http://www.google.com/patents/US20110023360?dq=raceway+integrated+ design + waller (accessed on 9 January 2012).
- Wigmosta, M.S.; Coleman, A.M.; Skaggs, R.J.; Huesemann, M.H.; Lane, L.J. National microalgae biofuel production potential and resource demand. Water Resour. Res. 2011, 47, 1–13. [Google Scholar] [CrossRef]
- Griffiths, M.J.; Harrison, S.T.L. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J. Appl. Phycol. 2009, 21, 493–507. [Google Scholar] [CrossRef]
- Andersen, R. Algal Culturing Techniques; Elsevier/Academic Press: Burlington, MA, USA, 2005. [Google Scholar]
- Richmond, A. Handbook of Microalgal Culture; Blackwell Science: Oxford, UK, 2003. [Google Scholar]
- Cuaresma, M.; Janssen, M.; Jan, E.; End, V.D.; Vílchez, C.; Wijffels, R.H. Bioresource Technology Luminostat operation: A tool to maximize microalgae photosynthetic efficiency in photobioreactors during the daily light cycle? Bioresour. Technol. 2011, 102, 7871–7878. [Google Scholar] [CrossRef] [PubMed]
- Posten, C. Design principles of photo-bioreactors for cultivation of microalgae. Eng. Life Sci. 2009, 9, 165–177. [Google Scholar] [CrossRef]
- Sukenik, A.; Zmora, O.; Carmeli, Y. Biochemical quality of marine unicellular algae with special emphasis on lipid composition II. Nannochloropsis sp. Aquaculture 1993, 117, 313–326. [Google Scholar]
- Guschina, I.A.; Harwood, J.L. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res. 2006, 45, 160–186. [Google Scholar] [CrossRef] [PubMed]
- Sukenik, A.; Carmeli, Y.; Berner, T. Regulation of fatty acid composon by irradiance level in the eustigmatophyte Nan-nochloropsis sp. J. Appl. Phycol. 1989, 25, 686–692. [Google Scholar] [CrossRef]
- Dunstan, G.A.; Volkman, J.K.; Barrett, S.M.; Garland, C.D. Changes in the lipid composition and maximisation of the polyunsaturated fatty acid content of three microalgae grown in mass culture. Cultures 1993, 5, 71–83. [Google Scholar]
- Hodgson, P.A.; Henderson, R.J.; Sargent, J.R.; Leftley, J.W.; Down, C. Patterns of variation in the lipid class and fatty acid composition of Nannochloropsis oculata (Eustigmatophyceae) during batch culture I. The growth cycle. Culture 1991, 2, 169–181. [Google Scholar]
- Pal, D.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. The effect of light, salinity, and nitrogen availability on lipid production by Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2011, 90, 1429–1441. [Google Scholar] [CrossRef] [PubMed]
- Sandnes, J.M.; Källqvist, T.; Wenner, D.; Gislerød, H.R. Combined influence of light and temperature on growth rates of Nannochloropsis oceanica: linking cellular responses to large-scale biomass production. J. Appl. Phycol. 2005, 17, 515–525. [Google Scholar] [CrossRef]
- Brock, T. Brock’s Biology of Microorganisms, 12th ed.; Pearson Benjamin Cummings Press: San Francisco, CA, USA, 2009. [Google Scholar]
- Voet, D. Biochemistry, 3rd ed.; Wiley Press: New York, NY, USA and Chichester, UK, 2003. [Google Scholar]
- Williams, P.J.L.B.; Laurens, L.M.L. Microalgae as biodiesel & biomass feedstocks: Review & analysis of the biochemistry, energetics & economics. Energy Environ. Sci. 2010, 3, 554. [Google Scholar] [CrossRef]
- Gwo, J.-C.; Chiu, J.-Y.; Chou, C.-C.; Cheng, H.-Y. Cryopreservation of a marine microalga, Nannochloropsis oculata (Eustigmatophyceae). Cryobiology 2005, 50, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Huesemann, M.H.; Hausmann, T.S.; Bartha, R.; Aksoy, M.; Weissman, J.C.; Benemann, J.R. Biomass productivities in wild type and pigment mutant of Cyclotella sp. (Diatom). Appl. Biochem. Biotechnol. 2009, 157, 507–526. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.H.; Scotten, H.L.; Bradshaw, J.S. Thermal Gradient Incubators for Small Aquatic Organisms. Limnol. Oceanogr. 1963, 8, 357–360. [Google Scholar] [CrossRef]
- Christie, W. Lipid Analysis: Isolation, Separation, Identification and Lipidomic Analysis, 4th ed.; Oily Press: Bridgwater, UK, 2010. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Van Wagenen, J.; Miller, T.W.; Hobbs, S.; Hook, P.; Crowe, B.; Huesemann, M. Effects of Light and Temperature on Fatty Acid Production in Nannochloropsis Salina. Energies 2012, 5, 731-740. https://doi.org/10.3390/en5030731
Van Wagenen J, Miller TW, Hobbs S, Hook P, Crowe B, Huesemann M. Effects of Light and Temperature on Fatty Acid Production in Nannochloropsis Salina. Energies. 2012; 5(3):731-740. https://doi.org/10.3390/en5030731
Chicago/Turabian StyleVan Wagenen, Jon, Tyler W. Miller, Sam Hobbs, Paul Hook, Braden Crowe, and Michael Huesemann. 2012. "Effects of Light and Temperature on Fatty Acid Production in Nannochloropsis Salina" Energies 5, no. 3: 731-740. https://doi.org/10.3390/en5030731