
Combined Effects of Soy Isoflavones and β-Carotene on Osteoblast Differentiation

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Cell Cultures
2.3. Measurement of Alkaline Phosphatase Activity in MC3T3-E1 Cells
2.4. Cell Growth of Preosteoblasts
2.5. Total RNA Isolation and Reverse Transcription Quantitative Real-Time PCR (RT-qPCR)
2.6. Statistical Analysis
3. Results
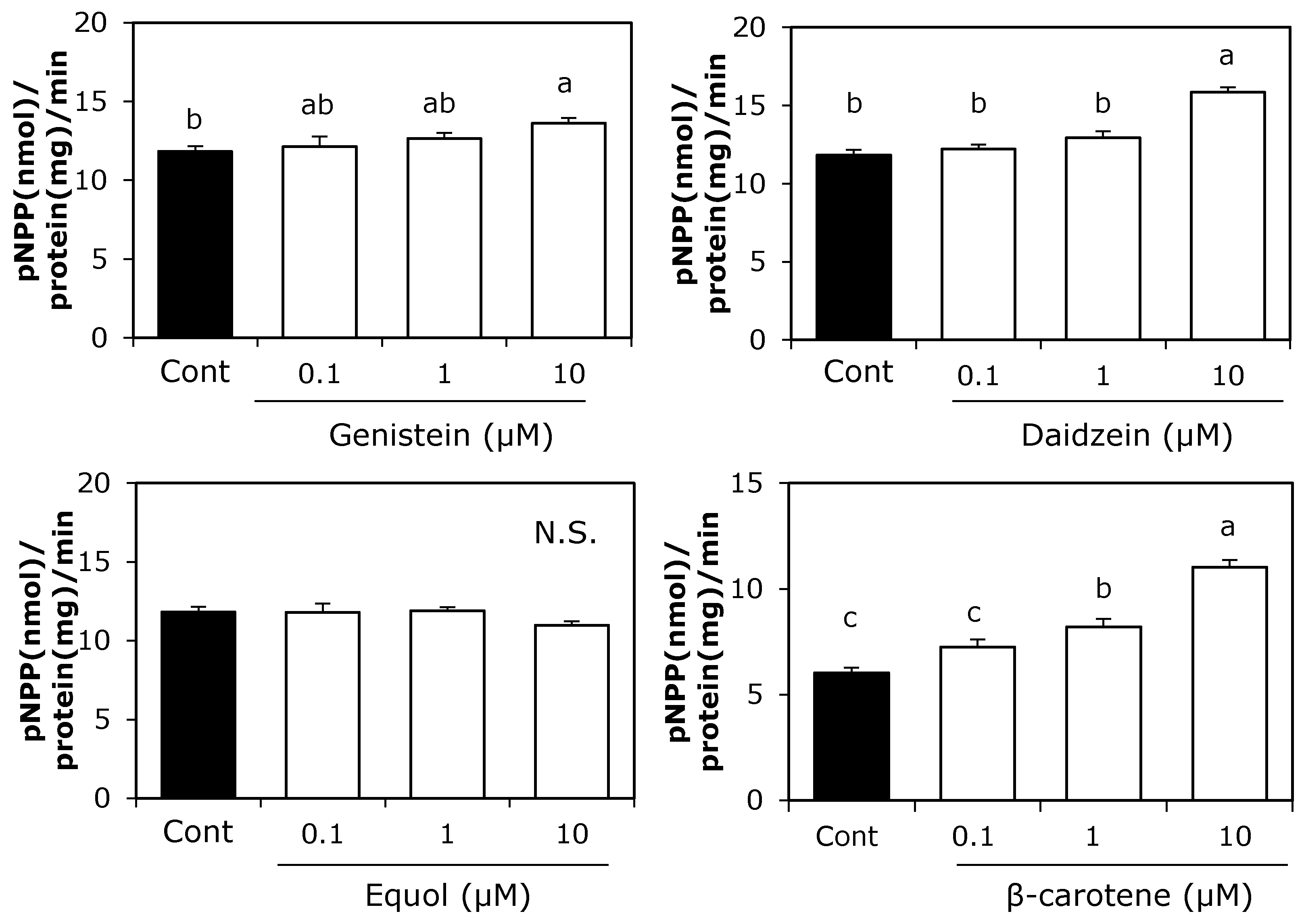
3.1. Effect of Soy Isoflavones and β-Carotene on ALP Activity in MC3T3-E1 Cells
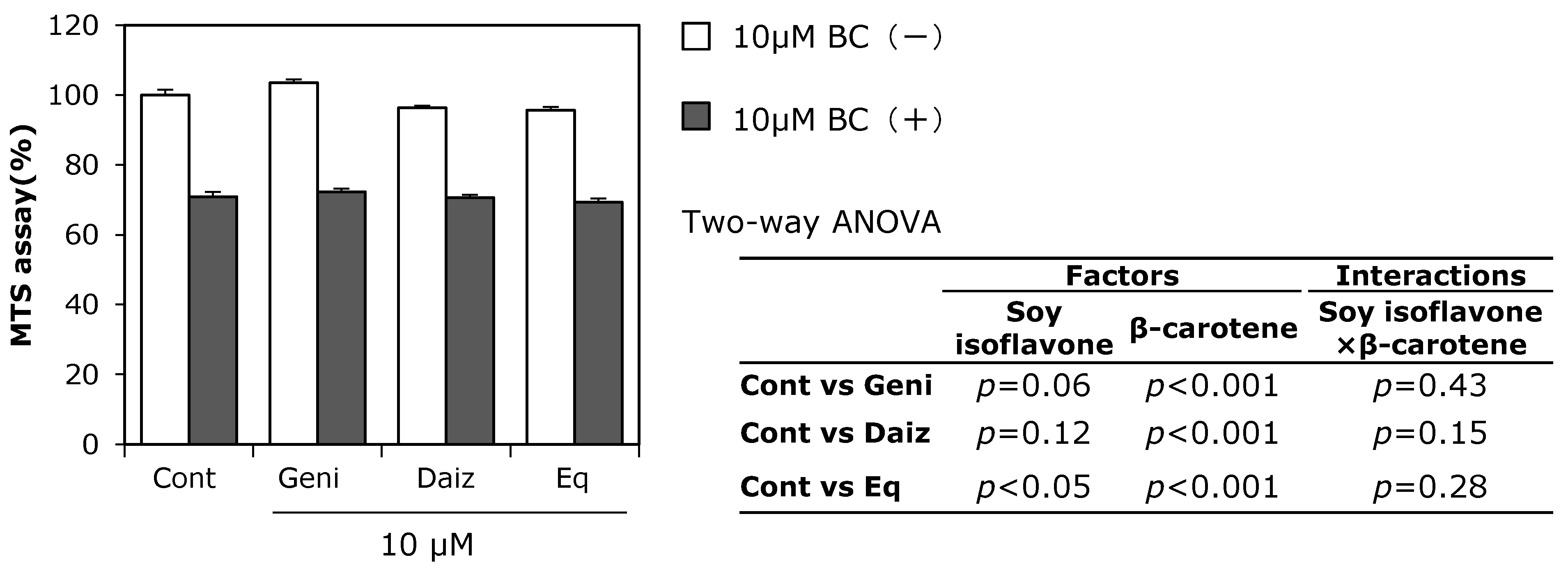
3.2. Combined Effect of Soy Isoflavones with β-Carotene on MC3T3-E1 Cell Growth
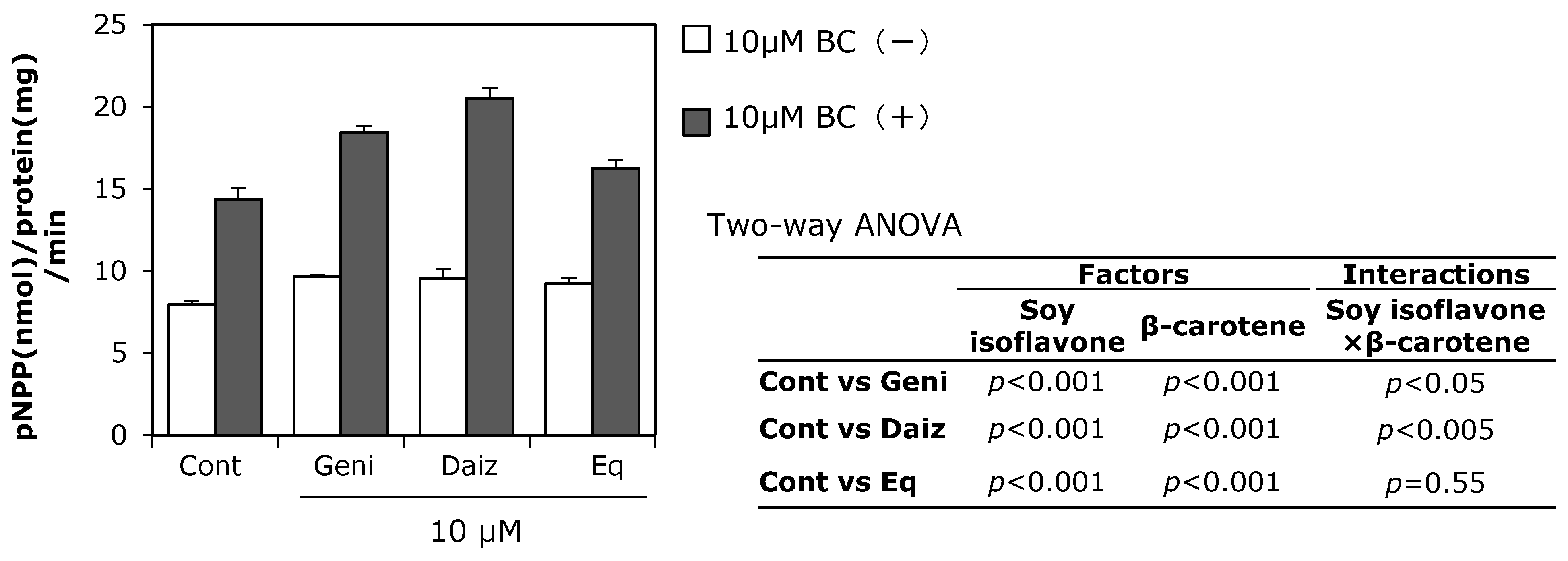
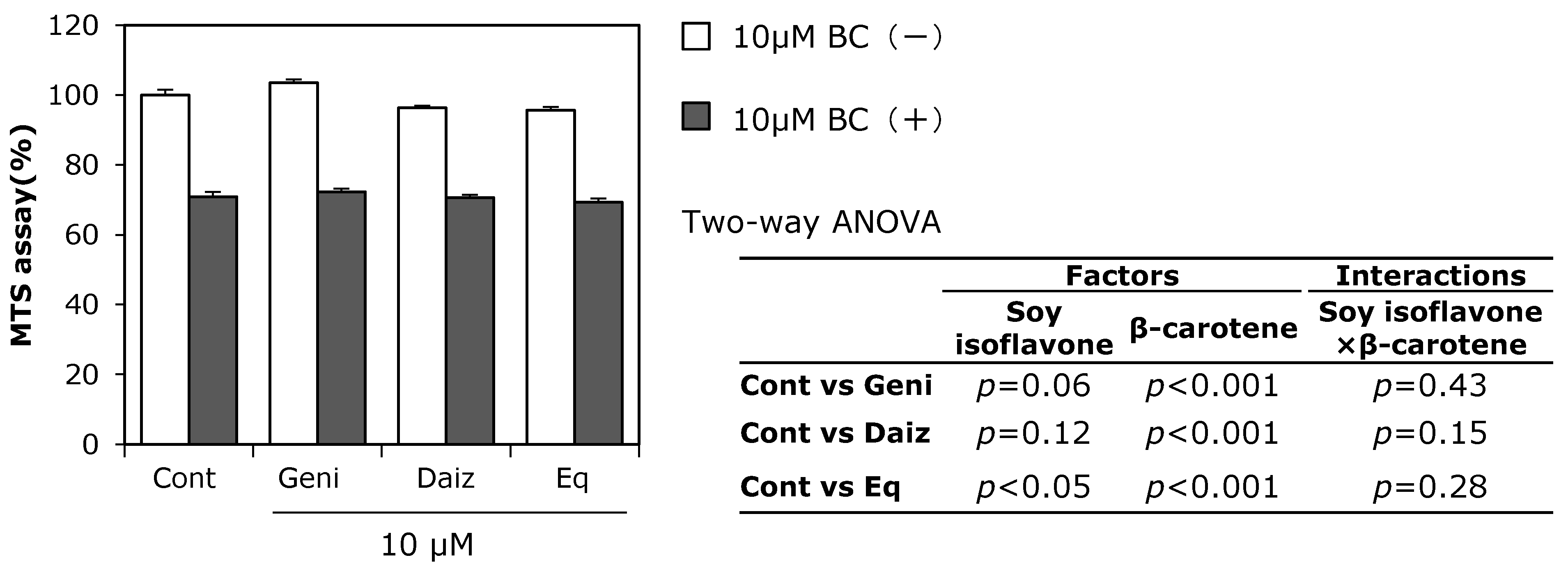
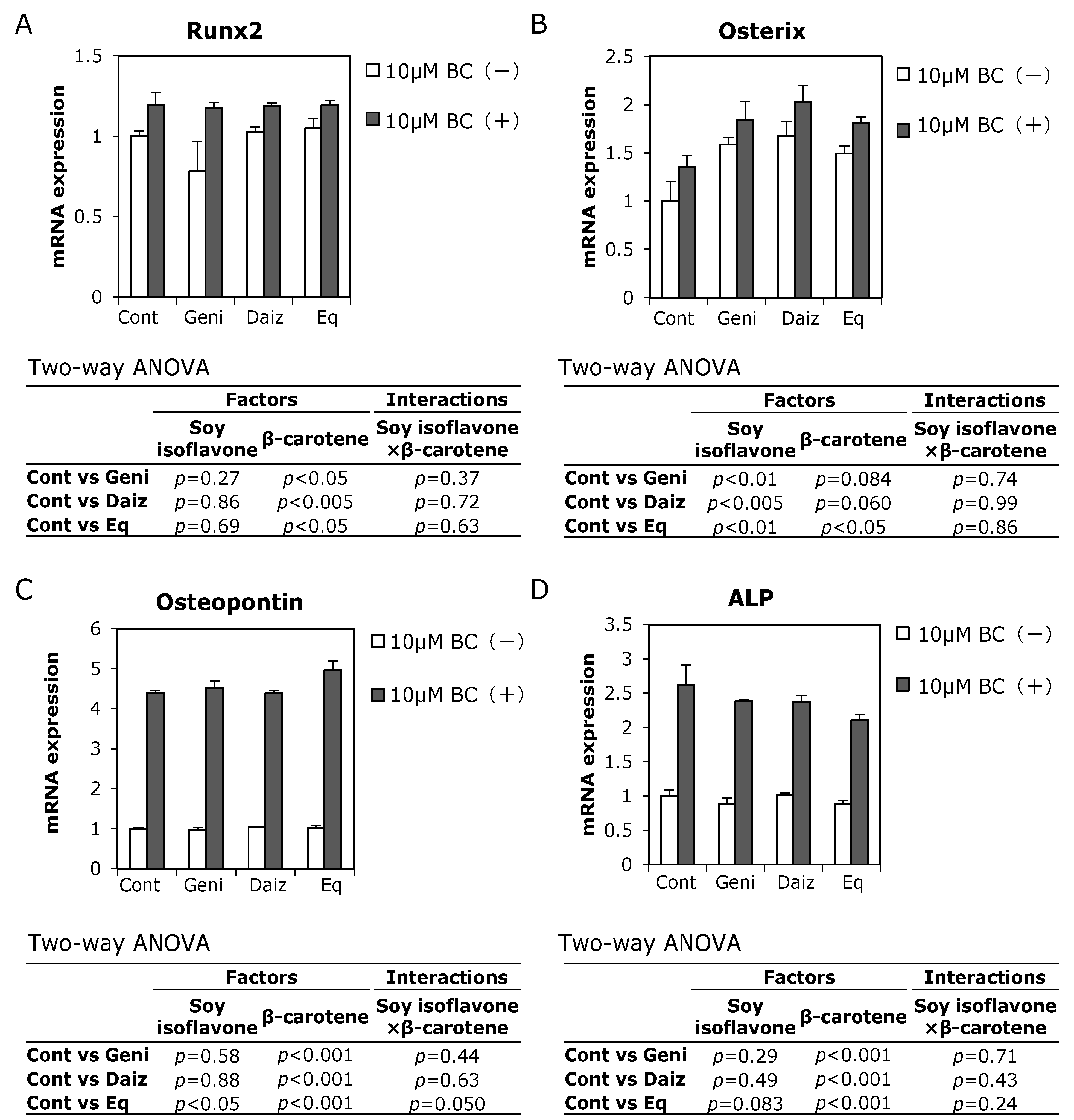
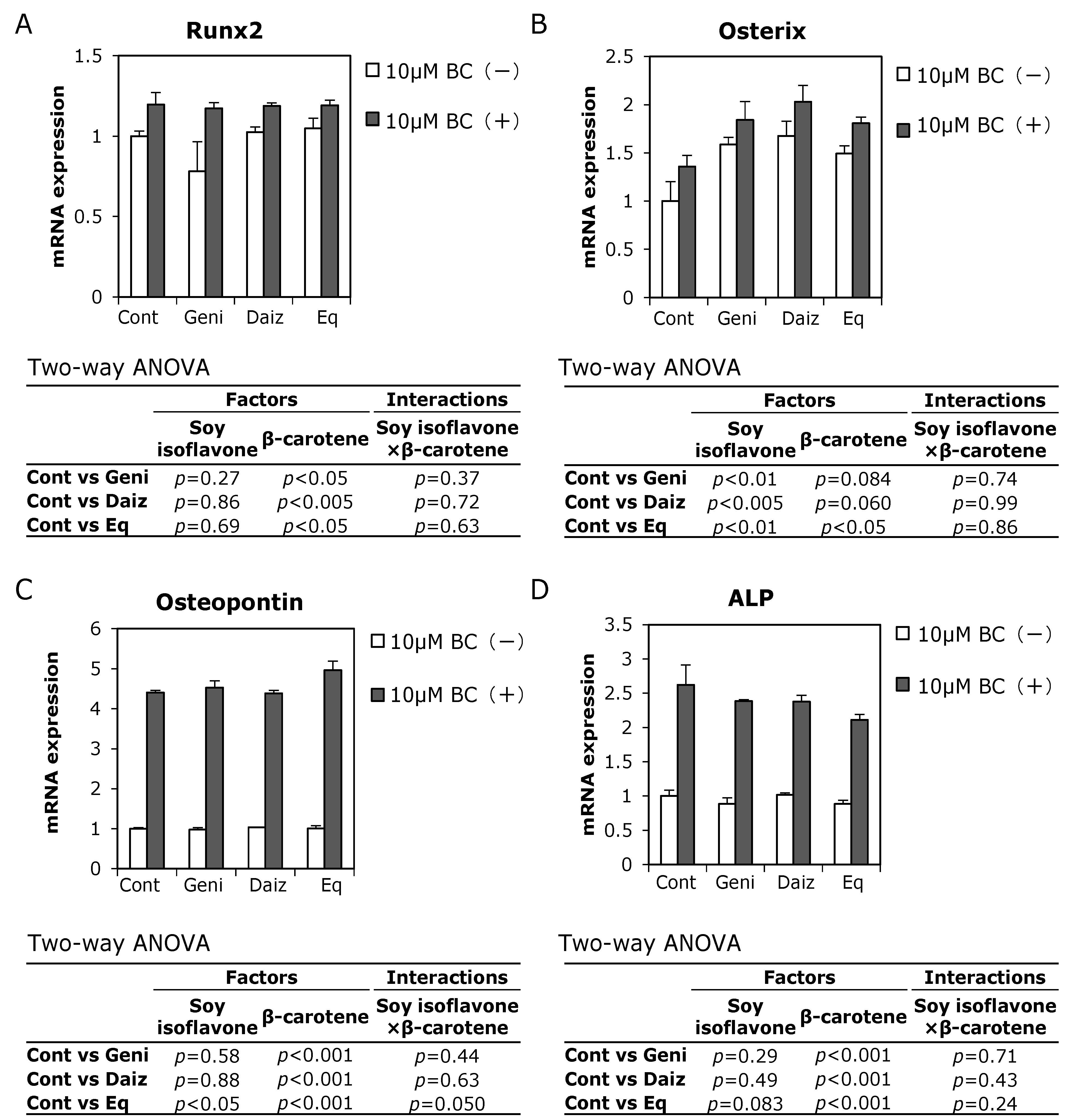
3.3. Combined Effects of Soy Isoflavones and β-Carotene on Expression of Osteoblast-Related Genes in MC3T3-E1 Cells
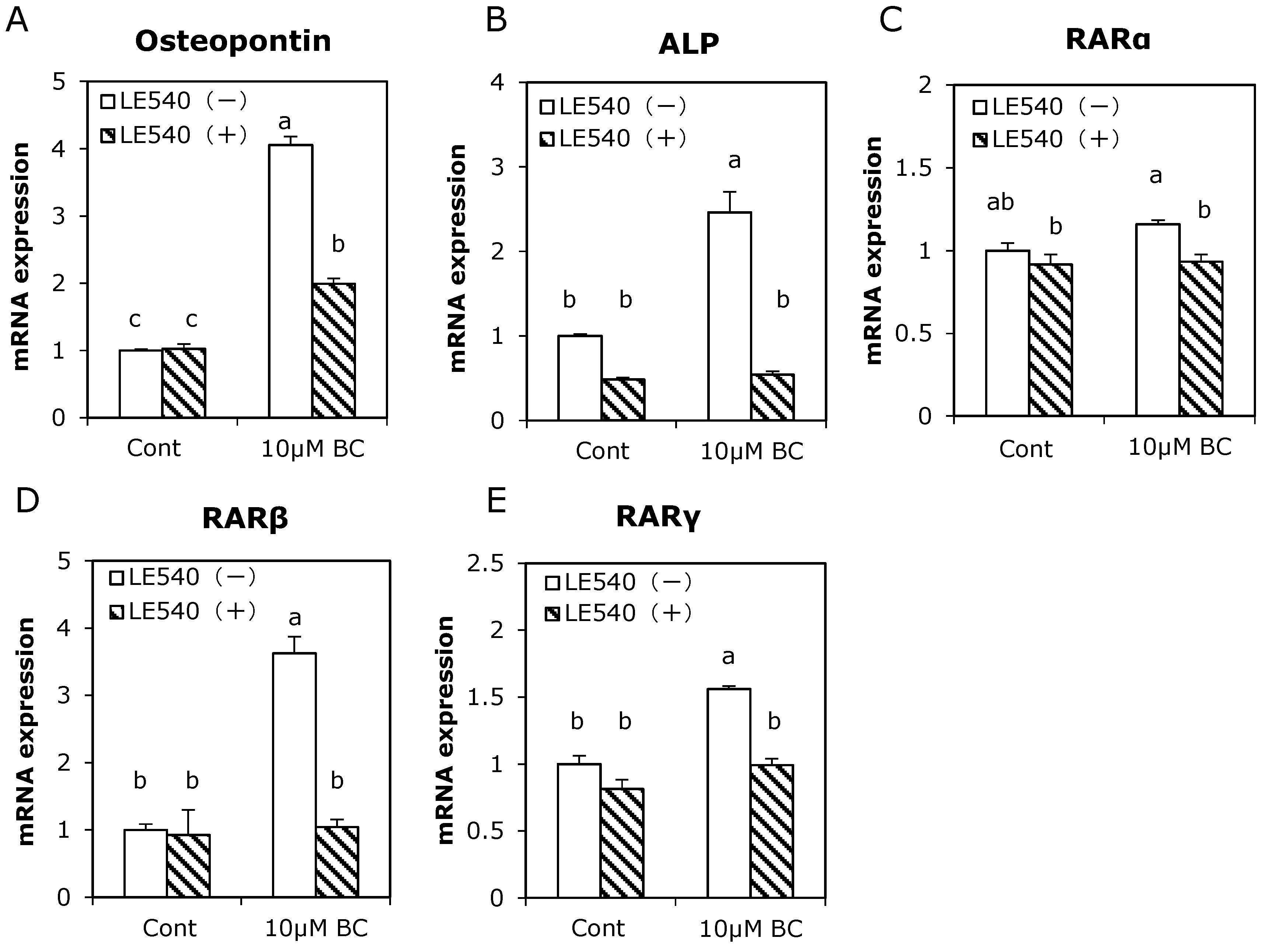
3.4. Effect of RAR Pan-Antagonist on Osteoblast-Related Gene Expression Induced by Treatment with β-Carotene

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wong, W.W.; Lewis, R.D.; Steinberg, F.M.; Murray, M.J.; Cramer, M.A.; Amato, P.; Young, R.L.; Barnes, S.; Ellis, K.J.; Shypailo, R.J.; et al. Soy isoflavone supplementation and bone mineral density in menopausal women: A 2-y multicenter clinical trial. Am. J. Clin. Nutr. 2009, 90, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Yang, H.P.; Yang, H.T.; Yang, T.C.; Shieh, M.J.; Huang, S.Y. One-year soy isoflavone supplementation prevents early postmenopausal bone loss but without a dose-dependent effect. J. Nutr. Biochem. 2006, 17, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.; Lydeking-Olsen, E. Dietary phytoestrogens and their effect on bone: Evidence from in vitro and in vivo, human observational, and dietary intervention studies. Am. J. Clin. Nutr. 2003, 78, 593–609. [Google Scholar]
- Pan, W.; Quarles, L.D.; Song, L.H.; Yu, Y.H.; Jiao, C.; Tang, H.B.; Jiang, C.H.; Deng, H.W.; Li, Y.J.; Zhou, H.H.; et al. Genistein stimulates the osteoblastic differentiation via NO/cGMP in bone marrow culture. J. Cell Biochem. 2005, 94, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, E.; Yamaguchi, M. Stimulatory effect of Daidzein in osteoblastic MC3T3-E1 cells. Biochem. Pharmacol. 2000, 59, 471–475. [Google Scholar] [CrossRef]
- Rassi, C.M.; Lieberherr, M.; Chaumaz, G.; Pointillart, A.; Cournot, G. Down-regulation of osteoclast differentiation by daidzein via caspase 3. J. Bone Miner. Res. 2002, 17, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.H.; Yamaguchi, M. Suppressive effect of genistein on rat bone osteoclasts: Apoptosis is induced through Ca2+ signaling. Biol. Pharm. Bull. 1999, 22, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.S. Absorption, metabolism, and transport of carotenoids. FASEB J. 1996, 10, 542–551. [Google Scholar] [PubMed]
- Eroglu, A.; Hruszkewycz, D.P.; dela Sena, C.; Narayanasamy, S.; Riedl, K.M.; Kopec, R.E.; Schwartz, S.J.; Curley, R.W., Jr.; Harrison, E.H. Naturally occurring eccentric cleavage products of provitamin A β-carotene function as antagonists of retinoic acid receptors. J. Biol. Chem. 2012, 287, 15886–15895. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M.; Nakamura, M.; Ogawa, K.; Ikoma, Y.; Yano, M. High serum carotenoids associated with lower risk for bone loss and osteoporosis in post-menopausal Japanese female subjects: Prospective cohort study. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.; Hannan, M.T.; Blumberg, J.; Cupples, L.A.; Kiel, D.P.; Tucker, K.L. Protective effect of total carotenoid and lycopene intake on the risk of hip fracture: A 17-year follow-up from the Framingham Osteoporosis Study. J. Bone Miner. Res. 2009, 24, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Ishimi, Y.; Ohmura, M.; Yamaguchi, M.; Ikegami, S. Vitamin A and carotenoids stimulate differentiation of mouse osteoblastic cells. J. Nutr. Sci. Vitaminol. 1997, 43, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, S.; Yamaguchi, M. Anabolic effect of beta-cryptoxanthin in osteoblastic MC3T3-E1 cells is enhanced with 17beta-estradiol, genistein, or zinc sulfate in vitro: the unique effect with zinc on Runx2 and alpha1(I) collagen mRNA expressions. Mol. Cell Biochem. 2008, 307, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Tadaishi, M.; Nishide, Y.; Tousen, Y.; Kruger, M.C.; Ishimi, Y. Cooperative effects of soy isoflavones and carotenoids on osteoclast formation. J. Clin. Biochem. Nutr. 2014, 54, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Sakai, A.; Mori, T.; Tanaka, S.; Moridera, K.; Nakamura, T. The anabolic action of intermittent PTH in combination with cathepsin K inhibitor or alendronate differs depending on the remodeling status in bone in ovariectomized mice. Bone 2009, 44, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, I.; Yamaguchi, T.; Yano, S.; Yamauchi, M.; Yamamoto, M.; Sugimoto, T. Adiponectin and AMP kinase activator stimulate proliferation, differentiation, and mineralization of osteoblastic MC3T3-E1 cells. BMC Cell Biol. 2007, 8. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, H.; Hao, W.; Fujita, Y.; Funayama, A.; Miyauchi, Y.; Hashimoto, K.; Miyamoto, K.; Iwasaki, R.; Sato, Y.; Kobayashi, T.; et al. Aldehyde-stress resulting from Aldh2 mutation promotes osteoporosis due to impaired osteoblastogenesis. J. Bone Miner. Res. 2012, 27, 2015–2023. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, T.; Kida, K.; Yamaguchi, A.; Hata, K.; Ichida, F.; Meguro, H.; Aburatani, H.; Nishimura, R.; Yoneda, T. BMP2 regulates Osterix through Msx2 and Runx2 during osteoblast differentiation. J. Biol. Chem. 2008, 283, 29119–29125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.K.; Uemura, T.; Nemoto, A.; Yabe, T.; Fujii, N.; Ushida, T.; Tateishi, T. Osteopontin involvement in integrin-mediated cell signaling and regulation of expression of alkaline phosphatase during early differentiation of UMR cells. FEBS Lett. 1997, 420, 112–116. [Google Scholar] [CrossRef]
- Collins, S.J.; Robertson, K.A.; Mueller, L. Retinoic acid-induced granulocytic differentiation of HL-60 myeloid leukemia cells is mediated directly through the retinoic acid receptor (RAR-alpha). Mol. Cell Biol. 1990, 10, 2154–2163. [Google Scholar] [CrossRef] [PubMed]
- Kanno, S.; Hirano, S.; Kayama, F. Effects of phytoestrogens and environmental estrogens on osteoblastic differentiation in MC3T3-E1 cells. Toxicology 2004, 196, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Castelo-Branco, C.; Cancelo Hidalgo, M.J. Isoflavones: Effects on bone health. Climacteric 2011, 14, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, R.; Inoue, D.; Shibata, M.; Saika, M.; Kido, S.; Ooka, H.; Tomiyama, H.; Sakamoto, Y.; Matsumoto, T. Estrogen promotes early osteoblast differentiation and inhibits adipocyte differentiation in mouse bone marrow stromal cell lines that express estrogen receptor (ER) alpha or beta. Endocrinology 2002, 143, 2349–2356. [Google Scholar] [PubMed]
- Chen, W.F.; Wong, M.S. Genistein modulates the effects of parathyroid hormone in human osteoblastic SaOS-2 cells. Brit. J. Nutr. 2006, 95, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Ernst, M.; Schmid, C.; Froesch, E.R. Enhanced osteoblast proliferation and collagen gene expression by estradiol. Proc. Natl. Acad. Sci. USA 1988, 85, 2307–2310. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.; Clerici, C.; Lephart, E.D.; Cole, S.J.; Heenan, C.; Castellani, D.; Wolfe, B.E.; Nechemias-Zimmer, L.; Brown, N.M.; Lund, T.D.; et al. S-equol, a potent ligand for estrogen receptor beta, is the exclusive enantiomeric form of the soy isoflavone metabolite produced by human intestinal bacterial flora. Am. J. Clin. Nutr. 2005, 81, 1072–1079. [Google Scholar] [PubMed]
- Wang, J.; Xu, J.; Wang, B.; Shu, F.R.; Chen, K.; Mi, M.T. Equol promotes rat osteoblast proliferation and differentiation through activating estrogen receptor. Genet. Mol. Res. 2014, 13, 5055–5063. [Google Scholar] [CrossRef] [PubMed]
- Hisada, K.; Hata, K.; Ichida, F.; Matsubara, T.; Orimo, H.; Nakano, T.; Yatani, H.; Nishimura, R.; Yoneda, T. Retinoic acid regulates commitment of undifferentiated mesenchymal stem cells into osteoblasts and adipocytes. J. Bone Miner. Metab. 2013, 31, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, S.; Kojima, F.; Momose, I.; Kawada, M.; Adachi, H.; Nishimura, Y. Decalpenic acid induces early osteoblastic markers in pluripotent mesenchymal cells via activation of retinoic acid receptor γ. Biochem. Biophys. Res. Commun. 2012, 422, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Lind, T.; Sundqvist, A.; Hu, L.; Pejler, G.; Andersson, G.; Jacobson, A.; Melhus, H. Vitamin A is a negative regulator of osteoblast mineralization. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishide, Y.; Tousen, Y.; Tadaishi, M.; Inada, M.; Miyaura, C.; Kruger, M.C.; Ishimi, Y. Combined Effects of Soy Isoflavones and β-Carotene on Osteoblast Differentiation. Int. J. Environ. Res. Public Health 2015, 12, 13750-13761. https://doi.org/10.3390/ijerph121113750
Nishide Y, Tousen Y, Tadaishi M, Inada M, Miyaura C, Kruger MC, Ishimi Y. Combined Effects of Soy Isoflavones and β-Carotene on Osteoblast Differentiation. International Journal of Environmental Research and Public Health. 2015; 12(11):13750-13761. https://doi.org/10.3390/ijerph121113750
Chicago/Turabian StyleNishide, Yoriko, Yuko Tousen, Miki Tadaishi, Masaki Inada, Chisato Miyaura, Marlena C. Kruger, and Yoshiko Ishimi. 2015. "Combined Effects of Soy Isoflavones and β-Carotene on Osteoblast Differentiation" International Journal of Environmental Research and Public Health 12, no. 11: 13750-13761. https://doi.org/10.3390/ijerph121113750