North American Wetlands and Mosquito Control

Abstract
:Acronyms
| BMP | Best Management Practices |
| Bti | Bacillus thuringensis var. israelensis |
| EEE | Eastern Equine Encephalitis |
| FIFRA | Federal Insecticide, Fungicide, and Rodenticide Act |
| FWS | Free Water Surface |
| USFWS | U.S. Fish and Wildlife Service |
| IGR | Insect Growth Regulator |
| IMM | Integrated Mosquito Management |
| IPN | Integrated Pest Management |
| Ls | Lysinibacillus sphaericus |
| MMF | Monomolecular Film |
| NOI | Notice of Intent |
| NPDES | National Pollutant Discharge Elimination System |
| OMWM | Open Marsh Water Management |
| PNW | Pacific Northwest |
| RIM | Rotational Impoundment Management |
| RRV | Ross River Virus |
| SLE | St. Louis Encephalitis |
| SSF | Sub-surface Flow |
| ULV | Ultra-low Volume |
| VEE | Venezuelan Encephalitis |
| WEE | Western Equine Encephalitis |
| WN | West Nile |
| WNV | West Nile Virus |
1. Introduction
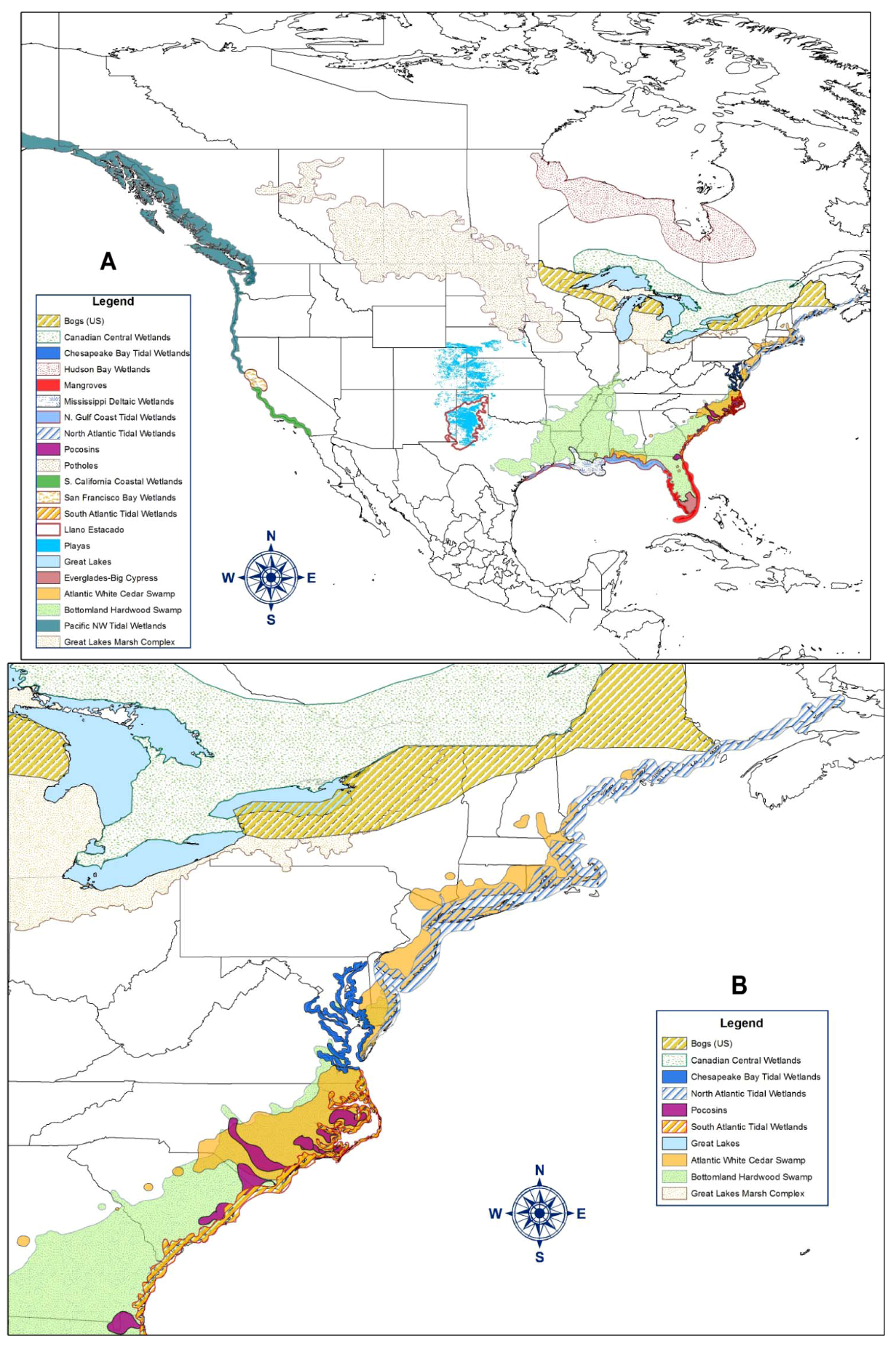
2. Types of Wetlands

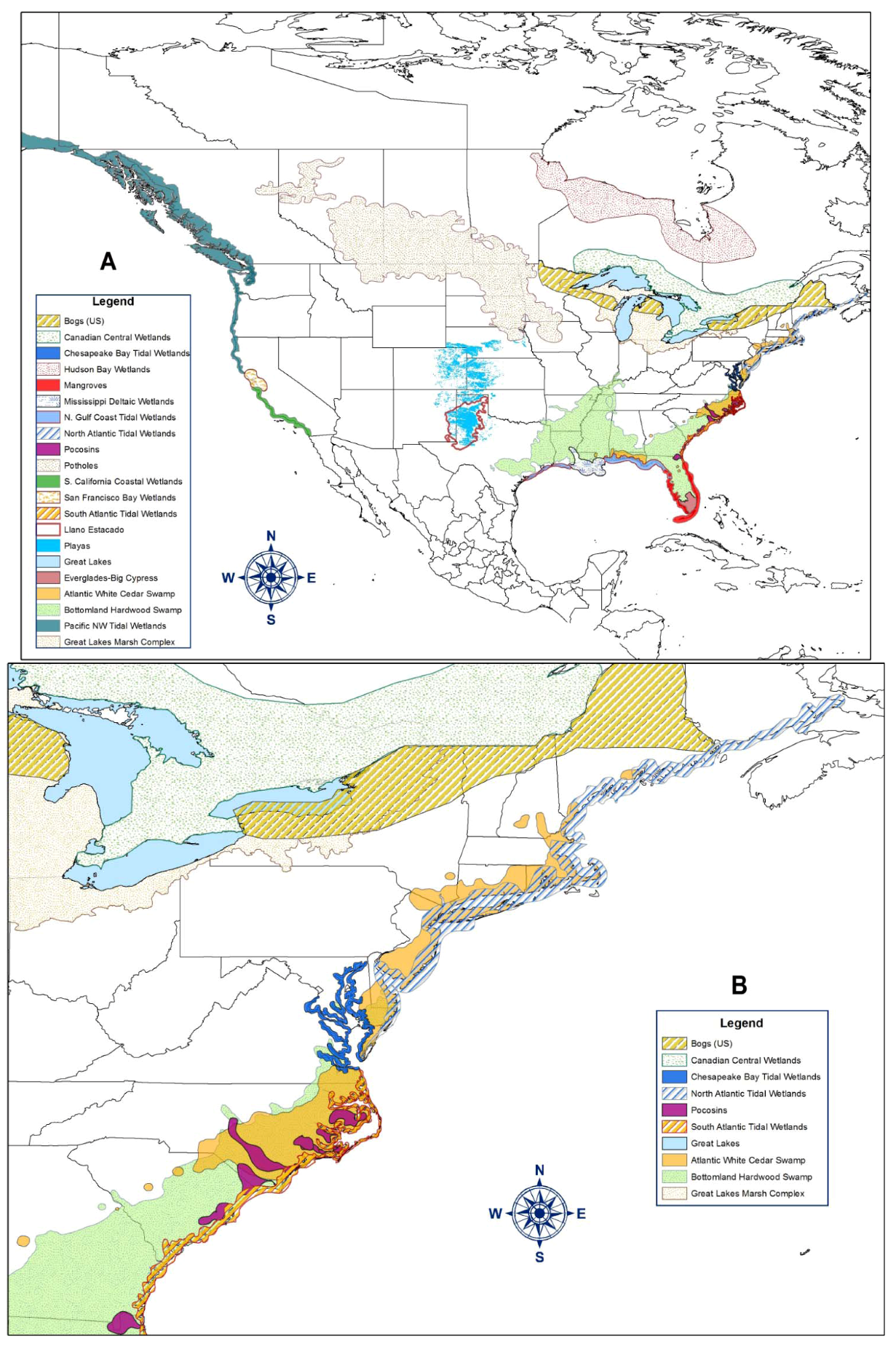{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | USFWS 1 classification | Major hydrologic influence | Flooding frequency |
|---|---|---|---|
| Mangrove | Estuarine forested/shrub | ocean tide | daily-seasonal |
| Tidal salt marsh | Estuarine intertidal emergent | ocean tide | daily-seasonal |
| Pacific Northwest tidal wetland | Estuarine intertidal emergent | ocean tide | daily-seasonal |
| Tidal brackish marsh | Estuarine intertidal emergent | tide/surface | daily-seasonal |
| Tidal freshwater wetland | Palustrine emergent | surface | daily-seasonal |
| Bottomland swamp | Palustrine emergent/ forested | river, precipitation, ground | semi-permanent |
| Atlantic white cedar wetland forest | Palustrine emergent/ forested | precipitation, ground | seasonal 2 |
| Riverine riparian floodplain wetland | Palustrine emergent/ forested | river, precipitation, | variable |
| Wet meadow | Palustrine emergent | ground | seasonal |
| Wet prairie | Palustrine emergent | ground | permanent, semi-permanent 2 |
| Playa | Palustrine emergent | surface | seasonal |
| Bog | Palustrine shrub | precipitation/runoff | variable/seasonal 2 |
| Pocosin | Palustrine shrub | ground | semi-permanent 2 |
| Fen | Palustrine shrub | ground | semi-permanent 2 |
| Carolina Bay | Palustrine shrub | precipitation/ground | permanent-seasonal |
| Pothole | Pond natural | precipitation | variable |
| Vernal pool | Pond natural | precipitation | seasonal |
| Mississippi deltaic plain wetlands | Mixed | various | variable |
| Everglades | Mixed | various | variable |
| Constructed wetlands | Mixed | - | permanent |
2.1. Coastal Wetlands
2.1.1. Mangroves
2.1.2. Tidal Salt Marshes
2.1.3. Pacific Northwest Tidal Wetlands
2.1.4. Tidal Brackish Marshes
2.1.5. Tidal Freshwater Wetlands
2.2. Freshwater (Non-Tidal) Wetlands
2.2.1. Wet Meadows
2.2.2. Wet Prairies
2.2.3. Potholes
2.2.4. Playas
2.2.5. Vernal Pools
2.2.6. Bottomland Swamps
2.2.7. Atlantic White Cedar Wetland Forests
2.2.8. Bogs and Pocosins (Including Carolina Bays)
2.2.9. Fens
2.2.10. Riverine, Riparian, Floodplain Wetlands
2.3. Wetland Complexes
2.3.1. The Florida Everglades
2.3.2. The Mississippi Deltaic Plain Wetlands (“Louisiana Wetlands”)
2.4. Constructed Wetlands
3. Mosquitoes
3.1. Mosquito Biology

3.2. Why Do Mosquitoes Bite?
3.3. Mosquitoes and Disease
| Mosquito Species | Habitat | Nuisance and/or Disease Associations | Associated Wetland Type | Range |
|---|---|---|---|---|
| Aedes abserratus | Freshwater cedar forests | Nuisance | Atlantic White Cedar wetland forest | North Atlantic Coast |
| Aedes atlanticus | Floodwater | Bottomland swamp | Southeastern US; Atlantic and Gulf Coasts | |
| Aedes atropalpus | Floodwater | Riverine | Northeastern US, Atlantic Coast | |
| Aedes aurifer | Freshwater cedar forests | Atlantic White Cedar wetland forest | North Atlantic Coast | |
| Aedes campestris | Floodwater | Potholes | Northwestern US | |
| Aedes canadensis | Floodwater: temporary shaded woodland pools and shaded pools adjacent to wooded areas | EEE, WN, dog heartworm | Vernal pools, Atlantic White Cedar wetland forests | Atlantic and Gulf Coasts, East of Rocky Mountains |
| Aedes cantator | Floodwater | Nuisance | Tidal salt marshes | Northern US |
| Aedes cinereus | Floodwater: freshwater cedar forests | WN, Nuisance | Atlantic White Cedar wetland forests, bogs and swamps | Atlantic Coast, Northern and Southeastern US |
| Aedes dorsalis | Floodwater: marshes and pools, overflow from wells | WEE, WN | Tidal salt marshes, coastal wetlands, potholes | Western US, Pacific Northwest, Northeastern US |
| Aedes excrucians | Floodwater: freshwater cedar forests | Nuisance | Vernal pools, Atlantic white cedar wetland forests | Atlantic and Gulf Coast, Northeastern and Northwestern US |
| Aedes infirmatus | Floodwater | EEE | Bottomland swamp, Mississippi deltaic plain | Gulf and South Atlantic Coasts, Southeastern US |
| Aedes flavescens | Floodwater | Nuisance | Potholes | Northeastern and Northwestern US |
| Aedes japonicus | Floodwater: containers, rock holes, shaded areas with water of high organic content | WN | Riverine | Eastern US |
| Aedes nigromaculis | Floodwater | Nuisance | Playas | Western US |
| Aedes sollicitans | Floodwater | EEE, nuisance | Mangrove, tidal salt marshes, tidal brackish | East and Gulf Coasts, Mississippi Deltaic Plain |
| Aedes squamiger | Floodwater | Nuisance | Tidal salt marshes, coastal wetlands | California, Pacific Northwest |
| Aedes sticticus | Floodwater | Nuisance, dog heartworm | Vernal pools | Eastern and Northwestern US |
| Aedes taeniorhynchus | Floodwater: salt marsh | Nuisance, dog heartworm | Mangrove, tidal salt marsh, tidal brackish, Florida Everglades, Mississippi deltaic plain | Atlantic, East and Gulf Coasts |
| Aedes triseriatus | Floodwater: treeholes, freshwater forests | Nuisance, EEE, WN, dog heartworm | Atlantic White Cedar wetland forest, Treeholes of deciduous trees | Eastern North America |
| Aedes trivitattus | Floodwater | Nuisance, dog heartworm | Meadows, swamps, woodlands | Eastern and Central US |
| Aedes vexans | Floodwater: freshwater cedar forests | Nuisance, EEE | Atlantic White Cedar wetland forests, Potholes, playas | Continental US |
| Anopheles atropos | Permanent water | Nuisance | Saltwater pools and marshes | Atlantic and Gulf Coasts |
| Anopheles crucians | Permanent water | Malaria | Bottomland swamp, Mississippi deltaic plain | Southeastern US; Gulf and Atlantic Coasts |
| Anopheles earlei | Semi-permanent and permanent water | Nuisance | Potholes, Bogs, marshes, woodland pools | Northern US |
| Anopheles quadrimaculatus and sibling species | Permanent Water: freshwater cypress swamps, river backwaters | Malaria | Bottomland swamp, Mississippi deltaic plain | Eastern US; Atlantic and Gulf Coasts |
| Coquillettidia perturbans | Permanent Water: Cattail ponds | EEE, nuisance | Tidal Freshwater (cattails), bottomland swamp, constructed wetlands, Mississippi, deltaic plain | Atlantic and Gulf Coasts, Eastern US; Northwestern US |
| Culex erraticus | Permanent Water: Freshwater cypress swamps | EEE, SLE, WN | Bottomland swamps | Eastern US; Atlantic and Gulf Coasts |
| Culex erythrothorax | Permanent water | WEE, WN | Constructed wetlands | West Coast |
| Culex nigripalpus | Permanent Water: ubiquitous in fresh water sources; sometimes found in brackish water | EEE, SLE, WN | Mangrove, bottomland swamp, Mississippi deltaic plain, constructed wetlands, Florida Everglades | Atlantic and Gulf Coasts |
| Culex pipiens | Permanent water, freshwater forests | WN | Atlantic White Cedar wetland forests, Constructed wetlands, playas | North Atlantic Coast, Northern US |
| Culex quinquefasciatus | Permanent water | SLE, WEE, WN, dog heartworm | Constructed wetlands, playas | Southern US |
| Culex restuans | Freshwater cedar forests | WN | Atlantic White Cedar wetland forests | North Atlantic coast |
| Culex salinarius | Permanent water | Nuisance, WN | Mangrove, tidal salt marshes, tidal brackish, Mississippi deltaic plain, constructed wetlands | Atlantic, Gulf and Pacific Coasts |
| Culex tarsalis | Permanent water | SLE, WEE, WN | Potholes, playas, constructed wetlands | Western and Southern US |
| Culex territans | Permanent water, freshwater forests | None for humans, feeds on cold-blooded vertebrates such as frogs | Atlantic White Cedar wetland forests, Potholes | North Atlantic Coast, Continental US |
| Culiseta inornata | Permanent water and temporarily flooded areas | WEE, WN | Tidal Salt Marshes (brackish upper marsh), Potholes | Atlantic and Gulf Coasts, Continental US |
| Culiseta melanura | Permanent Water: freshwater cypress swamps | EEE | Bottomland swamp; Atlantic White cedar wetland forests, Mississippi deltaic plain | Atlantic and Gulf Coasts; Eastern US |
| Culiseta morsitans | Freshwater cedar forests | EEE | Atlantic White Cedar wetland forests | North Atlantic Coast |
| Deinocerites cancer | Crab holes in tidal marshes | Nuisance | Tidal salt marshes | Eastern Coast of Florida |
| Deinocerites mathesoni | Crab holes | Nuisance | Tidal salt marshes | South Texas Coast |
| Deinocerites pseudes | Crab holes | Nuisance | Tidal salt marshes | South Texas Coast |
| Mansonia dyari | Permanent water | Nuisance | Constructed wetlands | Florida |
| Mansonia titillans | Permanent water | VEE, nuisance | Constructed wetlands | Florida to Texas |
| Psorophora ciliata | Floodwater | Nuisance | Rain-filled pools | Eastern US |
| Psorophora ferox | Floodwater | Nuisance | Woodlands, potholes | Eastern US |
| Psorophora signipennis | Floodwater | Nuisance | Playas | Central US |
| Uranotaenia lowii | Semi-permanent and permanent water; shallow margins of lakes | None for humans—Feeds on amphibians | Bottomland swamp | Southeastern US |
| Uranotaenia sapphirina | Freshwater Cedar forests, Permanent pools and ponds | None for humans | Atlantic White Cedar wetland forests, Bottomland swamp | North Atlantic Coast, Eastern US |
| Wyeomyia smithii | Pitcher plants | None for humans | Bogs, fens | Northeastern US |
| Wyeomyia mitchellae | Bromeliads | Nuisance | Florida Everglades | Southern Florida |
| Wyeomyia vanduzeei | Bromeliads | Nuisance | Florida Everglades | Southern Florida |
4. Mosquito Control
4.1. Strategy
4.2. Surveillance
4.3. Risk Assessment
4.4. Community Relations
4.5. Personal Protection
5. Transient Methods of Wetlands Mosquito Control
5.1. Chemical Control
5.2. Biological Control
5.3. Habitat Management
5.3.1. Water Management
5.3.2. Vegetation Control
5.3.3. Emergent Vegetation Control
5.3.4. Aquatic (Submerged and Floating) Vegetation Control
6. Long-Lasting Wetlands Modifications for Mosquito Control
6.1. Filling
6.2. Ditching
6.3. Runneling
6.4. Open Marsh Water Management (OMWM)

6.5. Impounding

6.6. Basin/Topography/Habitat Design
7. Integrated Mosquito Management (IMM)
8. Discussion
9. Concluding Remarks
Conflict of interest
Acknowledgements
References
- Mulrennan, J.A. Mosquito control—Its impact on the growth and development of Florida. J. Fl. Med. Assoc. 1986, 73, 310–311. [Google Scholar]
- Higgs, S. The 2005–2006 Chikungunya Epidemic in the Indian Ocean. Vector-borne Zoon. Dis. 2006, 6, 115–116. [Google Scholar] [CrossRef]
- Maartens, F; Sharp, B.; Curtis, B.; Mthembu, J.; Hatting, I. The impact of malaria control on perceptions of tourists and tourism operators concerning malaria prevalence in KwaZulu-Natal, 1999/2000 versus 2002/2003. Int. Soc. Travel Med. 2007, 14, 96–104. [Google Scholar] [CrossRef]
- Simmons, F.E.R.; Peng, Z. Skeeter syndrome. J. Allergy Clin. Immunol. 1999, 104, 705–707. [Google Scholar]
- Peng, Z.K.; Beckett, A.N.; Engler, R.J. Immune responses to mosquito saliva in 14 individuals with acute systemic allergic reactions to mosquito bites. J. Allergy Clin. Immunol. 2004, 144, 1189–1194. [Google Scholar]
- Rey, J.R.; Carlson, D.B.; Brockmeyer, R.E. Coastal wetland management in Florida: Environmental concerns and human health. Wetlands Ecol. Mgmt. 2012, 20, 197–211. [Google Scholar] [CrossRef]
- FDA (U.S. Food and Drug Administration). Protecting pets from mosquito-borne diseases. FDA Veterinarian 2002, 17, 1–3.
- Marra, P.P.; Griffing, S.; Caffrey, C.; Kilpatrick, A.M.; Mclean, R.; Brand, C.; Saito, E.; Dupuis, A.P.; Kramer, L.; Novak, R. West Nile virus and wildlife. BioScience 2004, 54, 393–402. [Google Scholar]
- Berg, J.A.; Felton, M.G.; Gecy, J.L.; Laderman, A.D.; Mayhew, C.R.; Mengler, J.L.; Meredith, W.H.; Read, N.R.; Rey, J.R.; Roberts, C.; Sakolsky, G.E.; Walton, W.E.; Wolfe, R.J. Mosquito control and wetlands. Wetland Sci. Pract. 2010, 27, 24–34. [Google Scholar]
- Meredith, W.H.; Saveikis, D.E.; Stachecki, C.J. Guidelines for “Open Marsh Water Management” in Delaware’s salt marshes—Objectives, system designs, and installation procedures. Wetlands 1985, 5, 119–133. [Google Scholar] [CrossRef]
- Carlson, D.B. Source reduction in Florida’s salt marshes: Management to reduce pesticide use and enhance the resource. J. Amer. Mosquito Control Assn. 2006, 22, 534–537. [Google Scholar] [CrossRef]
- Willott, E. Restoring nature, without mosquitoes? Restoration Ecol. 2004, 12, 147–153. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 3rd ed; John Wiley & Sons: New York, NY, USA, 2003. [Google Scholar]
- Ecology of Freshwater and Estuarine Wetlands; Batzer, D.P.; Sharitz, R.R. (Eds.) University of California Press: Berkeley, CA, USA, 2006.
- Coastal Wetlands: An Integrated Ecosystem Approach; Perillo, M.E.; Wolanski, E.; Cahoon, D.R.; Brinson, M.M. (Eds.) Elsevier: Oxford, UK, 2009.
- Wetland Habitats of North America: Ecology and Conservation Concerns; Batzer, D.P.; Baldwin, A.H. (Eds.) University of California Press: Berkeley, CA, USA, 2012.
- Wolanski, E.; Brinson, M.M.; Cahoon, D.R.; Perillo, G.M.E. A Synthesis of Coastal Wetlands Science. In Coastal Wetlands: An Integrated Ecosystem Approach; Perillo, M.E., Wolanski, E., Cahoon, D.R., Brinson, M.M., Eds.; Elsevier: Oxford, UK, 2009; pp. 2–42. [Google Scholar]
- Sharitz, R.R.; Pennings, S.C. Development of Wetland Plant Communities. In Ecology of Freshwater and Estuarine Wetlands; Batzer, D.P., Sharitz, R.R., Eds.; University of California Press: Berkeley, CA, USA, 2006; pp. 177–241. [Google Scholar]
- McKee, K.L. Neotropical Coastal Wetlands. In Wetland Habitats of North America: Ecology and Conservation Concerns; Batzer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 89–101. [Google Scholar]
- Provost, M.W. Mean high water mark and use of tidelands in Florida. Fla. Sci. 1973, 36, 50–66. [Google Scholar]
- Provost, M.W. Tidal datum planes circumscribing salt marshes. Bull. Mar. Sci. 1976, 26, 558–563. [Google Scholar]
- Carlson, D.B.; Vigliano, R.R. The effects of two different water management regimes on flooding and mosquito production in a salt marsh impoundment. J. Am. Mosquito Control Assoc. 1985, 1, 203–211. [Google Scholar]
- Bidlingmayer, W.L.; Schoof, H.F. The dispersal characteristics of the salt-marsh mosquito Aedes taeniorhynchus (Wiedemann) near Savannah, Georgia. Mosq. News 1957, 17, 202–212. [Google Scholar]
- Harden, F.W.; Chubb, H.S. Observations of Aedes taeniorhynchus dispersal in extreme South Florida and the Everglades National Park. Mosq. News 1960, 20, 249–255. [Google Scholar]
- Sudia, W.D.; Newhouse, V.F.; Beadle, I.D.; Miller, D.L.; Johnston, J.G., Jr.; Young, R.; Calisher, C.H.; Maness, K. Epidemic Venezuelan equine encephalitis in North America in 1971: Vector studies. Am. J. Epidemiol. 1975, 101, 17–35. [Google Scholar]
- Weaver, S.C.; Salas, R.; RicoHesse, R.; Ludwig, G.V.; Oberste, M.S.; Boshell, J.; Tesh, R.B. Re-emergence of epidemic Venezuelan equine encephalomyelitis in South America. Lancet 1996, 348, 436–440. [Google Scholar]
- Nayar, J.K.; Rosen, L.; Knight, J.W. Experimental vertical transmission of Saint Louis encephalitis-virus by Florida mosquitos. Amer. J. Trop. Med. Hyg. 1986, 35, 1296–1301. [Google Scholar]
- Turrell, M.J.; O’Guinn, M.L.; Dohm, D.J.; Jones, J.W. Vector competence of North American mosquitoes (Diptera: Culicidae) for West Nile virus. J. Med. Entomol. 2001, 38, 130–134. [Google Scholar] [CrossRef]
- Provost, M.W. Ecological control of salt marsh mosquitoes with side benefits to birds. Proc. Tall Timbers Conf. Ecol. Animal Control By Habitat Mgmt. 1969, 1968, 193–206. [Google Scholar]
- Kneib, R.T. The Role of Tidal Marshes in the Ecology of Estuarine Nekton. In Oceanography and Marine Biology: An Annual Review; Ansell, A.D., Gibson, R.N., Barnes, M., Eds.; UCL Press: Lodon, UK, 1997; pp. 163–220. [Google Scholar]
- Pennings, S.C.; Moore, D.J. Zonation of shrubs in Western Atlantic marshes. Oecologia 2001, 126, 587–594. [Google Scholar] [CrossRef]
- Zedler, J.B. The Ecology of Southern California Coastal Salt Marshes: A Community Profile; U.S. Fish and Wildlife Service: Washington, DC, USA, 1982. [Google Scholar]
- Wigland, C.; Roman, C.T. North Atlantic Coastal Tidal Wetlands. In Wetland Habitats of North America: Ecology and Conservation Concerns; Btazer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 13–28. [Google Scholar]
- Pennings, S.C.; Alber, M.; Clark, R.A.; Booth, M.; Burd, A.; Wei-Jun, C.; Craft, C.; Depratter, C.B.; Di Iorio, D.; Hopkinson, C.S.; Joye, S.B.; Meile, C.D.; Moore, W.S.; Silliman, B.; Thompson, V.; Wares, J.P. South Atlantic Tidal Wetlands. In Wetland Habitats of North America: Ecology and Conservation Concerns; Btazer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 45–61. [Google Scholar]
- Baldwin, A.H.; Batzer, D.P. Wetland Habitats of North America. In Wetland Habitats of North America: Ecology and Conservation Concerns; University of California Press: Berkeley, CA, USA, 2012; pp. 1–9. [Google Scholar]
- Kennish, M.J. Coastal salt marsh systems in the U.S.: A review of anthropogenic impacts. J. Coast. Res. 2001, 17, 731–748. [Google Scholar]
- Battaglia, L.L.; Woodrey, M.S.; Peterson, M.S.; Dillon, K.S.; Visser, J.M. Wetlands of the Northern Gulf Coast. In Wetland Habitats of North America: Ecology and Conservation Concerns; Batzer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 75–88. [Google Scholar]
- James-Pirri, M.; Ginsberg, H.; Erwin, R.; Taylor, J. Effects of open marsh water management on numbers of larval salt marsh mosquitoes. J. Med. Entomol. 2009, 46, 1392–1399. [Google Scholar] [CrossRef]
- MSMVCD (Marin Sonoma Mosquito and Vector Conrtrol District). The Salt Marsh Mosquito (Ochlerotatus squamiger). Available online: http://www.msmosquito.com/asquamig.html (accessed on 12 February 2012).
- Larsen, J. Characterizing Patterns of Wetland Occurrence in Oregon Using an Interactive Geodatabase: A Method for Conservation Planning.
- Brophy, L. Pacific Northwest Estuarine Wetlands 101. In Proceedings of the West Coast Symposium on the Effects of Tide Gates on Estuarine Habitats and Fishes, 31 October–2 November, Charleston, OR, USA; 2006. [Google Scholar]
- Callaway, J.C.; Borde, A.B.; Diefenderfer, H.L.; Parker, V.T.; Rybczyk, J.M.; Thom, R.M. Pacific Coast Tidal Wetlands. In Wetland Habitats of North America: Ecology and Conservation Concerns; Btazer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 103–116. [Google Scholar]
- NOAA, Estuaries of the United States: Vital Statistics of A National Resource Base; National Oceanographic and Atmospheric Administration, National Ocean Service: Rockville, MD, USA, 1990.
- Christy, J.A.; Putera, A.J. Lower Columbia River Natural Area Inventory; Columbia River Estuary Study Task Force: Astoria, OR, USA, 1993. [Google Scholar]
- Odum, W.E. National Coastal Ecosystems Team (U.S.). In The Ecology of Tidal Freshwater Marshes of The United States East Coast: A Community Profile; Fish and Wildlife Service, U.S. Department of the Interior: Washington, DC, USA, 1984. [Google Scholar]
- Hopkinson, C.S.; Gosselink, J.G.; Parrondo, R.T. Above ground production of seven marsh plant species in coastal Louisiana. Ecology 1978, 59, 760–769. [Google Scholar]
- Cooper, D.J.; Chimner, R.A.; Merritt, D.M. Western Mountain Wetlands. In Wetland Habitats of North America: Ecology and Conservation Concerns; Batzer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 313–328. [Google Scholar]
- U.S. Environmental Protection Agency. Wet Meadows. Available online: http://water.epa.gov/type/wetlands/wmeadows.cfm (accessed on 1 March 2012).
- NatureServe. NatureServe Explorer: An Online Encyclopedia Of Life. Available online: www.natureserve.org/explorer (accessed on 14 January 2010).
- United States Geological Survey. Ecosystems of South Florida. Available online: http://sofia.usgs.gov/publications/papers/pp1011/wetprairies.html (accessed on 29 March 2012).
- Loveless, C.M. A study of vegetation in the Florida Everglades. Ecology 1959, 40, 1–9. [Google Scholar] [CrossRef]
- Birnhak, B.I.; Crowder, J.P. An Evaluation of the Extent of Vegetative Habitat Alteration in South Florida 1943-1970. In South Florida Environmental Project: Ecological Report; U.S. Department of the Interior, Bureau of Sport Fisheries and Wildlife: Washington, DC, USA, 1974. [Google Scholar]
- Carter, M.R.; Burns, L.A.; Cavinder, T.R.; Dugger, K.R.; Fore, P.L.; Hicks, D.B.; Revells, H.L.; Schmidt, T.W. Ecosystems Analysis of the Big Cypress Swamp and Estuaries; US. Environmental Protection Agency: Washington, DC, USA, 1973. [Google Scholar]
- Weller, M.W. Freshwater Marshes: Ecology and Wildlife Management; University of Minnesota Press: Minneapolis, MN, USA, 1981. [Google Scholar]
- Galatowitsch, S. Northern Great Plains Wetlands. In Wetland Habitats of North America: Ecology and Conservation Concerns; Batzer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 283–298. [Google Scholar]
- Payne, N.F. Techniques for Wildlife Habitat Management of Wetlands; McGraw-Hill Publishers: New York, NY, USA, 1992. [Google Scholar]
- Meyer, M.I.; Swanson, G.A. Mosquitoes (Diptera: Culicidae) consumed by breeding Anatinae in south central North Dakota. Prairie Naturalist 1982, 14, 27–31. [Google Scholar]
- Parker, D.W. Emergence Phenologies and Patterns of Aquatic Insects Inhabiting a Prairie Pond.
- Smith, L.M. Playas of the Great Plains; University of Texas Press: Austin, TX, USA, 2003. [Google Scholar]
- Gurdak, J.J.; Roe, C.D. Recharge Rates and Chemistry Beneath Playas of the High Plains Aquifer—A Literature Review and Synthesis; U.S. Geological Survey Circular: Reston, VA, USA, 2009. [Google Scholar]
- Smith, L.M.; Haukos, D.A.; McMurry, S.T. High Plains Playas. In Wetland Habitats of North America: Ecology and Conservation Concerns; Batzer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 299–311. [Google Scholar]
- Haukos, D.A.; Smith, L.M. Plant Communities of Playa Wetlands in the Southern Great Plains; Museum of Texas Tech University: Lubbock, TX, USA, 2004. [Google Scholar]
- Mollhagen, T.R.; Urban, L.V.; Ramsey, R.H.; Wyatt, A.W.; McReynolds, C.D.; Ray, J.T. Assessment of Nonpoint-source Contamination of Playa Basins in the High Plains of Texas: Brazos Basin Watershed, Phase I; Texas Tech University, Water Resources Center: Lubbock, TX, USA, 1993. [Google Scholar]
- Venne, L.S.; Anderson, T.A.; Zhang, B.; Smith, L.M.; McMurry, S.T. Organochlorine pesticide concentrations in sediment and amphibian tissue in playa wetlands in the Southern High Plains, USA. Bull. Environ. Contam. Toxicol. 2008, 80, 497–501. [Google Scholar]
- Guerrant, G.O.; Fetzer, L.E., Jr.; Miles, J.W. Pesticide residues in Hale County, TX, before and after ultra-low volume aerial application of malathion. Pestic. Monit. J. 1970, 4, 14–20. [Google Scholar]
- McNew, R.M. Environmental Impact of Disease Vector Control.
- Owens, J.C.; Ward, C.R.; Huddleston, E.W.; Ashdown, D. Non-chemical methods of mosquito control for playa lakes in West Texas. Mosq. News 1970, 30, 571–579. [Google Scholar]
- Huddleston, E.W.; Riggs, V.C. Public Health Aspects of High Plains Water; Texas Water Development Board-Texas Tech University: Lubbock, TX, USA, 1965. [Google Scholar]
- Tiner, R.W.; Bergquist, H.C.; DeAlessio, G.P.; Starr, M.J. Geographically isolated wetlands of the United States. Wetlands 2003, 23, 494–516. [Google Scholar] [CrossRef]
- Colburn, E.A. Vernal Pools: Natural History and Conservation; McDonald and Woodward Publishing: Blacksburg, VA, USA, 2004. [Google Scholar]
- Lester, G.D. Louisiana Comprehensive Wildlife Conservation Strategy; Louisiana Department of Wildlife and Fisheries: Baton Rouge, LA, USA, 2005. [Google Scholar]
- Conner, W.H.; Buford, M.N.A. Southern Deepwater Swamps. In Southern Forested Wetlands: Ecology and Management; Messina, M.G., Conner, W.H., Eds.; Lewis Publishers: Boca Raton, FL, USA, 1998; pp. 261–290. [Google Scholar]
- King, S.L.; Battaglia, L.L.; Hupp, C.R.; Keim, R.F.; Lockaby, B.G. Floodplain Wetlands of the Southeastern Coastal Plain. In Wetland Habitats of North America: Ecology and Conservation Concerns; Batzer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 253–266. [Google Scholar]
- U.S. Environmental Protection Agency. Bottomland Hardwoods. Available online: http://water.epa.gov/type/wetlands/bottomland.cfm (accessed on 9 October 2011).
- Davis, H. Mosquito Populations and Arbovirus Activity in Cypress Domes. In Cypress Swamps; Ewel, C.C., Odum, H.T., Eds.; University of Florida Press: Gainesville, FL, USA, 1984. [Google Scholar]
- Laderman, A.D. The Ecology of the Atlantic White Cedar Wetlands: A Community Profile; U.S. Fish and Wildlife Service: Washington, DC, USA, 1989. [Google Scholar]
- Gorham, E. The Ecology and Biogeochemistry of Sphagnum Bogs in Central and Eastern North America. In Atlantic White Cedar Wetlands; Laderman, A.D., Ed.; Westview Press: Boulder, CO, USA, 1987; pp. 1–15. [Google Scholar]
- New Jersey Pinelands Commission, Comprehensive Management Plan for the Pinelands National Reserve and Pinelands Area; New Jersey Pinelands Commission: New Lisbon, NJ, USA, 1980.
- Taylor, N. Flora in the vicinity of New York: A contribution to plant geography. Mem. N. Y. Bot. Garden. 1916, 5, 1883–1967. [Google Scholar]
- Sakolsky, G.E.; Laderman, A. Mosquito Sampling Updated; Cedar Swamps of Cape Cod Massachusetts. In Proceedings of the Ecology and Management of White Cedar Ecosystems Symposium, Greenville, NC, USA, 9–11 June 2009.
- Carpenter, S.J.; LaCasse, W.J. Mosquitoes of North America (North of Mexico); University of California Press: Berkeley, CA, USA, 1955. [Google Scholar]
- Molaei, G.; Oliver, J.; Andreatis, T.G.; Armstrong, P.M.; Howard, J.J. Molecular identification of blood-meal sources in Culiseta melanura and Culiseta morsitans from an endemic focus of Eastern Equine Encephalitis virus in New York. Amer. J. Trop. Med. Hyg. 2006, 75, 1140–1147. [Google Scholar]
- Rochefort, L.; Strack, M.; Poulin, M.; Price, J.S.; Graf, M.; Desrochers, A.; Lavoie, C. Northern Peatlands. In Wetland Habitats of North America: Ecology and Conservation Concerns; Batzer, D.P., Baldwin, A.H., Eds.; University of California Press: Berkeley, CA, USA, 2012; pp. 119–134. [Google Scholar]
- Bedford, B.L.; Godwin, K.S. Fens of the United States: Distribution, characteristics, and scientific connection versus legal isolation. Wetlands 2003, 23, 608–629. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Fens. Available online: http://water.epa.gov/type/fens/wmeadows.cfm (accessed on 6 February 2012).
- Means, R.G. The Mosquitoes of New York; New York State Museum: New York, NY, USA, 1987. [Google Scholar]
- Kline, D.L.; Wood, J.R.; Cornell, J.A. Interactive effects of 1-octen-3-Ol and carbon dioxide on mosquito (Diptera, Culicidae) surveillance and control. J. Med. Entomol. 1991, 28, 254–258. [Google Scholar]
- LeBlanc, R. The Geological History of the Marshes of Coastal Louisiana. In Marsh Management in Coastal Louisiana: Effects And Issues; Duffy, W.G., Clark, D., Eds.; U.S. Fish Wildl. Serv. Biol. Rep: Washington, DC, USA, 1989. [Google Scholar]
- Gosselink, J.G. The Ecology of Delta Marshes of Coastal Louisiana: A Community Profile; U.S. Fish and Wildlife Service: Washington, DC, USA, 1984. [Google Scholar]
- Day, J.W., Jr.; Boesch, D.F.; Clairain, E.J.; Kemp, G.P.; Laska, S.B.; Mitsch, W.J.; Orth, K.; Mashriqui, H.; Reed, D.J.; Shabman, L.; Simenstad, C.A.; Streever, B.J.; Twilley, R.R.; Watson, C.C.; Wells, J.T.; Whigham, D.F. Restoration of the Mississippi Delta: Lessons from hurricanes Katrina and Rita. Science 2007, 315, 1679–1684. [Google Scholar]
- Barras, J.A. Land Area Change in Coastal Louisiana after the 2005 Hurricanes—A Series Of Three Maps: U.S. Geological Survey Open-File Report 2006-1274. Available online: http://pubs.usgs.gov/of/2006/1274/ (accessed on 14 February 2012).
- Turner, R.E.; Baustian, J.J.; Swenson, E.M.; Spicer, J.S. Wetland sedimentation from hurricanes Katrina and Rita. Science 2006, 314, 449–452. [Google Scholar]
- Kadlec, R.H.; Wallace, S.D. Treatment Wetlands, 2nd ed; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Crites, R.W.; Middlebrooks, R.J.; Reed, S.C. Natural Wastewater Treatment Systems; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Rey, J.R.; O’Meara, G.F.; O’Connell, S.M.; Cutwa-Francis, M.M. Mosquito production from four constructed treatment wetlands in peninsular Florida. J. Amer. Mosquito Control Assn. 2006, 22, 198–205. [Google Scholar]
- Vymazal, J.; Kröpfelová, L. Wastewater Treatment in Constructed Wetlands with Horizontal Sub-Surface Flow (Environmental Pollution); Springer: Berlin, Germany, 2008. [Google Scholar]
- Florida Coordinating Council on Mosquito Control, Florida Mosquito Control: The State of The Mission as Defined by Mosquito Controllers, Regulators, and Environmental Managers; Connelly, C.R.; Carlson, D.B. (Eds.) University of Florida, IFAS: Vero Beach, FL, USA, 2009.
- Carlson, D.B.; Knight, R.L. Mosquito production and hydrological capacity of southeast Florida impoundments used for wastewater retention. J. Amer. Mosquito Control Assn. 1987, 3, 74–83. [Google Scholar]
- Walton, W.E.; Workman, P.D. Effect of marsh design on the abundance of mosquitoes in experimental constructed wetlands in southern California. J. Amer. Mosquito Control Assn. 1998, 14, 95–107. [Google Scholar]
- Keiper, J.B.; Jiannino, J.A.; Sanford, M.R.; Walton, W.E. Effect of vegetation management on the abundance of mosquitoes at a constructed treatment wetland in southern California. Proc. Papers Mosq. Vector Control Assoc. Calif. 2003, 70, 35–43. [Google Scholar]
- Walton, W.E. Managing Mosquitoes in Surface-Flow Constructed Treatment Wetlands; University of California: Davis, CA, USA, 2003. [Google Scholar]
- Knight, R.L.; Walton, W.E.; O’Meara, G.F.; Reisen, W.K.; Wass, R. Strategies for effective mosquito control in constructed treatment wetlands. Ecol. Eng. 2003, 21, 211–232. [Google Scholar]
- Walton, W.E. Multipurpose Constructed Treatment Wetlands in the Arid Southwestern United States: Are the Benefits Worth the Risks? In Treatment Wetlands for Water Quality Improvement: Quebec 2000 Conference Proceedings (Selected Papers); Pries, J., Ed.; CH2M Hill Canada Limited, Pandora Press: Waterloo, ON, Canada, 2002; pp. 115–123. [Google Scholar]
- Foster, W.A. Mosquito sugar feeding and reproductive energetics. Ann. Rev. Entomol. 1995, 40, 443–474. [Google Scholar] [CrossRef]
- Spielman, A. Bionomics of autogenous mosquitoes. Ann. Rev. Entomol. 1971, 16, 231–248. [Google Scholar] [CrossRef]
- Klowden, M. J. Blood, sex, and the mosquito. BioScience 1995, 45, 326–331. [Google Scholar] [CrossRef]
- Savage, H.M.; Niebylski, M.L.; Smith, G.C.; Mitchell, C.J.; Craig, G.B., Jr. Host-feeding patterns of Aedes albopictus (Diptera: Culicidae) at a temperate North American site. J. Med. Entomol. 1993, 30, 27–34. [Google Scholar]
- Moore, C.G.; McLean, R.G.; Mitchell, C.J.; Nasci, R.S.; Tsai, T.F.; Calisher, C.H.; Marfin, A.A.; Moore, P.S.; Gubler, D.J. Guidelines for Arbovirus Surveillance Programs in the United States; U.S. Department of Health and Human Services: Fort Collins, CO, USA, 1993. [Google Scholar]
- Rutledge, C.R. Surveillance for Mosquito-Borne Viruses. Available online: http://edis.ifas.ufl.edu/in479 (accessed on 23 October 2011).
- Moore, D. Integrated Mosquito Surveillance and Environmental Monitoring to Asses Changes in Mosquito Populations. In Florida Coordinating Council on Mosquito Control. Florida Mosquito Control: The State of The Mission as Defined By Mosquito Controllers, Regulators, and Environmental Managers; Connelly, C.R., Carlson, D.B., Eds.; University of Florida: Vero Beach, FL, USA, 2009; pp. 28–37. [Google Scholar]
- Boyce, K.W.; Brown, D.A. Integrated vector management guidelines for adult mosquitoes. J. Amer. Mosquito Control Assn. 2003, 19, 448–451. [Google Scholar]
- Service, M.W. Mosquito Ecology: Field Sampling Methods, 2nd ed; Elsevier and Chapman and Hall: London, UK, 1993. [Google Scholar]
- Eisen, L.; Eisen, R. Using geographic information systems and decision support systems for the prediction, prevention, and control of vector-borne diseases. Ann. Rev. Entomol. 2011, 56, 41–61. [Google Scholar] [CrossRef]
- Callahan, M.A.; Sexton, K. Is cumulative risk assessment is the answer, what is the question? Env. Health Perspect. 2007, 115, 799–806. [Google Scholar] [CrossRef]
- Committee on Risk Assessment of Hazardous Air PollutantsBoard on Environmental Studies and ToxicologyCommission on Life SciencesNational Research CouncilScience and Judgement in Risk Assessment; National Academy Press: Washington, DC, USA, 1994.
- Dale, P.E.R.; Hulsman, K. A critical-review of salt-marsh management methods for mosquito-control. Rev. Aquatic Sci. 1990, 3, 281–311. [Google Scholar]
- Hoffsteter, P.; Bare, J.C.; Hammitt, J.K.; Murphy, P.A.; Rice, G.E. Tools for comparative analysis of alternatives: Competing or complementary perspectives? Risk Anal. 2002, 22, 833–851. [Google Scholar]
- California Department of Public Health, California Mosquito-Borne Virus Surveillance and Response Plan; California Department of Public Health, Mosquito and Vector Control Association of California and the University of California: Sacramento, CA, USA, 2009.
- Mosquito-Borne Virus Surveillance and Emergency Response Plan; Coachella Valley Mosquito and Vector Control District: Indio, CA, USA, 2010.
- Integrated Vector Management and Response Plan; Orange County Vector Control District: Garden Grove, CA, USA, 2010.
- Rutledge, C.R.; Day, J.F.; Lord, C.C.; Stark, L.M.; Tabachnick, W.J. West Nile virus infection rates in Culex nigripalpus (Diptera: Culicidae) do not reflect transmission rates in Florida. J. Med. Entomol. 2003, 40, 253–258. [Google Scholar] [CrossRef]
- Vitek, C.J.; Richards, S.L.; Mores, C.N.; Day, J.F.; Lord, C.C. Arbovirus transmission by Culex nigripalpus in Florida, 2005. J. Med. Entomol. 2008, 45, 483–493. [Google Scholar]
- Bustamante, D.M.; Lord, C.C. Sources of error in the estimation of mosquito infection rates used to assess risk of arbovirus transmission. Amer. J. Trop. Med. Hyg. 2010, 82, 1171–1184. [Google Scholar]
- Villari, P.; Spielman, A.; Komar, N.; McDowell, M.; Timperi, R.J. The economic burden imposed by a residual case of Eastern encephalitis. Amer. J. Trop. Med. Hyg. 1995, 52, 8–13. [Google Scholar]
- Utz, J.T.; Apperson, C.S.; MacCormack, J.N.; Salyers, M.; Dietz, E.J.; McPherson, J.T. Economic and social impacts of La Crosse encephalitis in western North Carolina. Amer. J. Trop. Med. Hyg. 2003, 69, 509–518. [Google Scholar]
- Zohrabian, A.; Meltzer, M.I.; Ratard, R.; Billah, K.; Molinari, N.A.; Roy, K.; Scott II, R.D.; Petersen, L.R. West Nile virus economic impact, Louisiana, 2002. Emerg. Infect. Diseases 2004, 10, 1736–1744. [Google Scholar] [CrossRef]
- Barber, L.M.; Schleier, J.J.; Peterson, R.K.D. Economic cost analysis of West Nile virus outbreak, Sacramento County, California, USA, 2005. Emerg. Infect. Diseases 2010, 16, 480–486. [Google Scholar] [CrossRef]
- Beams, B.F. Analysis of mosquito control agency public education programs in the United-States. J. Amer. Mosquito Control Assn. 1985, 1, 212–219. [Google Scholar]
- Swaddiwudhipong, W.; Chaovakiratipong, C.; Nguntra, P.; Koonchote, S.; Khumklam, P.; Lerdlukanavonge, P. Effect of health education on community participation in control of dengue hemorrhagic fever in an urban area of Thailand. SoutheastAsian J. Trop. Med. Public Health 1992, 23, 200–206. [Google Scholar]
- Kumar, R.; Hwang, J.S. Larvicidal efficiency of aquatic predators: A perspective for mosquito biocontrol. Zool. Studies 2006, 45, 447–466. [Google Scholar]
- Mani, T.R.; Arunachalam, N.; Rajendran, R.; Satyanarayana, K.; Dash, A.P. Efficacy of thermal fog application of deltacide, a synergized mixture of pyrethroids, against Aedes aegypti, the vector of dengue. Trop. Med. Int. Health. 2005, 10, 1298–1304. [Google Scholar] [CrossRef]
- Jensen, T.; Lawler, S.P.; Dritz, D.A. Effects of ultra-low volume pyrethrin, malathion, and permethrin on nontarget invertebrates, sentinel mosquitoes, and mosquitofish in seasonally impounded wetlands. J. Amer. Mosquito Control Assn. 1999, 15, 330–338. [Google Scholar]
- Lawler, S.P.; Jensen, T.; Dritz, D.A.; Wichterman, G. Field efficacy and nontarget effects of the mosquito Larvicides temephos, methoprene, and Bacillus thuringiensis var. israelensis in Florida mangrove swamps. J. Amer. Mosquito Control Assn. 1999, 15, 446–452. [Google Scholar]
- Petterson, E.L. Temporal pattern of mosquito flight activity. Behavior 1980, 72, 1–25. [Google Scholar]
- Nasci, R.S.; Edman, J.D. Vertical and temporal flight activity of the mosquito Culisetamelanura (Diptera, Culicidae) in southeastern Massachusetts. J. Med. Entomol. 1981, 18, 501–504. [Google Scholar]
- Williams, C.R.; Kokkinn, M.J. Daily patterns of locomotor and sugar-feeding activity of the mosquito Culex annulirostris from geographically isolated populations. Physiolog. Entomol. 2005, 30, 309–316. [Google Scholar] [CrossRef]
- Haufe, W. Synoptic correlation of weather with mosquito activity. Biometeorology 1967, 2, 523–540. [Google Scholar]
- Service, M. Effects of wind on the behaviour and distribution of mosquitoes and blackflies. Int. J. Biometeorol. 1980, 24, 347–353. [Google Scholar] [CrossRef]
- Freire, M.G.; Schweigmann, N.J. Effect of temperature on the flight activity of culicids in Buenos Aires City, Argentina. J. Nat. Hist. 2009, 43, 2167–2177. [Google Scholar]
- Roiz, D.; Rosa, R.; Arnoldi, D.; Rizzoli, A. Effects of temperature and rainfall on the activity and dynamics of host-seeking Aedes albopictus females in Northern Italy. Vector-BorneZoonot. Dis. 2010, 10, 811–816. [Google Scholar] [CrossRef]
- Marten, G.G. The potential of mosquito-indigestible phytoplankton for mosquito control. J. Amer. Mosquito Control Assn. 1987, 3, 105–106. [Google Scholar]
- Rey, J.R.; Hargraves, P.E.; O’Connell, S.M. Effect of selected marine and freshwater microalgae on development and survival of the mosquito Aedes Aegypti. Aquatic Ecology 2009, 43, 987–997. [Google Scholar] [CrossRef]
- Kerwin, J.L.; Washino, R.K. Field evaluation of Lagenidium giganteum (Oömycetes: Langenidiales) and description of a natural epizootic involving a new isolate of the fungus. J. Med. Entomol. 1988, 25, 452–460. [Google Scholar]
- Kerwin, J.L. Oomycetes, Lagenidium giganteum. J. Amer. Mosquito Control Assn. 2007, 23, 50–57. [Google Scholar] [CrossRef]
- Federici, B.A. Viral pathogens of mosquito larvae. Bull. Amer. Mosquito Control Assoc. 1985, 6, 62–74. [Google Scholar]
- Federici, B.A.; Park, H.W.; Bideshi, D.K.; Wirth, M.C.; Johnson, J.J. Recombinant bacteria for mosquito control. J. Exp. Biol. 2003, 206, 3877–3885. [Google Scholar]
- Becnel, J.J.; White, S.E.; Shapiro, A.M. Review of microsporidia-mosquito relationships: From the simple to the complex. Folia Parasitol. 2005, 52, 41–50. [Google Scholar]
- Tseng, M. Ascogregarine parasites as possible biocontrol agents of mosquitoes. J. Amer. Mosquito Control Assn. 2007, 23, 30–34. [Google Scholar] [CrossRef]
- Perez-Pacheco, R.; Santamarina-Mijares, A.; Vasquez-Lopez, A.; Martinez-Tomas, S.H.; Suarez-Espinosa, J. Effectiveness and survival of Romanomermis culicivorax in natural breeding sites of mosquito larvae. Agrociencia 2009, 43, 861–868. [Google Scholar]
- Collins, F.H.; Washino, R.K. Insect predators. Bull. Amer. Mosquito Control. Assoc. 1985, 6, 25–41. [Google Scholar]
- Mogi, M. Insects and other invertebrate predators. J. Amer. Mosquito Control Assn. 2007, 23, 93–109. [Google Scholar] [CrossRef]
- Collins, L.E.; Blackwell, A. The biology of Toxorhynchites mosquitoes and their potential as biocontrol agents. Biocontrol 2000, 21, 105N–116N. [Google Scholar]
- Focks, D.A. Toxorhynchites as biocontrol agents. J. Amer. Mosquito Control Assn. 2007, 23, 118–128. [Google Scholar] [CrossRef]
- Marten, G.G.; Bordes, E.S.; Nguyen, M. Use of cyclopoid copepods for mosquito control. Hydrobiologia 1994, 293, 491–496. [Google Scholar]
- Rey, J.R.; O’Connell, S.; Suarez, S.; Menendez, Z.; Lounibos, L.P.; Byer, G. Laboratory and field studies of Macrocyclops albidus (Crustacea: Copepoda) for biological control of mosquitoes in artificial containers in a subtropical environment. J. Vector Ecol. 2004, 29, 124–134. [Google Scholar]
- Walton, W.E.; Workman, P.D.; Pucko, S. Efficacy of larvivorous fish against Culex spp. in experimental wetlands. Proc Mosq. Vector. Control Assoc. Calif. 1996, 64, 96–101. [Google Scholar]
- Walton, W.E. Larvivorous fish including Gambusia. J. Amer. Mosquito Contr. Assn. 2007, 23, 184–220. [Google Scholar] [CrossRef]
- Chapman, H.C. Biological Control of Mosquitoes; American Mosquito Control Association(AMCA): Mount Laurel, NJ, USA, 1985; Bull No. 6. [Google Scholar]
- Floore, T. Biorational Control of Mosquitoes; American Mosquito Control Association (AMCA): Mount Laurel, NJ, USA, 2007; Bull No 7. [Google Scholar]
- Rey, J.R. Biological and Alternative Control. In Florida Mosquito Control: The State of The Mission as Defined by Mosquito Controllers, Regulators, and Environmental Managers; Connelly, C.R., Carlson, D.B., Eds.; University of Florida: Vero Beach, Fl, USA, 2009; pp. 123–130. [Google Scholar]
- Schleier, J.; Sing, S.; Peterson, R. Regional ecological risk assessment for the introduction of Gambusia affinis (western mosquitofish) into Montana watersheds. Biol. Invasions 2008, 10, 1277–1287. [Google Scholar] [CrossRef]
- Walton, W.E.; Mulla, M.S. Impacts and Fates of Microbial Pest Control Agents in the Aquatic Environment. In Dispersal of Living Organisms into Aquatic Ecosystems; Rosenfield, A., Mann, R., Eds.; University of Maryland Sea Grant College: College Park, MD, USA, 1992; pp. 205–237. [Google Scholar]
- Carney, R.M.; Husted, S.; Jean, C.; Glaser, C.; Kramer, V. Efficacy of aerial spraying of mosquito adulticide in reducing incidence of West Nile virus, California, 2005. Emerg. Infect. Dis. 2008, 14, 747–754. [Google Scholar] [CrossRef]
- Elnaiem, D.; Kelley, K.; Wright, S.; Laffey, R.; Yoshimura, G.; Reed, M.; Goodman, G.; Thiemann, T.; Reimer, L.; Reisen, W.; Brown, D. Impact of aerial spraying of pyrethrin insecticide on Culex pipiens and Culex tarsalis (Diptera: Culicidae) abundance and West Nile virus infection rates in an urban/suburban area of Sacramento County, California. J. Med. Entomol. 2008, 45, 751–757. [Google Scholar] [CrossRef]
- Macedo, P.A.; Schleier, J.J.; Reed, M.; Kelley, K.; Goodman, G.W.; Brown, D.A.; Peterson, R.K.D. Evaluation of efficacy and human health risk of aerial ultra-low volume applications of pyrethrins and piperonyl butoxide for adult mosquito management in response to West Nile virus activity in Sacramento County, California. J. Amer. Mosquito Contr. Assn.. 2010, 26, 57–66. [Google Scholar]
- Juliano, S.A. Population dynamics. J. Amer. Mosquito Contr. Assn. 2007, 23, 265–275. [Google Scholar]
- Pike, G.H. Plague minnow or mosquito fish? A review of the biology and impacts of introduced Gambusia species. Ann. Rev. Ecol. Evol. Syst. 2008, 39, 171–191. [Google Scholar] [CrossRef]
- Meffe, G.K.; Snelson, F.F. An Ecological Overview of Poeciliid Fishes. In Ecology and Evolution of Livebearing Fishes (Poeciliidae); Meffe, G.K., Snelson, F.F., Eds.; Prentice Hall: Englewood Cliffs, NJ, USA, 1989. [Google Scholar]
- Gratz, N.S.; Legner, E.F.; Meffe, G.K.; Bay, E.C.; Service, M.W.; Swanson, Jr., C.; Cech, J.J.; Laird, M. Comments on adverse assessments of Gambusia affinis. J. Amer. Mosquito Contr. Assn. 1996, 12, 752–752. [Google Scholar]
- Rupp, H.R. Adverse assessments of Gambusia affinis: An alternate view for mosquito control practitioners. J. Amer. Mosquito Contr. Assn. 1996, 12, 155–159. [Google Scholar]
- Gerberich, J.; Laird, M. Biocontrol and Other Innovative Components, and Future Directions. In Integrated Mosquito Control Methodologies Volume 2; Laird, M., Miles, J., Eds.; Academic Press: London, UK, 1985. [Google Scholar]
- Ahmed, S.S.; Linden, A.L.; Cech, J.J. A rating system and annotated bibliography for the selection of appropriate indigenous fish species for mosquito and weed control. Bull. Soc. Vector Ecol. 1988, 13, 1–59. [Google Scholar]
- Sakolsky-Hoopes, G.; Doane, J.W. Preliminary evaluation of the use of native banded sunfish to control the mosquito vector of Eastern Equine Encephalitis. Environment Cape Cod 1998, 1, 41–47. [Google Scholar]
- Daniels, C.W. Reports of the Malaria Commission of the Royal Society; Harrison: London, UK, 1901; pp. 28–33, Series 5. [Google Scholar]
- Riviére, F.; Thirel, R. The predation of Aedes (Stegomyia) aegypti and Ae. (St) polynesiensis larvae [Dip, Culicidae] by the copepod Mesocyclops leuckarti-pilosa [Crustacea]—Preliminary experiments as biological-control agent. Entomophaga 1981, 26, 427–439. [Google Scholar] [CrossRef]
- Nam, V.S.; Yen, N.T.; Kay, B.H.; Marten, G.G.; Reid, J.W. Eradication of Aedes aegypti from a village in Vietnam, using copepods and community participation. Amer. J. Trop. Med. Hyg. 1998, 59, 657–660. [Google Scholar]
- Kay, B.; Nam, V.S. New strategy against Aedes aegypti in Vietnam. Lancet 2005, 365, 613–617. [Google Scholar]
- Marten, G.G. Elimination of Aedes albopictus from tire piles by introducing Macrocyclops albidus (Copepoda, Cyclopidae). J. Amer. Mosquito Contr. Assn. 1990, 6, 689–693. [Google Scholar]
- Marten, G.G.; Nguyen, M.; Ngo, G. Copepod predation on Anopheles quadrimaculatus larvae in rice fields. J. Vector Ecol. 2000, 25, 1–6. [Google Scholar]
- Marten, G.G.; Reid, J.W. Cyclopoid copepods. J. Amer. Mosquito Contr. Assn. 2007, 23, 65–92. [Google Scholar] [CrossRef]
- Lizárraga-Partida, M.L.; Mendez-Gomez, E.; Rivas-Montano, A.M.; Vargas-Hernandez, E.; Portillo-Lopez, A.; Gonzalez-Ramirez, A.R.; Huq, A.; Colwell, R.R. Association of Vibrio cholerae with plankton in coastal areas of Mexico. Env. Microbiol. 2009, 11, 201–208. [Google Scholar] [CrossRef]
- Signoretto, C.; Burlacchini, G.; Pruzzo, C.; Canepari, P. Persistence of Enterococcus faecalis in aquatic environments via surface interactions with copepods. Appl. Environ. Microbiol. 2005, 71, 2756–2761. [Google Scholar] [CrossRef]
- Colwell, R.R.; Huq, A.; Islam, M.S.; Aziz, K.M.A.; Yunus, M.; Khan, N.H.; Mahmud, A.; Sack, R.B.; Nair, G.B.; Chakraborty, J.; Sack, D.A.; Russek-Cohen, E. Reduction of cholera in Bangladeshi villages by simple filtration. Proc. Natl. Acad. Sci. USA 2003, 100, 1051–1055. [Google Scholar]
- O’Meara, G.F.; Cutwa-Francis, M.; Rey, J.R. Seasonal variation in the abundance of Culex nigripalpus and Culex quinquefasciatus in wastewater ponds at two Florida dairies. J. Amer. Mosquito Contr. Assn. 2010, 26, 160–166. [Google Scholar] [CrossRef]
- Magenheimer, J.F.; Moore, T.R.; Chmura, G.L.; Daoust, R.J. Methane and carbon dioxide flux from a macrotidal salt marsh, Bay of Fundy, New Brunswick. Estuaries 1996, 19, 139–145. [Google Scholar] [CrossRef]
- Batzer, D.P.; Sharitz, R.R. Ecology of Freshwater and Estuarine Wetlands; University of California Press: Berkeley, CA, USA, 2006. [Google Scholar]
- Gedan, K.; Silliman, B.; Bertness, M. Centuries of human-driven change in salt marsh ecosystems. Ann. Rev. Mar. Sci. 2009, 1, 17–141. [Google Scholar]
- Orr, B.K.; Resh, V.H. Experimental test of the influence of aquatic macrophyte cover on the survival of Anopheles larvae. J. Amer. Mosquito Contr. Assn. 1989, 5, 579–585. [Google Scholar]
- Clements, A.N. The Biology of Mosquitoes; Chapman and Hall: New York, NY, USA, 1992; Volume 1. [Google Scholar]
- Jiannino, J.A.; Walton, W.E. Evaluation of vegetation management strategies for controlling mosquitoes in a southern California constructed wetland. J. Amer. Mosquito Control Assn. 2004, 20, 18–26. [Google Scholar]
- Workman, P.D.; Walton, W.E. Emergence patterns of Culex mosquitoes at an experimental constructed treatment wetland in southern California. J. Amer. Mosquito Control Assn. 2000, 16, 124–130. [Google Scholar]
- Thullen, J.S.; Sartoris, J.J.; Walton, W.E. Effects of vegetation management in constructed wetland treatment cells on water quality and mosquito production. Ecol. Eng. 2002, 18, 441–457. [Google Scholar] [CrossRef]
- Lawler, S.P.; Reimer, L.; Thiemann, T.; Fritz, J.; Parise, K.; Feliz, D.; Elnaiem, D.E. Effects of vegetation control on mosquitoes in seasonal freshwater wetlands. J. Amer. Mosquito Control Assn. 2007, 23, 66–70. [Google Scholar] [CrossRef]
- de Szalay, F.A.; Euliss, N.H., Jr.; Batzer, D.P. Seasonal and Semipermanent Wetlands of California: Invertebrate Community Ecology and Responses to Management Methods. In Invertebrates in Freshwater Wetlands of North America; Batzer, D.P., Rader, R.B., Wissinger, S.A., Eds.; John Wiley and Sons: New York, NY, USA, 1999; pp. 829–855. [Google Scholar]
- Batzer, D.P.; Resh, V.H. Wetland management strategies that enhance waterfowl habitats can also control mosquitoes. J. Amer. Mosquito Control Assn. 1992, 8, 117–125. [Google Scholar]
- Flores, C.; Bounds, D.; Ruby, D. Does prescribed fire benefit wetland vegetation? Wetlands 2011, 31, 35–44. [Google Scholar] [CrossRef]
- Kwasny, D.C.; Wolder, M.; Isola, C.R. Technical Guide to Best Management Practices for Mosquito Control in Managed Wetlands; Central Valley Joint Venture, U.S. Bureau of Reclamation: Sacramento, CA, USA, 2004. [Google Scholar]
- Mayhew, C.R.; Raman, D.R.; Gerhardt, R.R.; Burns, R.T.; Younger, M.S. Periodic draining reduces mosquito emergence from free-water surface constructed wetlands. Trans. Amer. Soc. Agric. Eng. 2004, 47, 567–573. [Google Scholar]
- Carlson, D.B.; O’Meara, G.F. Mosquito Control through Source Reduction. In Florida Coordinating Council on Mosquito Control. Florida Mosquito Control: The State of the Mission as Defined by Mosquito Controllers, Regulators, and Environmental Managers; Connelly, C.R., Carlson, D.B., Eds.; University of Florida: Vero Beach, Fl, USA, 2009; pp. 38–56. [Google Scholar]
- Room, P.M.; Harley, K.L.S.; Forno, I.W.; Sands, D.P.A. Successful biological control of the floating weed salvinia. Nature 2000, 294, 78–80. [Google Scholar]
- Newman, R.M. Biological control of Eurasian watermilfoil by aquatic insects: Basic insights from an applied problem. Arch. Hydrobiol. 2004, 159, 145–184. [Google Scholar]
- Ajuonu, O.; Byrne, M.; Hill, M.; Neuenschwander, P.; Korie, S. The effect of two biological control agents, the weevil Neochetina eichhorniae and the mirid Eccritotarsus catarinensis on water hyacinth, Eichhornia crassipes, grown in culture with water lettuce, Pistia stratioes. BioControl 2009, 54, 155–162. [Google Scholar]
- Carlson, D.B.; O’Bryan, P.D.; Rey, J.R. The Management of Florida’s (USA) Salt Marsh impoundments for Mosquito Control and Natural Resource Enhancement. In Global Wetlands: Old World and New; Mitsch, W.J., Ed.; Elsevier: New York, NY, USA, 1994. [Google Scholar]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Dahl, C.; Madon, M.; Kaiser, A. Mosquitoes and Their Control, 2nd ed; Kluwer Academic Publishers: New York, NY, USA, 2010. [Google Scholar]
- Kentula, M.E. Perspectives on setting success criteria for wetland restoration. Ecol. Eng. 2000, 15, 199–209. [Google Scholar] [CrossRef]
- Committee on Mitigating Wetland LossesBoard on Environmental Studies and ToxicologyWater Science and Technology BoardDivision of Earth and Life StudiesNational Research CouncilCompensating for Wetlands Losses Under the Clean Water Act; National Academy Press: Washington, DC, USA, 2001.
- Provost, M.W. Source reduction in salt-marsh mosquito control: Past and future. Mosq. News 1977, 37, 689–698. [Google Scholar]
- Brockmeyer, R.E.; Rey, J.R.; Virnstein, R.W.; Gilmore, R.G.; Ernest, L. Rehabilitation of impounded estuarine wetlands by hydrologic reconnection to the Indian River Lagoon, Florida. Wetlands. Ecol. Mgmt. 1997, 4, 93–109. [Google Scholar]
- Bourne, W.S.; Cottam, C. Some Biological Effects of Ditching Tidewater Marshes; U.S. Fish and Wildlife Servervice: Washington, DC, USA, 1950. [Google Scholar]
- Crain, C.M.; Gedan, K.B.; Dionne, M. Tidal Restrictions and Mosquito Ditching in New England Marshes. In Human Impacts on Salt Marshes a Global Perspective; Silliman, B.R., Grosholtz, E.D., Bertness, M.D., Eds.; University of Californi : Berkely, CA, USA, 2009; pp. 149–169. [Google Scholar]
- Teal, J.; Teal, M. Life and Death of a Salt Marsh; Ballantine Books: New York, NY, USA, 1969. [Google Scholar]
- Smith, T.J., III; Tiling, G.; Leasure, P.S. Restoring coastal wetlands that were ditched for mosquito control: A preliminary assessment of hydro-leveling as a restoration technique. J. Coast. Conserv. 2007, 11, 67–74. [Google Scholar]
- Rey, J.R.; Peterson, M.S.; Kain, T.R.; Vose, F.E.; Crossman, R.A. Fish populations and physical conditions in ditched and impounded marshes in east-central Florida. NE Gulf Sci. 1990, 11, 163–170. [Google Scholar]
- Lesser, C.R.; Murohey, F.J.; Lake, R.W. Some effects of grid system mosquito-control ditching on salt-marsh biota in Delaware. Mosq. News 1976, 36, 69–77. [Google Scholar]
- LaSalle, R.; Knight, K.L. The Effects of Ditching on the Mosquito Populations in Some Sections of a Juncus Salt Marsh in Carteret County, North Carolina; Water Resources Research Institute of the University of North Carolina: Chapel Hill, NC, USA, 1973. [Google Scholar]
- Clarke, J.A.; Harrington, B.A.; Hruby, T.; Wasserman, F.E. The effect of ditching for mosquito control on salt marsh use by birds in Rowley, Massachusetts. J. Field Ornithol. 1984, 55, 160–180. [Google Scholar]
- Adamowicz, S.C.; Roman, C.T. New England salt marsh pools: A quantitative analysis of geomorphic and geographic features. Wetlands 2005, 25, 279–288. [Google Scholar] [CrossRef]
- Alsemsgeest, G.; Dale, P.; Alsemgeest, D. Evaluating the risk of potential acid sulfate soils and habitat modification for mosquito control (runneling); Comparing methods and managing the risk. Environ. Mgmt. 2005, 36, 152–161. [Google Scholar] [CrossRef]
- Dale, P.E.R.; Dale, P.T.; Hulsman, K.; Kay, B.H. Runnelling to control salt-marsh mosquitos—long-term efficacy and environmental impacts. J. Amer. Mosquito Control Assn. 1993, 9, 174–181. [Google Scholar]
- Dale, P.E.; Knight, J.M. Wetlands and mosquitoes: A review. Wetlands Ecol. Manag. 2008, 16, 255–276. [Google Scholar] [CrossRef]
- Soukup, M.A.; Portnoy, J.W. Impacts from mosquito control-induced sulfur mobilization in a Cape-Cod estuary. Environ. Conserv. 1986, 13, 47–50. [Google Scholar] [CrossRef]
- Morton, R.M.; Beumer, J.P.; Pollock, R.B. Fishes of a subtropical Australian saltmarsh and their predation upon mosquitoes. Environ. Biol. Fishes. 1988, 21, 185–194. [Google Scholar]
- Chapman, H.F.; Dale, P.E.R.; Kay, B.H. A method for assessing the effects of runneling on salt marsh grapsid crab populations. J. Amer. Mosquito Control Assn. 1998, 14, 61–68. [Google Scholar]
- Dale, P.E.; Knight, J.M. Managing salt marshes for mosquito control: Impacts of runnelling, Open Marsh Water Management and grid-ditching in sub-tropical Australia. Wetlands Ecol. Manag. 2006, 14, 211–220. [Google Scholar] [CrossRef]
- Dale, P.E. Assessing impacts of habitat modification on a subtropical salt marsh: 20 years of monitoring. Wetlands Ecol. Mgmt. 2008, 16, 77–87. [Google Scholar] [CrossRef]
- Ferrigno, F.; Jobbins, D.M. Open marsh water management. Proc. NJ Mosquito Exterm. Assoc. 1968, 55, 104–115. [Google Scholar]
- Shisler, J.K.; Lesser, F.H.; Gooley, B. Practical application of rotary ditcher in pond construction. Mosq. News 1978, 38, 112–115. [Google Scholar]
- Candeletti, R.; Candeletti, T.; Kent, R. The amphibious rotary excavator: New equipment for salt marsh management in NJ. Proc. N. J. Mosq. Cont. Assoc. 1988, 75, 102–108. [Google Scholar]
- Wolfe, R. Open marsh water management: a review of system designs and installation guidelines for mosquito control and integration in wetland habitat management. Proc. N. J. Mosq. Cont. Assoc. 2005, 92, 3–14. [Google Scholar]
- Wolfe, R.J. Effects of open marsh water management on selected tidal marsh resources: A review. J. Amer. Mosquito Control Assn. 1996, 12, 701–712. [Google Scholar]
- Bruder, K.W. The establishment of unified open marsh water management standards in New Jersey. Proc. N. J. Mosq. Cont. Assoc. 1980, 67, 72–76. [Google Scholar]
- Rochlin, I.; James-Pirri, M.-J.; Adamowicz, S.C.; Wolfe, R.J.; Capotosto, P.; Dempsey, M.E.; Iwanejko, T.; Ninivaggi, D.V. Integrated Marsh Management (IMM): A new perspective on mosquito control and best management practices for salt marsh restoration. Wetl. Ecol. Manag. 2012, 20, 219–232. [Google Scholar] [CrossRef]
- Rochlin, I.; Iwanejko, Y.; Dempsey, M.E.; Ninivaggi, D.V. Geostatistical evaluation of integrated marsh management impact on mosquito vectors using before-after-control-impact (BACI) design. Int. J. Health Geographics 2009, 8, 35. [Google Scholar] [CrossRef]
- Resh, V.H.; Balling, S.S. Tidal circulation alteration for salt-marsh mosquito-control. Environ. Manag. 1983, 7, 79–84. [Google Scholar]
- James-Pirri, M.J.; Erwin, R.M.; Prosser, D.J.; Taylor, J. Responses of salt marsh ecosystems to mosquito control management practices along the Atlantic Coast (U.S.A.). Restoration Ecol. 2011, 20, 395–404. [Google Scholar]
- le May, L.E. The Impact of Drainage Ditches on Salt Marsh Flow Patterns, Sedimentation and Morphology: Rowley River, Massachusetts.
- Pepper, M.A.; Shriver, G.W. Effects of open marsh water management on the reproductive success and nesting ecology of seaside sparrows in tidal marshes. Waterbirds 2010, 33, 381–388. [Google Scholar]
- Roman, C.T.; Raposa, K.B.; Adamowicz, S.C.; James-Pirri, M.; Catena, J.G. Quantifying vegetation and nekton response to tidal restoration of a New England salt marsh. Restoration Ecol. 2002, 10, 450–460. [Google Scholar] [CrossRef]
- Rey, J.R.; Kain, T.; Stahl, R. Wetland impoundments of east-central Florida. Fl. Scientist 1991, 54, 33–40. [Google Scholar]
- Hull, J.B.; Dove, W.E. Experimental diking for control of sand fly and mosquito breeding in Florida salt water marshes. J. Econ. Entomol. 1939, 32, 309–312. [Google Scholar]
- Clements, B.W.; Rogers, A.J. Studies of impounding for control of salt marsh mosquitoes in Florida, 1958-1963. Mosq. News 1964, 24, 264–276. [Google Scholar]
- Chapman, H.C.; Ferrigno, F. A three year study of mosquito breeding in natural and impounded salt marsh areas of New Jersey. Proc. N. J. Mosq. Exterm. Assoc. 1956, 65, 59–66. [Google Scholar]
- Darsie, R.F.; Springer, P.F. Three-year Investigation of Mosquito Breeding in Natural And Impounded Tidal Marshes in Delaware; University of Delaware Agricultural Experiment Station: Newark, NJ, USA, 1957. [Google Scholar]
- Carlson, P.; Sargent, B.; Arnold, H.; Yarbro, L.; David, J. The effects of water management practice on impoundment water quality. Bull. Fl. Anti-Mosquito Assoc. 1989, 1, 13–14. [Google Scholar]
- Rey, J.R.; Shaffer, J.; Kain, T.E.; Crossman, R.A. Sulfide variation in the pore and surface waters of artificial salt marsh ditches and a natural tidal creek. Estuaries 1992, 15, 257–269. [Google Scholar] [CrossRef]
- Harrington, R.W.; Harrington, E.S. Food selection among fishes invading a high subtropical salt marsh: From onset of flooding through the progress of a mosquito brood. Ecology 1961, 42, 646–665. [Google Scholar] [CrossRef]
- Harrington, R.W.; Harrington, E.S. Effects on fishes and their forage organisms of impounding a Florida salt marsh to prevent breeding by salt marsh mosquitoes. Bull. Mar. Sci. 1982, 32, 523–531. [Google Scholar]
- Snelson, F.F. A Study of Diverse Coastal Ecosystems on the Atlantic Coast of Florida: Ichthyological Studies. Final Report to NASA Administration Kennedy Space Center; NASA: Houston, TX, USA, 1976. [Google Scholar]
- Snelson, F.F. Ichthyofauna of the northern part of the Indian River Lagoon System, Florida. Fl. Scientist 1983, 46, 187–205. [Google Scholar]
- Gilmore, R.G.; D.W.Cooke, D.W.; Donohoe, C.J. A comparison of the fish populations and habitat in open and closed salt marsh impoundments in east-central Florida. NE Gulf Sci. 1982, 5, 25–37. [Google Scholar]
- Rey, J.R.; Kain, T.; Crossman, R.A.; Peterson, M.S.; Shaffer, J.; Vose, F.E. Zooplankton of impounded marshes and shallow areas of a subtropical lagoon. Fl. Scientist 1991, 54, 191–203. [Google Scholar]
- Rose, R.I. Pesticides and public health: Integrated methods of mosquito management. Emerg. Infect. Diseases 2001, 7, 17–23. [Google Scholar]
- Rochlin, I.; James-Pirri, M.-J.; Adamowicz, S.C.; Wolfe, R.J.; Capotosto, P.; Dempsey, M.E. Integrated Marsh Management (IMM): A new perspective on mosquito control and best management practices for salt marsh restoration. Wetlands Ecol. Mgmt. 2012, 20, 219–232. [Google Scholar] [CrossRef]
- Rupp, H.R. Adverse assessments of Gambusia affinis: An alternate view for mosquito control practitioners. J. Amer. Mosquito Control Assn. 1996, 12, 155–159. [Google Scholar]
- Mitsch, W.J., Gosselink. Wetlands, 4th ed; John Wiley & Sons: Hoboken, NJ, USA, 2007. [Google Scholar]
- Anderson, C.J.; Mitsch, W.J. Sediment, carbon, and nutrient accumulation at two 10-year-old created riverine marshes. Wetlands 2006, 26, 779–792. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Day, J.W.; Zhang, L.; Lane, R.R. Nitrate-nitrogen retention in wetlands in the Mississippi river basin. Ecol. Eng. 2005, 24, 267–278. [Google Scholar]
- Mitsch, W.J.; Day, J.W. Restoration of wetlands in the Mississippi-Ohio-Missouri (MOM) River Basin: Experience and needed research. Ecol. Eng. 2006, 26, 55–69. [Google Scholar] [CrossRef]
- Marquat-Pyatt, S.T. Are there similar sources of environmental concern? Comparing industrialized countries. Soc. Sci. Quart. 2008, 89, 1312–1335. [Google Scholar]
- Harden, F. An historical perspective of Florida’s mangrove swamps: 1565-1996. Bull. Fl. Mosquito Control Assoc. 1997, 3, 1–6. [Google Scholar]
- Tebeau, C.W. Man in the Everglades; University of Miami Press: Coral Gables, FL, USA, 1968. [Google Scholar]
- Collins, J.N.; Resh, V.H. Guidelines for the Ecological Control of Mosquitoes in Non-tidal Wetlands of the San Francisco Bay Area; University of California Mosquito Research Program: Berkeley, CA, USA, 1989. [Google Scholar]
- Walton, W.E. Protocol for Mosquito Sampling for Mosquito Best Management Practices on State of California-Managed Wildlife Areas; Integrated Pest Management Committee, Mosquito and Vector Control Association of California: Sacramento, CA, USA, 2005. [Google Scholar]
- World Health Organization, Using Climate to Predict Infectious Disease Outbreaks: A review; World Health Organization: Geneva, Switzerland, 2004.
- Tong, S.; Dale, P.; Nicholls, N.; Mackenzie, J.S.; Wolff, R.; McMichael, A.J. Climate variability, social and environmental factors, and Ross River virus transmission: Research development and future research needs. Env. Health Persp. 2008, 116, 1591–1597. [Google Scholar]
- Barker, C.M.; Reisen, W.K.; Kramer, V.L. California State Mosquito-borne Virus Surveillance and Response Plan: A retrospective evaluation using conditional simulations. Amer. J. Trop. Med. Hyg. 2003, 68, 508–518. [Google Scholar]
- Vezzani, D.; Eiras, D.F.; Wisnivesky, C. Dirofilariasis in Argentina: historical review and first report of Dirofilaria immitis in a natural mosquito population. Vet. Parasitol. 2006, 136, 259–273. [Google Scholar] [CrossRef]
- Watson, R.T.; Patz, J.; Gubler, D.J.; Parson, E.A.; Vincent, J.H. Environmental health implications of global climate change. J. Env. Monit. 2005, 7, 834–843. [Google Scholar] [CrossRef]
- Tabachnick, W.J. Challenges in predicting climate and environmental effects on vector-borne disease episystems in a changing world. J. Exp. Biol. 2010, 213, 946–954. [Google Scholar] [CrossRef]
- McMichael, A.J.; Campbell-Lendrum, D.H.; Corvalan, C.F.; Ebi, K.L.; Githeko, A.; Scheraga, J.D.; Woodward, A. Climate Change and Human Health: Risks and Responses; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rey, J.R.; Walton, W.E.; Wolfe, R.J.; Connelly, C.R.; O'Connell, S.M.; Berg, J.; Sakolsky-Hoopes, G.E.; Laderman, A.D. North American Wetlands and Mosquito Control. Int. J. Environ. Res. Public Health 2012, 9, 4537-4605. https://doi.org/10.3390/ijerph9124537
Rey JR, Walton WE, Wolfe RJ, Connelly CR, O'Connell SM, Berg J, Sakolsky-Hoopes GE, Laderman AD. North American Wetlands and Mosquito Control. International Journal of Environmental Research and Public Health. 2012; 9(12):4537-4605. https://doi.org/10.3390/ijerph9124537
Chicago/Turabian StyleRey, Jorge R., William E. Walton, Roger J. Wolfe, C. Roxanne Connelly, Sheila M. O'Connell, Joe Berg, Gabrielle E. Sakolsky-Hoopes, and Aimlee D. Laderman. 2012. "North American Wetlands and Mosquito Control" International Journal of Environmental Research and Public Health 9, no. 12: 4537-4605. https://doi.org/10.3390/ijerph9124537