Epistasis between IL1A, IL1B, TNF, HTR2A, 5-HTTLPR and TPH2 Variations Does Not Impact Alcohol Dependence Disorder Features

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Inclusion and Exclusion Criteria
2.2. Assessment
2.3. Statistics
Principal analysis
Secondary analysis
3. Results
4. Discussion
Abbreviations:
| 5-HTTLPR | promoter of the gene of the serotonin transporter; |
| CIDI | Composite International Diagnostic Interview; |
| GAS | Global Assessment Scale; |
| GGT | gamma-glutamyltransferase; |
| GMDR | general multifactor dimensionality reduction; |
| HAM-A | Hamilton Anxiety Rating Scale; |
| HAM-D | Hamilton Depression Rating Scale; |
| Hb | hemoglobin; |
| HDL | high density lipoprotein; |
| HTc | hematocrit; |
| HTR2A | serotonin receptor 2A; |
| IL1A | interleukin 1, alpha; |
| IL1B | interleukin 1, beta; |
| LDL | low density lipoprotein; |
| MDR | multifactor dimensionality reduction; |
| SCAN | Schedules for Clinical Assessment in Neuropsychiatry; |
| SGOT (or AST) | glutamic oxaloacetic transaminase; |
| SGPT (or ALT) | glutamic pyruvic transaminase; |
| T levels | thyroid hormonal levels; |
| T3 | Triiodothyronine; |
| T4 | Thyroxine; |
| TNF | tumor necrosis factor; |
| TPH2 | tryptophan hydroxylase 2; |
| TSH | Thyrotropin. |
Acknowledgments
References
- Serretti, A; Artioli, P. The pharmacogenomics of selective serotonin reuptake inhibitors. Pharmacogenomics J 2004, 4, 233–244. [Google Scholar]
- Fisher, PM; Munoz, KE; Hariri, AR. Identification of neurogenetic pathways of risk for psychopathology. Am. J. Med. Genet. C Semin. Med. Genet 2008, 2, 147–153. [Google Scholar]
- D’Souza, UM; Craig, IW. Functional polymorphisms in dopamine and serotonin pathway genes. Hum. Mutat 2006, 1, 1–13. [Google Scholar]
- Lesch, KP. Alcohol dependence and gene x environment interaction in emotion regulation: Is serotonin the link? Eur. J. Pharmacol 2005, 1, 113–124. [Google Scholar]
- Vengeliene, V; Bilbao, A; Molander, A; Spanagel, R. Neuropharmacology of alcohol addiction. Br. J. Pharmacol 2008, 2, 299–315. [Google Scholar]
- Serretti, A; Calati, R; Mandelli, L; de Ronchi, D. Serotonin transporter gene variants and behavior: a comprehensive review. Curr. Drug Target 2006, 12, 1659–1669. [Google Scholar]
- Pinto, E; Reggers, J; Gorwood, P; Boni, C; Scantamburlo, G; Pitchot, W; Ansseau, M. The short allele of the serotonin transporter promoter polymorphism influences relapse in alcohol dependence. Alcohol Alcoholism 2008, 4, 398–400. [Google Scholar]
- Dick, DM; Plunkett, J; Hamlin, D; Nurnberger, J, Jr; Kuperman, S; Schuckit, M; Hesselbrock, V; Edenberg, H; Bierut, L. Association analyses of the serotonin transporter gene with lifetime depression and alcohol dependence in the Collaborative Study on the Genetics of Alcoholism (COGA) sample. Psychiatr. Genet 2007, 1, 35–38. [Google Scholar]
- Marques, FZ; Hutz, MH; Bau, CH. Influence of the serotonin transporter gene on comorbid disorders among alcohol-dependent individuals. Psychiatr. Genet 2006, 3, 125–131. [Google Scholar]
- Kohnke, MD; Kolb, W; Lutz, U; Maurer, S; Batra, A. The serotonin transporter promotor polymorphism 5-HTTLPR is not associated with alcoholism or severe forms of alcohol withdrawal in a German sample. Psychiatr. Genet 2006, 6, 227–228. [Google Scholar]
- Rasmussen, H; Bagger, Y; Tanko, LB; Christiansen, C; Werge, T. Lack of association of the serotonin transporter gene promoter region polymorphism, 5-HTTLPR, including rs25531 with cigarette smoking and alcohol consumption. Am. J. Med. Genet. B Neuropsychiatr. Genet 2009, 150, 575–580. [Google Scholar]
- Serretti, A; Drago, A; De Ronchi, D. HTR2A gene variants and psychiatric disorders: a review of current literature and selection of SNPs for future studies. Curr. Med. Chem 2007, 19, 2053–2069. [Google Scholar]
- Polina, ER; Contini, V; Hutz, MH; Bau, CH. The serotonin 2A receptor gene in alcohol dependence and tobacco smoking. Drug Alcohol Depend 2009, 1, 128–131. [Google Scholar]
- Hwu, HG; Chen, CH. Association of 5HT2A receptor gene polymorphism and alcohol abuse with behavior problems. Am. J. Med. Genet 2000, 6, 797–800. [Google Scholar]
- Nakamura, T; Matsushita, S; Nishiguchi, N; Kimura, M; Yoshino, A; Higuchi, S. Association of a polymorphism of the 5HT2A receptor gene promoter region with alcohol dependence. Mol. Psychiat 1999, 1, 85–88. [Google Scholar]
- Himmerich, H; Fulda, S; Linseisen, J; Seiler, H; Wolfram, G; Himmerich, S; Gedrich, K; Kloiber, S; Lucae, S; Ising, M; Uhr, M; Holsboer, F; Pollmacher, T. Depression, comorbidities and the TNF-alpha system. Eur. Psychiat 2008, 6, 421–429. [Google Scholar]
- Zill, P; Preuss, UW; Koller, G; Bondy, B; Soyka, M. SNP- and haplotype analysis of the tryptophan hydroxylase 2 gene in alcohol-dependent patients and alcohol-related suicide. Neuropsychopharmacology 2007, 8, 1687–1694. [Google Scholar]
- Mokrovic, G; Matosic, A; Hranilovic, D; Stefulj, J; Novokmet, M; Oreskovic, D; Balija, M; Marusic, S; Cicin-Sain, L. Alcohol dependence and polymorphisms of serotonin-related genes: association studies. Coll. Antropol 2008, 32, 127–131. [Google Scholar]
- Gacek, P; Conner, TS; Tennen, H; Kranzler, HR; Covault, J. Tryptophan hydroxylase 2 gene and alcohol use among college students. Addict. Biol 2008, 3, 440–448. [Google Scholar]
- Yu, YW; Chen, TJ; Hong, CJ; Chen, HM; Tsai, SJ. Association study of the interleukin-1 beta (C-511T) genetic polymorphism with major depressive disorder, associated symptomatology, and antidepressant response. Neuropsychopharmacology 2003, 6, 1182–1185. [Google Scholar]
- Pastor, IJ; Laso, FJ; Romero, A; Gonzalez-Sarmiento, R. Interleukin-1 gene cluster polymorphisms and alcoholism in Spanish men. Alcohol Alcoholism 2005, 3, 181–186. [Google Scholar]
- Serretti, A; Liappas, I; Mandelli, L; Albani, D; Forloni, G; Malitas, P; Piperi, C; Zisaki, A; Tzavellas, EO; Papadopoulou-Daifoti, Z; Prato, F; Batelli, S; Pesaresi, M; Kalofoutis, A. Interleukin-1 alpha and beta, TNF-alpha and HTTLPR gene variants study on alcohol toxicity and detoxification outcome. Neurosci. Lett 2006, 1, 107–112. [Google Scholar]
- Serretti, A; Liappas, I; Mandelli, L; Albani, D; Forloni, G; Malitas, P; Piperi, C; Politis, A; Tzavellas, EO; Papadopoulou-Daifoti, Z; Zisaki, A; Prato, F; Batelli, S; Polito, L; De Ronchi, D; Kalofoutis, A. TPH2 gene variants and anxiety during alcohol detoxification outcome. Psychiat. Res 2009, 1, 106–114. [Google Scholar]
- Wing, JK; Babor, T; Brugha, T; Burke, J; Cooper, JE; Giel, R; Jablenski, A; Regier, D; Sartorius, N. SCAN, schedules for clinical assessment in neuropsychiatry. Arch. Gen. Psychiat 1990, 6, 589–593. [Google Scholar]
- Ye, J; Wang, L; Zhang, X; Tantishaiyakul, V; Rojanasakul, Y. Inhibition of TNF-alpha gene expression and bioactivity by site-specific transcription factor-binding oligonucleotides. Am. J. Physiol. Lung Cell Mol. Physiol 2003, 2, L386–L394. [Google Scholar]
- Liebowitz, MR. Social phobia. Mod. Probl. Pharm 1987, 22, 141–173. [Google Scholar]
- Marks, IM; Mathews, AM. Brief standard self-rating for phobic patients. Behav. Res. Ther 1979, 3, 263–267. [Google Scholar]
- Cooper, JE. The Leyton Obsessional Inventory. Psychol. Med 1970, 1, 48–64. [Google Scholar]
- Pilowsky, I. Dimensions of hypochondriasis. Br. J. Psychiat 1967, 494, 89–93. [Google Scholar]
- Lou, XY; Chen, GB; Yan, L; Ma, JZ; Zhu, J; Elston, RC; Li, MD. A generalized combinatorial approach for detecting gene-by-gene and gene-by-environment interactions with application to nicotine dependence. Am. J. Hum. Genet 2007, 6, 1125–1137. [Google Scholar]
- Drago, A; de Ronchi, D; Serretti, A. Incomplete coverage of candidate genes: a poorly considered bias. Curr. Genomics 2007, 7, 476–483. [Google Scholar]
- Walther, DJ; Peter, JU; Bashammakh, S; Hortnagl, H; Voits, M; Fink, H; Bader, M. Synthesis of serotonin by a second tryptophan hydroxylase isoform. Science 2003, 299, 76. [Google Scholar]
- Zill, P; Baghai, TC; Zwanzger, P; Schule, C; Eser, D; Rupprecht, R; Moller, HJ; Bondy, B; Ackenheil, M. SNP and haplotype analysis of a novel tryptophan hydroxylase isoform (TPH2) gene provide evidence for association with major depression. Mol. Psychiat 2004, 11, 1030–1036. [Google Scholar]
- Zhou, Z; Peters, EJ; Hamilton, SP; McMahon, F; Thomas, C; McGrath, PJ; Rush, J; Trivedi, MH; Charney, DS; Roy, A; Wisniewski, S; Lipsky, R; Goldman, D. Response to Zhang et al. 2005: loss-of-function mutation in tryptophan hydroxylase-2 identified in unipolar major depression. Neuron 2005, 48, 704–705. [Google Scholar]
- Zhou, Z; Roy, A; Lipsky, R; Kuchipudi, K; Zhu, G; Taubman, J; Enoch, MA; Virkkunen, M; Goldman, D. Haplotype-based linkage of tryptophan hydroxylase 2 to suicide attempt, major depression, and cerebrospinal fluid 5-hydroxyindoleacetic acid in 4 populations. Arch. Gen. Psychiat 2005, 10, 1109–1118. [Google Scholar]
- Harvey, M; Shink, E; Tremblay, M; Gagne, B; Raymond, C; Labbe, M; Walther, DJ; Bader, M; Barden, N. Support for the involvement of TPH2 gene in affective disorders. Mol. Psychiat 2004, 11, 980–981. [Google Scholar]
- de Luca, V; Likhodi, O; van Tol, HH; Kennedy, JL; Wong, AH. Tryptophan hydroxylase 2 gene expression and promoter polymorphisms in bipolar disorder and schizophrenia. Psychopharmacology (Berl) 2005, 3, 378–382. [Google Scholar]
- Hoebel, BG; Hernandez, L; McClelland, RC; Schwartz, D. Dexfenfluramine and feeding reward. Clin. Neuropharmacol 1988, 11, S72–S85. [Google Scholar]
- Jadhav, SA; Gaikwad, RV; Gaonkar, RK; Thorat, VM; Gursale, SC; Balsara, JJ. Dose-dependent response of central dopaminergic systems to buspirone in mice. Indian J. Exp. Biol 2008, 10, 704–714. [Google Scholar]
- Urwin, RE; Nunn, KP. Epistatic interaction between the monoamine oxidase A and serotonin transporter genes in anorexia nervosa. Eur. J. Hum. Genet 2005, 3, 370–375. [Google Scholar]
- van Gestel, S; Forsgren, T; Claes, S; Del-Favero, J; van Duijn, CM; Sluijs, S; Nilsson, LG; Adolfsson, R; van Broeckhoven, C. Epistatic effect of genes from the dopamine and serotonin systems on the temperament traits of novelty seeking and harm avoidance. Mol. Psychiat 2002, 5, 448–450. [Google Scholar]
- Sullivan, PF. Spurious genetic associations. Biol. Psychiat 2007, 10, 1121–1126. [Google Scholar]
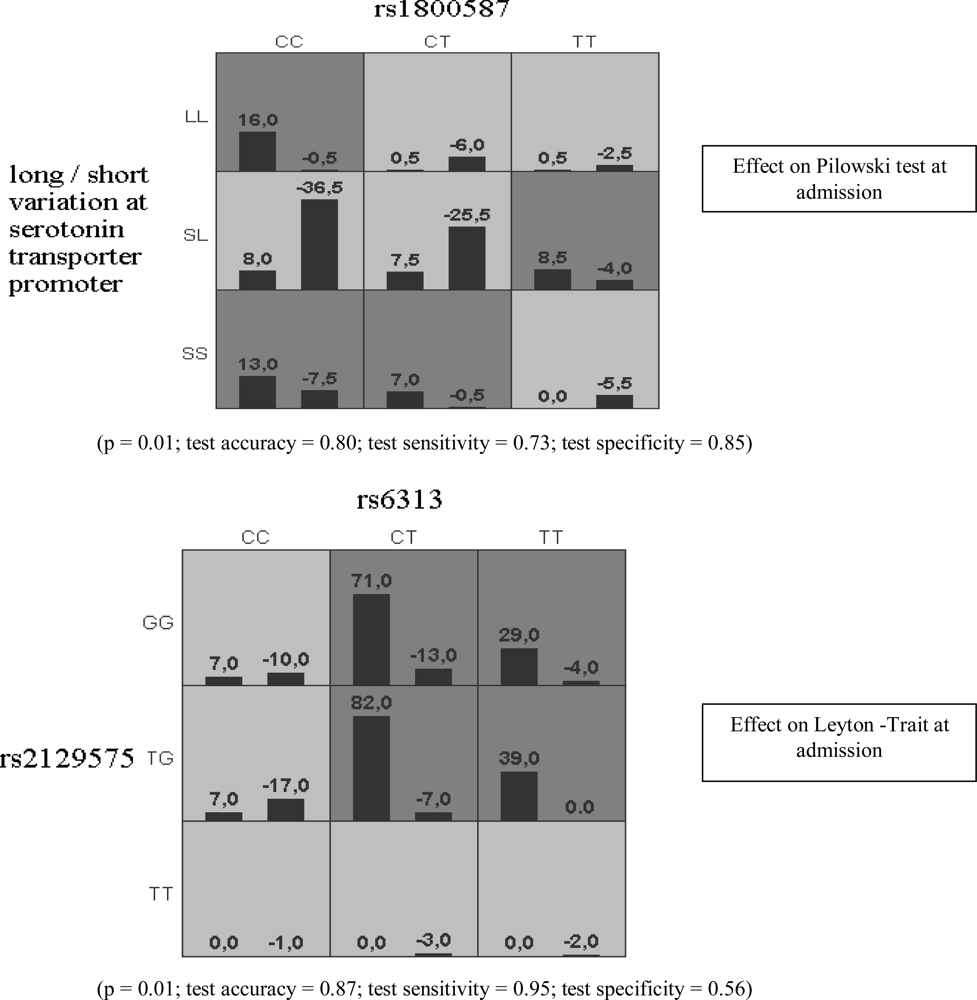

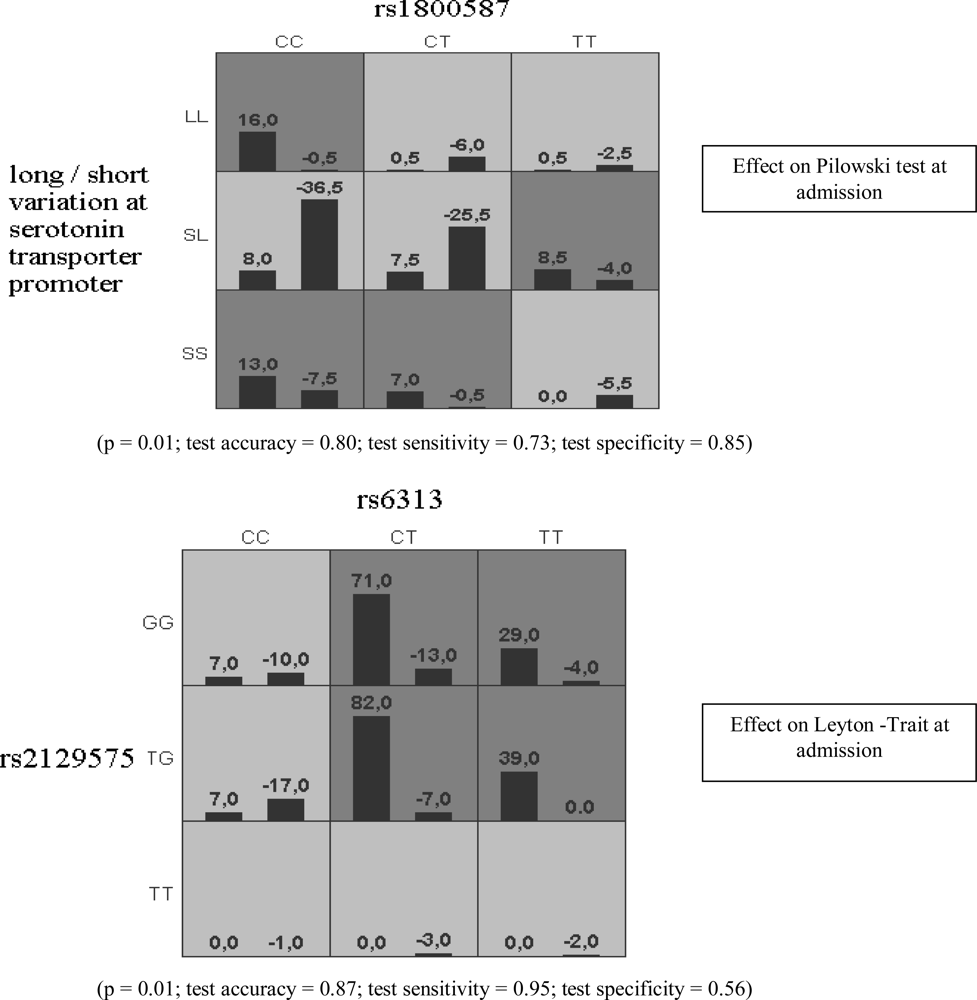{kind=link}
| Variable | Total | Females | Males |
|---|---|---|---|
| Sex | Cases = 64 (100%) | Cases = 15 (21%) | Cases = 49 (79%) |
| Controls = 47 (100%) | Controls = 39 (52%) | Controls = 8 (48%) | |
| Age (years) | Cases = 45.60 ± 9.89 | Cases = 45.42 ± 9.96 | Cases = 45.64 ± 9.94 |
| Controls = 47 ± 11.52 | Controls = 48.79 ± 10.98 | Controls = 43.87 ± 13.96 | |
| Weight (kg) | 74.42 ± 11.45 | 62.94 ± 0.96 | 77.11 ± 9.83 |
| Alcohol (gr/day) | 265.27 ± 138.32 | 152.32 ± 55.66 | 291.77 ± 138.7 |
| Age of onset | 26.91 ± 10.14 | 29 ± 8.75 | 26.42 ± 10.45 |
| Smoking (cigs/day) | 11.40 ± 15.84 | 9.74 ± 10.47 | 11.79 ± 16.89 |
| Variable | Admission | 2nd week | 3rd week | 4th week | P (adm vs 4th week) |
|---|---|---|---|---|---|
| HDRS | 39.45 ± 6.86 | 31.35 ± 7.11 | 14.5 ± 6.81 | 6.65 ± 5.20 | < 0.001 |
| HARS | 34.44 ± 9.80 | 27.54 ± 8.47 | 14.35 ± 7.11 | 6.71 ± 5.29 | < 0.001 |
| GAS | 46.30 ± 5.06 | 56.7 ± 5.51 | 74.3 ± 8.68 | 83.90 ± 8.27 | < 0.001 |
| Liebowitz Social Anxiety Scale (social fear) | 51 ± 9.45 | – | – | 30.16 ± 6.04 | < 0.001 |
| Liebowitz Social Anxiety Scale (social avoidance) | 54.91 ± 11.04 | – | – | 31.89 ± 7.63 | < 0.001 |
| Mark & Mathews Scale | 49.30 ± 11.59 | – | – | 18.86 ± 5.91 | < 0.001 |
| Leyton Obsessional Inventory | 14.5 ± 3.34 | – | – | 10.04 ± 3.17 | < 0.001 |
| Pilowski scale | 9.14 ± 2.72 | – | – | 4.92 ± 2.16 | < 0.001 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Drago, A.; Liappas, I.; Petio, C.; Albani, D.; Forloni, G.; Malitas, P.; Piperi, C.; Politis, A.; Tzavellas, E.O.; Zisaki, K.K.; et al. Epistasis between IL1A, IL1B, TNF, HTR2A, 5-HTTLPR and TPH2 Variations Does Not Impact Alcohol Dependence Disorder Features. Int. J. Environ. Res. Public Health 2009, 6, 1980-1990. https://doi.org/10.3390/ijerph6071980
Drago A, Liappas I, Petio C, Albani D, Forloni G, Malitas P, Piperi C, Politis A, Tzavellas EO, Zisaki KK, et al. Epistasis between IL1A, IL1B, TNF, HTR2A, 5-HTTLPR and TPH2 Variations Does Not Impact Alcohol Dependence Disorder Features. International Journal of Environmental Research and Public Health. 2009; 6(7):1980-1990. https://doi.org/10.3390/ijerph6071980
Chicago/Turabian StyleDrago, Antonio, Ioannis Liappas, Carmine Petio, Diego Albani, Gianluigi Forloni, Petros Malitas, Christina Piperi, Antonis Politis, Elias O. Tzavellas, Katerina K. Zisaki, and et al. 2009. "Epistasis between IL1A, IL1B, TNF, HTR2A, 5-HTTLPR and TPH2 Variations Does Not Impact Alcohol Dependence Disorder Features" International Journal of Environmental Research and Public Health 6, no. 7: 1980-1990. https://doi.org/10.3390/ijerph6071980