Investigation into the Effect of Molds in Grasses on Their Content of Low Molecular Mass Thiols

Abstract
:1. Introduction
2. Experimental Section
2.1. Chemicals
2.2. Plant Material and Cultivation
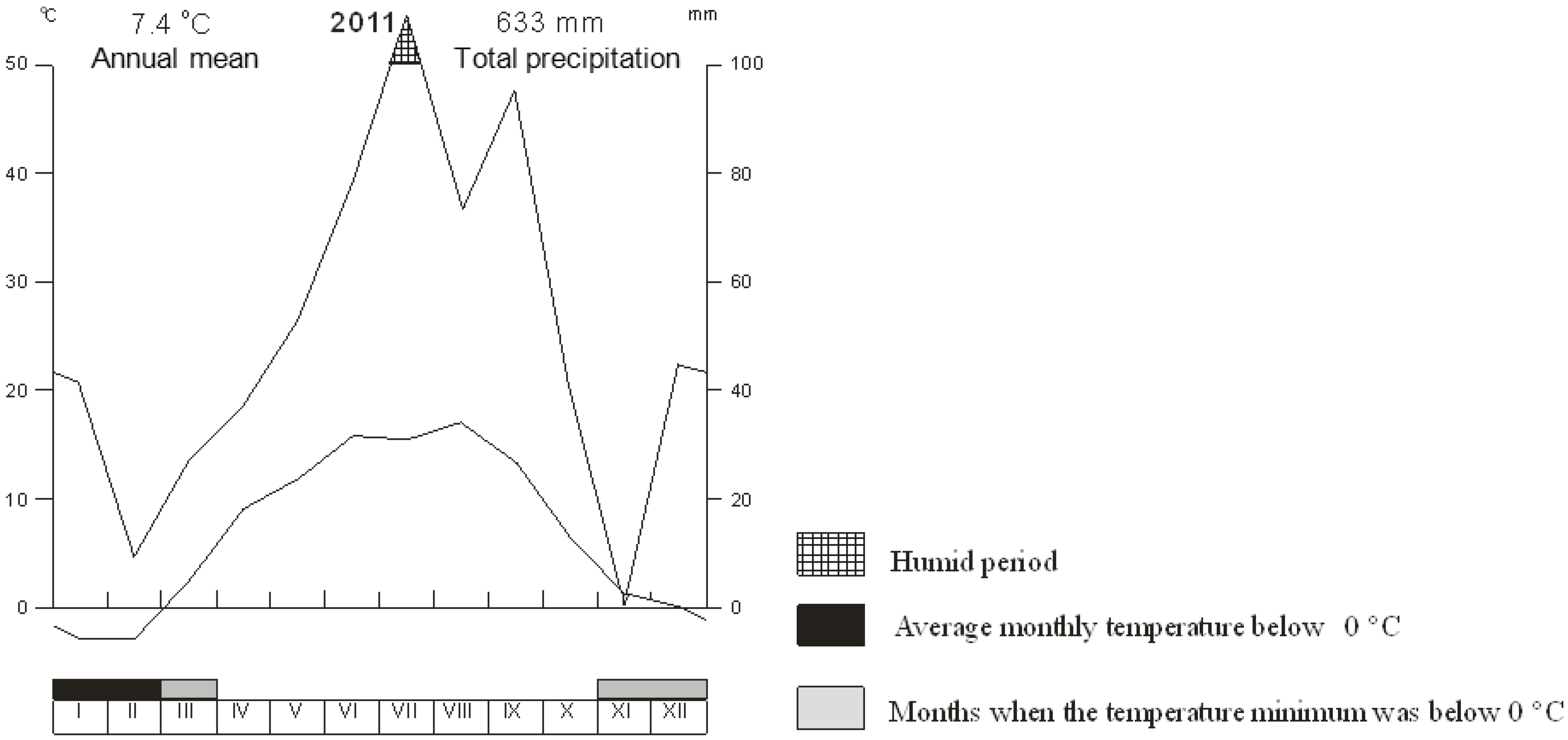
2.2.1. Description of the Site
2.2.2. Experimental Plots
2.2.3. Control Pot Experiment
2.2.4. Description of the Species
2.3. Description of Fungal Disease Identification and Assessment
2.4. Sample Preparation for Fresh Weight Analysis
2.5. Sample Preparation for Low Molecular Mass Determination
2.6. Determination of Low Molecular Mass Thiols
2.7. Descriptive Statistics
3. Results and Discussion
3.1. Mold Influence on Plants

3.2. Plant Development


{kind=link}
{kind=link}
{kind=link}
| Date evaluated | Lolium perenne | Festulolium pabulare | Festulolium braunii | |||
|---|---|---|---|---|---|---|
| No fungicide | Fungicide 1 | No fungicide | Fungicide 1 | No fungicide | Fungicide 1 | |
| 31.5.2011 | 7.3 | 7.3 | 7.7 | 7.7 | 8.0 | 7.3 |
| 12.7.2011 | 7.3 | 7.7 | 8.0 | 8.0 | 8.0 | 8.0 |
| 26.9.2011 | 3.0 | 3.0 | 4.0 | 4.0 | 3.0 | 3.0 |
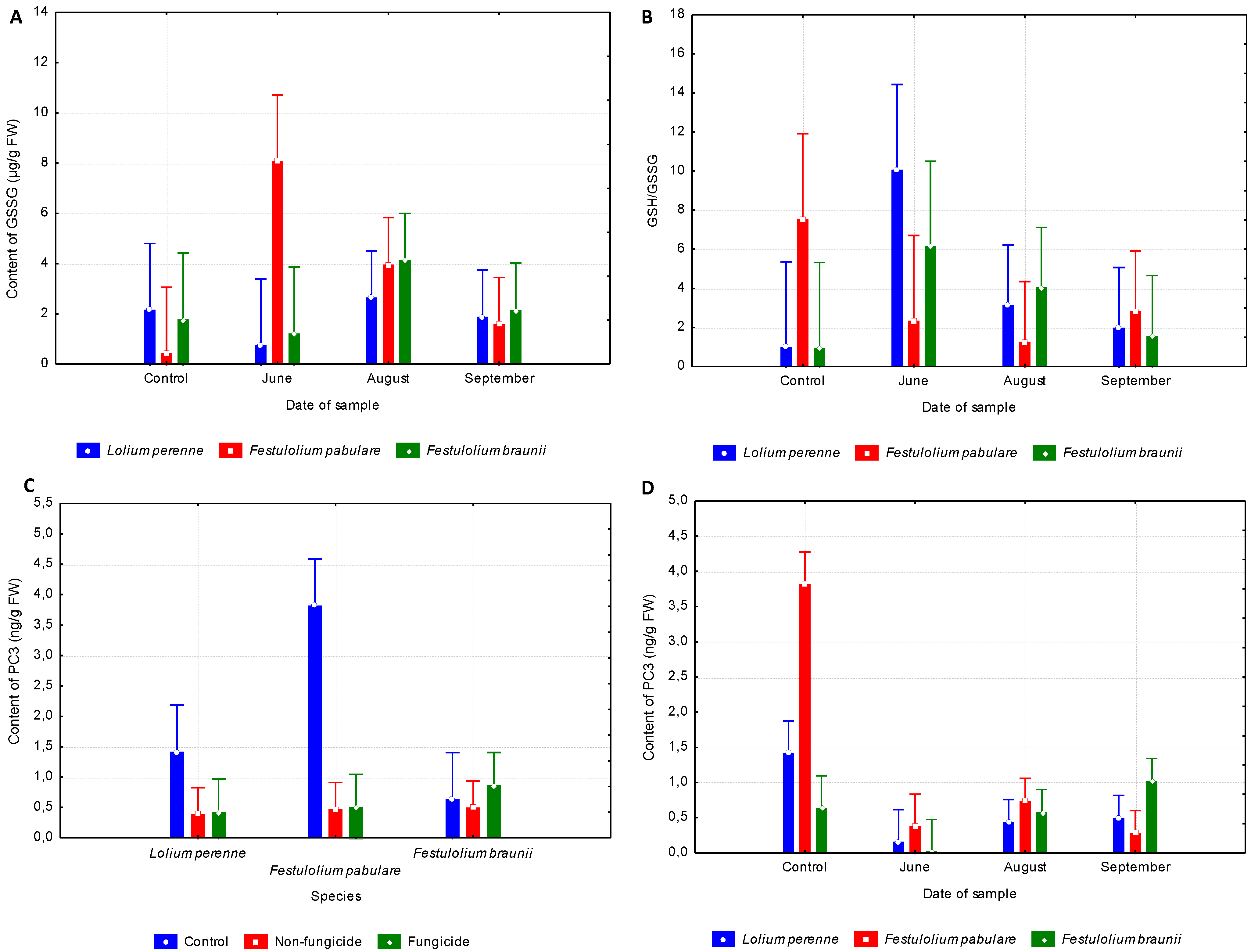
3.3. Low Molecular Mass Thiols
| Factor | ||||||||
|---|---|---|---|---|---|---|---|---|
| Species (S) | CYS | GSH | GSSG | GSH/GSSG | PC2 | PC3 | PC4 | PC5 |
| Lolium perenne | 0.65 | 2.16 | 2.16 | 1.00 | 0.239 | 1.42 | 0.74 | 1.02 |
| Festulolium pabulare | 1.43 | 3.23 | 0.43 | 7.56 | 0.28 | 3.82 | 0.44 | 0.05 |
| Festulolium braunii | 6.27 | 1.71 | 1.78 | 0.96 | 0.47 | 0.64 | 0.30 | 0.03 |
| Factor | ||||||||
|---|---|---|---|---|---|---|---|---|
| CYS | GSH | GSSG | GSH/GSSG | PC2 | PC3 | PC4 | PC5 | |
| Species (S) | ||||||||
| Lolium perenne | 2.9 ± 1.6 | 8.3 ± 4.5 | 1.5 ± 0.9 | 5.5 ± 2.5 | 0.5 ± 0.3 | 0.4 ± 0.1 | 0.6 ± 0.3a | 0.8 a ± 0.4 |
| Festulolium pabulare | 6.2 ± 1.7 | 9.0 ± 5.0 | 4.7 ± 1.8 | 1.7 ± 0.4 | 0.5 ± 0.1 | 0.5 ± 0.2 | 2.7 ± 0.3b | 2.8 a ± 1.1 |
| Festulolium braunii | 8.2 ± 3.9 | 7.4 ± 2.3 | 2.9 ± 1.1 | 3.3 ± 1.5 | 0.5 ± 0.3 | 0.5 ± 0.3 | 1.4 ± 0.2a | 0.3 a ± 0.2 |
| Date of sample (D) | ||||||||
| June | 7.1 ± 0.8 | 11.3 ± 3.8 | 3.4 ± 2.4 | 6.2 ± 2.2 | 0.3 ± 0.1 | 0.2 ± 0.1 | 1.6 ± 1.0 | 0.6 ± 0.2 |
| August | 2.5 ± 0.3 | 11.0 ± 3.2 | 4.1 ± 0.5 | 2.9 ± 1.1 | 0.7 ± 0.3 | 0.5 ± 0.2 | 1.3 ± 0.5 | 2.0 ± 1.0 |
| September | 7.6 ± 4.4 | 2.3 ± 0.8 | 1.7 ± 0.6 | 1.4 ± 0.1 | 0.5 ± 0.3 | 0.6 ± 0.3 | 1.9 ± 0.4 | 1.3 ± 1.3 |
| Factor | ||||||||
|---|---|---|---|---|---|---|---|---|
| CYS | GSH | GSSG | GSH/GSSG | PC2 | PC3 | PC4 | PC5 | |
| Species (S) | ||||||||
| Lolium perenne | 1.8 ± 0.6 | 7.0 ± 3.5 | 2.3 ± 0.6 | 2.6 ± 0.9 | 0.7 ± 0.2 | 0.5 ± 0.1 | 1.8 ± 0.7 | 0.7 ± 0.3 a |
| Festulolium pabulare | 4.8 ± 1.1 | 4.2 ± 0.6 | 2.8 ± 0.7 | 2.0 ± 0.8 | 1.2 ± 0.5 | 0.5 ± 0.1 | 3.7 ± 1.4 | 2.6 ± 0.9 b |
| Festulolium braunii | 9.7 ± 4.9 | 9.2 ± 3.7 | 3.1 ± 0.7 | 2.8 ± 1.0 | 0.7 ± 0.2 | 0.8 ± 0.2 | 1.2 ± 0.3 | 0.3 ± 0.2 a |
| Date of sample (D) | ||||||||
| August | 6.2 ± 2.9 | 9.8 ± 2.7 | 3.6 ± 0.4a | 2.8 ± 0.8 | 0.8 ± 0.2 | 0.6 ± 0.1 | 2.7 ± 1.1 | 1.2 ± 0.6 |
| September | 4.6 ± 2.4 | 3.8 ± 1.0 | 1.9 ± 0.4b | 2.1 ± 0.5 | 1.0 ± 0.3 | 0.6 ± 0.1 | 1.8 ± 0.3 | 1.2 ± 0.7 |
| Treatment (T) | ||||||||
| No fungicide | 5.0 ± 2.3 | 6.6 ± 2.4 | 2.9 ± 0.6 | 2.1 ± 0.6 | 0.6 ± 0.2 | 0.6 ± 0.1 | 1.6 ± 0.3 | 1.7 ± 0.7 |
| Fungicide | 5.8 ± 3.0 | 7.0 ± 2.5 | 2.6 ± 0.5 | 2.8 ± 0.8 | 1.2 ± 0.3 | 0.6 ± 0.1 | 2.9 ± 1.1 | 0.7 ± 0.4 |
3.4. Interactions
| Factor | CYS | GSH | GSSG | GSH/GSSG | PC2 | PC3 | PC4 | PC5 |
|---|---|---|---|---|---|---|---|---|
| D × S | 0.9915 | 0.4084 | 0.0393 | 0.0750 | 0.9928 | 0.0005 | 0.8305 | 0.5962 |
| S × T | 0.9835 | 0.9008 | 0.5880 | 0.2044 | 0.5229 | 0.0017 | 0.4169 | 0.5111 |

4. Conclusions
Acknowledgements
References
- Ryant, P.; Skladanka, J. The effect of applications of various forms of sulfur on the yields and quality of grass forage. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2009, 59, 208–216. [Google Scholar]
- Skladanka, J.; Nedelnik, J.; Adam, V.; Dolezal, P.; Moravcova, H.; Dohnal, V. Forage as a primary source of mycotoxins in animal diets. Int. J. Environ. Res. Public Health 2011, 8, 37–50. [Google Scholar]
- Barnes, R.F.; Nelson, C.J.; Moore, K.J. Forages: The Science of Grassland Agriculture, 6th ed; Iowa State University Press: Iowa City, IA, USA, 2007; p. 791. [Google Scholar]
- Malinowski, D.P.; Belesky, D.P.; Hill, N.S.; Baligar, V.C.; Fedders, J.M. Influence of phosphorus on the growth and ergot alkaloid content of neotyphodium coenophialum-infected tall fescue (festuca arundinacea schreb.). Plant Soil 1998, 198, 53–61. [Google Scholar] [CrossRef]
- Anderson, M.E. Glutathione: An overview of biosynthesis and modulation. Chem.-Biol. Interact. 1998, 112, 1–14. [Google Scholar] [CrossRef]
- Asensi, M.; Sastre, J.; Pallardo, F.V.; Lloret, A.; Lehner, M.; Garcia-de-la Asuncion, J.; Vina, J. Ratio of reduced to oxidized glutathione as indicator of oxidative stress status and DNA damage. Oxidants and Antioxidants, Pt A 1999, 299, 267–276. [Google Scholar] [CrossRef]
- Garrido, T.; Mendoza, J.; Riveros, R.; Saez, L. Acute and chronic effect of copper on levels of reduced and oxidized glutathione and nutrient uptake of tomato plants. J. Plant Nutr. Soil Sci. 2010, 173, 920–926. [Google Scholar] [CrossRef]
- Bielawski, W.; Joy, K.W. Reduced and oxidized glutathione and glutathione-reductase activity in tissues of pisum-sativum. Planta 1986, 169, 267–272. [Google Scholar] [CrossRef]
- Ogawa, K. Glutathione-associated regulation of plant growth and stress responses. Antioxid. Redox Signal. 2005, 7, 973–981. [Google Scholar] [CrossRef]
- Paradiso, A.; Berardino, R.; de Pinto, M.C.; di Toppi, L.S.; Storelli, M.M.; Tommasi, F.; De Gara, L. Increase in ascorbate-glutathione metabolism as local and precocious systemic responses induced by cadmium in durum wheat plants. Plant Cell Physiol. 2008, 49, 362–374. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- McGovern, J.J.; Isselbacher, K.; Rose, P.J.; Grossman, M.S. Observations on the glutathione (gsh) stability of red blood cells. AMA J. Dis. Child. 1958, 96, 502–502. [Google Scholar]
- Manso, C.; Wroblewski, F. Glutathione reductase activity in blood and body fluids. J. Clin. Invest. 1958, 37, 214–218. [Google Scholar] [CrossRef]
- Pisciotta, A.V.; Daly, M. Studies on agranulocytosis. 3. The reduced glutathione (gsh) content of leukocytes of normals and patients recovered from agranulocytosis. Blood 1960, 16, 1572–1578. [Google Scholar]
- Pisciotta, A.V.; Daly, M. Reduced glutathione (gsh) content of leukocytes in various hematologic diseases. Blood 1960, 15, 421–422. [Google Scholar]
- Mullineaux, P.M.; Rausch, T. Glutathione, photosynthesis and the redox regulation of stress-responsive gene expression. Photosynth. Res. 2005, 86, 459–474. [Google Scholar] [CrossRef]
- Schneider, A.; Martini, N.; Rennenberg, H. Reduced glutathione (gsh) transport into cultured tobacco cells. Plant Physiol. Biochem. 1992, 30, 29–38. [Google Scholar]
- Herschbach, C.; Rennenberg, H. Influence of glutathione (gsh) on net uptake of sulfate and sulfate transport in tobacco plants. J. Exp. Bot. 1994, 45, 1069–1076. [Google Scholar] [CrossRef]
- Gelhaye, E.; Rouhier, N.; Jacquot, J.P. Evidence for a subgroup of thioredoxin h that requires gsh/grx for its reduction. FEBS Lett. 2003, 555, 443–448. [Google Scholar] [CrossRef]
- Rausch, T.; Gromes, R.; Liedschulte, V.; Muller, I.; Bogs, J.; Galovic, V.; Wachter, A. Novel insight into the regulation of gsh biosynthesis in higher plants. Plant Biol. 2007, 9, 565–572. [Google Scholar] [CrossRef]
- Liedschulte, V.; Wachter, A.; An, Z.G.; Rausch, T. Exploiting plants for glutathione (gsh) production: Uncoupling gsh synthesis from cellular controls results in unprecedented gsh accumulation. Plant Biotechnol. J. 2010, 8, 807–820. [Google Scholar] [CrossRef]
- Suh, M.C.; Choi, D.; Liu, J.R. Cadmium resistance in transgenic tobacco plants expressing the nicotiana glutinosa l. Metallothionein-like gene. Mol. Cells 1998, 8, 678–684. [Google Scholar]
- Liu, J.Y.; Lu, T.; Zhao, N.M. Classification and nomenclature of plant metallothionein-like proteins based on their cysteine arrangement patterns. Acta Bot. Sinica 2000, 42, 649–652. [Google Scholar]
- He, H.Z.; Zhu, C.M.; Lu, T.; Zhang, R.Q.; Zhao, N.M.; Liu, J.Y. Modeling the cysteine rich domain of plant metallothionein-like protein. Acta Bot. Sinica 2002, 44, 1155–1159. [Google Scholar]
- Lu, T.; Liu, J.Y.; Zhang, R.Q.; Zhao, N.M. Modeling rice rgmt as a plant metallothionein-like protein by the distance geometry and homology methods. Acta Bot. Sinica 2003, 45, 1297–1306. [Google Scholar]
- Plocke, D.J. Cadmium-binding peptide complexes from schizosaccharomyces-pombe. Methods Enzymol. 1991, 205, 603–610. [Google Scholar]
- Grill, E.; Gekeler, W.; Winnacker, E.L.; Zenk, H.H. Homo-phytochelatins are heavy metal-binding peptides of homo-glutathione containing fabales. FEBS Lett. 1986, 205, 47–50. [Google Scholar] [CrossRef]
- Rauser, W.E. Phytochelatins. Annu. Rev. Biochem. 1990, 59, 61–86. [Google Scholar] [CrossRef]
- Cobbett, C.S. Phytochelatin biosynthesis and function in heavy-metal detoxification. Curr. Opin. Plant Biol. 2000, 3, 211–216. [Google Scholar]
- Cobbett, C.S. Heavy metal detoxification in plants: Phytochelatin biosynthesis and function. IUBMB Life 2001, 51, 183–188. [Google Scholar] [CrossRef]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef]
- Pal, R.; Rai, J.P.N. Phytochelatins: Peptides involved in heavy metal detoxification. Appl. Biochem. Biotechnol. 2010, 160, 945–963. [Google Scholar] [CrossRef]
- Diopan, V.; Shestivska, V.; Adam, V.; Macek, T.; Mackova, M.; Havel, L.; Kizek, R. Determination of content of metallothionein and low molecular mass stress peptides in transgenic tobacco plants. Plant Cell Tissue Organ Cult. 2008, 94, 291–298. [Google Scholar] [CrossRef]
- Janouskova, M.; Pavlikova, D.; Macek, T.; Vosatka, M. Influence of arbuscular mycorrhiza on the growth and cadmium uptake of tobacco with inserted metallothionein gene. Appl. Soil Ecol. 2005, 29, 209–214. [Google Scholar] [CrossRef]
- Janouskova, M.; Pavlikova, D.; Macek, T.; Vosatka, M. Arbuscular mycorrhiza decreases cadmium phytoextraction by transgenic tobacco with inserted metallothionein. Plant Soil 2005, 272, 29–40. [Google Scholar]
- Kotrba, P.; Macek, T.; Ruml, T. Heavy metal-binding peptides and proteins in plants. A review. Collect. Czech. Chem. Commun. 1999, 64, 1057–1086. [Google Scholar] [CrossRef]
- Macek, T.; Mackova, M.; Pavlikova, D.; Szakova, J.; Truksa, M.; Cundy, S.; Kotrba, P.; Yancey, N.; Scouten, W.H. Accumulation of cadmium by transgenic tobacco. Acta Biotechnol. 2002, 22, 101–106. [Google Scholar] [CrossRef]
- Macek, T.; Mackova, M.; Truksa, M.; Cundy, A.S.; Kotrba, P.; Yancey, N.; Schouten, W.H. Preparation of transgenic tobacco with a yeast metallothionein combined with a polyhistidine tail. Chem. Listy 1996, 90, 690–690. [Google Scholar]
- Pavlikova, D.; Macek, T.; Mackova, M.; Sura, M.; Szakova, J.; Tlustos, P. The evaluation of cadmium, zinc and nickel accumulation ability of transgenic tobacco bearing different transgenes. Plant Soil Environ. 2004, 50, 513–517. [Google Scholar]
- Shestivska, V.; Adam, V.; Prasek, J.; Macek, T.; Mackova, M.; Havel, L.; Diopan, V.; Zehnalek, J.; Hubalek, J.; Kizek, R. Investigation of the antioxidant properties of metallothionein in transgenic tobacco plants using voltammetry at a carbon paste electrode. Int. J. Electrochem. Sci. 2011, 6, 2869–2883. [Google Scholar]
- Supalkova, V.; Petrek, J.; Baloun, J.; Adam, V.; Bartusek, K.; Trnkova, L.; Beklova, M.; Diopan, V.; Havel, L.; Kizek, R. Multi-instrumental investigation of affecting of early somatic embryos of spruce by cadmium(ii) and lead(ii) ions. Sensors 2007, 7, 743–759. [Google Scholar]
- Diopan, V.; Stejskal, K.; Galiova, M.; Adam, V.; Kaiser, J.; Horna, A.; Novotny, K.; Liska, M.; Havel, L.; Zehnalek, J.; et al. Determination of plant thiols by liquid chromatography coupled with coulometric and amperometric detection in lettuce treated by lead(ii) ions. Electroanalysis 2010, 22, 1248–1259. [Google Scholar] [CrossRef]
- Potesil, D.; Petrlova, J.; Adam, V.; Vacek, J.; Klejdus, B.; Zehnalek, J.; Trnkova, L.; Havel, L.; Kizek, R. Simultaneous femtomole determination of cysteine, reduced and oxidized glutathione, and phytochelatin in maize (zea mays l.) kernels using high-performance liquid chromatography with electrochemical detection. J. Chromatogr. A 2005, 1084, 134–144. [Google Scholar] [CrossRef]
- Petrlova, J.; Mikelova, R.; Stejskal, K.; Kleckerova, A.; Zitka, O.; Petrek, J.; Havel, L.; Zehnalek, J.; Adam, V.; Trnkova, L.; et al. Simultaneous determination of eight biologically active thiol compounds using gradient elution-liquid chromatography with coul-array detection. J. Sep. Sci. 2006, 29, 1166–1173. [Google Scholar]
- Zitka, O.; Krystofova, O.; Sobrova, P.; Adam, V.; Zehnalek, J.; Beklova, M.; Kizek, R. Phytochelatin synthase activity as a marker of metal pollution. J. Hazard. Mater. 2011, 192, 794–800. [Google Scholar] [CrossRef]
- Zitka, O.; Skutkova, H.; Krystofova, O.; Sobrova, P.; Adam, V.; Zehnalek, J.; Havel, L.; Beklova, M.; Hubalek, J.; Provaznik, I.; et al. Rapid and ultrasensitive method for determination of phytochelatin(2) using high performance liquid chromatography with electrochemical detection. Int. J. Electrochem. Sci. 2011, 6, 1367–1381. [Google Scholar]
- Studer, B.; Boller, B.; Bauer, E.; Posselt, U.K.; Widmer, F.; Kolliker, R. Consistent detection of qtls for crown rust resistance in italian ryegrass (lolium multiflorum lam.) across environments and phenotyping methods. Theor. Appl. Genet. 2007, 115, 9–17. [Google Scholar] [CrossRef]
- Kleckerova, A.; Sobrova, P.; Krystofova, O.; Sochor, J.; Zitka, O.; Babula, P.; Adam, V.; Docekalova, H.; Kizek, R. Cadmium(ii) and zinc(ii) ions effects on maize plants revealed by spectroscopy and electrochemistry. Int. J. Electrochem. Sci. 2011, 6, 6011–6031. [Google Scholar]
- Krizkova, S.; Krystofova, O.; Trnkova, L.; Hubalek, J.; Adam, V.; Beklova, M.; Horna, A.; Havel, L.; Kizek, R. Silver(i) ions ultrasensitive detection at carbon electrodes—Analysis of waters, tobacco cells and fish tissues. Sensors 2009, 9, 6934–6950. [Google Scholar] [CrossRef]
- Krizkova, S.; Ryant, P.; Krystofova, O.; Adam, V.; Galiova, M.; Beklova, M.; Babula, P.; Kaiser, J.; Novotny, K.; Novotny, J. Multi-instrumental analysis of tissues of sunflower plants treated with silver(i) ions—Plants as bioindicators of environmental pollution. Sensors 2008, 8, 445–463. [Google Scholar]
- Krystofova, O.; Shestivska, V.; Galiova, M.; Novotny, K.; Kaiser, J.; Zehnalek, J.; Babula, P.; Opatrilova, R.; Adam, V.; Kizek, R. Sunflower plants as bioindicators of environmental pollution with lead(ii) ions. Sensors 2009, 9, 5040–5058. [Google Scholar]
- Krystofova, O.; Trnkova, L.; Adam, V.; Zehnalek, J.; Hubalek, J.; Babula, P.; Kizek, R. Electrochemical microsensors for the detection of cadmium(ii) and lead(ii) ions in plants. Sensors 2010, 10, 5308–5328. [Google Scholar] [CrossRef]
- Memon, A.R.; Schroder, P. Implications of metal accumulation mechanisms to phytoremediation. Environ. Sci. Pollut. Res. 2009, 16, 162–175. [Google Scholar] [CrossRef]
- Verkleij, J.A.C.; Golan-Goldhirsh, A.; Antosiewisz, D.M.; Schwitzguebel, J.P.; Schroder, P. Dualities in plant tolerance to pollutants and their uptake and translocation to the upper plant parts. Environ. Exp. Bot. 2009, 67, 10–22. [Google Scholar] [CrossRef]
- Abhilash, P.C.; Jamil, S.; Singh, N. Transgenic plants for enhanced biodegradation and phytoremediation of organic xenobiotics. Biotechnol. Adv. 2009, 27, 474–488. [Google Scholar] [CrossRef]
- Lyubenova, L.; Schroder, P. Plants for waste water treatment—Effects of heavy metals on the detoxification system of typha latifolia. Bioresour. Technol. 2011, 102, 996–1004. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Liu, J.H. Transgenic alfalfa plants co-expressing glutathione s-transferase (gst) and human cyp2e1 show enhanced resistance to mixed contaminates of heavy metals and organic pollutants. J. Hazard. Mater. 2011, 189, 357–362. [Google Scholar] [CrossRef]
- Grossmann, K.; Retzlaff, G. Bioregulatory effects of the fungicidal strobilurin kresoxim-methyl in wheat (triticum aestivum). Pestic. Sci. 1997, 50, 11–20. [Google Scholar] [CrossRef]
- Glaab, J.; Kaiser, W.M. Increased nitrate reductase activity in leaf tissue after application of the fungicide kresoxim-methyl. Planta 1999, 207, 442–448. [Google Scholar] [CrossRef]
- Grossmann, K.; Kwiatkowski, J.; Casper, G. Regulation of phytohormone levels, leaf senescence and transpiration by the strobilurin kresoxim-methyl in wheat (triticum aestivum). J. Plant Physiol. 1999, 154, 805–808. [Google Scholar] [CrossRef]
- Nason, M.A.; Farrar, J.; Bartlett, D. Strobilurin fungicides induce changes in photosynthetic gas exchange that do not improve water use efficiency of plants grown under conditions of water stress. Pest Manag. Sci. 2007, 63, 1191–1200. [Google Scholar] [CrossRef]
- Wu, Y.X.; von Tiedemann, A. Physiological effects of azoxystrobin and epoxiconazole on senescence and the oxidative status of wheat. Pest. Biochem. Physiol. 2001, 71, 1–10. [Google Scholar] [CrossRef]
- Wu, Y.X.; von Tiedemann, A. Impact of fungicides on active oxygen species and antioxidant enzymes in spring barley (hordeum vulgare l.) exposed to ozone. Environ. Pollut. 2002, 116, 37–47. [Google Scholar] [CrossRef]
- Fernandez-Ortuno, D.; Tores, J.A.; De Vicente, A.; Perez-Garcia, A. Mechanisms of resistance to qol fungicides in phytopathogenic fungi. Int. Microbiol. 2008, 11, 1–9. [Google Scholar]
- Mari, M.; Morales, A.; Colell, A.; Garcia-Ruiz, C.; Fernandez-Checa, J.C. Mitochondrial glutathione, a key survival antioxidant. Antioxid. Redox Signal. 2009, 11, 2685–2700. [Google Scholar] [CrossRef]
- Giesler, L.J.; Yuen, G.Y.; Horst, G.L. The microclimate in tall fescue turf as affected by canopy density and its influence on brown patch disease. Plant Dis. 1996, 80, 389–394. [Google Scholar] [CrossRef]
- von Boberfeld, W.O.; Banzhaf, K. Yield and forage quality of different xfestulolium cultivars in winter. J. Agron. Crop Sci. 2006, 192, 239–247. [Google Scholar] [CrossRef]
- Becana, M.; Matamoros, M.A.; Udvardi, M.; Dalton, D.A. Recent insights into antioxidant defenses of legume root nodules. New Phytol. 2010, 188, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Shen, G.M.; Zhu, C.; Shangguan, L.N.; Du, Q.Z. The cd-tolerant rice mutant cadh-5 is a high cd accumulator and shows enhanced antioxidant activity. J. Plant Nutr. Soil Sci. 2012, 175, 309–318. [Google Scholar] [CrossRef]
- Raab, A.; Feldmann, J.; Meharg, A.A. The nature of arsenic-phytochelatin complexes in holcus lanatus and pteris cretica. Plant Physiol. 2004, 134, 1113–1122. [Google Scholar] [CrossRef]
- Hunaiti, A.A.; Al-Oqlah, A.; Shannag, N.M.; Abukhalaf, I.K.; Silvestrov, N.A.; von Deutsch, D.A.; Bayorh, M.A. Toward understanding the influence of soil metals and sullfate content on plant thiols. J. Toxicol. Env. Health Part A 2007, 70, 559–567. [Google Scholar] [CrossRef]
- Jia, Y.; Tang, S.R.; Wang, R.G.; Ju, X.H.; Ding, Y.Z.; Tu, S.X.; Smith, D.L. Effects of elevated co2 on growth, photosynthesis, elemental composition, antioxidant level, and phytochelatin concentration in lolium mutiforum and lolium perenne under cd stress. J. Hazard. Mater. 2010, 180, 384–394. [Google Scholar] [CrossRef]
- Supalkova, V.; Huska, D.; Diopan, V.; Hanustiak, P.; Zitka, O.; Stejskal, K.; Baloun, J.; Pikula, J.; Havel, L.; Zehnalek, J.; et al. Electroanalysis of plant thiols. Sensors 2007, 7, 932–959. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Skladanka, J.; Adam, V.; Zitka, O.; Krystofova, O.; Beklova, M.; Kizek, R.; Havlicek, Z.; Slama, P.; Nawrath, A. Investigation into the Effect of Molds in Grasses on Their Content of Low Molecular Mass Thiols. Int. J. Environ. Res. Public Health 2012, 9, 3789-3805. https://doi.org/10.3390/ijerph9113789
Skladanka J, Adam V, Zitka O, Krystofova O, Beklova M, Kizek R, Havlicek Z, Slama P, Nawrath A. Investigation into the Effect of Molds in Grasses on Their Content of Low Molecular Mass Thiols. International Journal of Environmental Research and Public Health. 2012; 9(11):3789-3805. https://doi.org/10.3390/ijerph9113789
Chicago/Turabian StyleSkladanka, Jiri, Vojtech Adam, Ondrej Zitka, Olga Krystofova, Miroslava Beklova, Rene Kizek, Zdenek Havlicek, Petr Slama, and Adam Nawrath. 2012. "Investigation into the Effect of Molds in Grasses on Their Content of Low Molecular Mass Thiols" International Journal of Environmental Research and Public Health 9, no. 11: 3789-3805. https://doi.org/10.3390/ijerph9113789