Physiological and Biochemical Responses of Pearl Millet (Pennisetum glaucum L.) Seedlings Exposed to Silver Nitrate (AgNO3) and Silver Nanoparticles (AgNPs)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sliver Nanoparticles and Silver Nitrate
2.2. Plant Material and Sterilization
2.3. Growth Parameters
2.4. Estimation of Photosynthetic Pigments
2.5. Biochemical Profiling
2.5.1. Hydrogen Peroxide Content and Lipid Peroxidation
2.5.2. Proline Content
2.5.3. Superoxide Dismutase (SOD) Activity
2.5.4. Estimation of Catalases, Peroxidases, Ascorbate Peroxidase Activities, and Protein Contents
2.5.5. Glutathion Reductase (GR) and Guaiacol Peroxidase (GPX) Activity
2.5.6. Estimation of Total Phenolic Content (TPC)
2.5.7. Estimation of Total Flavonoids Content (TFC)
2.6. Experimental Design and Statistical Analysis
3. Results
3.1. Growth Parameters
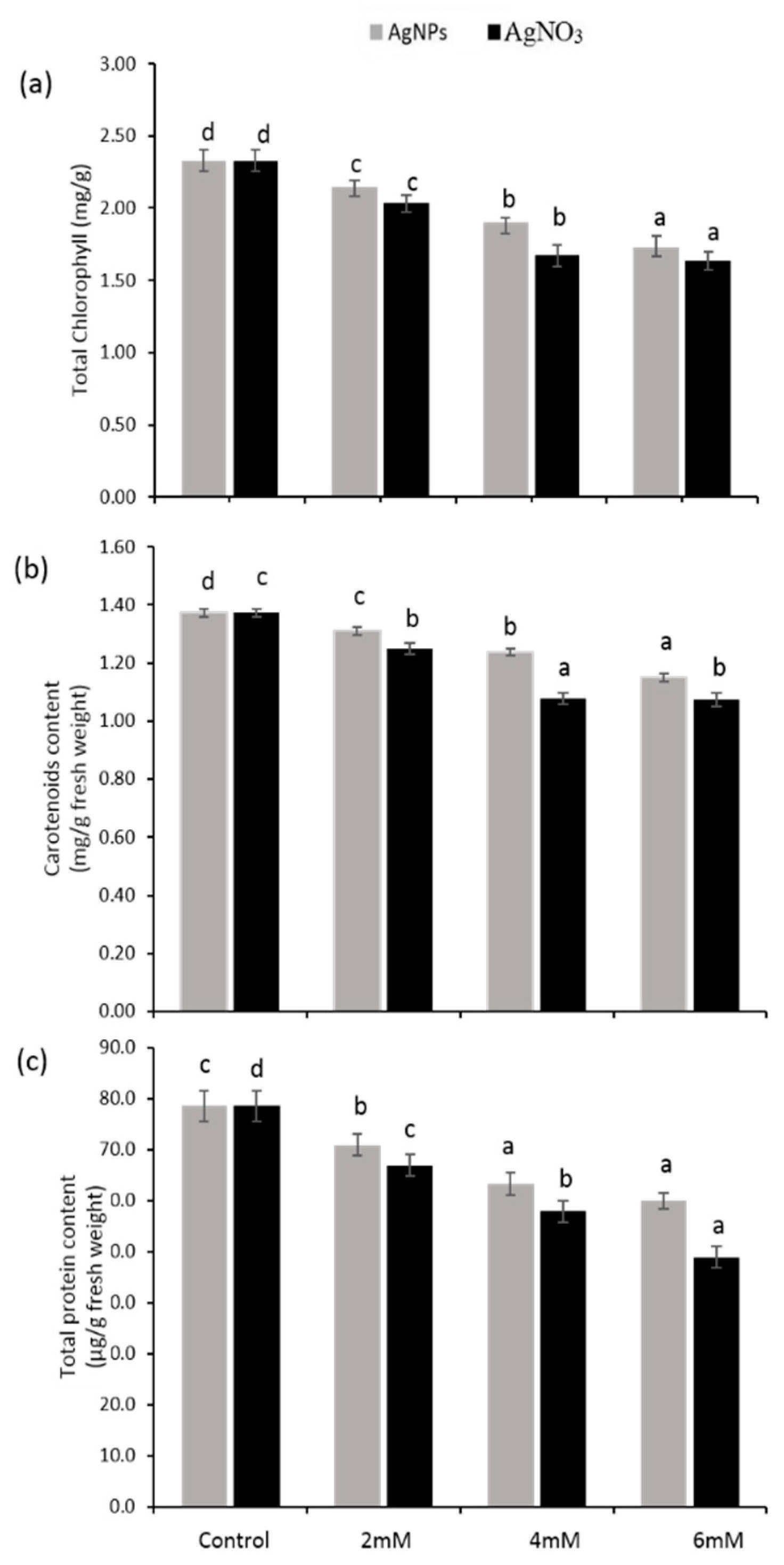
3.2. Photosynthetic Pigments and Protein Contents
3.3. Oxidative Damage
3.4. Lipid Peroxidation as MDA Contents
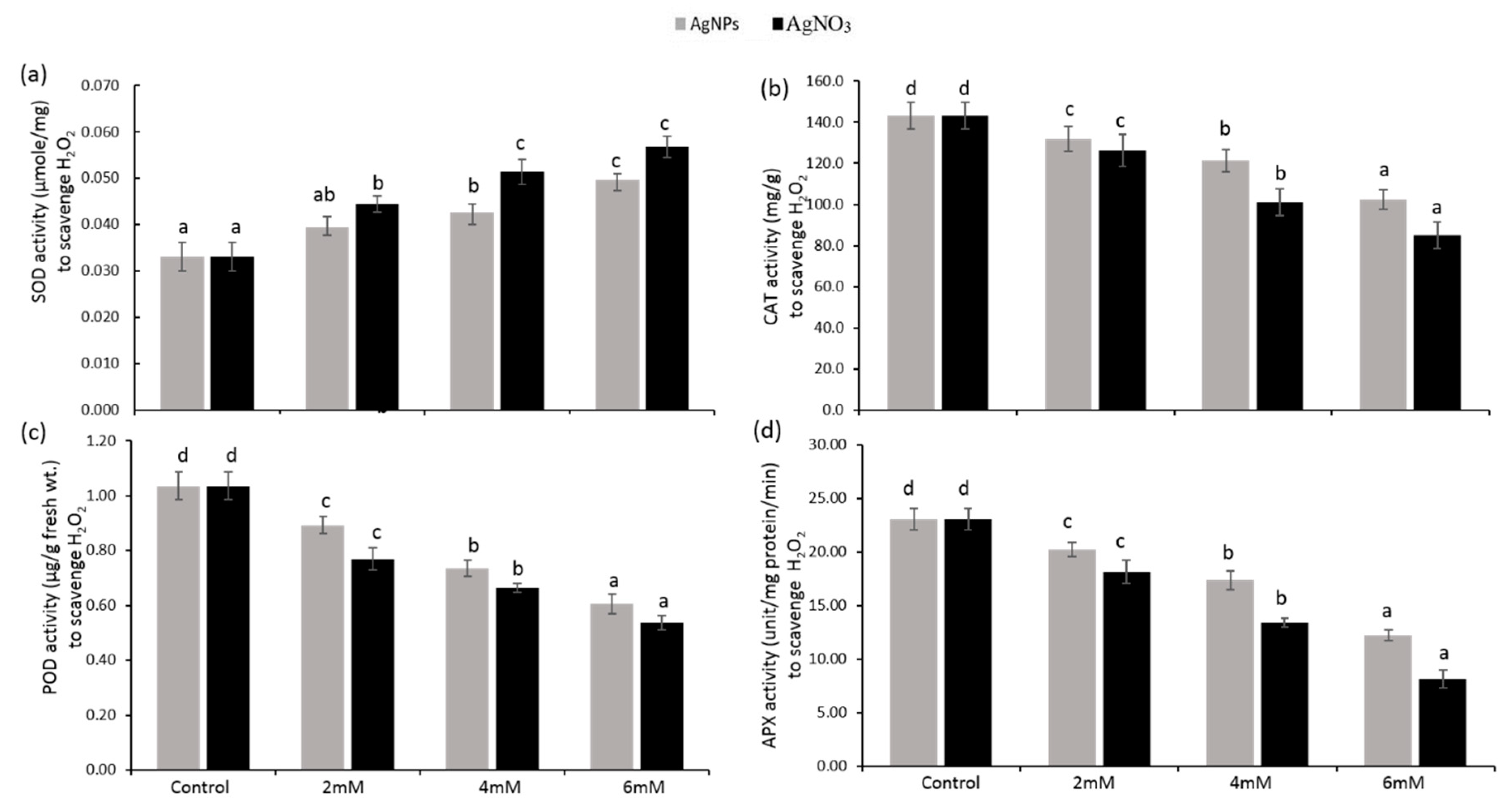
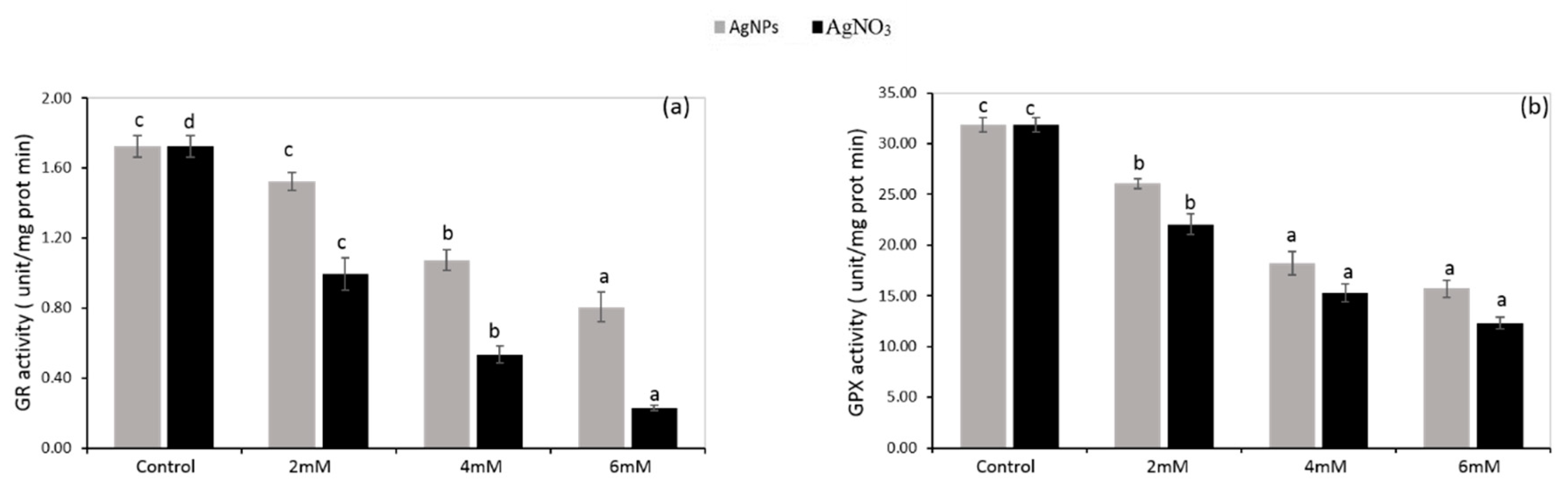
3.5. Enzymatic Antioxidants
3.6. Non-Enzymatic Antioxidants
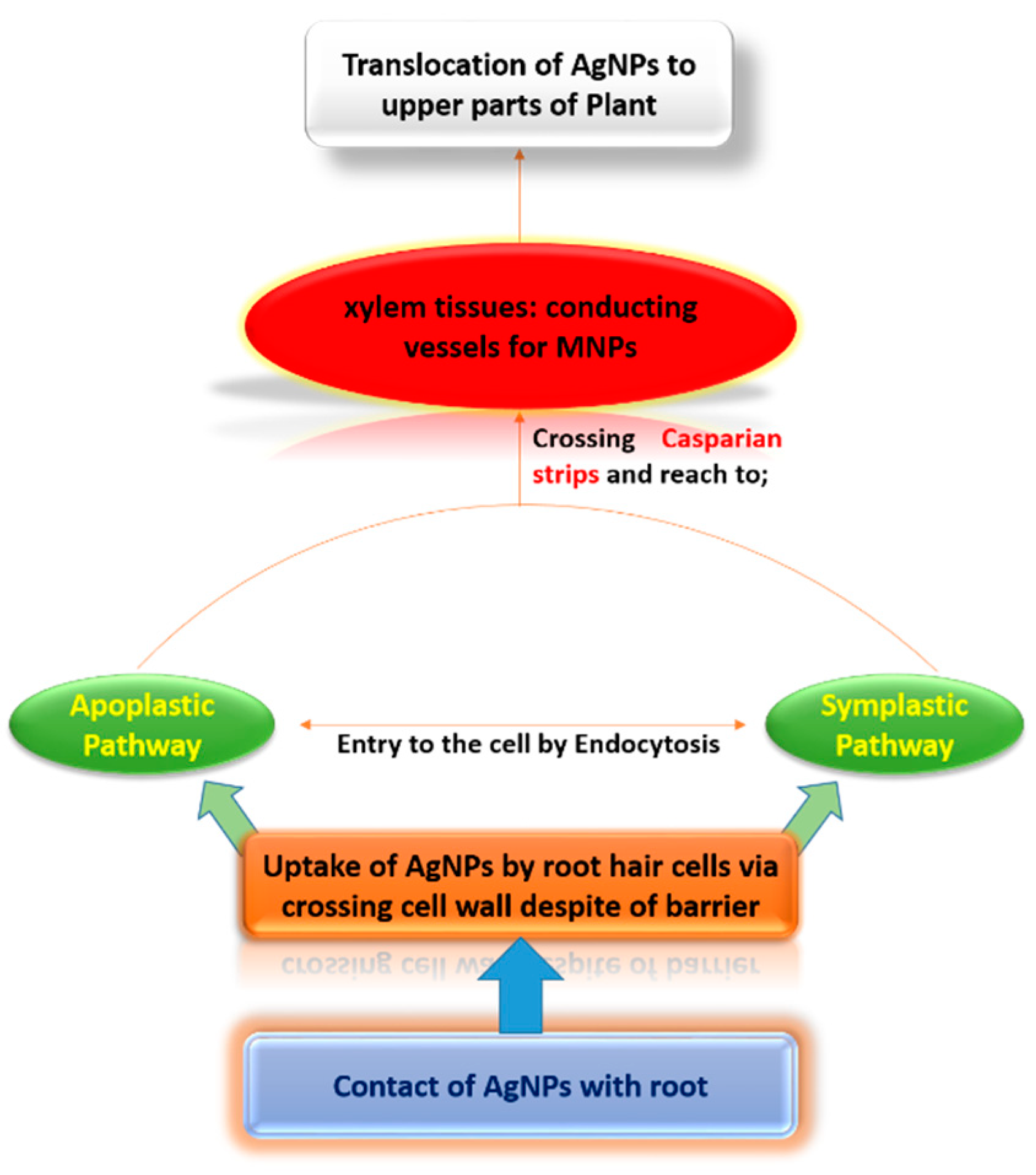
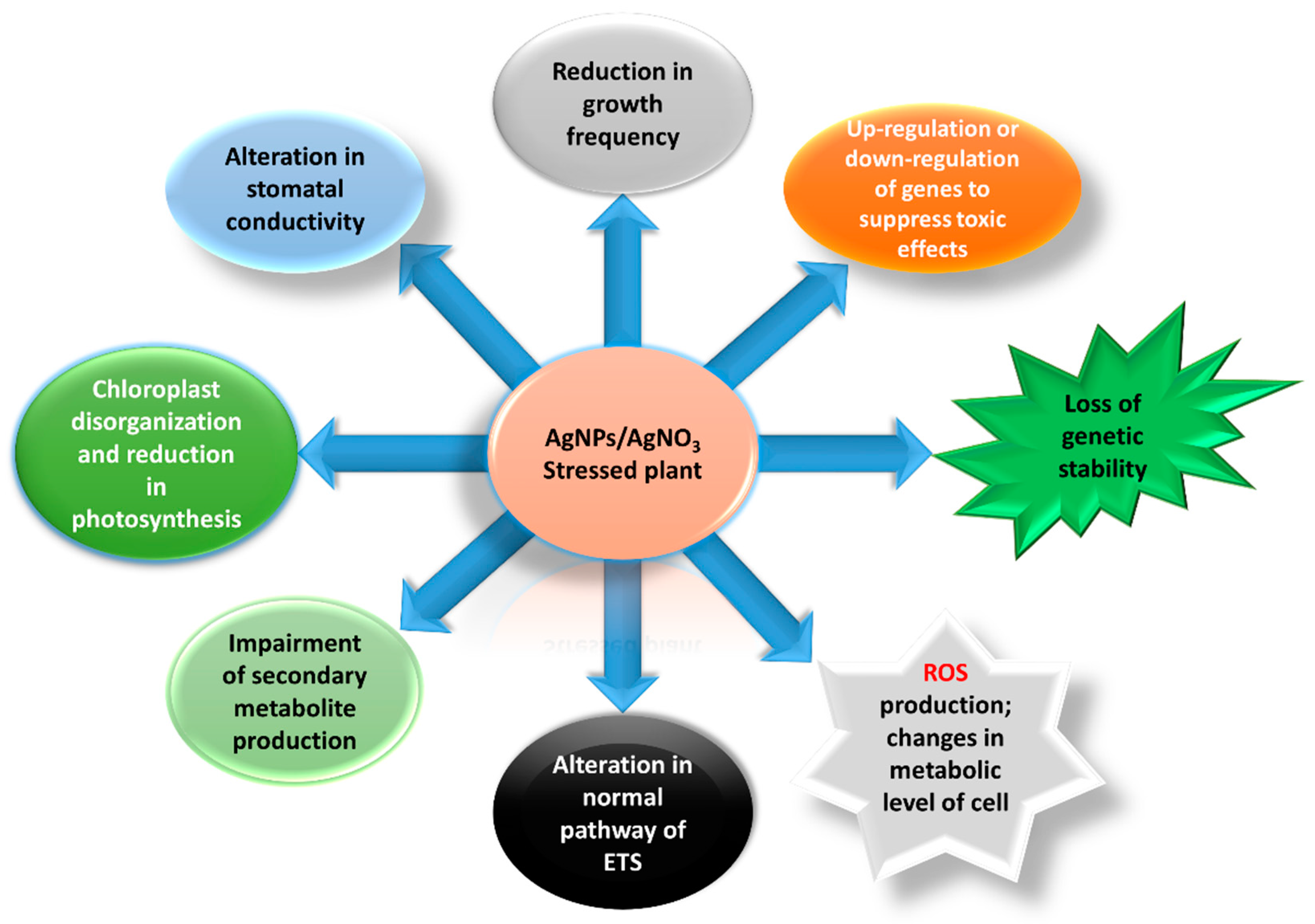
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Basavaraj, G.; Rao, P.P.; Bhagavatula, S.; Ahmed, W. Availability and utilization of pearl millet in India. Available online: http://ejournal.icrisat.org/Volume8/IMPI/Availability_and_utilization.pdf (accessed on 26 June 2019).
- Zhou, S.; Wang, C.; Yin, G.; Zhang, Y.; Shen, X.; Pennerman, K.K.; Zhang, J.; Yan, H.; Zhang, C.; Zhang, X. Phylogenetics and diversity analysis of Pennisetum species using Hemarthria EST-SSR markers. Grassl. Sci. 2019, 65, 13–22. [Google Scholar] [CrossRef]
- Badi, S.; Hoseney, R. Use of sorghum and pearl millet flours in cookies. Cereal Chem. 1976, 53, 733–738. [Google Scholar]
- Thiombiano, L.; Meshack, M. Scaling up Conservation Agriculture in Africa: Strategies and Approaches; The FAO Subregional Office for Eastern Africa: Kirkos Sub-city, Ethiopia, 2009. [Google Scholar]
- Wang, C.; Yan, H.; Li, J.; Zhou, S.; Liu, T.; Zhang, X.; Huang, L. Genome survey sequencing of purple elephant grass (Pennisetum purpureum Schum ‘Zise’) and identification of its SSR markers. Mol. Breed. 2018, 38, 94. [Google Scholar] [CrossRef]
- Hakeem, K.R.; Jawaid, M.; Alothman, O.Y. Agricultural Biomass Based Potential Materials; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Austin, L.A.; Mackey, M.A.; Dreaden, E.C.; El-Sayed, M.A. The optical, photothermal, and facile surface chemical properties of gold and silver nanoparticles in biodiagnostics, therapy, and drug delivery. Arch. Toxicol. 2014, 88, 1391–1417. [Google Scholar] [CrossRef] [Green Version]
- Majdalawieh, A.; Kanan, M.C.; El-Kadri, O.; Kanan, S.M. Recent advances in gold and silver nanoparticles: Synthesis and applications. J. Nanosci. Nanotechnol. 2014, 14, 4757–4780. [Google Scholar] [CrossRef] [PubMed]
- Siripattanakul-Ratpukdi, S.; Fürhacker, M. Issues of silver nanoparticles in engineered environmental treatment systems. Water Air Soil Pollut. 2014, 225, 1939. [Google Scholar] [CrossRef]
- Almutairi, Z.M.; Alharbi, A. Effect of silver nanoparticles on seed germination of crop plants. J. Adv. Agric. 2015, 4, 283–288. [Google Scholar] [CrossRef]
- Mueller, N.C.; Nowack, B. Exposure modeling of engineered nanoparticles in the environment. Environ. Sci. Technol. 2008, 42, 4447–4453. [Google Scholar] [CrossRef]
- Bouwmeester, H.; Dekkers, S.; Noordam, M.Y.; Hagens, W.I.; Bulder, A.S.; De Heer, C.; Ten Voorde, S.E.; Wijnhoven, S.W.; Marvin, H.J.; Sips, A.J. Review of health safety aspects of nanotechnologies in food production. Regul. Toxicol. Pharmacol. 2009, 53, 52–62. [Google Scholar] [CrossRef]
- Kaegi, R.; Sinnet, B.; Zuleeg, S.; Hagendorfer, H.; Mueller, E.; Vonbank, R.; Boller, M.; Burkhardt, M. Release of silver nanoparticles from outdoor facades. Environ. Pollut. 2010, 158, 2900–2905. [Google Scholar] [CrossRef]
- Tripathi, A.; Liu, S.; Singh, P.K.; Kumar, N.; Pandey, A.C.; Tripathi, D.K.; Chauhan, D.K.; Sahi, S. Differential phytotoxic responses of silver nitrate (AgNO3) and silver nanoparticle (AgNps) in Cucumis sativus L. Plant Gene 2017, 11, 255–264. [Google Scholar] [CrossRef]
- Yin, L.; Colman, B.P.; McGill, B.M.; Wright, J.P.; Bernhardt, E.S. Effects of silver nanoparticle exposure on germination and early growth of eleven wetland plants. PLoS ONE 2012, 7, e47674. [Google Scholar] [CrossRef] [PubMed]
- García-López, J.; Zavala-García, F.; Olivares-Sáenz, E.; Lira-Saldívar, R.; Díaz Barriga-Castro, E.; Ruiz-Torres, N.; Ramos-Cortez, E.; Vázquez-Alvarado, R.; Niño-Medina, G. Zinc Oxide Nanoparticles Boosts Phenolic Compounds and Antioxidant Activity of Capsicum annuum L. during Germination. Agronomy 2018, 8, 215. [Google Scholar] [CrossRef]
- Haverkamp, R.; Marshall, A. The mechanism of metal nanoparticle formation in plants: Limits on accumulation. J. Nanoparticle Res. 2009, 11, 1453–1463. [Google Scholar] [CrossRef]
- Ejaz, M.; Raja, N.I.; Ahmad, M.S.; Hussain, M.; Iqbal, M. Effect of silver nanoparticles and silver nitrate on growth of rice under biotic stress. IET Nanobiotechnol. 2018, 12, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Tripathi, A.; Singh, S.; Singh, Y.; Vishwakarma, K.; Yadav, G.; Sharma, S.; Singh, V.K.; Mishra, R.K.; Upadhyay, R. Uptake, accumulation and toxicity of silver nanoparticle in autotrophic plants, and heterotrophic microbes: A concentric review. Front. Microbiol. 2017, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Rahmatpour, S.; Shirvani, M.; Mosaddeghi, M.R.; Nourbakhsh, F.; Bazarganipour, M. Dose–response effects of silver nanoparticles and silver nitrate on microbial and enzyme activities in calcareous soils. Geoderma 2017, 285, 313–322. [Google Scholar] [CrossRef]
- Dwivedi, R.; Singh, V.P.; Kumar, J.; Prasad, S.M. Differential physiological and biochemical responses of two Vigna species under enhanced UV-B radiation. J. Radiat. Res. Appl. Sci. 2015, 8, 173–181. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Sagisaka, S. The occurrence of peroxide in a perennial plant, Populus gelrica. Plant Physiol. 1976, 57, 308–309. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ullah, N.; Haq, I.U.; Safdar, N.; Mirza, B. Physiological and biochemical mechanisms of allelopathy mediated by the allelochemical extracts of Phytolacca latbenia (Moq.) H. Walter. Toxicol. Ind. Health 2015, 31, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. [3] Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Chance, B.; Maehly, A. [136] Assay of Catalases and Peroxidases; Elsevier: Amsterdam, The Netherlands, 1955. [Google Scholar]
- Klapheck, S.; Zimmer, I.; Cosse, H. Scavenging of hydrogen peroxide in the endosperm of Ricinus communis by ascorbate peroxidase. Plant Cell Physiol. 1990, 31, 1005–1013. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Smith, I.K.; Vierheller, T.L.; Thorne, C.A. Assay of glutathione reductase in crude tissue homogenates using 5, 5′-dithiobis (2-nitrobenzoic acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef]
- Velioglu, Y.; Mazza, G.; Gao, L.; Oomah, B. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Lopez-Contreras, J.J.; Zavala-Garcia, F.; Urias-Orona, V.; Martinez-Avila, G.C.G.; Rojas, R.; Guillermo, N.-M. Chromatic, phenolic and antioxidant properties of Sorghum bicolor genotypes. Not. Bot. Horti Agrobot. Cluj-Napoca 2015, 43, 366–370. [Google Scholar]
- Harris, A.T.; Bali, R. On the formation and extent of uptake of silver nanoparticles by live plants. J. Nanoparticle Res. 2008, 10, 691–695. [Google Scholar] [CrossRef]
- Fayez, K.; El-Deeb, B.; Mostafa, N. Toxicity of biosynthetic silver nanoparticles on the growth, cell ultrastructure and physiological activities of barley plant. Acta Physiol. Plant. 2017, 39, 155. [Google Scholar] [CrossRef]
- Parveen, A.; Rao, S. Cytotoxicity and genotoxicity of biosynthesized gold and silver nanoparticles on human cancer cell lines. J. Clust. Sci. 2015, 26, 775–788. [Google Scholar] [CrossRef]
- Shams, G.; Ranjbar, M.; Amiri, A. Effect of silver nanoparticles on concentration of silver heavy element and growth indexes in cucumber (Cucumis sativus L. negeen). J. Nanoparticle Res. 2013, 15, 1630. [Google Scholar] [CrossRef]
- Sarabi, M.; Safipour Afshar, A.; Mahmoodzadeh, H. Physiological Analysis of Silver Nanoparticles and AgNO3 Effect to Brassica napus L. J. Chem. Health Risks 2015, 5, 285–294. [Google Scholar]
- Yasur, J.; Rani, P.U. Environmental effects of nanosilver: Impact on castor seed germination, seedling growth, and plant physiology. Environ. Sci. Pollut. Res. 2013, 20, 8636–8648. [Google Scholar] [CrossRef] [PubMed]
- Krizkova, S.; Ryant, P.; Krystofova, O.; Adam, V.; Galiova, M.; Beklova, M.; Babula, P.; Kaiser, J.; Novotny, K.; Novotny, J. Multi-instrumental analysis of tissues of sunflower plants treated with silver (I) ions–plants as bioindicators of environmental pollution. Sensors 2008, 8, 445–463. [Google Scholar] [CrossRef]
- Pandey, C.; Khan, E.; Mishra, A.; Sardar, M.; Gupta, M. Silver nanoparticles and its effect on seed germination and physiology in Brassica juncea L.(Indian mustard) plant. Adv. Sci. Lett. 2014, 20, 1673–1676. [Google Scholar] [CrossRef]
- Sharma, P.; Bhatt, D.; Zaidi, M.; Saradhi, P.P.; Khanna, P.; Arora, S. Silver nanoparticle-mediated enhancement in growth and antioxidant status of Brassica juncea. Appl. Biochem. Biotechnol. 2012, 167, 2225–2233. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Upadhyay, N.; Singh, J.; Liu, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Tripathi, D.K.; Sharma, S. Differential Phytotoxic Impact of Plant Mediated Silver Nanoparticles (AgNPs) and Silver Nitrate (AgNO3) on Brassica sp. Front. Plant Sci. 2017, 8, 1501. [Google Scholar] [CrossRef]
- Genty, B.; Harbinson, J.; Briantais, J.-M.; Baker, N.R. The relationship between non-photochemical quenching of chlorophyll fluorescence and the rate of photosystem 2 photochemistry in leaves. Photosynth. Res. 1990, 25, 249–257. [Google Scholar] [CrossRef]
- Jiang, H.S.; Yin, L.Y.; Ren, N.N.; Zhao, S.T.; Li, Z.; Zhi, Y.; Shao, H.; Li, W.; Gontero, B. Silver nanoparticles induced reactive oxygen species via photosynthetic energy transport imbalance in an aquatic plant. Nanotoxicology 2017, 11, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Atli, G.; Alptekin, Ö.; Tükel, S.; Canli, M. Response of catalase activity to Ag+, Cd2+, Cr6+, Cu2+ and Zn2+ in five tissues of freshwater fish Oreochromis niloticus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 143, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Mehrian, S.K.; Heidari, R.; Rahmani, F. Effect of silver nanoparticles on free amino acids content and antioxidant defense system of tomato plants. Indian J. Plant Physiol. 2015, 20, 257–263. [Google Scholar] [CrossRef]
- Mehta, C.; Srivastava, R.; Arora, S.; Sharma, A. Impact assessment of silver nanoparticles on plant growth and soil bacterial diversity. 3 Biotech 2016, 6, 254. [Google Scholar]
- Zancan, S.; Suglia, I.; La Rocca, N.; Ghisi, R. Effects of UV-B radiation on antioxidant parameters of iron-deficient barley plants. Environ. Exp. Bot. 2008, 63, 71–79. [Google Scholar] [CrossRef]
- Zhou, S.; Wang, C.; Frazier, T.P.; Yan, H.; Chen, P.; Chen, Z.; Huang, L.; Zhang, X.; Peng, Y.; Ma, X. The first Illumina-based de novo transcriptome analysis and molecular marker development in Napier grass (Pennisetum purpureum). Mol. Breed. 2018, 38, 95. [Google Scholar] [CrossRef]
- Cekić, S.; Zlatanović, G.; Cvetković, T.; Petrović, B. Oxidative stress in cataractogenesis. Bosn. J. Basic Med. Sci. 2010, 10, 265. [Google Scholar] [CrossRef] [PubMed]
- Krishnaraj, C.; Jagan, E.; Ramachandran, R.; Abirami, S.; Mohan, N.; Kalaichelvan, P. Effect of biologically synthesized silver nanoparticles on Bacopa monnieri (Linn.) Wettst. plant growth metabolism. Process Biochem. 2012, 47, 651–658. [Google Scholar] [CrossRef]
- Hegedüs, A.; Erdei, S.; Horváth, G. Comparative studies of H2O2 detoxifying enzymes in green and greening barley seedlings under cadmium stress. Plant Sci. 2001, 160, 1085–1093. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, J.; Wu, Q.; Huang, Y. Effects of Silver Nanoparticles on Seed Germination and Seedling Growth of Radish (Raphanus sativus L.). In Proceedings of the 2nd International Conference on Civil, Materials and Environmental Sciences, London, UK, 13–14 March 2015; Atlantis Press: London, UK, 2015. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Tripathi, D.; Yadav, S.; Kalaji, H.; Brestic, M. Phytotoxic effect of silver nanoparticles in Triticum aestivum: Improper regulation of photosystem I activity as the reason for oxidative damage in the chloroplast. Photosynthetica 2019, 57, 209–216. [Google Scholar] [CrossRef]
- Oukarroum, A.; Barhoumi, L.; Pirastru, L.; Dewez, D. Silver nanoparticle toxicity effect on growth and cellular viability of the aquatic plant Lemna gibba. Environ. Toxicol. Chem. 2013, 32, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Li, P.; Huang, Q.; Zhang, H. The different response mechanisms of Wolffia globosa: Light-induced silver nanoparticle toxicity. Aquat. Toxicol. 2016, 176, 97–105. [Google Scholar] [CrossRef] [PubMed]
- McShan, D.; Ray, P.C.; Yu, H. Molecular toxicity mechanism of nanosilver. J. Food Drug Anal. 2014, 22, 116–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuesombat, P.; Hannongbua, S.; Akasit, S.; Chadchawan, S. Effect of silver nanoparticles on rice (Oryza sativa L. cv. KDML 105) seed germination and seedling growth. Ecotoxicol. Environ. Saf. 2014, 104, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Mahdavian, K.; Ghorbanli, M.; Kalantari, K.M. The Effects of Ultraviolet Radiation on the Contents of Chlorophyll, Flavonoid, Anthocyanin and Proline in Capsicum annuum L. Turk. J. Bot. 2008, 32, 25–33. [Google Scholar]
- Kishor, P.K.; Sangam, S.; Amrutha, R.; Laxmi, P.S.; Naidu, K.; Rao, K.; Rao, S.; Reddy, K.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | AgNPs | AgNO3 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Growth Parameters | ||||||||||
| RL | SL | FW | DW | RWC | RL | SL | FW | DW | RWC | |
| Control | 12.37 ± 0.96 c | 7.47 ± 0.23 c | 0.131 ± 0.006 c | 0.020 ± 0.0005 d | 95.17 ± 0.79 d | 12.37 ± 0.96 c | 7.47 ± 0.23 c | 0.131 ± 0.006 c | 0.020 ± 0.0005 d | 95.17 ± 0.79 d |
| 2 mM | 8.87 ± 0.99 (28) b | 6.33 ± 0.26 (15) b | 0.112 ± 0.003 (14) b | 0.018 ± 0.0001(10) c | 90.01 ± 1.47 (10) c | 7.27 ± 0.62 (41) b | 5.87 ± 0.22 c (21) b | 0.081 ± 0.003 (38) b | 0.0164 ± 0.0001 (20) c | 83.02 ± 1.68 (13) c |
| 4 mM | 7.20 ± 0.45 (41) bc | 5.90 ± 0.23 (21) b | 0.075 ± 0.002 (43) a | 0.016 ± 0.0002 (20) b | 80.54 ± 1.83 (15) b | 5.17 ± 0.33 (58) a | 5.10 ± 0.20c (32) a | 0.068 ± 0.001 (48) ab | 0.0143 ± 0.0002 c (30) b | 72.04 ± 2.16 (24) b |
| 6 Mm | 4.80 ± 0.35 (61) a | 5.13 ± 0.18 (31) a | 0.069 ± 0.002 (47) a | 0.0132 ± 0.0002 (35)a | 70.28 ± 1.59 (26) a | 3.90 ± 0.21 (68) a | 4.77 ± 0.14 c (36) a | 0.058 ± 0.002(55) a | 0.0114 ± 0.0003 c (45)a | 63.10 ± 1.49 (28) a |
| Treatment | AgNPs | AgNO3 | ||||
|---|---|---|---|---|---|---|
| Non-Enzymatic Antioxidants | ||||||
| Proline | TFCs | TPCs | Proline | TFCs | TPCs | |
| Control | 1.22 ± 0.037 a | 2.06 ± 0.043 (41) d | 1.56 ± 0.032 a | 1.22 ± 0.037 a | 0.59 ± 0.023 a | 1.22 ± 0.023 (52) d |
| 2 mM | 1.59 ± 0.027 (23) b | 0.71 ± 0.022 (17) b | 1.75 ± 0.021 (11) b | 1.72 ± 0.032 (29) b | 0.84 ± 0.015 (30) b | 1.84 ± 0.021(15) b |
| 4 Mm | 1.83 ± 0.026 (33) c | 0.85 ± 0.020 (30) c | 1.92 ± 0.024 (19) c | 1.93 ± 0.026 (37) c | 0.94 ± 0.017 (37) c | 2.00 ± 0.055 (22) c |
| 6 Mm | 2.06 ± 0.043 (41) d | 1.13 ± 0.023 (48) d | 2.32 ± 0.028 (33) d | 2.16 ± 0.023 (44) d | 1.22 ± 0.023 (52) d | 2.53 ± 0.026 (38) d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, I.; Raza, M.A.; Khalid, M.H.B.; Awan, S.A.; Raja, N.I.; Zhang, X.; Min, S.; Wu, B.C.; Hassan, M.J.; Huang, L. Physiological and Biochemical Responses of Pearl Millet (Pennisetum glaucum L.) Seedlings Exposed to Silver Nitrate (AgNO3) and Silver Nanoparticles (AgNPs). Int. J. Environ. Res. Public Health 2019, 16, 2261. https://doi.org/10.3390/ijerph16132261
Khan I, Raza MA, Khalid MHB, Awan SA, Raja NI, Zhang X, Min S, Wu BC, Hassan MJ, Huang L. Physiological and Biochemical Responses of Pearl Millet (Pennisetum glaucum L.) Seedlings Exposed to Silver Nitrate (AgNO3) and Silver Nanoparticles (AgNPs). International Journal of Environmental Research and Public Health. 2019; 16(13):2261. https://doi.org/10.3390/ijerph16132261
Chicago/Turabian StyleKhan, Imran, Muhammad Ali Raza, Muhammad Hayder Bin Khalid, Samrah Afzal Awan, Naveed Iqbal Raja, Xinquan Zhang, Sun Min, Bing Chao Wu, Muhammad Jawad Hassan, and Linkai Huang. 2019. "Physiological and Biochemical Responses of Pearl Millet (Pennisetum glaucum L.) Seedlings Exposed to Silver Nitrate (AgNO3) and Silver Nanoparticles (AgNPs)" International Journal of Environmental Research and Public Health 16, no. 13: 2261. https://doi.org/10.3390/ijerph16132261