Over-Expression of CYP2E1 mRNA and Protein: Implications of Xenobiotic Induced Damage in Patients with De Novo Acute Myeloid Leukemia with inv(16)(p13.1q22); CBFβ-MYH11


Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Selection
2.2. Conventional Cytogenetic Analysis
2.3. Real-Time Quantitative Reverse Transcriptase Polymerase Chain Reaction (qRT-PCR) of CBFβ-MYH11 Transcripts, CYP2E1, CYP1A2 and NQO1
2.4. Immunohistochemical Analysis
2.5. Cell Culture, Treatment and Quantitative Real-Time RT-PCR Assay
2.6. Statistical Analysis
3. Results
3.1. Clinical Findings

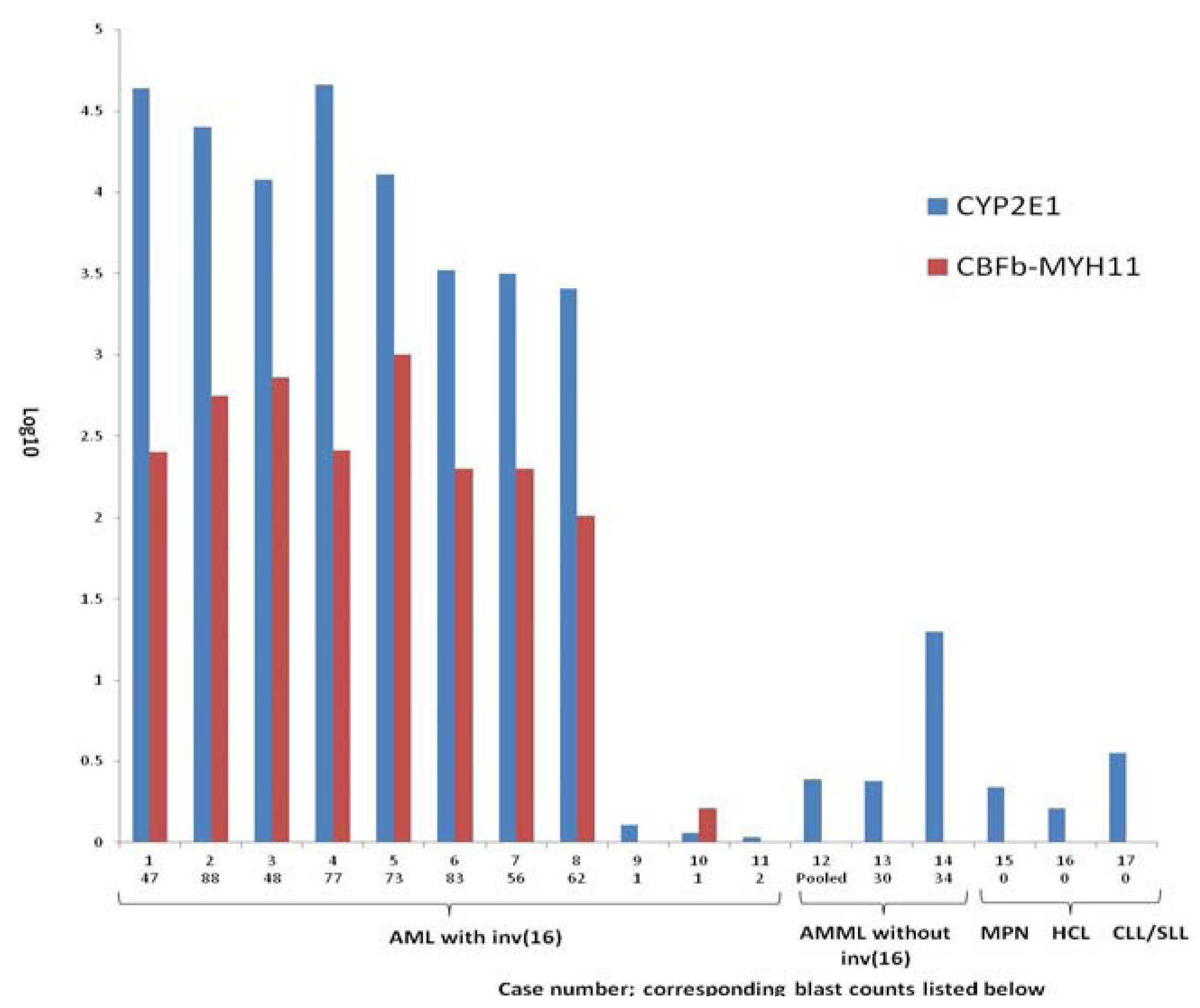
{kind=link}
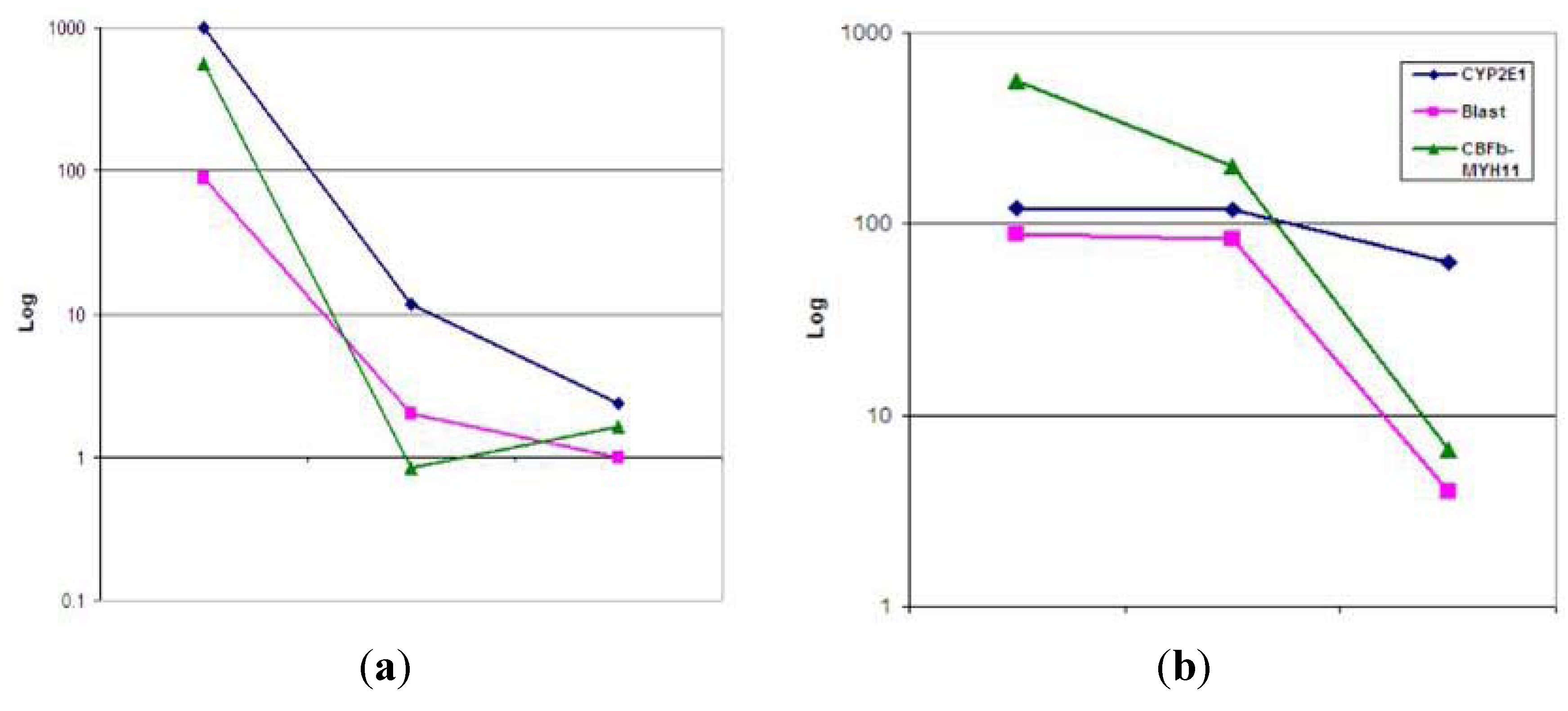{kind=link}
{kind=link}
{kind=link}
| Age/Sex | Blast % | Eo % | Log10 2E1/actin | Log10 CBFB/MYH11 | F/U | Other genetic mutations | Conventional cytogenetics on BM |
|---|---|---|---|---|---|---|---|
| 45/M | 47 | 7 | 4.64 | 2.40 | Alive | None | 46,XY,inv(16)(p13.1q22) [1] 47,XY,inv(16)(p13.1q22),+22 [2] |
| 40/M | 88 | 3 | 4.40 | 2.75 | Dead | NRAS | 46,XY,inv(16)(p13.1q22) [20] |
| 10/F | 48 | 12 | 4.08 | 2.86 | Alive | None | 46,XX,inv(16)(p13.1q22) [19]/46,XX,+8,inv(16)(p13.2q22) [1] |
| 24/M | 77 | 6 | 4.66 | 2.41 | Alive | None | 46,XY,inv(16)(p13q22) [20] |
| 36/M | 73 | 8 | 4.11 | 3.00 | Alive | NRAS | 46,XY,inv(16)(p13q22) [15] 46,XY [5] |
| 51/M | 83 | 0 | 3.52 | 2.30 | Alive | FLT3 | 46,XX,inv(16)(p13.1q22) [20] |
| 65/M | 56 | 10 | 3.50 | 2.30 | Alive | None | 46,XY,inv(16)(p13.1q22) [20] |
| 35/M | 62 | 5 | 3.41 | 2.01 | Alive | NRAS | 46,XY,inv(16)(p13.1q22) [20] |
| 53/M | 1 | 3 | 0.11 | 0.00 | Alive | None | 46,XY,inv(16)(p13.1q22) [1]/46,XY, t(6;20)(p21.3;q11.2) inv(16)(p13.1q22) [9] /46,XY [10] |
| 26/M | 1 | 1 | 0.06 | 0.21 | Alive | None | BM, 46,XY; Soft tissue myeloid sarcoma showed inv(16) by FISH (Conventional karyotyping ND) |
| 45/F | 2 | 3 | 0.03 | 0.00 | Alive | None | BM, 46,XY [19]; history of inv(16); current BM is in CR |
3.2. Real-Time qRT-PCR Findings of CBFβ-MYH11 Transcripts, CYP2E1, CYP1A2 and NQO1

| Category | CYP2E1/ABL | CYP1A2/ABL * | NQO1/ABL * |
|---|---|---|---|
| Mean (n = 7) | 25.32 | 0.77 | 52.11 |
| R2 | −0.506 | 0.934 |

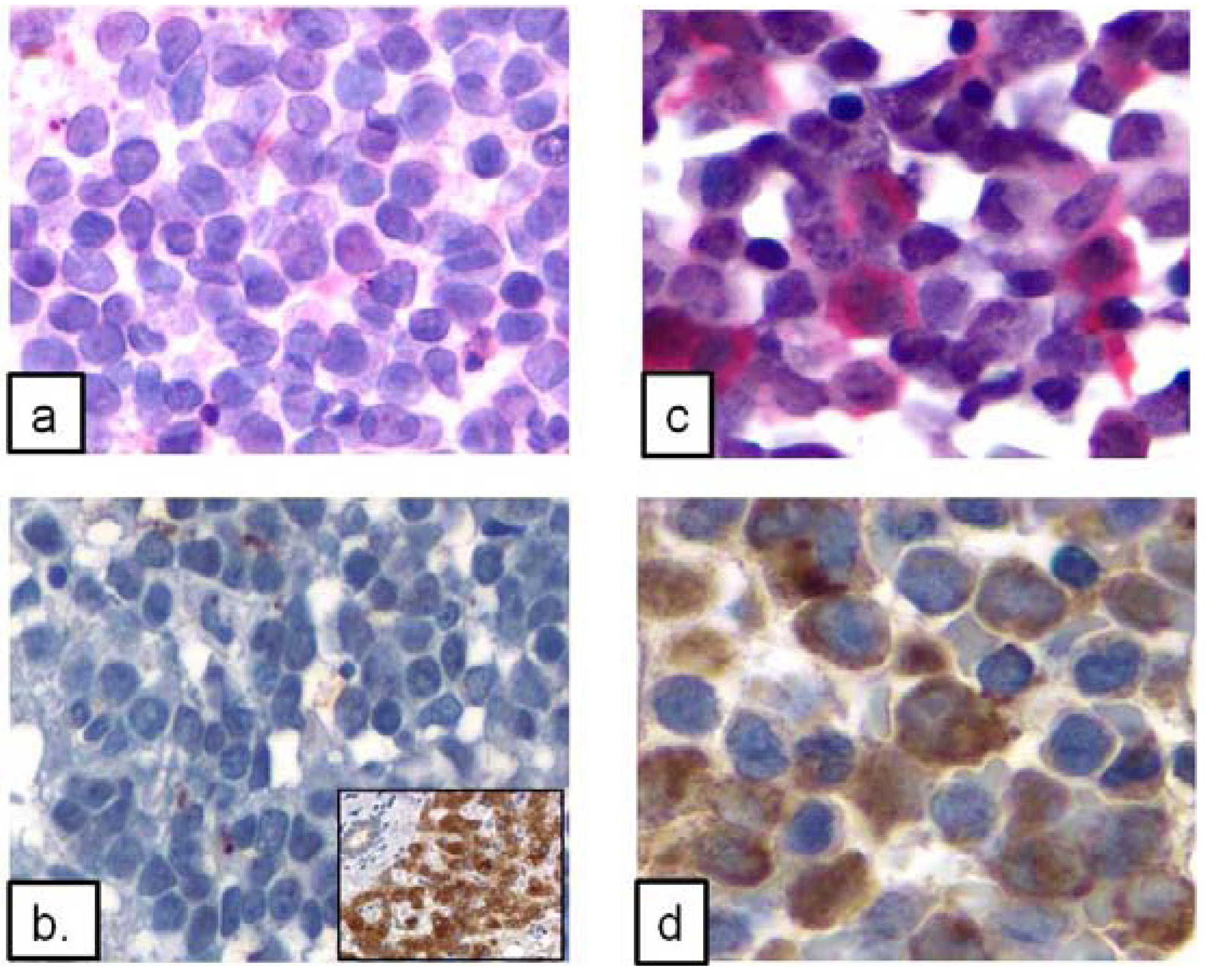
3.3. Immunohistochemical Findings
| Category | AML with inv(16) (n = 27) | AMML without inv(16) * (n = 10) | Other types of AML * (n = 22) |
|---|---|---|---|
| Strong | 20 | 1 | 7 |
| Weak/Focal | 5 | 3 | 3 |
| Negative | 2 | 6 | 12 |
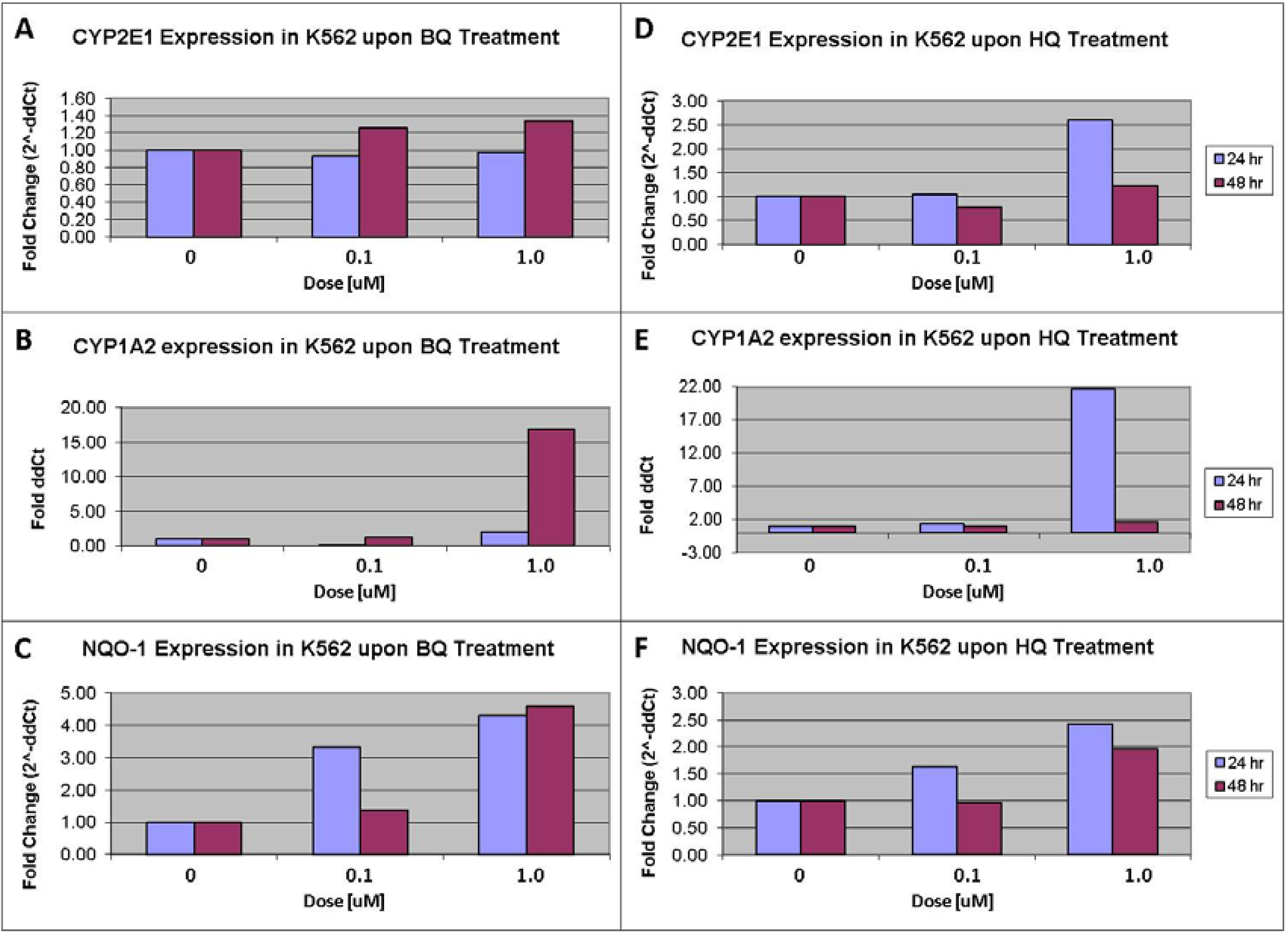
3.4. Cell Culture, Treatment and Quantitative Real-Time RT-PCR Assay
4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Huff, J. Benzene-induced cancers: Abridged history and occupational health impact. Int. J. Occup. Environ. Health 2007, 13, 213–221. [Google Scholar]
- Kok, P.W.; Ong, C.N. Blood and urinary benzene determined by headspace gas chromatography with photoionization detection: Application in biological monitoring of low-level nonoccupational exposure. Int. Arch. Occup. Environ. Health 1994, 66, 195–201. [Google Scholar] [CrossRef]
- Brugnone, F.; Perbellini, L.; Giuliari, C.; Cerpelloni, M.; Soave, M. Blood and urine concentrations of chemical pollutants in the general population. Med. Lav. 1994, 85, 370–389. [Google Scholar]
- Bennett, J.M. The Myelodysplastic Syndromes: Pathobiology and Clinical Management. In Basic and Clinical Oncology; Cheson, B., Ed.; Marcel Dekker: New York, NY, USA, 2002; Volume 27, pp. 36–40. [Google Scholar]
- Smith, M.T.; Wang, Y.; Kane, E.; Rollinson, S.; Wiemels, J.L.; Roman, E.; Roddam, P.; Cartwright, R.; Morgan, G. Low NAD(P)H: Quinone oxidoreductase 1 activity is associated with increased risk of acute leukemia in adults. Blood 2001, 97, 1422–1426. [Google Scholar]
- Rothman, N.; Smith, M.T.; Hayes, R.B.; Traver, R.D.; Hoener, B.; Campleman, S.; Li, G.L.; Dosemeci, M.; Linet, M.; Zhang, L. Benzene poisoning, a risk factor for hematological malignancy, is associated with the NQO1 609C→T mutation and rapid fractional excretion of chlorzoxazone. Cancer Res. 1997, 57, 2839–2842. [Google Scholar]
- Aydin-Sayitoglu, M.; Hatirnaz, O.; Erensoy, N.; Ozbek, U. Role of CYP2D6, CYP1A1, CYP2E1, GSTT1, and GSTM1 genes in the susceptibility to acute leukemias. Am. J. Hematol. 2006, 81, 162–170. [Google Scholar] [CrossRef]
- Zhang, J.; Yin, L.H.; Liang, G.Y.; Liu, R.; Fan, K.H.; Pu, Y.P. Detection of CYP2E1, a genetic biomarker of susceptibility to benzene metabolism toxicity in immortal human lymphocytes derived from the Han Chinese population. Biomed. Environ. Sci. 2010, 24, 300–309. [Google Scholar]
- Bolufer, P.; Collado, M.; Barragan, E.; Calasanz, M.J.; Colomer, D.; Tormo, M.; González, M.; Brunet, S.; Batlle, M.; Cervera, J. Profile of polymorphisms of drug-metabolising enzymes and the risk of therapy-related leukaemia. Br. J. Haematol. 2007, 136, 590–596. [Google Scholar] [CrossRef]
- Han, X.; Zheng, T.; Foss, F.M.; Lan, Q.; Holford, T.R.; Rothman, N.; Ma, S.; Zhang, Y. Genetic polymorphisms in the metabolic pathway and non-Hodgkin lymphoma survival. Am. J. Hematol. 2010, 85, 51–56. [Google Scholar]
- Ingelman-Sundberg, M.; Ekström, G.; Tindberg, N. Lipid peroxidation dependent on ethanol-inducible cytochrome p-450 from rat liver. Adv. Biosci. 1988, 71, 43–47. [Google Scholar]
- Weltman, M.D.; Farrell, G.C.; Hall, P.; Ingelman-Sundberg, M.; Liddle, C. Hepatic cytochrome p450 2e1 is increased in patients with nonalcoholic steatohepatitis. Hepatology 1998, 27, 128–133. [Google Scholar] [CrossRef]
- Larson, R.A.; Wang, Y.; Banerjee, M.; Wiemels, J.; Hartford, C.; Beau, M.M.L.; Smith, M.T. Prevalence of the inactivating 609C→T polymorphism in the NAD(P)H: Quinone oxidoreductase (NQO1) gene in patients with primary and therapy-related myeloid leukemia. Blood 1999, 94, 803–807. [Google Scholar]
- Rothman, N.; Li, G.L.; Dosemeci, M.; Bechtold, W.E.; Marti, G.E.; Wang, Y.Z.; Linet, M.; Xi, L.; Lu, W.; Smith, M.T. Hematotoxocity among Chinese workers heavily exposed to benzene. Am. J. Ind. Med. 1996, 29, 236–246. [Google Scholar] [CrossRef]
- Wiemels, J.L.; Pagnamenta, A.; Taylor, G.M.; Eden, O.B.; Alexander, F.E.; Greaves, M.F. A lack of a functional NAD(P)H: Quinone oxidoreductase allele is selectively associated with pediatric leukemias that have MLL fusions. Cancer Res. 1999, 59, 4095–4099. [Google Scholar]
- Sun, X.; Zhang, W.; Ramdas, L.; Stivers, D.N.; Jones, D.M.; Kantarjian, H.M.; Estey, E.H.; Vadhan-Raj, S.; Medeiros, L.J.; Bueso-Ramos, C.E. Comparative analysis of genes regulated in acute myelomonocytic leukemia with and without Inv (16)(p13q22) using microarray techniques, real-time PCR, immunohistochemistry, and flow cytometry immunophenotyping. Mod. Pathol. 2007, 20, 811–820. [Google Scholar]
- Larson, R.A.; Williams, S.F.; le Beau, M.M.; Bitter, M.A.; Vardiman, J.W.; Rowley, J.D. Acute myelomonocytic leukemia with abnormal eosinophils and inv(16) or t(16;16) has a favorable prognosis. Blood 1986, 68, 1242–1249. [Google Scholar]
- Grimwade, D.; Hills, R.K.; Moorman, A.V.; Walker, H.; Chatters, S.; Goldstone, A.H.; Wheatley, K.; Harrison, C.J.; Burnett, A.K. Refinement of cytogenetic classification in acute myeloid leukemia: Determination of prognostic significance of rare recurring chromosomal abnormalities among 5876 younger adult patients treated in the United Kingdom Medical Research Council Trials. Blood 2010, 116, 354–365. [Google Scholar] [CrossRef]
- Kanagal-Shamanna, R.; Bueso-Ramos, C.E.; Chen, S.S.; Abruzzo, L.V.; de Lima, M.J.; Sargent, R.L.; Medeiros, L.J.; Lu, G. Donor-derived isolated del (20q) after hematopoietic stem cell transplantation: Report of two cases and review of the literature. J. Hematopathol. 2011. [Google Scholar] [CrossRef]
- An International System for Human Cytogenetic Nomenclature: Recommendations of the International Standing Committee on Human Cytogenetic Nomenclature; Shaffer, L.G.; Slovak, M.L.; Campbell, L.J. (Eds.) Karger: Basel, Switzerland, 2009.
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar]
- Xu, M.; Li, D.; Lu, Y.; Chen, G.Q. Leukemogenic AML1-ETO fusion protein increases carcinogen-DNA adduct formation with upregulated expression of cytochrome P450-1A1 gene. Exp. Hematol. 2007, 35, 1249–1255. [Google Scholar] [CrossRef]
- McDonald, T.A.; Holland, N.T.; Skibola, C.; Duramad, P.; Smith, M.T. Hypothesis: Phenol and hydroquinone derived mainly from diet and gastrointestinal flora activity are causal factors in leukemia. Leukemia 2001, 15, 10–20. [Google Scholar]
- Smith, M.T. The mechanism of benzene-induced leukemia: A hypothesis and speculations on the causes of leukemia. Environ. Health Perspect. 1996, 104, 1219–1225. [Google Scholar]
- Smith, M.T. Benzene, NQO1, and genetic susceptibility to cancer. Proc. Natl. Acad. Sci. USA 1999, 96, 7624–7626. [Google Scholar] [CrossRef]
- Chattopadhyay, M.; Kodela, R.; Nath, N.; Street, C.R.; Velázquez-Martínez, C.A.; Boring, D.; Kashfi, K. Hydrogen sulfide-releasing aspirin modulates xenobiotic metabolizing enzymes in vitro and in vivo. Biochem. Pharmacol. 2012, 83, 733–740. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Talalay, P. NAD(P)H: Quinone acceptor oxidoreductase 1 (NQO1), a multifunctional antioxidant enzyme and exceptionally versatile cytoprotector. Arch. Biochem. Biophys. 2010, 501, 116–123. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kanagal-Shamanna, R.; Zhao, W.; Vadhan-Raj, S.; Nguyen, M.H.; Fernandez, M.H.; Medeiros, L.J.; Bueso-Ramos, C.E. Over-Expression of CYP2E1 mRNA and Protein: Implications of Xenobiotic Induced Damage in Patients with De Novo Acute Myeloid Leukemia with inv(16)(p13.1q22); CBFβ-MYH11. Int. J. Environ. Res. Public Health 2012, 9, 2788-2800. https://doi.org/10.3390/ijerph9082788
Kanagal-Shamanna R, Zhao W, Vadhan-Raj S, Nguyen MH, Fernandez MH, Medeiros LJ, Bueso-Ramos CE. Over-Expression of CYP2E1 mRNA and Protein: Implications of Xenobiotic Induced Damage in Patients with De Novo Acute Myeloid Leukemia with inv(16)(p13.1q22); CBFβ-MYH11. International Journal of Environmental Research and Public Health. 2012; 9(8):2788-2800. https://doi.org/10.3390/ijerph9082788
Chicago/Turabian StyleKanagal-Shamanna, Rashmi, Weiqiang Zhao, Saroj Vadhan-Raj, Martin H. Nguyen, Michael H. Fernandez, L. Jeffrey Medeiros, and Carlos E. Bueso-Ramos. 2012. "Over-Expression of CYP2E1 mRNA and Protein: Implications of Xenobiotic Induced Damage in Patients with De Novo Acute Myeloid Leukemia with inv(16)(p13.1q22); CBFβ-MYH11" International Journal of Environmental Research and Public Health 9, no. 8: 2788-2800. https://doi.org/10.3390/ijerph9082788