Microbial Biodiesel Production by Direct Transesterification of Rhodotorula glutinis Biomass

Department of Bioengineering, Tatung University, Taipei 10452, Taiwan
*
Author to whom correspondence should be addressed.
Energies 2018, 11(5), 1036; https://doi.org/10.3390/en11051036
Submission received: 8 March 2018
/
Revised: 11 April 2018
/
Accepted: 19 April 2018
/
Published: 24 April 2018
(This article belongs to the Special Issue Biofuel and Bioenergy Technology)
Abstract
:(1) Background: Lipids derived from oleaginous microbes have become promising alternative feedstocks for biodiesel. This is mainly because the lipid production rate from microbes is one to two orders of magnitude higher than those of energy crops. However, the conventional process for converting these lipids to biodiesel still requires a large amount of energy and organic solvents; (2) Methods: In this study, an oleaginous yeast, Rhodotorula glutinis, was used for direct transesterification without lipid pre-extraction to produce biodiesel, using sulfuric acid or sodium hydroxide as a catalyst. Such processes decreased the amount of energy and organic solvents required simultaneously; (3) Results: When 1 g of dry R. glutinis biomass was subject to direct transesterification in 20 mL of methanol catalyzed by 0.6 M H2SO4 at 70 °C for 20 h, the fatty acid methyl ester (FAME) yield reached 111%. Using the same amount of biomass and methanol loading but catalyzed by 1 g/L NaOH at 70 °C for 10 h, the FAME yield reached 102%. The acid-catalyzed process showed a superior moisture tolerance; when the biomass contained 70% moisture, the FAME yield was 43% as opposed to 34% of the base-catalyzed counterpart; (4) Conclusions: Compared to conventional transesterification, which requires lipid pre-extraction, direct transesterification not only simplifies the process and shortens the reaction time, but also improves the FAME yield.
1. Introduction
Biodiesel is one of the most promising renewable fuels in transportation. It can be used as a drop-in replacement fuel for existing diesel vehicles and boiler engines without major modifications. It is also compatible with current fuel infrastructure [1]. Biodiesel is defined as the fatty acid, alkyl monoesters, derived from renewable feedstocks such as vegetable oils, animal fats, and waste cooking oil. Compared to conventional petrodiesel, biodiesel is highly degradable, non-toxic, and cleaner in exhaust emissions, with the exception of NOx [2]. The combustion properties of biodiesel are similar to those of petroleum diesel.
Currently, commercial biodiesel is produced by the transesterification of plant oils with short-chain alcohols, using alkaline catalysts such as NaOH and KOH; most of these plant oils are edible, including rapeseed, sunflower, palm, and soybean oil [3]. The biodiesel derived from these oils (first generation biodiesel) has two major drawbacks: (1) the high cost of feedstocks renders biodiesel unable to compete commercially with petrodiesel [4]; (2) the utilization of these edible plants for fuel production may endanger the world’s food supply [5]. These problems led to the development of the second generation which mainly utilizes non-edible plant oils such as jatropha, jojoba, and waste cooking oil [6]. However, the supply of these non-edible oils is not likely to meet the global demand for biodiesel. Recently, the use of oil-accumulating microbes as feedstocks for biodiesel production has drawn a lot of attention [7]; the biodiesel derived from these oleaginous microalgae, bacteria, yeasts, and fungi is commonly referred to as the third-generation.
These microbial lipids, also known as single cell oils (SCO), have certain advantages over oils derived from plants. Firstly, these oleaginous microbes can be cultivated all year round with suitable substrates on non-arable lands; secondly, the oil production rate is one to two orders of magnitude higher than those of conventional energy crops [8]. In addition, the lipid compositions of many oleaginous microbes are not too different from those of vegetable oils [9]. This suggests that the fuel properties of biodiesel derived from microbial origins should be similar to those derived from vegetables oils. Among the oleaginous microbes, microalgae have been extensively studied for their potential as alternative feedstocks for biodiesel. Microalgae are mostly phototrophic and capable of accumulating a large amount of biomass and lipids rapidly [10]. Although microalgae are very effective in producing SCO, if grown photo synthetically, they require a large area of land to be cultivated, and are subject to daily and seasonal variations [9,10]. Compared to microalgae, oleaginous yeasts could be a better alternative for the production of SCO. The growth rates of these yeasts are faster than those of microalgae; the duplication time could be less than 1 h in certain cases [11]. Unlike phototrophic microalgae, the growth of oleaginous yeasts is not affected by the variation in weather and sunlight and is less prone to contamination by other microorganisms. Several yeasts are known for their ability to accumulate SCO, such as Cryptococcus curvatus, Lipomyces starkeyi, Rhodosporidium toruloides, and Rhodotorula glutinis [12]. R. glutinis caught our attention as an ideal SCO-producing strain because it could be cultivated with a variety of low cost carbon sources such as crude glycerol [13] and lignocellulosic biomass hydrolysate [14]. R. glutinis is also very effective in accumulating SCO with oil content of up to 72% [9]. R. glutinis is also capable of synthesizing carotenoids including β-carotene, torulene, and torularhodin [15]; the composition of these carotenoids depends on the cultivation conditions.
The conventional process for converting yeast SCO to biodiesel includes the following steps in this order: cell disruption, oil extraction, separation and transesterification. In order to decrease the energy expenditure and the amount of solvents used in these steps, many researchers combine the aforementioned steps into one, which is usually described as direct transesterification [16]. The simplified process reduces the overall high cost of biodiesel derived from microbial origins. A fatty acid methyl esters (FAME) yield of up to 98% was reached by direct transesterification of dry R. toruloides biomass catalyzed by either H2SO4 or HCl [17]. Using the same dry biomass, a similar yield was also obtained by direct transesterification using NaOH [18].
In this work, dry R. glutinis biomass was converted to FAME by direct transesterification with either acidic or basic catalysts. The effects of catalyst amount, reaction temperature, incubation time, and methanol loading on FAME yield were studied. In order to evaluate the feasibility of using wet biomass directly, the influences of moisture content on the FAME yield were also examined. Finally, under optimized reaction conditions, the yields and FAME compositions from direct transesterification were compared with those from the conventional processes.
2. Results and Discussion
2.1. Effect of Catalyst Concentration on Transesterification
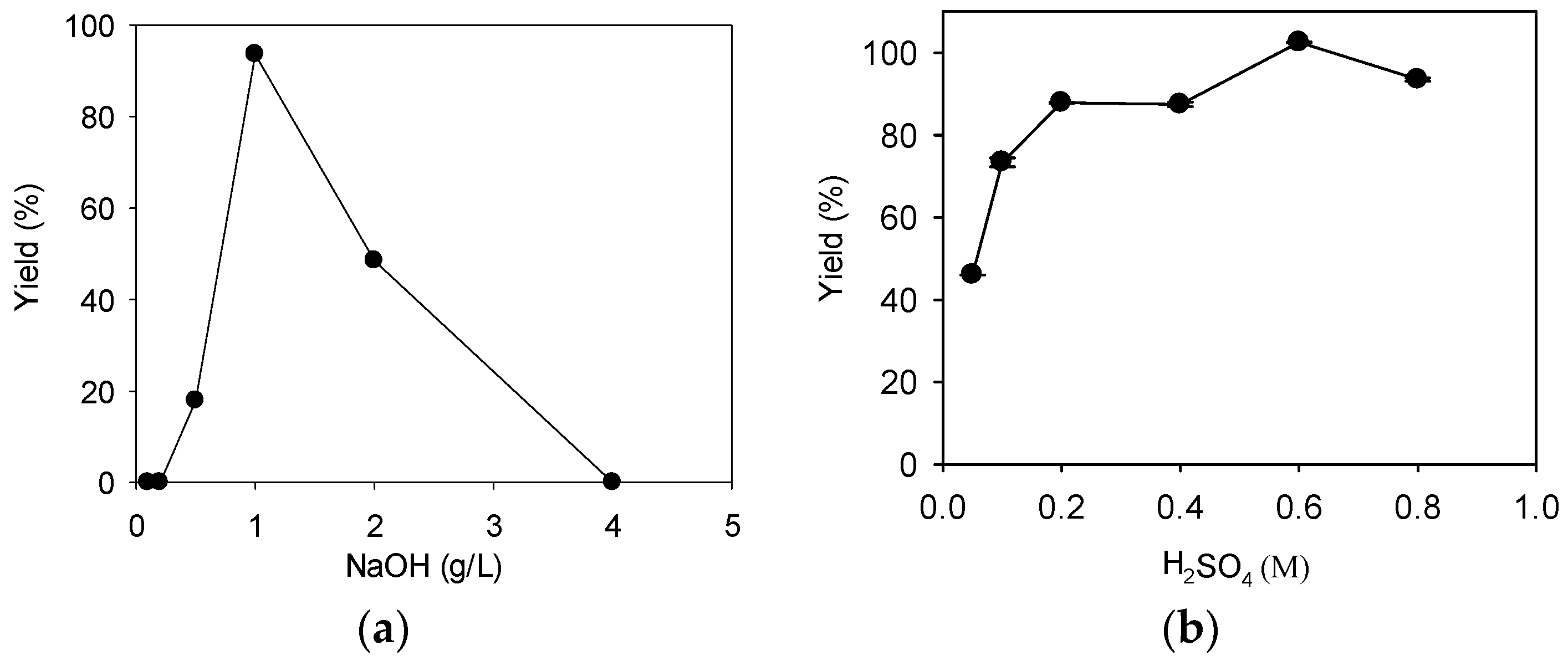
The average biomass concentration of R. glutinis from fermentation was 16 ± 5 g/L, with a lipid content of 39± 6%. Simple calculation revealed that 1 g of FAME is expected for 1 g of microbial lipid under complete transesterification. Thus the highest theoretical yield is 100%.When 0.1 and 0.2 g/L NaOH were used as catalysts, no FAME product was detected (Figure 1a). The added NaOH could be consumed by the saponification side reaction because there was an excess of yeast lipids. The FAME yield increased to 18% when the catalyst concentration was 0.5 g/L; the highest FAME yield of 94% was observed with 1 g/L NaOH. However, the FAME yield started to decrease rapidly as the catalyst concentration increased further. No FAME was observed with 4 g/L NaOH; a high alkaline catalyst concentration may facilitate saponification, thus leading to this low FAME yield [19]. Our results indicated that only a narrow range of NaOH concentration was suitable for FAME production. Similar results were observed by others using dry R. toruloides biomass as a feedstock [18]. The NaOH concentration of 1 g/L was selected for later experiments.
When 0.05 M H2SO4 was used as a catalyst, the FAME yield was only 46% (Figure 1b). The FAME yield increased with the concentration of H2SO4, and the highest FAME yield of 103% was observed with 0.6 M H2SO4. However, a further increase in the H2SO4 concentration to 0.8 M led to a decrease in FAME yield; a similar observation was made by others using a dry R. toruloides biomass as a feedstock, and the decrease in FAME yield was explained as a result of side reactions such as polymerization under harsh conditions [17]. The transesterification reaction mixture appeared orange when H2SO4 concentration was below 0.1 M, but the color changed to light yellow as the catalyst concentration increased further; the color change may be due to the oxidation of the caroteinoids present in the biomass [20]. The H2SO4 concentration of 0.6 M was selected for later experiments.
2.2. Effect of Reaction Time and Temperature on Transesterification
The FAME yield obtained by NaOH catalysis was only 49% after 2 h of incubation, while increasing dramatically up to a yield of 102% after 10 h of incubation (Figure 2). Further extension of the reaction time did not improve the yield, indicating that the reaction reached completion after 10 h. When H2SO4 was used as a catalyst, the FAME yield increased with the reaction time of up to 20 h with a yield of 111%. More FAME was formed than the available lipid which could be explained by the following reasons: (1) The lipid content within the biomass was underestimated; (2) some phopholipids were also converted to FAME [21,22]. Further increase in the reaction time beyond 20 h did not improve the FAME yield. For later experiments, the reaction times of 10 and 20 h were used for NaOH and H2SO4 catalysts, respectively.
With a 10 h reaction time and NaOH as a catalyst; the FAME yields at 50 and 60 °C were only 61% and 94%, respectively; even increasing the reaction duration up to 20 h did not enhance the FAME yield. Thus, 70 °C was optimal for FAME production catalyzed by NaOH. A similar observation was also obtained with catalysis by H2SO4. A previous study reported an optimal temperature of 50 °C for converting the dry biomass of oleaginous R. toruloides directly to biodiesel within a lidded test tube [18]. The difference in the optimal reaction temperature might be attributed to a different reaction setup. In later experiments, for both NaOH and H2SO4, a reaction temperature of 70 °C was used.
2.3. Effect of Methanol Loading on Transesterification
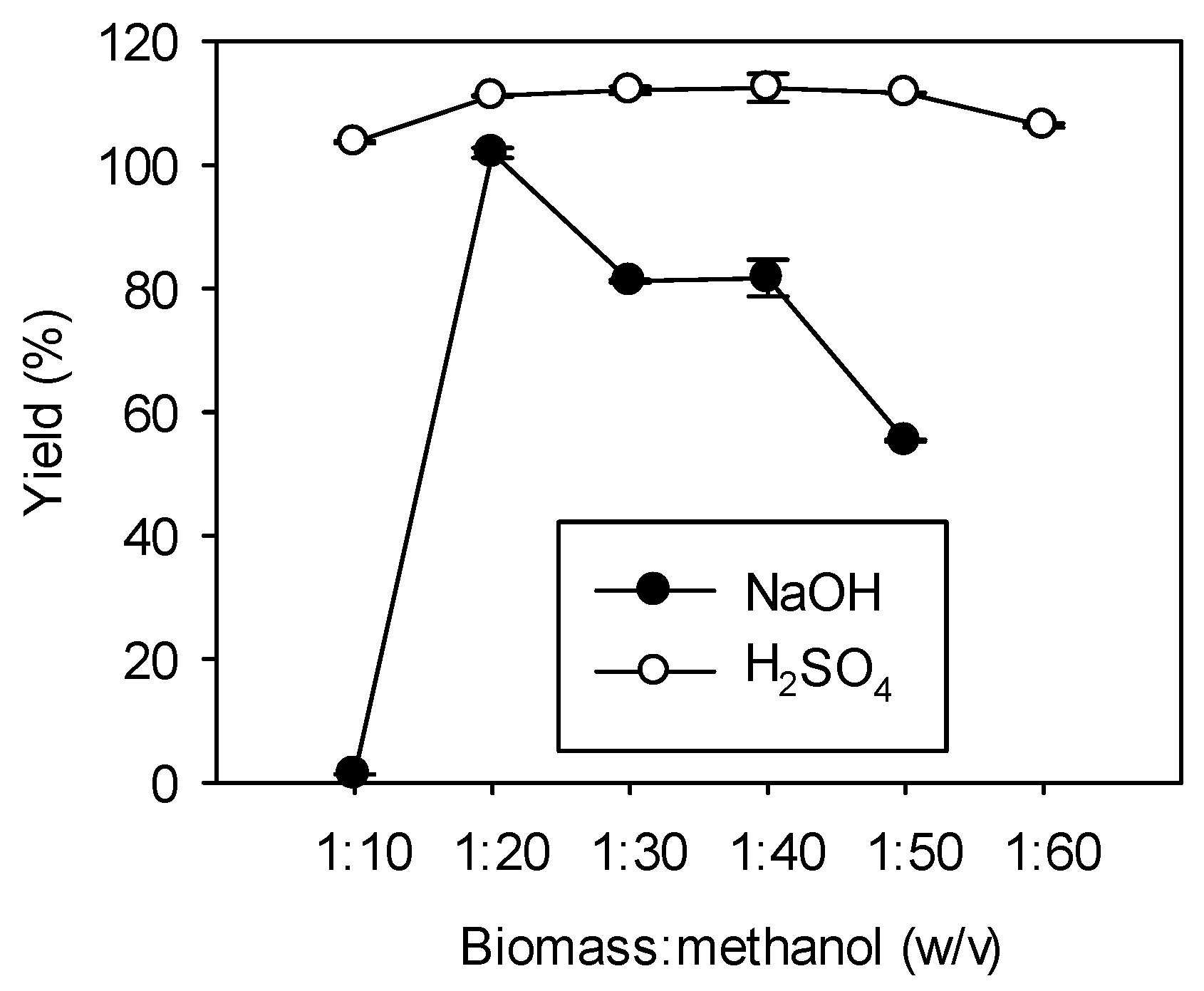
Methanol serves as one of the reactants and also acts as a solvent to weaken and disrupt the cell walls [23]. The effect of methanol loading on transesterification is shown in Figure 3. When NaOH was used as a catalyst, almost no FAME was formed with a biomass to methanol ratio (w/v) of 1:10; this could be because themethanol loading is insufficient to disrupt the cell walls effectively. When the biomass to methanol ratio was increased to 1:20, the highest yield of 102% was obtained. Nevertheless, the yield started to decrease as the ratio was further increased; similar results were also observed by others [24]. One possible explanation for the decreased yield at high methanol loading is that excess methanol dilutes the biomass and catalyst concentrations, thus giving rise to a lower conversion [18,24]. On the other hand, when H2SO4 was employed as a catalyst, the methanol loading had little influence on the FAME yield within the range we studied. For both catalysts, a biomass to methanol ratio (w/v) of 1:20 was used for later experiments.
2.4. Effect of Moisture on Transesterification
In an esterification reaction, the moisture content should be minimized because the presence of water favors the generation of free fatty acids from the hydrolysis of triglycerides, thus enhancing the degree of saponification and lowering the FAME yield. Previous studies on the transesterification of vegetable oils have shown that the presence of water even at very low concentrations has a detrimental effect on the FAME yield [25]. To determine the effect of moisture content on the FAME yield, various amounts of water were added back to 1 g of dry biomass; the moisture content was defined by the following equation:
moisture content (%) = amount of added water (g)/(amount of added water (g) +
1 g of biomass)
1 g of biomass)
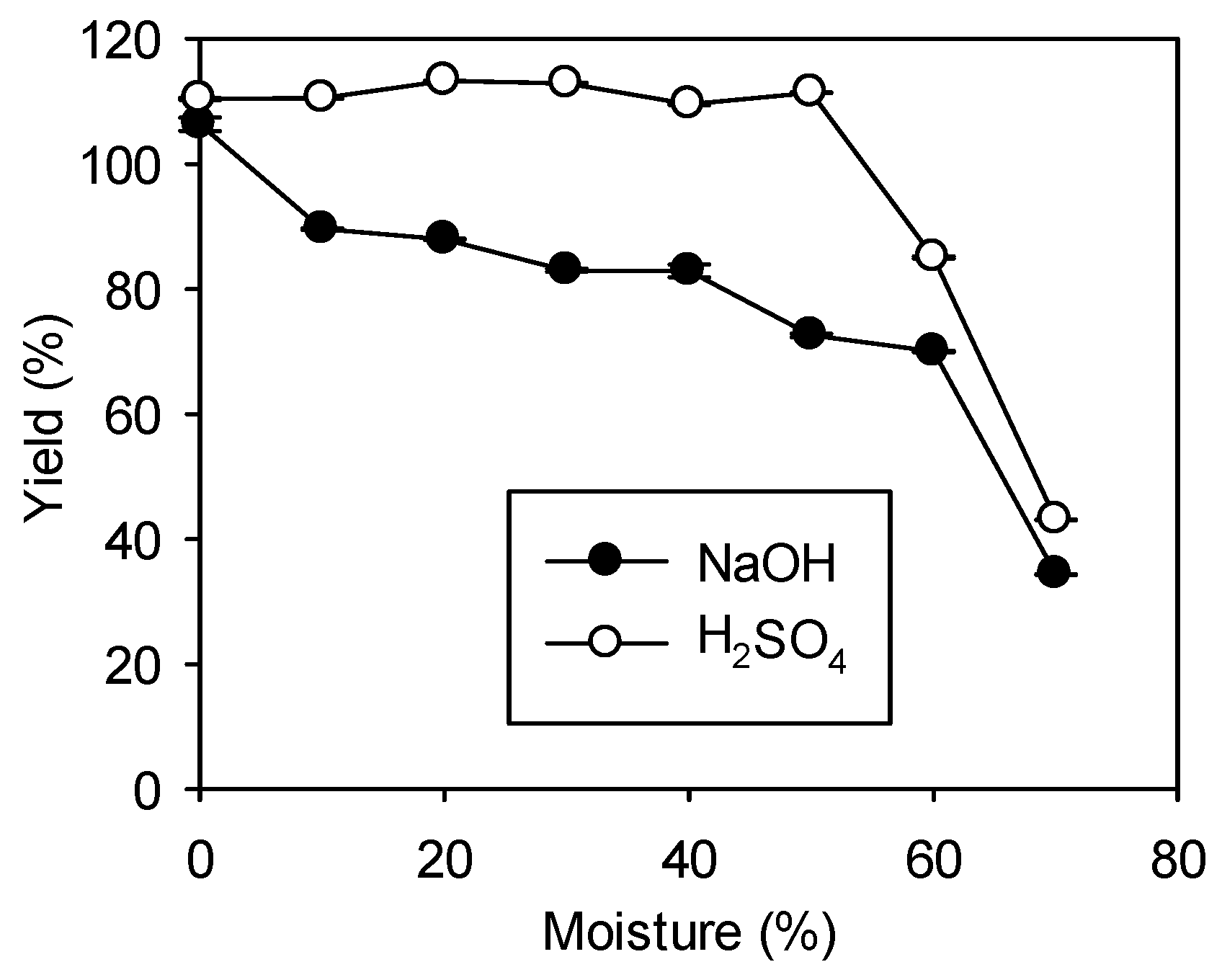
Using NaOH as a catalyst, the FAME yield was decreased with an increase in the moisture content (Figure 4); the FAME yield was 34% when the biomass contained 70% moisture. Our results were quite different from those reported for R. toruloides (also using NaOH as a catalyst), which showed that there was almost no formation of FAME with moisture content above 10% [18]. In the case of H2SO4, the process tolerated moisture fairly well up to 50%. However, the FAME yield decreased rapidly to 85% and 43% when the biomass contained 60% and 70% moisture, respectively. Similar results were obtained by others using dry microalgae biomass, Chaetoceros gracilis, as a feed stock with a higher methanol loading of 1:40 [21]. We have also examined the feasibility of using a wet R. glutinis biomass collected from centrifugation without drying (the moisture content was 72%) for direct transesterification. However, this included a heating step at 80°C for 10 min before centrifugation. Consequently, the FAME yield could reach 73%. Yellapu et al. reported that the yield could be further improved by pre-treating the wet biomass with surfactants such as N-lauroyl sarcosine, and the reaction time may also be shortened by carrying out the reaction under sonication [26].
2.5. Comparison of Yields and Compositions of FAME Derived from Different Transesterification Methods
The yields and compositions of FAME produced by different transesterification methods are listed in Table 1. The conventional methods resulted in lower FAME yields of 78% and 85% using NaOH and H2SO4 as catalysts, respectively; the acidic method had a yield similar to that of the previous report (81%), using the same biomass [27]. Direct transesterification, regardless of the catalyst used, resulted in significantly higher yields in a shorter time (approximately 4 h less) than conventional methods, which involved cell disruption, lipid extraction, and transesterification in that order. For direct transesterification, the compositions of FAME obtained with either catalyst were almost the same; the oleic acid methyl ester was the dominant FAME, followed by linoleic, and then palmitic acid methyl ester. However, a previous report showed that the FAME composition was catalyst dependent, especially for oleic and linolenic acid methyl esters [18]. The content of linoleic acid methyl ester from direct transesterification was about 20% higher than the linoleic acid content of R. glutinis cultured with glycerol as a carbon source [13,14]. The higher amount of linoleic acid methyl ester was explained by the fact that some FAME was derived from the phospholipids in the cell membrane [22,23]. When conventional methods were used, the linoleic acid methyl ester was the dominant FAME, followed by oleic acid methyl ester instead.
3. Materials and Methods
3.1. Microorganism and Medium
The seed medium for culturing R. glutinis BCRC (Bioresource Collection and Research Center, Hsinchu, Taiwan) 22,360 contained 3 g of yeast extract, 3 g of malt extract, 5 g of peptone, and 10 g of dextrose per liter. The fermentation medium contained 80 g of crude glycerol (Yu-Hwa Biodiesel Company, Changhua, Taiwan), 2 g of yeast extract, 2 g of (NH4)2SO4, 1 g of KH2PO4, 0.5 g of MgSO4·7H2O, 0.1 g of CaCl2, and 0.1 g of NaCl per liter. The crude glycerol, dark brown in color and slightly viscous, was used directly without any pretreatment or purification.
3.2. Production of Biomass
Under aerobic conditions, 300 mL of seed culture was prepared in a seed medium after a 24-h incubation at 24 °C with orbital shaking at 150 rpm. The seed culture was inoculated into a 5-L stirred-tank fermentor (BIOSTAT® A plus, Sartorius, Gottingen, Germany) containing 2.7 L of fermentation medium (3 L working volume). The pH was maintained at 5.5 by automatically feeding 1.0 N NaOH or 1.0 N HCl solution into the medium. The fermentor was operated at 24 °C with dissolved oxygen controlled at 30, ±2% of the saturation level. The agitation during the process was limited to a range from 200 to 400 rpm to avoid potential damage resulting from high shear force. The biomass was harvested after 72 h of incubation by centrifugation at 12,021× g, and then the pellets were washed with deionized water to remove any residual fermentation medium. The biomass was stored at −80 °C for 24 h before lyophilized in a Freezemobile 12 XL freeze dryer (Virtis). The dried biomass was stored in a refrigerator for later use.
3.3. Total Lipid Analysis
The extraction of lipids from a biomass was modified from the procedure reported by Bligh and Dyer [28]. Fifty milliliters of chloroform/methanol mixture (2:1, v/v) was mixed with 0.5 g of finely ground powder of dry biomass. The suspension was then subject to ultrasonication with a Misonix XL2020 sonicator for 6 min on an ice bath; the ultrasonic horn was directly immersed in the suspension and the on/off cycle was set to 60/60 s in order to minimize the heat generated. The suspension was incubated at room temperature for 1 h on a rocking mixer at 100 rpm. The solvent phase was collected by centrifugation at 12,100× g for 10 min; the biomass was extracted with an additional 30 mL of chloroform/methanol mixture for 1 h. The combined solvent phase was evaporated at 65 °C under vacuum (337 mbar), and then the remaining lipid was weighted. The lipid content was defined as below:
lipid content (%) = weight of lipid (g)/weight of biomass (g) × 100
3.4. Direct Transesterification
One gram of dry biomass and various amounts of methanol (biomass to methanol ratio in the range of 1:10 to 1:60, w/v) were added to a 100 mL round-bottom flask fitted with a condenser. NaOH (from 0.1 to 4 g/L) or H2SO4 (from 0.05 to 0.8 M) dissolved in methanol was used as a catalyst. The suspension was heated at 40 to 70 °C in a water bath under atmospheric pressure with vigorous mixing for 2 to 30 h, followed by a centrifugation at 12,100× g for 10 min and a collection of the supernatant. The residual biomass was then washed with 10 mL of n-hexane and the washing fraction was collected after centrifugation as described above. The supernatant and the washing fraction were combined and extracted with equal volume of n-hexane. The n-hexane layer was collected and the solvent was evaporated at 65 °C under vacuum (337 mbar). The biodiesel product was collected and weighted. The experiments were performed in triplicate; the average from triplicate was plotted in the figures and the standard deviation was indicated with an error bar.
3.5. Analysis of Fatty Acid Methyl Esters
The content of FAME in the transesterification product was measured according to the standard of Taiwan, CNS-15051. The biodiesel product was first treated with sodium sulfate followed by centrifugation at 2040× g for 5 min. Fifty microliters of the treated sample was mixed with 1 mL of 10 mg/mL methyl heptadecanoate in hexane as an internal standard. One microliter of the sample was injected into a gas chromatograph (GC-2014, Shimadzu, Kyoto, Japan) equipped with a flame-ionization detector (FID). A BPX70 capillary column (30 m × 0.25 mm i.d.; SGE Analytical Science, Ringwood, Australia) with nitrogen as carrier gas was used. The injector and FID temperatures were both set at 250 °C. The oven temperature was initially held at 150 °C for 30 s and then increased to 180 °C at 10 °C/min, finally to 198 °C at 1.5 °C/min. The content of FAME was calculated using the following equation:
where ΣA is the summation of peak area of all the FAME peaks (from C14:0 to C24:1); AEI is the peak area of internal standard, methyl heptadecanoate; CEI is the concentration of methyl heptadecanoate, VEI is the volume of methyl heptadecanoate, and m is the mass of the biodiesel sample. The FAME yield was calculated using the following equation:
4. Conclusions
In this study, the oleaginousyeast, R. glutinis, was used for the direct transesterification without lipid pre-extraction to produce FAME. The use of chloroform, a highly toxic organic solvent often applied in lipid extraction, was avoided. The highest FAME yield of 111% was obtained with 0.6 M H2SO4 as a catalyst and a methanol loading of 1:20 (biomass to methanol, w/v), the reaction was carried out at 70 °C for 20 h. Using the same methanol loading and temperature, a slightly lower yield of 102% was obtained with 1 g/L NaOH after 10 h of incubation. Although basic catalysts are often avoided because of the potential saponification caused by the free fatty acids present in SCO, the NaOH catalyst was still able to achieve similar FAME yield in half the reaction time required by the H2SO4 catalyst. However, the acid-catalyzed direct transesterification tolerates moisture content much better than the base-catalyzed process; thus, the H2SO4 catalyst is more suitable for wet biomass feedstocks. Compared to conventional transesterification, the direct process is not only simpler, but also improves the FAME yield by 24–29% with a shorter reaction time and a lower energy consumption. The reaction processes reported in this work simplify for the production of SCO-derived biodiesel and reduce its cost. Our preliminary work also indicates that wet biomass can be used directly as a feedstock with some minor adjustments in reaction conditions, suggesting that further process simplification and cost reduction are possible.
Author Contributions
Chi-Yang Yu and I-Ching Kuan designed the experiments and analyzed the data. Wei-Chen Kao and Chun-Ling Chen performed the experiments using the basic and acidic catalyst, respectively. Chi-Yang Yu and I-Ching Kuan prepared the manuscript. All authors read and approved the final manuscript. All authors contributed equally to this work.
Acknowledgments
We thank Hong-Wei Yen (Department of Chemical and Materials Engineering, Tunghai University, Taiwan) and Yaw-Nan Chang (Department of Biotechnology, National Formosa University, Taiwan) for providing R. glutinis BCRC 22360.This work was supported by the Ministry of Science and Technology (MOST 105-2621-M-036-001-MY2) in Taiwan.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sitepu, I.R.; Garay, L.A.; Sestric, R.; Levin, D.; Block, D.E.; German, J.B.; Boundy-Mills, K.L. Oleaginous yeasts for biodiesel: Current and future trends in biology and production. Biotechnol. Adv. 2014, 32, 1336–1360. [Google Scholar] [CrossRef] [PubMed]
- Canakci, M.; Sanli, H. Biodiesel production from various feedstocks and their effects on the fuel properties. J. Ind. Microbiol. Biotechnol. 2008, 35, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, H.; Kondo, A.; Noda, H. Biodiesel fuel production by transesterification of oils. J. Biosci. Bioeng. 2001, 92, 405–416. [Google Scholar] [CrossRef]
- Demirbas, A. Importance of biodiesel as transportation fuel. Energy Policy 2007, 35, 4661–4670. [Google Scholar] [CrossRef]
- Escobar, J.C.; Lora, E.S.; Venturini, O.J.; Yáñez, E.E.; Castillo, E.F.; Almazan, O. Biofuels: Environment, technology and food security. Renew. Sustain. Energy Rev. 2009, 13, 1275–1287. [Google Scholar] [CrossRef]
- Kulkarni, M.G.; Dalai, A.K. Waste cooking oil-an economical source for biodiesel: A review. Ind. Eng. Chem. Res. 2006, 45, 2901–2913. [Google Scholar] [CrossRef]
- Huang, C.; Chen, X.-F.; Xiong, L.; Chen, X.-D.; Ma, L.-L.; Chen, Y. Single cell oil production from low-cost substrates: The possibility and potential of its industrialization. Biotechnol. Adv. 2013, 31, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Yeh, K.-L.; Aisyah, R.; Lee, D.-J.; Chang, J.-S. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Yang, J.; Xu, X.; Zhang, L.; Nie, Q.; Xian, M. Biodiesel production from oleaginous microorganisms. Renew. Energy 2009, 34, 1–5. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Yousuf, A.; Khan, M.R.; Islam, M.A.; Wahid, Z.A.; Pirozzi, D. Technical difficulties and solutions of direct transesterification process of microbial oil for biodiesel synthesis. Biotechnol. Lett. 2017, 39, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C.; Cohen, Z. Microbial and algal oils: Do they have a future for biodiesel or as commodity oils? Lipid Technol. 2008, 20, 155–160. [Google Scholar] [CrossRef]
- Yen, H.-W.; Zhang, Z. Effects of dissolved oxygen level on cell growth and total lipid accumulation in the cultivation of Rhodotorula glutinis. J. Biosci. Bioeng. 2011, 112, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.-W.; Chang, J.-T. Growth of oleaginous Rhodotorula glutinis in an internal-loop airlift bioreactor by using lignocellulosic biomass hydrolysate as the carbon source. J. Biosci. Bioeng. 2015, 119, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula glutinis—Potential source of lipids, carotenoids, and enzymes for use in industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef] [PubMed]
- Cheirsilp, B.; Louhasakul, Y. Industrial wastes as a promising renewable source for production of microbial lipid and direct transesterification of the lipid into biodiesel. Bioresour. Technol. 2013, 142, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhao, Z. Biodiesel production by direct methanolysis of oleaginous microbial biomass. J. Chem. Technol. Biotechnol. 2007, 82, 775–780. [Google Scholar] [CrossRef]
- Thliveros, P.; Uçkun Kiran, E.; Webb, C. Microbial biodiesel production by direct methanolysis of oleaginous biomass. Bioresour. Technol. 2014, 157, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y.C.; Guo, Y. Transesterification of neat and used frying oil: Optimization for biodiesel production. Fuel Process. Technol. 2006, 87, 883–890. [Google Scholar] [CrossRef]
- Boon, C.S.; McClements, D.J.; Weiss, J.; Decker, E.A. Factors influencing the chemical stability of carotenoids in foods. Crit. Rev. Food Sci. Nutr. 2010, 50, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Wahlen, B.D.; Willis, R.M.; Seefeldt, L.C. Biodiesel production by simultaneous extraction and conversion of total lipids from microalgae, cyanobacteria, and wild mixed-cultures. Bioresour. Technol. 2011, 102, 2724–2730. [Google Scholar] [CrossRef] [PubMed]
- Vicente, G.; Bautista, L.F.; Rodríguez, R.; Gutiérrez, F.J.; Sádaba, I.; Ruiz-Vázquez, R.M.; Torres-Martínez, S.; Garre, V. Biodiesel production from biomass of an oleaginous fungus. Biochem. Eng. J. 2009, 48, 22–27. [Google Scholar] [CrossRef]
- Zhang, X.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y.; Valéro, J.R. Ultrasonication aided in-situ transesterification of microbial lipids to biodiesel. Bioresour. Technol. 2014, 169, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.D.; Gude, V.G.; Mannarswamy, A.; Cooke, P.; Munson-McGee, S.; Nirmalakhandan, N.; Lammers, P.; Deng, S. Optimization of microwave-assisted transesterification of dry algal biomass using response surface methodology. Bioresour. Technol. 2011, 102, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Canakci, M.; Van Gerpen, J. Biodiesel production via acid catalysis. Trans. ASAE 1999, 42, 1203–1210. [Google Scholar] [CrossRef]
- Yellapu, S.K.; Kaur, R.; Tyagi, R.D. Detergent assisted ultrasonication aided in situ transesterification for biodiesel production from oleaginous yeast wet biomass. Bioresour. Technol. 2017, 224, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.-C.; Tao, J.; Xie, F.; Dai, Y.-J.; Zhao, M. Biodiesel generation from oleaginous yeast Rhodotorula glutinis with xylose assimilating capacity. Afr. J. Biotechnol. 2007, 6, 2130–2134. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The effect of catalyst concentration on fatty acid methyl ester (FAME) yield. (a) NaOH; (b) H2SO4. For both catalysts, direct transesterification on 1 g of dry R. glutinis biomass was carried out with a methanol loading of 1:20 (biomass: methanol, w/v) at 70 °C for 15 h. Data were obtained by averaging three individual measurements and the standard deviations were indicated with error bars.
Figure 1.
The effect of catalyst concentration on fatty acid methyl ester (FAME) yield. (a) NaOH; (b) H2SO4. For both catalysts, direct transesterification on 1 g of dry R. glutinis biomass was carried out with a methanol loading of 1:20 (biomass: methanol, w/v) at 70 °C for 15 h. Data were obtained by averaging three individual measurements and the standard deviations were indicated with error bars.
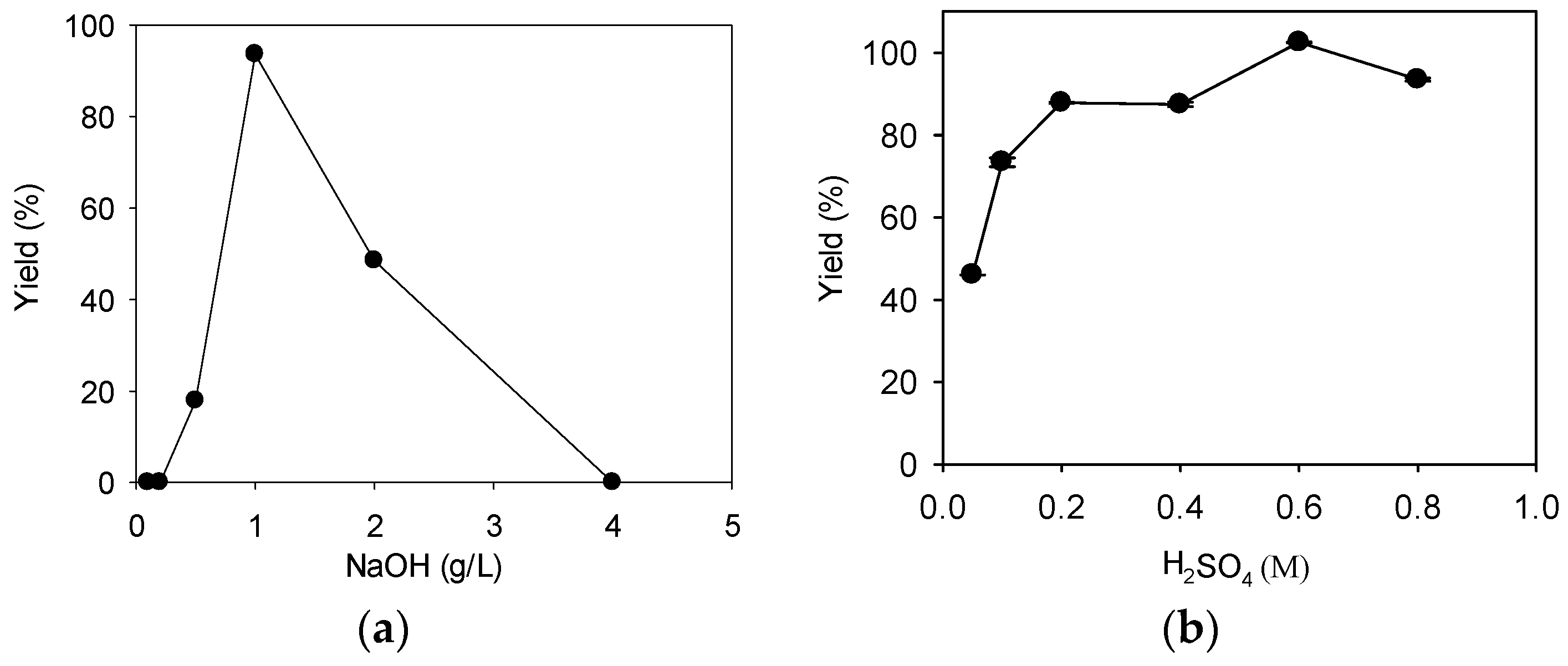
Figure 2.
The effect of reaction time on FAME yield. Direct transesterification on 1 g of dry R. glutinis biomass was carried out with a methanol loading of 1:20 (biomass: methanol, w/v) at 70°C using 1 g/L NaOH or 0.6 M H2SO4 as a catalyst. Data were obtained by averaging three individual measurements and the standard deviations were indicated with error bars.
Figure 2.
The effect of reaction time on FAME yield. Direct transesterification on 1 g of dry R. glutinis biomass was carried out with a methanol loading of 1:20 (biomass: methanol, w/v) at 70°C using 1 g/L NaOH or 0.6 M H2SO4 as a catalyst. Data were obtained by averaging three individual measurements and the standard deviations were indicated with error bars.

Figure 3.
The effect of methanol loading on transesterification. Direct transesterification on 1 g of dry R. glutinis biomass was carried out at 70 °C with 1 g/L NaOH and 0.6 M H2SO4 for 10 and 20 h, respectively. Data were obtained by averaging three individual measurements and the standard deviations were indicated with error bars.
Figure 3.
The effect of methanol loading on transesterification. Direct transesterification on 1 g of dry R. glutinis biomass was carried out at 70 °C with 1 g/L NaOH and 0.6 M H2SO4 for 10 and 20 h, respectively. Data were obtained by averaging three individual measurements and the standard deviations were indicated with error bars.

Figure 4.
The effect of moisture on FAME yield. Direct transesterification on 1 g of dry R. glutinis biomass was carried out at 70 °C with 1 g/L NaOH and 0.6 M H2SO4 for 10 and 20 h, respectively. A methanol loading of 1:20 (biomass: methanol, w/v) was used for both catalysts. Data were obtained by averaging three individual measurements and the standard deviations were indicated with error bars.
Figure 4.
The effect of moisture on FAME yield. Direct transesterification on 1 g of dry R. glutinis biomass was carried out at 70 °C with 1 g/L NaOH and 0.6 M H2SO4 for 10 and 20 h, respectively. A methanol loading of 1:20 (biomass: methanol, w/v) was used for both catalysts. Data were obtained by averaging three individual measurements and the standard deviations were indicated with error bars.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Yields and compositions of FAME derived from R. glutinis using different transesterification methods.
Table 1.
Yields and compositions of FAME derived from R. glutinis using different transesterification methods.
| Method | FAME Yield (%) a | FAME Composition (%) | |||||
|---|---|---|---|---|---|---|---|
| C16:0 | C16:1 | C18:0 | C18:1 | C18:2 | C18:3 | ||
| Direct acid-catalyzed | 111.0 ± 0.1 | 16.8 | 0.7 | 4.1 | 42.2 | 33.8 | 2.5 |
| Direct base-catalyzed | 102.0 ± 0.8 | 18.8 | 0.7 | 4 | 41.1 | 32.9 | 2.6 |
| Conventional acid-catalyzed b | 85.2 ± 0.9 | 18.1 | 1.6 | 3 | 33.3 | 41.1 | 4.1 |
| Conventional base-catalyzed c | 77.9 ± 0.7 | 21 | 1 | 2.5 | 30.5 | 41.2 | 3.7 |
a Data were obtained by averaging three individual measurements. b The lipid was first extracted as described in Methods (Total lipid analysis) except that 1 g of dry biomass was used and reagents required were scaled-up accordingly. Transesterification of extracted lipid was carried out with 0.6 M H2SO4 for 20 h at 70 °C, using a methanol loading of 1:20 (biomass: methanol, w/v). c The lipid was first extracted as described in Methods (Total lipid analysis) except that 1 g of dry biomass was used and the reagents required were scaled-up accordingly. The transesterification of extracted lipid was carried out with 1 g/L NaOH for 10 h at 70 °C, using a methanol loading of 1:20 (biomass: methanol, w/v).
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kuan, I.-C.; Kao, W.-C.; Chen, C.-L.; Yu, C.-Y. Microbial Biodiesel Production by Direct Transesterification of Rhodotorula glutinis Biomass. Energies 2018, 11, 1036. https://doi.org/10.3390/en11051036
AMA Style
Kuan I-C, Kao W-C, Chen C-L, Yu C-Y. Microbial Biodiesel Production by Direct Transesterification of Rhodotorula glutinis Biomass. Energies. 2018; 11(5):1036. https://doi.org/10.3390/en11051036
Chicago/Turabian StyleKuan, I-Ching, Wei-Chen Kao, Chun-Ling Chen, and Chi-Yang Yu. 2018. "Microbial Biodiesel Production by Direct Transesterification of Rhodotorula glutinis Biomass" Energies 11, no. 5: 1036. https://doi.org/10.3390/en11051036
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.