
Genomic Evidence for Bacterial Determinants Influencing Obesity Development


Abstract
:1. Introduction
2. Methods
2.1. Collection of Data on Lactobacilli Universal Stress Proteins and Lactobacilli Genomes
2.2. Visual Analytics of Information on Genome Annotation and Protein Annotation
2.3. Evaluation of the Genomic Context of Genes Encoding Universal Stress Proteins
3. Results
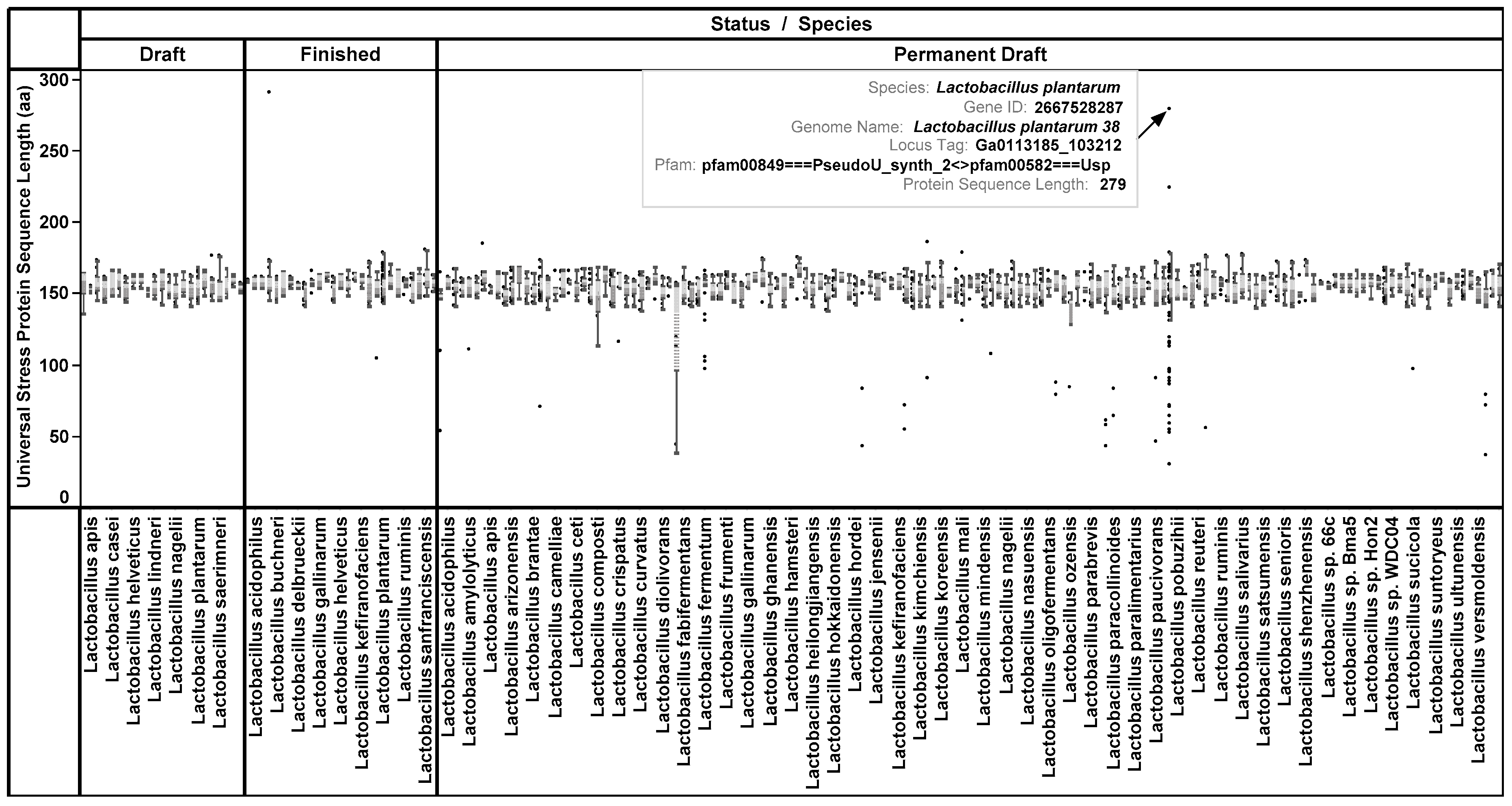
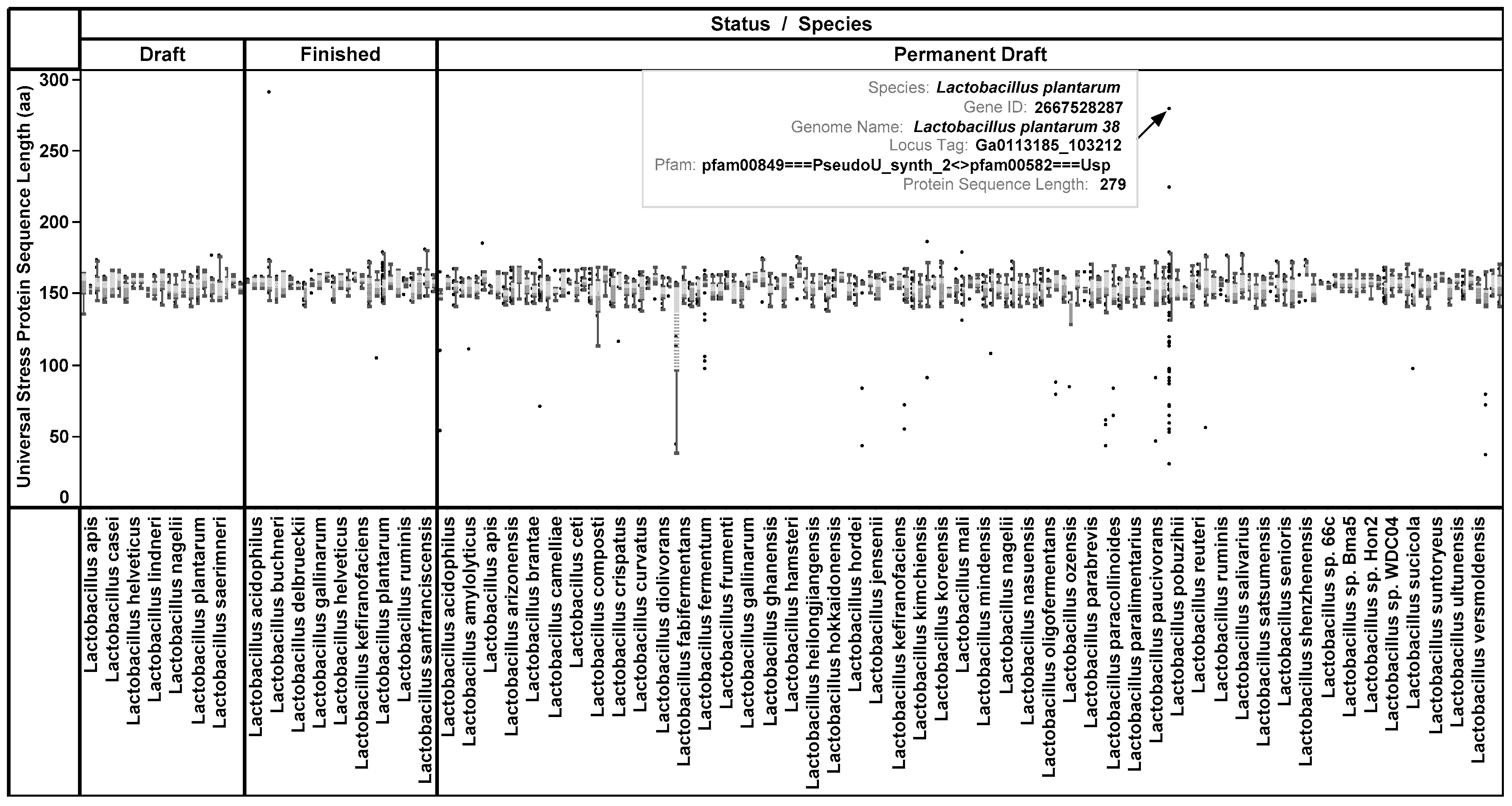
3.1. Protein Length and Protein Domain Architecture
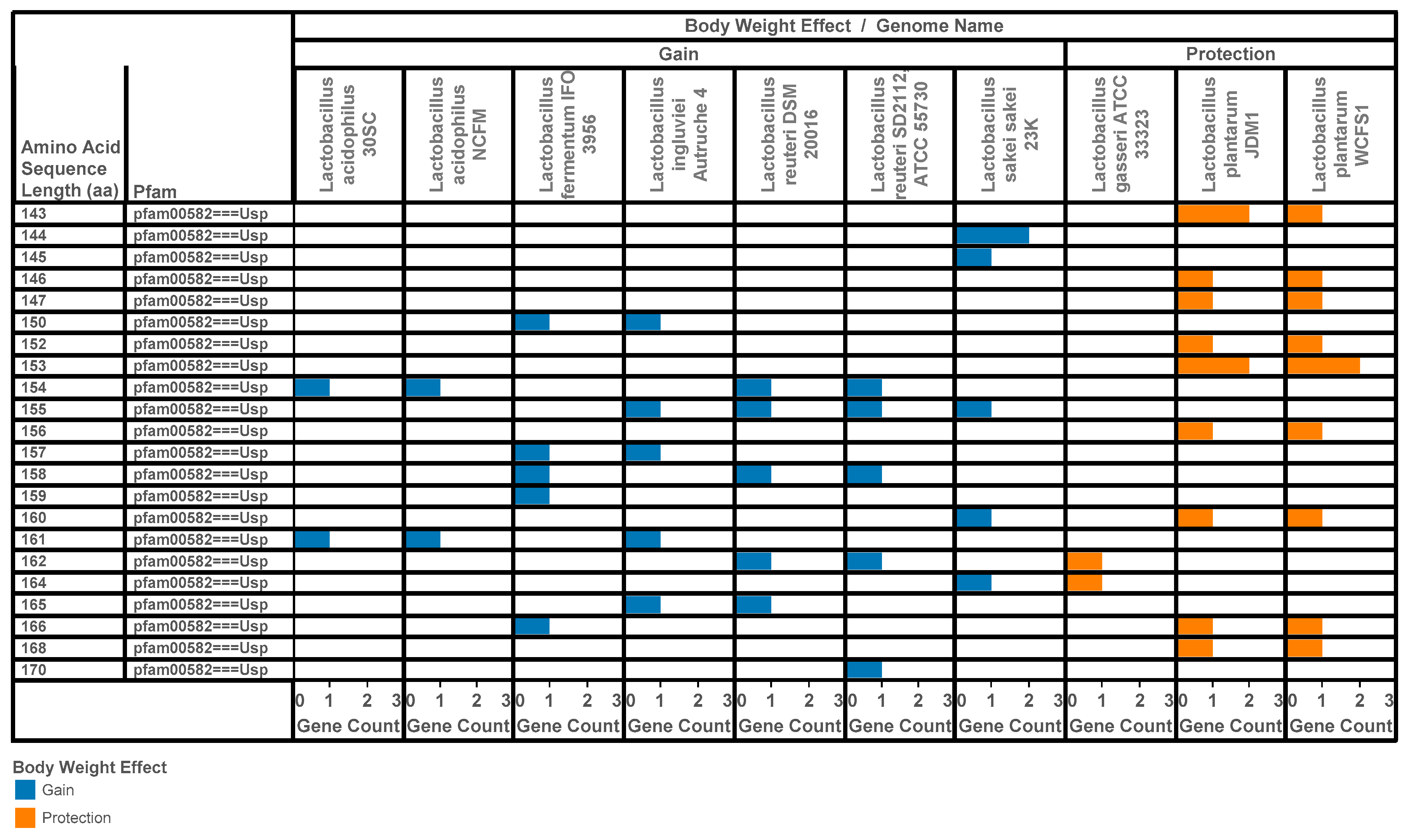
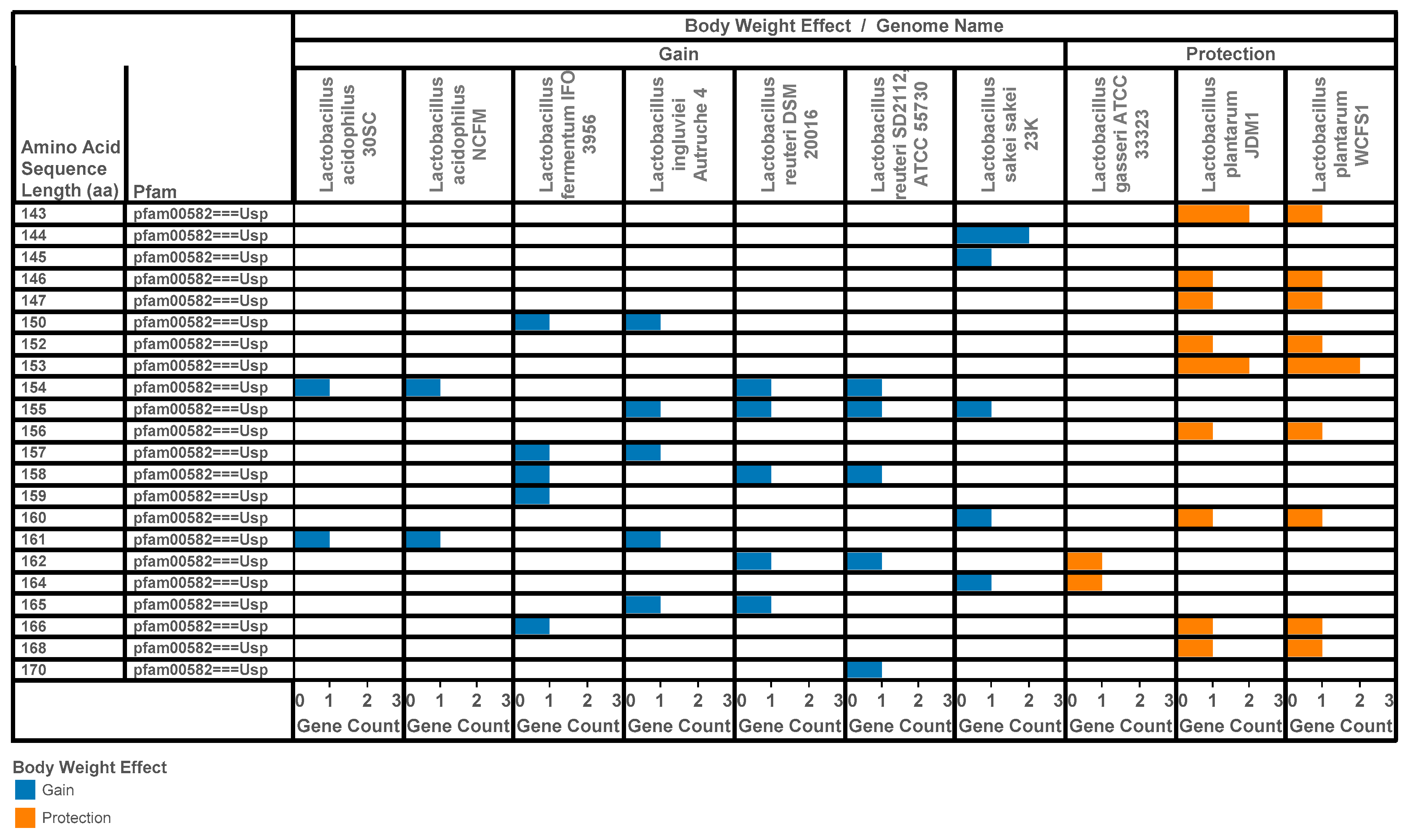
3.2. Identification of Lactobacilli Universal Stress Proteins Encoded in Operons
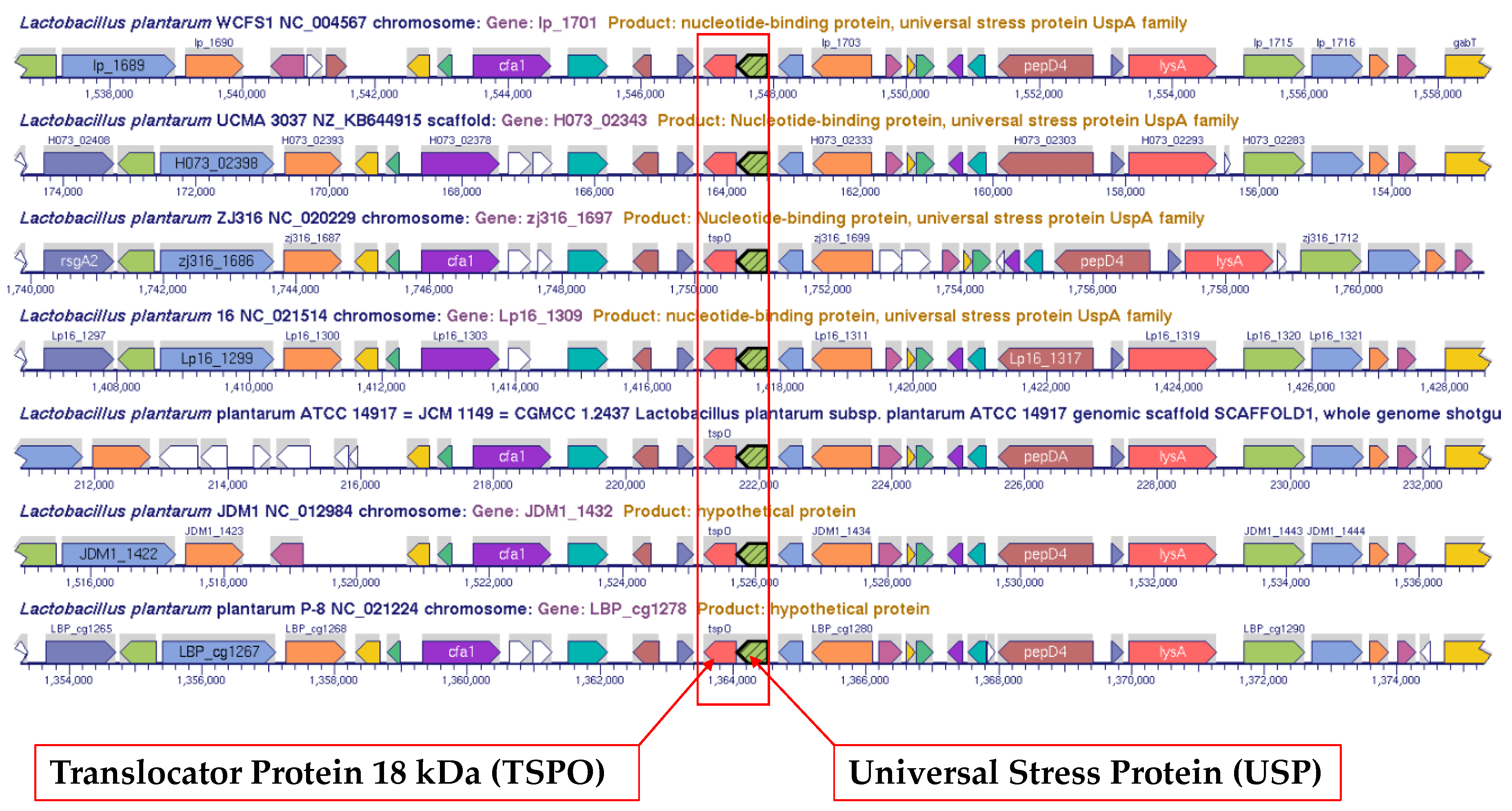
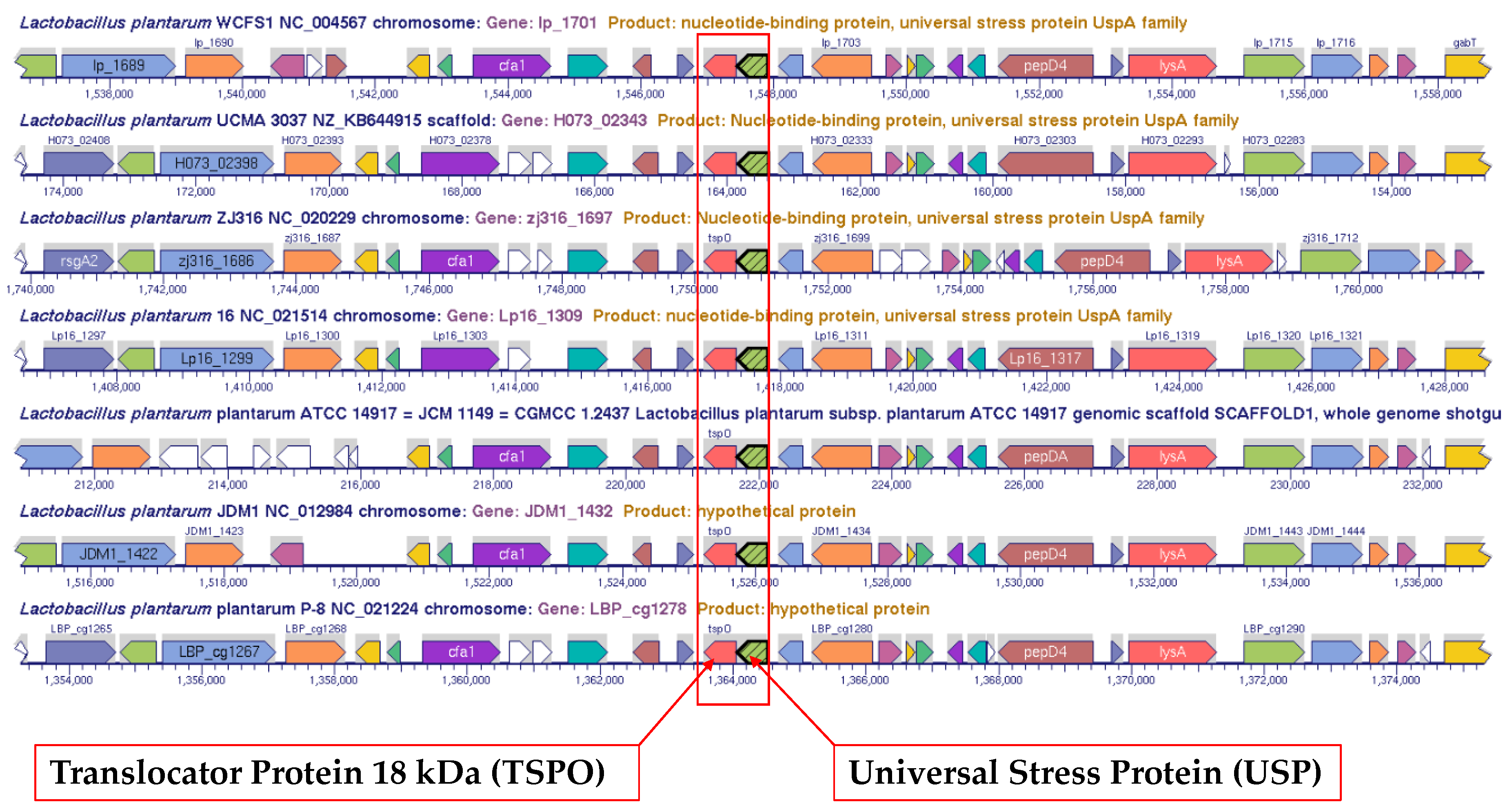
3.3. Alignment of Genomic Regions for Two-Gene Operons that include Gene for Universal Stress Protein
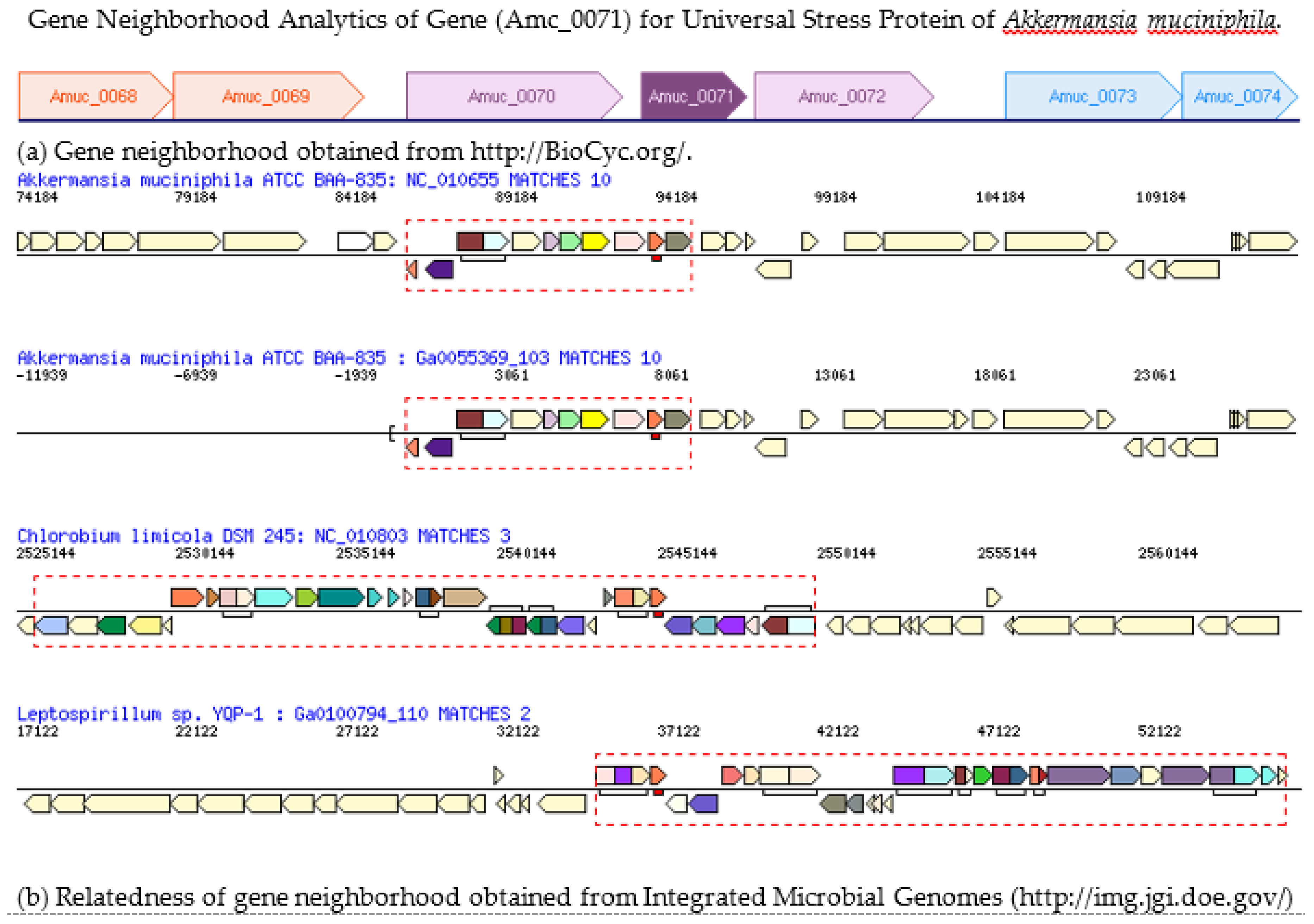
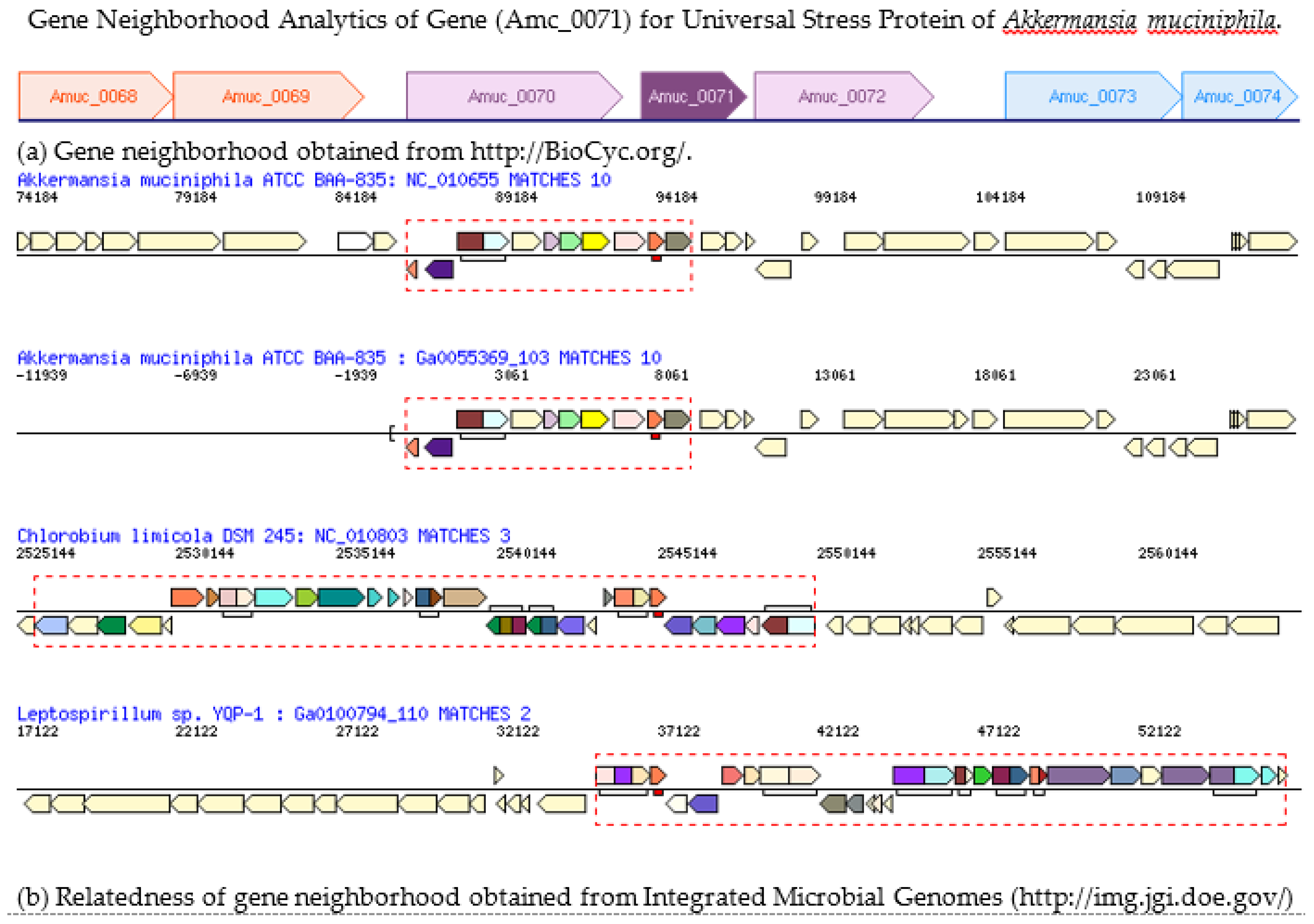
3.4. Gene Neighborhood of Gene for the Universal Stress Protein in Akkermansia muciniphila Genome
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Conte, C.; Cammarota, G.; Haley, A.P.; Styriak, I.; Gaspar, L.; Fusek, J.; Rodrigo, L.; Kruzliak, P. Probiotics in prevention and treatment of obesity: A critical view. Nutr. Metab. 2016, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Ouwerkerk, J.; Aalvink, S.; Belzer, C.; De Vos, W. Preparation and preservation of viable Akkermansia muciniphila cells for therapeutic interventions. Benef. Microbes 2017, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Lara, M.J.; Robles-Sanchez, C.; Ruiz-Ojeda, F.J.; Plaza-Diaz, J.; Gil, A. Effects of probiotics and synbiotics on obesity, insulin resistance syndrome, type 2 diabetes and non-alcoholic fatty liver disease: A review of human clinical trials. Int. J. Mol. Sci. 2016, 17, 928. [Google Scholar] [CrossRef] [PubMed]
- Cardinelli, C.S.; Sala, P.C.; Alves, C.C.; Torrinhas, R.S.; Waitzberg, D.L. Influence of intestinal microbiota on body weight gain: A narrative review of the literature. Obes. Surg. 2015, 25, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Vrieze, A.; Holleman, F.; Zoetendal, E.; De Vos, W.; Hoekstra, J.; Nieuwdorp, M. The environment within: How gut microbiota may influence metabolism and body composition. Diabetologia 2010, 53, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Angelakis, E.; Merhej, V.; Raoult, D. Related actions of probiotics and antibiotics on gut microbiota and weight modification. Lancet Infect. Dis. 2013, 13, 889–899. [Google Scholar] [CrossRef]
- Drissi, F.; Merhej, V.; Angelakis, E.; El Kaoutari, A.; Carriere, F.; Henrissat, B.; Raoult, D. Comparative genomics analysis of Lactobacillus species associated with weight gain or weight protection. Nutr. Diabetes 2014, 4, e109. [Google Scholar] [CrossRef] [PubMed]
- Deng, T.; Lyon, C.J.; Bergin, S.; Caligiuri, M.A.; Hsueh, W.A. Obesity, inflammation, and cancer. Ann. Rev. Pathol. Mech. Dis 2016, 11, 421–449. [Google Scholar] [CrossRef] [PubMed]
- Hubert, H.B.; Feinleib, M.; McNamara, P.M.; Castelli, W.P. Obesity as an independent risk factor for cardiovascular disease: A 26-year follow-up of participants in the Framingham Heart Study. Circulation 1983, 67, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, C.; Wilding, J. Obesity and cardiovascular risk in type 2 diabetes. In Advanced Nutrition and Dietetics in Diabetes; John Wiley & Sons, Ltd: New York, NY, USA, 2015; pp. 119–128. [Google Scholar]
- Lebeer, S.; Verhoeven, T.L.; Vélez, M.P.; Vanderleyden, J.; De Keersmaecker, S.C. Impact of environmental and genetic factors on biofilm formation by the probiotic strain Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 2007, 73, 6768–6775. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.N.; Bergendahl, L.T.; Marsh, J.A. Operon gene order is optimized for ordered protein complex assembly. Cell Rep. 2016, 14, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Isokpehi, R.D.; Udensi, U.K.; Anyanwu, M.N.; Mbah, A.N.; Johnson, M.O.; Edusei, K.; Bauer, M.A.; Hall, R.A.; Awofolu, O.R. Knowledge building insights on biomarkers of arsenic toxicity to keratinocytes and melanocytes. Biomark. Insights 2012, 7, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Isokpehi, R.D.; Udensi, U.K.; Simmons, S.S.; Hollman, A.L.; Cain, A.E.; Olofinsae, S.A.; Hassan, O.A.; Kashim, Z.A.; Enejoh, O.A.; Fasesan, D.E. Evaluative profiling of arsenic sensing and regulatory systems in the human microbiome project genomes. Microbiol. Insights 2014, 7, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.S.; Isokpehi, R.D.; Mbah, A.N.; Hollman, A.L.; Bernard, C.O.; Simmons, S.S.; Ayensu, W.K.; Garner, B.L. Functional annotation analytics of Bacillus genomes reveals stress responsive acetate utilization and sulfate uptake in the biotechnologically relevant Bacillus megaterium. Bioinform. Biol. Insights 2012, 6, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Gury, J.; Seraut, H.; Tran, N.P.; Barthelmebs, L.; Weidmann, S.; Gervais, P.; Cavin, J.-F. Inactivation of PadR, the repressor of the phenolic acid stress response, by molecular interaction with Usp1, a universal stress protein from Lactobacillus plantarum, in Escherichia coli. Appl. Environ. Microbiol. 2009, 75, 5273–5283. [Google Scholar] [CrossRef] [PubMed]
- Licandro-Seraut, H.; Gury, J.; Tran, N.P.; Barthelmebs, L.; Cavin, J.-F. Kinetics and intensity of the expression of genes involved in the stress response tightly induced by phenolic acids in Lactobacillus plantarum. J. Mol. Microbiol. Biotechnol. 2007, 14, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Fong, J.H.; Geer, L.Y.; Panchenko, A.R.; Bryant, S.H. Modeling the evolution of protein domain architectures using maximum parsimony. J. Mol. Biol. 2007, 366, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; Gwadz, M. CDD: Specific functional annotation with the Conserved Domain Database. Nucleic Acids Res. 2009, 37, D205–D210. [Google Scholar] [CrossRef] [PubMed]
- Toll-Riera, M.; Albà, M.M. Emergence of novel domains in proteins. BMC Evolutionary Biology 2013, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A. The Pfam protein families database: towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.C.; McKay, D.B. Structure of the universal stress protein of Haemophilus influenzae. Structure 2001, 9, 1135–1141. [Google Scholar] [CrossRef]
- Veenman, L.; Vainshtein, A.; Yasin, N.; Azrad, M.; Gavish, M. Tetrapyrroles as endogenous TSPO ligands in eukaryotes and prokaryotes: comparisons with synthetic ligands. Int. J. Mol. Sci. 2016, 17, 880. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liu, J.; Liu, N.; Kuhn, L.A.; Garavito, R.M.; Ferguson-Miller, S. Translocator protein 18 kDa (TSPO): An old protein with new functions? Biochemistry (Mosc). 2016, 55, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.N.; Zhao, A.H.; Hussein, M.; Stocco, D.M.; Selvaraj, V. Translocator protein (TSPO) affects mitochondrial fatty acid oxidation in steroidogenic cells. Endocrinology 2016, 157, 1110–1121. [Google Scholar] [CrossRef] [PubMed]
- IMG/M. Website for Integrated Microbial Genomes & Microbiomes. Available online: https://img.jgi.doe.gov/ (accessed on 30 December 2016).
- BioCyc. BioCyc Pathway/Genome Database Collection. Available online: http://biocyc.org/ (accessed on 30 December 2016).
- BioCyc. Genome Overview Page of Lactobacillus plantarum WCFS1 in BioCyc.org. Available online: http://biocyc.org/LPLA220668/NEW-IMAGE?type=GENOME-OVERVIEW (accessed on 30 December 2016).
- BioCyc. BioCyc Web Page for Universal Stress Protein with Locus Tag lp_2993 in the Genome of Lactobacillus plantarum WCFS1. Available online: https://biocyc.org/gene?orgid=LPLA220668&id=G137Z-2535#tab=TU (accessed on 30 December 2016).
- PATRIC. PathoSystems Resource Integration Center’s File Transfer Protocol (FTP) Server. Available online: ftp://ftp.patricbrc.org/patric2 (accessed on 30 December 2016).
- NCSA. Blue Waters Supercomputer. Available online: https://bluewaters.ncsa.illinois.edu/ (accessed on 30 December 2016).
- Isokpehi, R.D. Interactive Visual Representations on Data on Universal Stress Protein of Lactobacilli. Available online: https://public.tableau.com/profile/publish/lactobacusp/abstract (accessed on 30 December 2016).
- Schneeberger, M.; Everard, A.; Gómez-Valadés, A.G.; Matamoros, S.; Ramírez, S.; Delzenne, N.M.; Gomis, R.; Claret, M.; Cani, P.D. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci. Rep. 2015, 5, 16643. [Google Scholar] [CrossRef] [PubMed]
- Heid, I.M.; Jackson, A.U.; Randall, J.C.; Winkler, T.W.; Qi, L.; Steinthorsdottir, V.; Thorleifsson, G.; Zillikens, M.C.; Speliotes, E.K.; Mägi, R. Meta-analysis identifies 13 new loci associated with waist-hip ratio and reveals sexual dimorphism in the genetic basis of fat distribution. Nat. Genet. 2010, 42, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-T.; Monda, K.L.; Taylor, K.C.; Lange, L.; Demerath, E.W.; Palmas, W.; Wojczynski, M.K.; Ellis, J.C.; Vitolins, M.Z.; Liu, S. Genome-wide association of body fat distribution in African ancestry populations suggests new loci. PLoS Genet. 2013, 9, e1003681. [Google Scholar] [CrossRef] [PubMed]
- IMG/M. Alignment of chromosomal cassette containing gene for universal stress protein of Akkermansia muciniphila. Available online: https://img.jgi.doe.gov/cgi-bin/m/main.cgi?section=GeneCassette&page=geneCassette&gene_oid=642612113&type=pfam (accessed on 30 December 2016).
- Pothuraju, R.; Sharma, R.; Kavadi, P.; Chagalamarri, J.; Jangra, S.; Bhakri, G.; De, S. Anti-obesity effect of milk fermented by Lactobacillus plantarum NCDC 625 alone and in combination with herbs on high fat diet fed C57BL/6J mice. Benef. Microbes 2016, 7, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xiao, X.; Dong, Y.; Xu, T.; Wu, F. Dietary supplementation with Lactobacillus plantarum dy-1 fermented barley suppresses body weight gain in high-fat diet-induced obese rats. J. Sci. Food Agric. 2016, 96, 4907–4917. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-H.; Song, J.-L.; Park, E.-S.; Ju, J.; Kim, H.-Y.; Park, K.-Y. Anti-obesity effects of starter fermented Kimchi on 3T3-L1 adipocytes. Prev. Nutr. and Food Sci. 2015, 20, 298. [Google Scholar] [CrossRef] [PubMed]
- Takemura, N.; Okubo, T.; Sonoyama, K. Lactobacillus plantarum strain No. 14 reduces adipocyte size in mice fed high-fat diet. Exp. Biol. Med. 2010, 235, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Yeliseev, A.A.; Kaplan, S. TspO of Rhodobacter sphaeroides. A structural and functional model for the mammalian peripheral benzodiazepine receptor. J. Biol. Chem. 2000, 275, 5657–5667. [Google Scholar] [CrossRef] [PubMed]
- Girard, C.; Liu, S.; Adams, D.; Lacroix, C.; Sineus, M.; Boucher, C.; Papadopoulos, V.; Rupprecht, R.; Schumacher, M.; Groyer, G. Axonal regeneration and neuroinflammation: Roles for the translocator protein 18 kDa. J. Neuroendocrinol. 2012, 24, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Serra, D.; Mera, P.; Malandrino, M.I.; Mir, J.F.; Herrero, L. Mitochondrial fatty acid oxidation in obesity. Antioxid. Redox Sign. 2013, 19, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Lassance, L.; Haghiac, M.; Minium, J.; Catalano, P.; Hauguel-de Mouzon, S. Obesity-induced down-regulation of the mitochondrial translocator protein (TSPO) impairs placental steroid production. J. Clin. Endocrinol. Metab. 2014, 100, E11–E18. [Google Scholar] [CrossRef] [PubMed]
- Doublié, S.; Bricogne, G.; Gilmore, C.; Carter Jr, C.W. Tryptophanyl-tRNA synthetase crystal structure reveals an unexpected homology to tyrosyl-tRNA synthetase. Structure 1995, 3, 17–31. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus Tag | Protein Length (aa) | Genome Name | Operon Neighbor Gene Product |
|---|---|---|---|
| LSA0042 | 155 | Lactobacillus sakei sakei 23K | 2-dehydropantoate 2-reductase |
| LAR_1223 | 170 | Lactobacillus reuteri JCM 1112 | dipeptidase PepV |
| lp_1322 | 168 | Lactobacillus plantarum WCFS1 | dipeptidase PepV |
| JDM1_1117 | 168 | Lactobacillus plantarum JDM1 | dipeptidase PepV |
| lp_2993 | 143 | Lactobacillus plantarum WCFS1 | manganese transport protein |
| JDM1_2397 | 143 | Lactobacillus plantarum JDM1 | manganese transport protein |
| LSA0247 | 144 | Lactobacillus sakei sakei 23K | manganese transport protein |
| lp_1701 | 152 | Lactobacillus plantarum WCFS1 | sensory protein |
| JDM1_1432 | 152 | Lactobacillus plantarum JDM1 | sensory protein |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isokpehi, R.D.; Simmons, S.S.; Johnson, M.O.; Payton, M. Genomic Evidence for Bacterial Determinants Influencing Obesity Development. Int. J. Environ. Res. Public Health 2017, 14, 345. https://doi.org/10.3390/ijerph14040345
Isokpehi RD, Simmons SS, Johnson MO, Payton M. Genomic Evidence for Bacterial Determinants Influencing Obesity Development. International Journal of Environmental Research and Public Health. 2017; 14(4):345. https://doi.org/10.3390/ijerph14040345
Chicago/Turabian StyleIsokpehi, Raphael D., Shaneka S. Simmons, Matilda O. Johnson, and Marinelle Payton. 2017. "Genomic Evidence for Bacterial Determinants Influencing Obesity Development" International Journal of Environmental Research and Public Health 14, no. 4: 345. https://doi.org/10.3390/ijerph14040345