
Biodegradation of Mixed PAHs by PAH-Degrading Endophytic Bacteria

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Isolation of PAH-Degrading Endophytic Bacteria
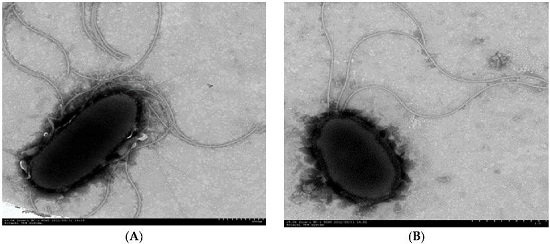
2.2. Identification of PAH-Degrading Endophytic Bacteria
2.3. Biodegradation of PAHs Using Endophytic Bacteria
2.3.1. Degradation of PHE
2.3.2. Capacities of Degrading Other PAHs
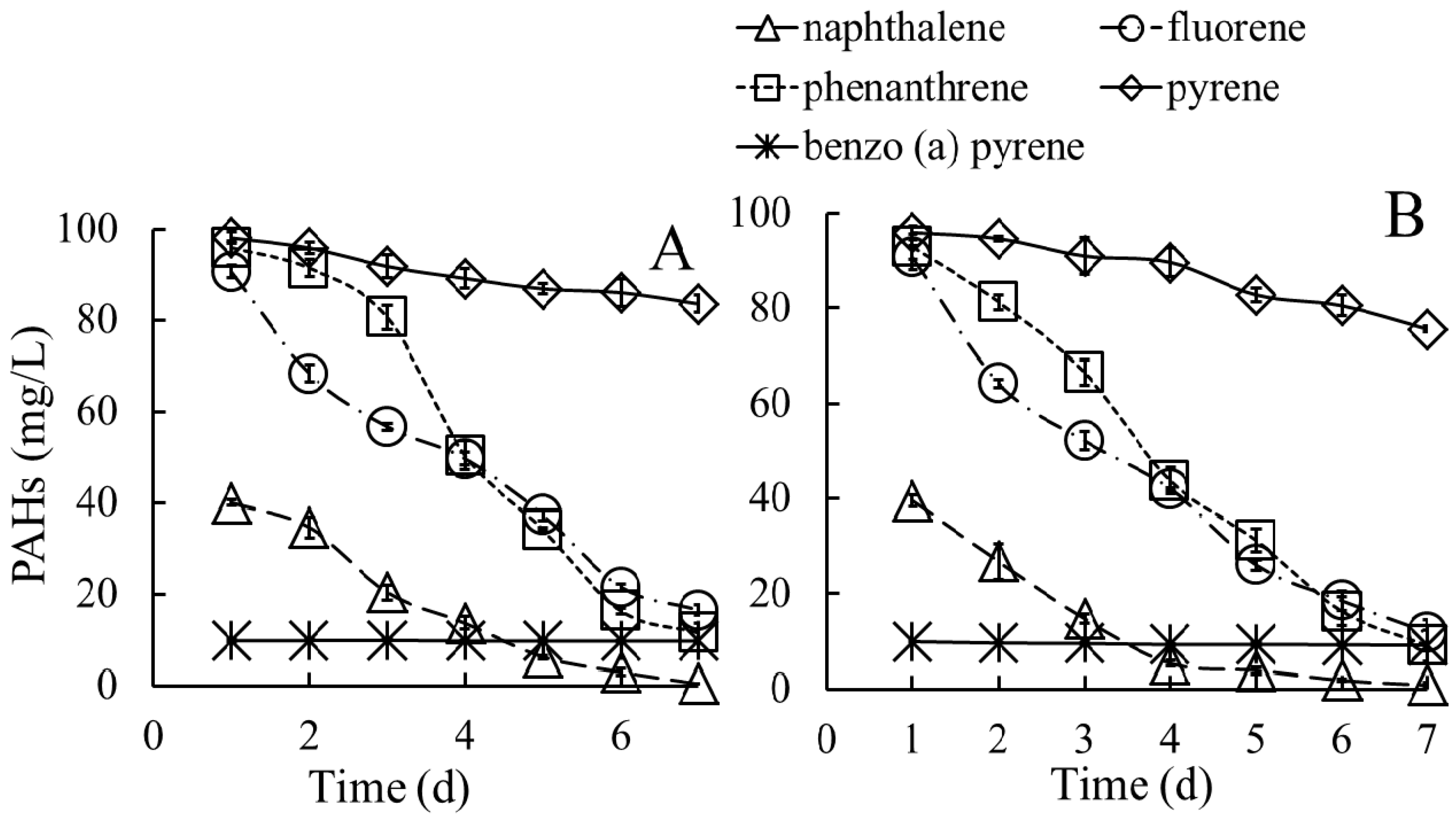
2.3.3. Capacities of Degrading a Mixture of PAHs
2.4. Effects of Additional Nutrients on Degradation of PHE
2.5. Detection of PAH Residues Using HPLC
2.6. Statistical Analyses
3. Results and Discussion
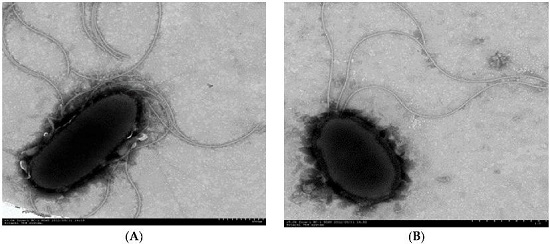
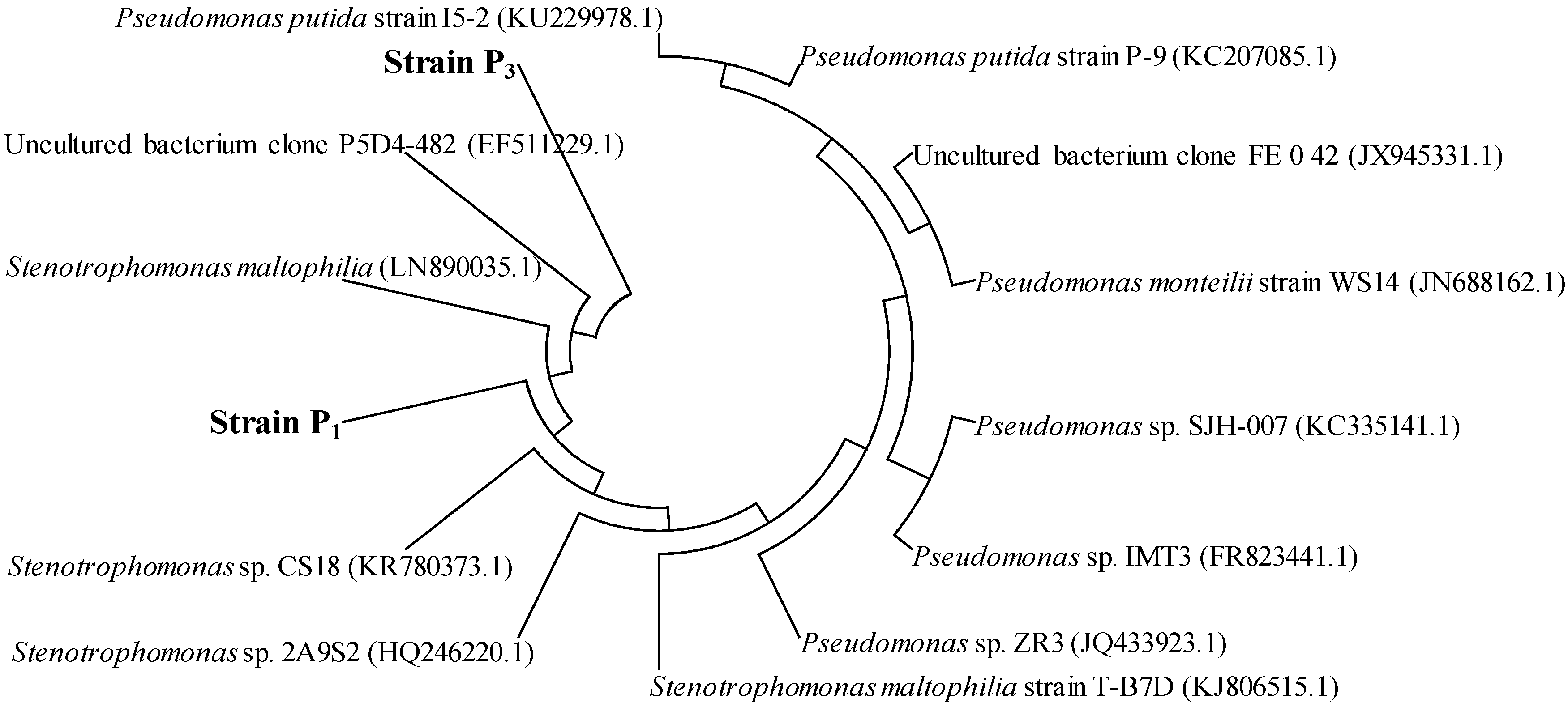
3.1. Isolation and Identification of PAH-Degrading Endophytic Bacteria
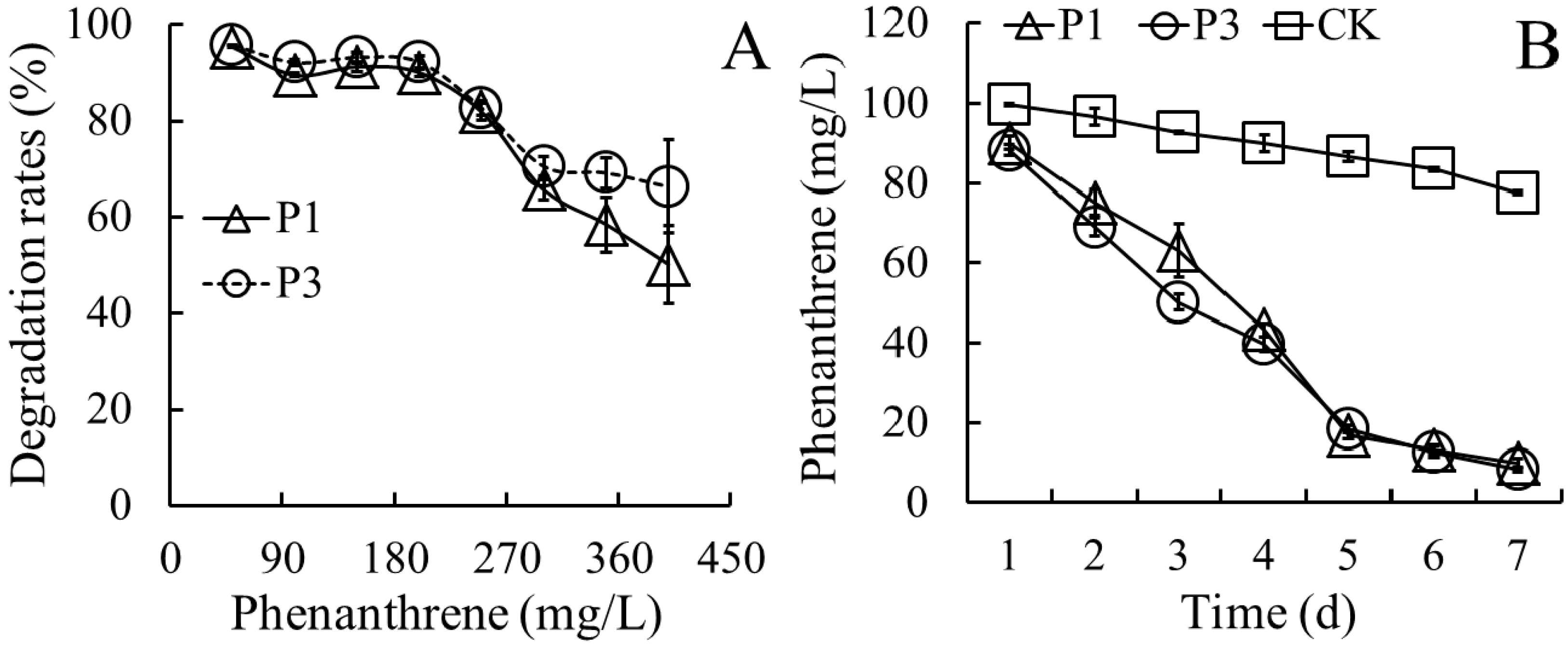
3.2. Biodegradation Kinetics of PHE by Strains P1 and P3
- CPHE = 167.61 × e −0.4075 t (by strain P1, r = 0.9738)
- CPHE = 156.21 × e −0.4099 t (by strain P3, r = 0.9871)
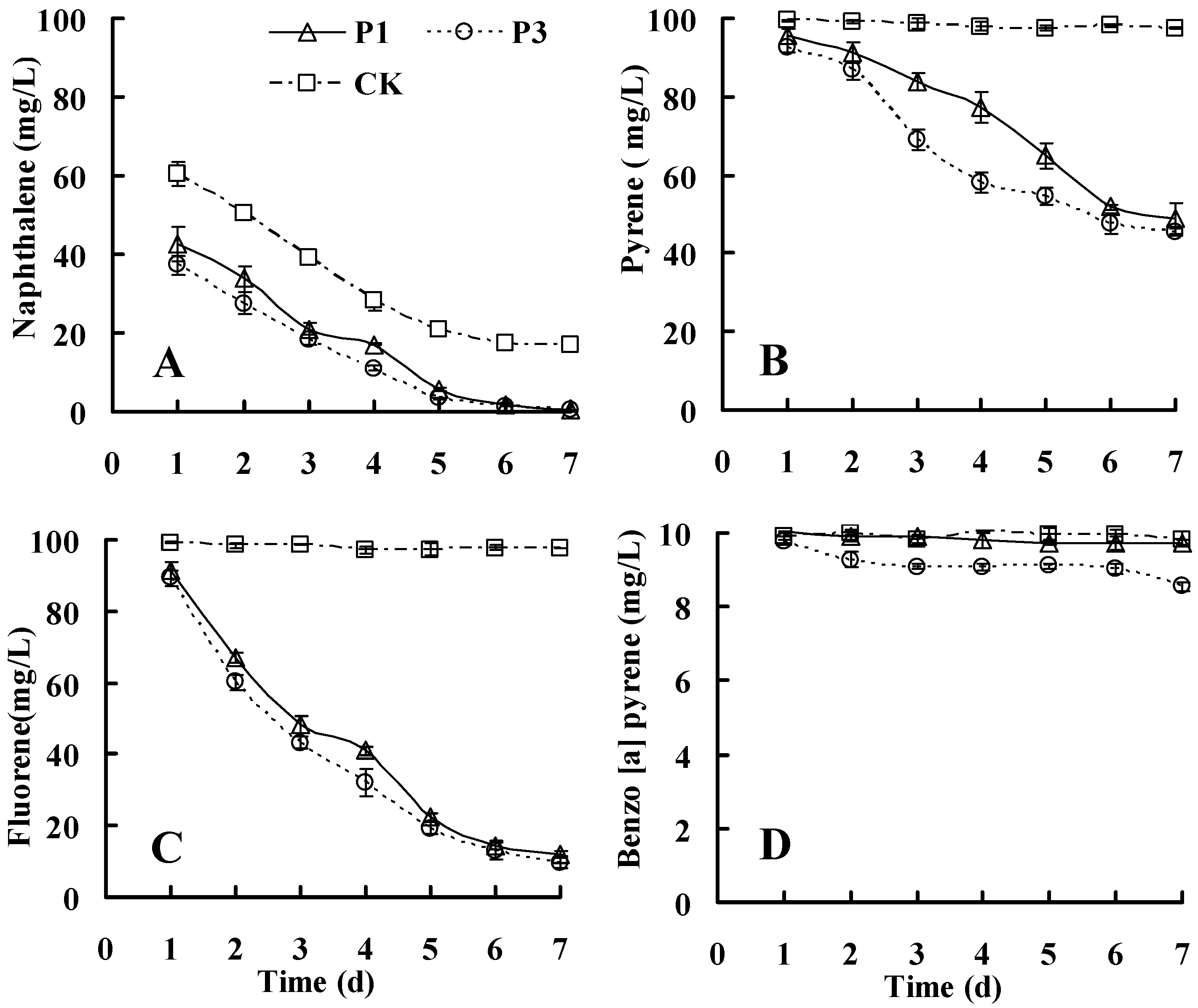
3.3. Biodegradation Kinetics of Other PAHs
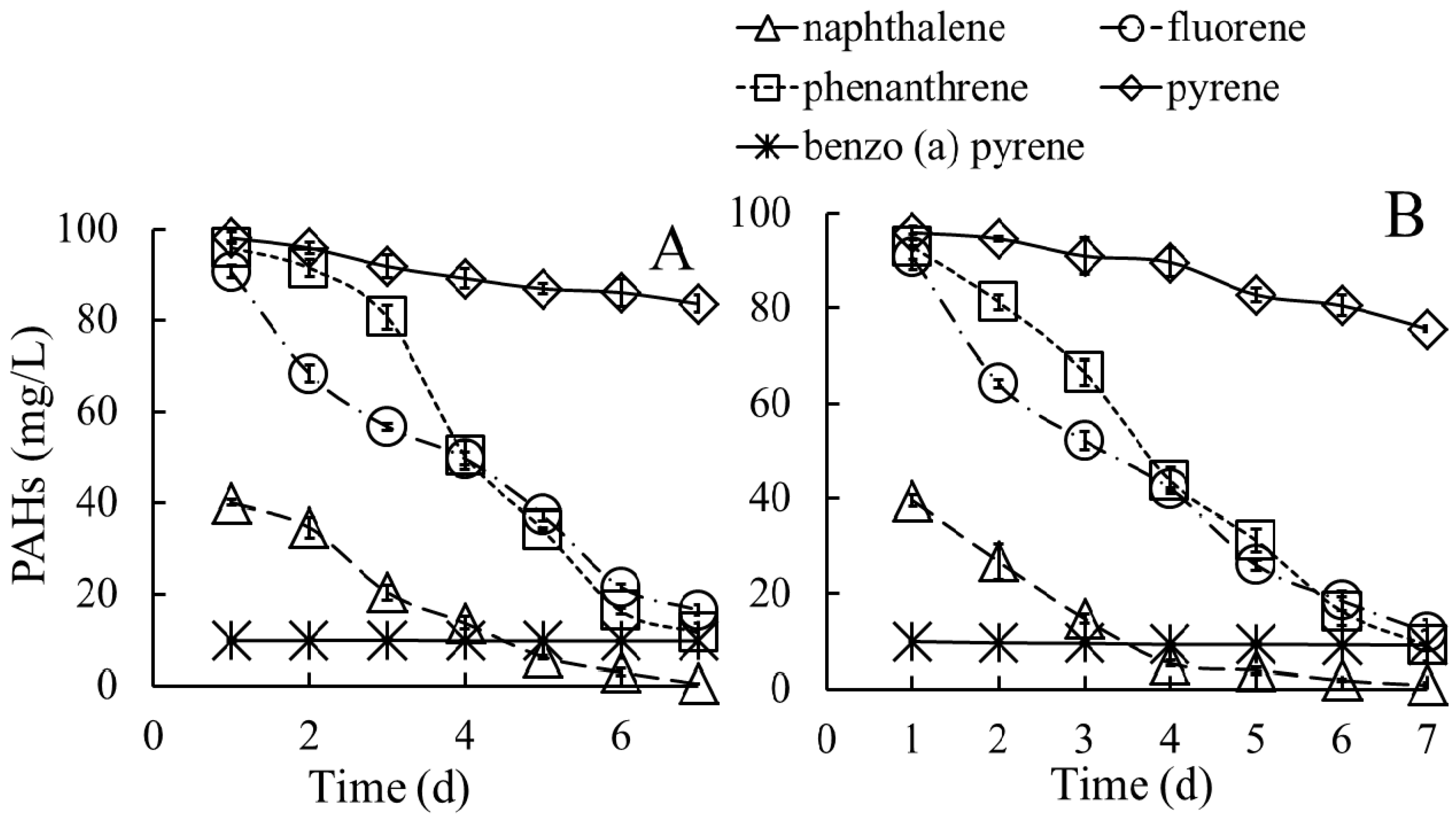
3.3.1. Biodegradation of PAHs in Medium Containing Individual PAHs
3.3.2. Biodegradation of PAHs in Media Containing a Mixture of PAHs
3.4. Additional Carbon and Nitrogen Nutrients Enhance the Biodegradation of PHE
3.5. Potential Application for PAH-Degrading Endophytic Bacteria
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tao, S.; Cui, Y.H.; Xu, F.L.; Li, B.G.; Cao, J.; Liu, W.X.; Schmitt, G.; Wang, X.J.; Shen, W.R.; Qing, B.P.; et al. Polycyclic aromatic hydrocarbons (PAHs) in agricultural soil and vegetables from Tianjin. Sci. Total Environ. 2004, 320, 11–24. [Google Scholar] [CrossRef]
- Tang, L.; Tang, X.Y.; Zhu, Y.G.; Zheng, M.H.; Miao, Q.L. Contamination of polycyclic aromatic hydrocarbons (PAHs) in urban soils in Beijing, China. Environ. Int. 2005, 31, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.T.; Miao, Y.; Zhang, Y.; Li, Y.C.; Wu, M.H.; Yu, G. Polycyclic aromatic hydrocarbons (PAHs) in urban soils of the megacity Shanghai: Occurrence, source apportionment and potential human health risk. Sci. Total Environ. 2013, 447, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Orecchio, S. PAHs associated with the leaves of Quercus ilex L.: Extraction, GC-MS analysis, distribution and sources: Assessment of air quality in the Palermo (Italy) area. Atmos. Environ. 2007, 41, 8669–8680. [Google Scholar] [CrossRef]
- Maliszewska-Kordybach, B.; Smreczak, B.; Klimkowicz-Pawlas, A.; Terelak, H. Monitoring of the total content of polycyclic aromatic hydrocarbons (PAHs) in arable soils in Poland. Chemosphere 2008, 73, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tian, Z.J.; Zhu, H.L.; Cheng, Z.N.; Kang, M.L.; Luo, C.L.; Li, J.; Zhang, G. Polycyclic aromatic hydrocarbons (PAHs) in soils and vegetation near an e-waste recycling site in south China: Concentration, distribution, source, and risk assessment. Sci. Total Environ. 2012, 439, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Macek, T.; Kotrba, P.; Svatos, A.; Novakova, M.; Demnerova, K.; Mackova, M. Novel roles for genetically modified plants in environmental protection. Trends Biotechnol. 2008, 26, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Bacosa, H.P.; Inoue, C. Polycyclic aromatic hydrocarbons (PAHs) biodegradation potential and diversity of microbial consortia enriched from tsunami sediments in Miyagi. J. Hazard Mater. 2015, 283, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Maiti, A.; Das, S.; Bhattacharyya, N. High gelatinase activity of a newly isolated polycyclic aromatic hydrocarbon degrading bacteria Bacillus weihenstephanensis strain AN1. J. Pharm. Res. 2013, 6, 199–204. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Suto, K.; Inoue, C. Bacterial community dynamics during the preferential degradation of aromatic hydrocarbons by a microbial consortium. Int. Biodeterior. Biodegrad. 2012, 74, 109–115. [Google Scholar] [CrossRef]
- Balachandran, C.; Duraipandiyan, V.; Balakrishna, K.; Ignacimuthu, S. Petroleum and polycyclic aromatic hydrocarbons (PAHs) degradation and naphthalene metabolism in Streptomyces sp. (ERI-CPDA-1) isolated from oil contaminated soil. Bioresour. Technol. 2012, 112, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Song, X.H.; Xu, Y.; Li, G.M.; Zhang, Y.; Huang, T.W.; Hu, Z. Isolation, characterization of Rhodococcus sp. P14 capable of degrading high-molecular-weight polycyclic aromatic hydrocarbons and aliphatic hydrocarbons. Mar. Pollut. Bull. 2011, 62, 2122–2128. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Li, Q.X.; Xu, T.; Cui, Z.L.; Sun, Y.; Li, J. Sphingobium sp. FB3 degrades a mixture of polycyclic aromatic hydrocarbons. Int. Biodeterior. Biodegrad. 2014, 87, 44–51. [Google Scholar] [CrossRef]
- Sun, K.; Liu, J.; Gao, Y.; Gu, Y.; Wang, W. Isolation, plant colonization potential, and phenanthrene degradation performance of the endophytic bacterium Pseudomonas sp. Ph6-gfp. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Bacosa, H.P.; Suto, K.; Inoue, C. Degradation potential and microbial community structure of heavy oil-enriched microbial consortia from mangrove sediments in Okinawa, Japan. J. Environ. Sci. Health A 2013, 48, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Weyens, N.; Truyens, S.; Dupae, J.; Newman, L.; Taghavi, S.; van der Lelie, D.; Carleer, R.; Vangronsveld, J. Potential of the TCE-Degrading endophyte pseudomonas putida W619-TCE to improve plant growth and reduce TCE phytotoxicity and evapotranspiration in poplar cuttings. Environ. Pollut. 2010, 158, 2915–2919. [Google Scholar] [CrossRef] [PubMed]
- Newman, L.A.; Reynolds, C.M. Bacteria and phytoremediation: New uses for endophytic bacteria in plants. Trends Biotechnol. 2005, 23, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dashti, N.; Khanafer, M.; El-Nemr, I.; Sorkhoh, N.; Ali, N.; Radwan, S. The potential of oil-utilizing bacterial consortia associated with legume root nodules for cleaning oily soils. Chemosphere 2009, 74, 1354–1359. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Wang, E.T.; Chen, W.F.; Chen, W.X. Genetic diversity and potential for promotion of plant growth detected in nodule endophytic bacteria of soybean grown in Heilongjiang province of China. Soil Biol. Biochem. 2008, 40, 238–246. [Google Scholar] [CrossRef]
- Wen, J.; Gao, D.; Zhang, B.; Liang, H. Co-metabolic degradation of pyrene by indigenous white-rot fungus Pseudotrametes gibbosa from the northeast China. Int. Biodeterior. Biodegrad. 2011, 65, 600–604. [Google Scholar] [CrossRef]
- Suto, M.; Takebayashi, M.; Saito, K.; Tanaka, M.; Yokota, A.; Tomita, F. Endophytes as producers of xylanase. J. Biosci. Bioeng. 2002, 93, 88–90. [Google Scholar] [CrossRef]
- Guo, C.L.; Zhou, H.W.; Wong, Y.S.; Tam, N.F.Y. Isolation of PAH-degrading bacteria from mangrove sediments and their biodegradation potential. Mar. Pollut. Bull. 2005, 51, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.Z.; Ni, X.; Waigi, M.G.; Liu, J.; Sun, K.; Gao, Y.Z. Microbial biodegradation of mixed PAHs through PAH-degrading endophytic bacteria. Int. J. Environ. Res. Public Health 2016, 13, 633. [Google Scholar] [CrossRef] [PubMed]
- Byers, H.K.; Stackebrandt, E.; Hayward, C.; Blackall, L.L. Molecular investigation of a microbial mat associated with the Great Artesian Basin. FEMS Microbiol. Ecol. 1998, 25, 391–403. [Google Scholar] [CrossRef]
- GenBank Database. Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 8 August 2016).
- Bacosa, H.; Suto, K.; Inoue, C. Preferential utilization of petroleum oil hydrocarbon components by microbial consortia reflects degradation pattern in aliphatic-aromatic hydrocarbon binary mixtures. World J. Microbiol. Biotechnol. 2011, 27, 1109–1117. [Google Scholar] [CrossRef]
- Juhasz, A.L.; Stanley, G.A.; Britz, M.L. Microbial degradation and detoxification of high molecular weight polycyclic aromatic hydrocarbons by Stenotrophomonas maltophilia strain VUN 10003. Lett. Appl. Microbiol. 2000, 30, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.; Patel, J.; Madamwar, D. Biodegradation of phenanthrene in bioaugmented microcosm by consortium ASP developed from coastal sediment of Alang-Sosiya ship breaking yard. Mar. Pollut. Bull. 2013, 74, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, N.; Xue, M.; Lu, S.T.; Tao, S. Effects of soil organic matter on the development of the microbial polycyclic aromatic hydrocarbons (PAHs) degradation potentials. Environ. Pollut. 2011, 159, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Xu, L.; Jia, L.Y. Characterization of pyrene degradation by Pseudomonas sp. strain Jpyr-1 isolated from active sewage sludge. Bioresour. Technol. 2013, 140, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Zou, S.C.; Lin, L.; Luan, T.G.; Qiu, R.L.; Tam, N.F.Y. Effects of pyrene and fluoranthene on the degradation characteristics of phenanthrene in the cometabolism process by Sphingomonas sp. strain PheB4 isolated from mangrove sediments. Mar. Pollut. Bull. 2010, 60, 2043–2049. [Google Scholar] [CrossRef] [PubMed]
- Ambrosoli, R.; Petruzzelli, L.; Minati, J.L.; Marsan, F.A. Anaerobic PAH degradation in soil by a mixed bacterial consortium under denitrifying conditions. Chemosphere 2005, 60, 1231–1236. [Google Scholar] [CrossRef] [PubMed]
- Warshawsky, D.; LaDow, K.; Schneider, J. Enhanced degradation of benzo(a)pyrene by Mycobacterium sp. in conjunction with green alga. Chemosphere 2007, 69, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Su, L.H.; Zhou, H.Y.; Guo, G.Z.; Zhao, A.H.; Zhao, Y.C. Anaerobic biodegradation of PAH in river sediment treated with different additives. Procedia Environ. Sci. 2012, 16, 311–319. [Google Scholar] [CrossRef]
- Ahmed, R.Z.; Ahmed, N. Effect of yeast extract on fluoranthene degradation and aromatic ring dioxygenase expressing bacterial community structure of a fluoranthene degrading bacterial consortium. Int. Biodeterior. Biodegrad. 2014, 88, 56–61. [Google Scholar] [CrossRef]
- Chen, B.L.; Yuan, M.X. Enhanced dissipation of polycyclic aromatic hydrocarbons in the presence of fresh plant residues and their extracts. Environ. Pollut. 2012, 161, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Murai, Y.; Tatsumi, K.; Iimura, Y. Biodegradation of polycyclic aromatic hydrocarbons by Sphingomonas sp. enhanced by water-extractable organic matter from manure compost. Sci. Total Environ. 2009, 407, 5805–5810. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.G.; Bao, M.T.; Fan, X.N.; Liang, S.K.; Sun, P.Y. Rhamnolipids enhance marine oil spill bioremediation in laboratory system. Mar. Pollut. Bull. 2013, 71, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.S.; Syeda, K.; Shann, J.; Yadav, J.S. A novel P450-initiated biphasic process for sustainable biodegradation of benzo(a)pyrene in soil under nutrient-sufficient conditions by the white rot fungus Phanerochaete chrysosporium. J. Hazard Mater. 2013, 261, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Vauramo, S.; Jaaskelainen, V.; Setala, H. Environmental fate of polycyclic aromatic hydrocarbons under different plant traits in urban soil as affected by nitrogen deposition. Appl. Soil Ecol. 2011, 47, 167–175. [Google Scholar] [CrossRef]
- Brandenburg, K.S.; Rodriguez, K.J.; McAnulty, J.F.; Murphy, C.J.; Abbott, N.L.; Schurr, M.J.; Czuprynski, C.J. Tryptophan inhibits biofilm formation by Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013, 57, 1921–1925. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Chen, X.; He, L. Characteristics of an endophytic pyrene-degrading bacterium of Enterobacter sp. 12J1 from Allium macrostemon Bunge. Int. Biodeterior. Biodegrad. 2008, 62, 88–95. [Google Scholar] [CrossRef]
- Afzal, M.; Khan, Q.M.; Sessitsch, A. Endophytic bacteria: Prospects and applications for the phytoremediation of organic pollutants. Chemosphere 2014, 117, 232–242. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAHs | Degradation Analysis | Strain P1 | Strain P3 |
|---|---|---|---|
| NAP | Degradation kinetics equations | CNAP = 189.09 × e −0.8072 t (r = 0.9353) | CNAP = 138.99 × e −0.7813 t (r = 0.9681) |
| Half-life (days) | 0.86 | 0.89 | |
| PHE | Degradation kinetics equations | CPHE = 167.61 × e −0.4075 t (r = 0.9738) | CPHE = 156.21 × e −0.4099 t (r = 0.9871) |
| Half-life (days) | 1.70 | 1.69 | |
| FLR | Degradation kinetics equations | CFLR = 137.99 × e −0.354 t (r = 0.9906) | CFLR = 132.24 × e −0.3784 t (r = 0.9981) |
| Half-life (days) | 1.96 | 1.83 | |
| PYR | Degradation kinetics equations | CPYR = 115.73 × e −0.1215 t (r = 0.9763) | CPYR = 104.13 × e −0.127 t (r = 0.9829) |
| Half-life (days) | 5.70 | 5.46 | |
| B(a)P | Degradation kinetics equations | CB(a)P = 10.036 × e −0.0051 t (r = 0.9356) | CB(a)P = 9.7435 × e −0.0164 t (r = 0.8885) |
| Half-life (days) | 135.88 | 42.26 |
| PAHs | Degradation Analysis | Strain P1 | Strain P3 |
|---|---|---|---|
| NAP | Degradation kinetics equations | CNAP = 150.31 × e −0.7287 t (r = 0.9386) | CNAP = 89.541 × e −0.6571 t (r = 0.9943) |
| Half-life (days) | 0.95 | 1.05 | |
| PHE | Degradation kinetics equations | CPHE = 192.68 × e −0.3815 t (r = 0.9656) | CPHE = 178.8 × e −0.3916 t (r = 0.9736) |
| Half-life (days) | 1.82 | 1.77 | |
| FLR | Degradation kinetics equations | CFLR = 128.82 × e −0.2803 t (r = 0.9810) | CFLR = 134.63 × e −0.3325 t (r = 0.9917) |
| Half-life (days) | 2.47 | 2.08 | |
| PYR | Degradation kinetics equations | CPYR = 100.26 × e −0.0269 t (r = 0.9893) | CPYR = 102.21 × e −0.0406 t (r = 0.9794) |
| Half-life (days) | 25.76 | 17.07 | |
| B(a)P | Degradation kinetics equations | CB(a)P = 9.8059 × e ‒0.0021 t (r = 0.6901) | CB(a)P = 9.8931 × e −0.012 t (r = 0.9415) |
| Half-life (days) | 330 | 57.75 |
| Extra Carbon Sources | Extra Nitrogen Sources | ||||
|---|---|---|---|---|---|
| Carbon Sources | Degradation (%) | Nitrogen Sources | Degradation (%) | ||
| Strain P1 | Strain P3 | Strain P1 | Strain P3 | ||
| CK | 90.5 ± 0.2 e | 90.4 ± 0.5 h | CK | 90.5 ± 0.2 c | 90.4 ± 0.5 cd |
| Glucose | 98.7 ± 0.1 a | 98.7 ± 0.1 ab | NH4Cl | 90.0 ± 1.2 c | 91.6 ± 1.5 cd |
| Fructose | 99.0 ± 0.1 a | 98.8 ± 0.2 ab | NH4NO3 | 89.7 ± 1.4 c | 91.1 ± 0.7 cd |
| Sucrose | 97.7 ± 0.2 a | 97.9 ± 0.4 b | (NH4)2SO4 | 91.4 ± 0.8 c | 90.1 ± 1.1 d |
| Yeast | 98.4 ± 0.1 a | 99.3 ± 0.1 a | (NH4)2HPO4 | 91.2 ± 1.6 c | 90.9 ± 1.4 cd |
| Soluble starch | 94.1 ± 0.5 c | 97.8 ± 0.2 b | Peptone | 96.7 ± 0.9 b | 99.0 ± 0.2 a |
| Glycerine | 98.6 ± 0.1 a | 94.8 ± 0.3 c | Urea | 90.1 ± 1.1 c | 89.6 ± 0.9 d |
| Malic acid | 95.8 ± 0.5 b | 92.8 ± 0.7 de | Beef extract | 99.1 ± 0.2 a | 99.1 ± 0.1 a |
| Oxalic acid | 95.2 ± 0.5 b | 93.0 ± 0.3 de | Tryptophan | 76.1 ± 2.6 e | 66.2 ± 2.5 e |
| Citric acid | 96.1 ± 0.5 b | 93.3 ± 0.6 d | Arginine | 84.6 ± 1.3 d | 90.1 ± 2.5 d |
| Mannite | 92.7 ± 0.6 d | 98.4 ± 0.3 ab | Cysteine | 91.4 ± 1.0 c | 92.5 ± 1.6 c |
| Sorbitol | 92.5 ± 0.5 d | 98.0 ± 0.2 b | Proline | 94.7 ± 1.1 b | 95.7 ± 0.5 b |
| Catechol | 91.2 ± 0.5 de | 92.1 ± 0.4 ef | |||
| Phthalate | 90.9 ± 0.5 e | 90.8 ± 1.0 gh | |||
| Salicylic acid | 90.5 ± 1.0 e | 91.6 ± 0.7 fg | |||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Ni, X.; Waigi, M.G.; Liu, J.; Sun, K.; Gao, Y. Biodegradation of Mixed PAHs by PAH-Degrading Endophytic Bacteria. Int. J. Environ. Res. Public Health 2016, 13, 805. https://doi.org/10.3390/ijerph13080805
Zhu X, Ni X, Waigi MG, Liu J, Sun K, Gao Y. Biodegradation of Mixed PAHs by PAH-Degrading Endophytic Bacteria. International Journal of Environmental Research and Public Health. 2016; 13(8):805. https://doi.org/10.3390/ijerph13080805
Chicago/Turabian StyleZhu, Xuezhu, Xue Ni, Michael Gatheru Waigi, Juan Liu, Kai Sun, and Yanzheng Gao. 2016. "Biodegradation of Mixed PAHs by PAH-Degrading Endophytic Bacteria" International Journal of Environmental Research and Public Health 13, no. 8: 805. https://doi.org/10.3390/ijerph13080805