Bioprocessing Data for the Production of Marine Enzymes

Abstract
:1. Introduction
2. Batch Processes
2.1. Submerged processes
2.1.1. Proteases
2.1.2. Chitinases
2.1.3. DNA polymerases
2.1.4. Agarases
2.1.5. Haloperoxidases
2.1.6. Amylase
2.1.7. Multiple enzymatic activities
2.1.8. Superoxide dismutase
2.1.9. Other enzymes
2.2. Immobilized processes
2.2.1. Proteases
2.2.2. Peroxidase
2.3. Solid-state processes
2.3.1. Use of organic media
2.3.2. Use of inert media
2.3.3. Advantages and disadvantages of solid-state processes
3. Fed-batch Processes
3.1. Sulfite oxidase
3.2. Protease
3.3. Cellulase and xylanse
3.4. Glutamate dehydrogenase
3.5. ADP ribosyl cyclase
4. Continuous Processes
4.1. Suspended cell systems
4.2. Immobilized cell systems
5. Shake Flask Cultivations
6. Conclusions and Future Directions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Pomponi, SA. The bioprocess–Technological potential of the sea. J Biotechnol 1999, 70, 5–13. [Google Scholar]
- Ghosh, D; Saha, M; Sana, B; Mukherjee, J. Marine Enzymes. Adv Biochem Eng Biotechnol 2005, 96, 189–218. [Google Scholar]
- Lang, S; Huners, M; Verena, L. Bioprocess engineering data on the cultivation of marine prokaryotes and fungi. Adv Biochem Eng Biotechnol 2005, 97, 29–62. [Google Scholar]
- Shluer, ML; Kargi, F. Bioprocess Engineering: Basic Concepts, 2nd ed; Prentice Hall of India: New Delhi, India, 2003; pp. 245–301. [Google Scholar]
- Blanch, HW; Clark, DS. Biochemical Engineering; Marcel Dekker: New York, NY, USA, 1996; pp. 276–331. [Google Scholar]
- Niehaus, F; Bertoldo, C; Kahler, M; Antranikian, G. Extremophiles as a source of novel enzymes for industrial application. Appl Microbiol Biot 1999, 51, 711–729. [Google Scholar]
- Estrada-Badillo, C; Marquez-Rocha, FJ. Effect of agitation rate on biomass and protease production by a marine bacterium Vibrio harveyi cultured in a fermentor. World J Microb Biot 2003, 19, 129–133. [Google Scholar]
- Kumar, CG; Joo, HS; Koo, YM; Paik, SR; Chang, CS. Thermostable alkaline protease from a novel marine haloalkalophilic Bacillus clausii isolate. World J Microb Biot 2004, 20, 351–357. [Google Scholar]
- Narinx, E; Baise, E; Gerday, C. Subtilisin from psychrophilic Antarctic bacteria: Characterization and site-directed mutagenesis of residues possibly involved in the adaptation to cold. Protein Eng 1997, 10, 1271–1279. [Google Scholar]
- Weiner, RM; Shi, J; Coyne, VE. Thermostable alkaline metalloprotease produced by a hyphomonas and preparation thereof. US Patent No 5,589,373, 1996. [Google Scholar]
- Fenice, M; Leuba, JL; Federici, F. Chitinolytic enzyme activity of Penicillium janthinellum P9 in bench-top bioreactor. J Ferment Technol 1998, 86, 620–623. [Google Scholar]
- Kao, PM; Chen, CI; Huang, SC; Chang, YC; Tsai, PJ; Liu, YC. Effects of shear stress and mass transfer on chitinase production by Paenibacillus sp. CHE-N1. Biochem Eng J 2007, 34, 172–178. [Google Scholar]
- Liu, BL; Kao, PM; Tzeng, YM; Feng, KC. Production of chitinase from Verticillium lecanii F091 using submerged fermentation. Enzyme Microb Tech 2003, 33, 410–415. [Google Scholar]
- Gelfand, DH; Lawyer, FC; Stoffel, S. Purified thermostable nucleic acid polymerase enzyme from Thermotoga maritime. US Patent No 5,624,833, 1997. [Google Scholar]
- Baheri, HR; Hill, GA; Roesler, WJ. Modelling plasmid instability in batch and continuous fermentors. Biochem Eng J 2001, 8, 45–50. [Google Scholar]
- Slater, MR; Huang, F; Hartnett, JR. Thermophilic DNA polymerases from Thermotoga neapolitana. U.S. Patent No 6,001,645, 1999. [Google Scholar]
- Sugano, Y; Nagae, H; Inagaki, K; Yamamoto, T; Terada, I; Yamazaki, Y. Production and characteristics of some new β-agarases from a marine bacterium, Vibrio sp. strain JT0107. J Ferment Technol 1995, 79, 549–554. [Google Scholar]
- Byun, DS; Kim, DS; Godber, JS; Nam, SW; Oh, MJ; Shim, HS; Kim, HR. Isolation and characterization of marine bacterium producing arylsulfatase. J Microbiol Biotechn 2004, 14, 1134–1141. [Google Scholar]
- Rorrer, GL; Tucker, MP; Cheney, DP; Maliakal, S. Bromoperoxidase activity in microplantlet suspension cultures of the macrophytic red alga Ochtodes secundiramea. Biotechnol Bioeng 2001, 74, 389–395. [Google Scholar]
- Posten, C. Design principles of photo-bioreactors for cultivation of microalgae. Eng Life Sci 2009, 9, 165–177. [Google Scholar]
- Wong, BL; Shen, YQ; Chen, YP. Halogenation of cephalexin with haloperoxidase from Rathayibacter biopuresis. U.S. Patent No 5,589,354, 1996. [Google Scholar]
- Wong, BL; Shen, YQ; Chen, YP. Chlorination of cephalexin with chloroperoxidase from Rathayibacter biopuresis. US Patent No 5,695,951, 1997. [Google Scholar]
- Wong, BL; Shen, YQ; Chen, YP. Rathayibacter biopuresis produces cephalosporin haloperoxidase. US Patent No 5,854,054, 1998. [Google Scholar]
- Li, H; Chi, Z; Duan, X; Wang, L; Sheng, J; Wu, L. Glucoamylase production by the marine yeast Aureobasidium pullulans N13d and hydrolysis of potato starch granules by the enzyme. Process Biochem 2007, 42, 462–465. [Google Scholar]
- Gohel, V; Maisuria, V; Chhatpar, HS. Utilization of various chitinous sources for production of mycolytic enzymes by Pantoea dispersa in bench-top fermenter. Enzyme Microb Technol 2007, 40, 1608–1614. [Google Scholar]
- Weiner, RM; Hutcheson, SW; Abdel-Hafez, AAW; Howard, M; Taylor, LE; Ekborg, NA. Alginases, systems containing alginases and methods of cloning, purifying and/or utilizing alginases. US Patent No 7,439,034, 2008. [Google Scholar]
- Orozco, MR; Hernandez-Saavedra, NY; Valle, FA; Gonzalez, BA; Ochoa, JL. Cell yield and superoxide dismutase activity of the marine yeast Debaryomyces hansenii under different culture conditions. J Mar Biotechnol 1998, 6, 255–259. [Google Scholar]
- Slater, MR; Huang, F; Hartnett, JR; Bolchakova, E; Storts, DR; Otto, P; Miller, KM; Novikov, A; Velikodvorskaya, GA. Thermophilic DNA polymerases from Thermotoga neapolitana. U.S. Patent No 6,077,664, 2000. [Google Scholar]
- Daniels, EG. Process for producing prostaglandin acids. US Patent No 3,840,434, 1974. [Google Scholar]
- Müller, WEG; Wiens, M; Batel, R; Steffen, R; Schröder, HC; Borojevic, R; Custodio, MR. Establishment of a primary cell culture from a sponge: Primmorphs from Suberites domuncula. Mar Ecol Prog Ser 1999, 178, 205–219. [Google Scholar]
- Moll, BA. Methods and compositions for producing metabolic products for algae. US Patent No 5,270,175, 1993. [Google Scholar]
- Qureshi, MH; Kato, C; Horikoshi, K. Purification of a ccb-type quinol oxidase specifically induced in a deep-sea barophilic bacterium, Shewanella sp. strain DB-172F. Extremophiles 1998, 2, 93–99. [Google Scholar]
- Elibol, M; Moreira, AR. Production of extracellular alkaline protease by immobilization of the marine bacterium Teredinobacter turnirae. Process Biochem 2003, 38, 1445–1450. [Google Scholar]
- Beshay, U; Moreira, A. Repeated batch production of alkaline protease using porous sintered glass as carriers. Process Biochem 2003, 38, 1463–1469. [Google Scholar]
- Beshay, U; Moreira, A. Protease production from marine microorganism by immobilized cells. AIChE Ann Meet Conf Proc 2004, 8717–8729. [Google Scholar]
- Irvine, RL; Venkatadri, R. Method of producing extracellular products from aerobic microorganisms. US Patent No 5,342,765, 1994. [Google Scholar]
- Hölker, U; Lenz, J. Solid-state fermentation — are there any biotechnological advantages. Curr Opin Microbiol 2005, 8, 301–306. [Google Scholar]
- Ooijkaas, LP; Weber, FJ; Buitelaar, RM; Tramper, J; Rinzema, A. Defined media and inert supports: Their potential as solid-state fermentation production systems. Trends Biotechnol 2000, 18, 356–360. [Google Scholar]
- Nagendra Prabhu, G; Chandrasekaran, M. L-glutaminase production by marine Vibrio costicola under solid-state fermentation using different substrates. J Mar Biotechnol 1996, 4, 176–179. [Google Scholar]
- Suresh, PV; Chandrasekaran, M. Impact of process parameters on chitinase production by an alkalophilic marine Beauveria bassiana in solid state fermentation. Process Biochem 1999, 34, 257–267. [Google Scholar]
- Suresh, PV; Chandrasekaran, M. Utilization of prawn waste for chitinase production by the marine fungus Beauveria bassiana by solid state fermentation. World J Microb Biot 1998, 14, 655–660. [Google Scholar]
- Chellappan, S; Jasmin, C; Basheer, SM; Elyas, KK; Bhat, SG; Chandrasekaran, M. Production, purification and partial characterization of a novel protease from marine Engyodontium album BTMFS10 under solid state fermentation. Process Biochem 2006, 41, 956–961. [Google Scholar]
- Elibol, M; Moreira, AR. Optimizing some factors affecting alkaline protease production by a marine bacterium Teredinobacter turnirae under solid substrate fermentation. Process Biochem 2005, 40, 1951–1956. [Google Scholar]
- Sheng, J; Chi, Z; Yan, K; Wang, X; Gong, F; Li, J. Use of response surface methodology for optimizing process parameters for high inulinase production by the marine yeast Cryptococcus aureus G7a in solid-state fermentation and hydrolysis of inulin. Bioprocess Biosyst Eng 2009, 32, 333–339. [Google Scholar]
- Prabhu, GN; Chandrasekaran, M. Polystyrene-An inert carrier for L-glutaminase production by marine Vibrio costicola under solid-state fermentation. World J Microb Biot 1995, 11, 683–684. [Google Scholar]
- Nagendra Prabhu, G; Chandrasekaran, M. Impact of process parameters on L-glutaminase production by marine Vibrio costicola in solid state fermentation using polystyrene as an inert support. Process Biochem 1997, 32, 285–289. [Google Scholar]
- Sabu, A; Keerthi, TR; Kumar, SR; Chandrasekaran, M. L-glutaminase production by marine Beauveria sp. under solid state fermentation. Process Biochem 2000, 35, 705–710. [Google Scholar]
- Longobardi, GP. Fed-batch versus batch fermentation. Bioprocess Eng 1994, 10, 185–194. [Google Scholar]
- Gu, MB; Park, MH; Kim, DI. Growth rate control in fed-batch cultures of recombinant Saccharomyces cerevisiae producing hepatitis B surface antigen (HBsAg). Appl Microbiol Biotechnol 1991, 35, 46–50. [Google Scholar]
- Muffler, K; Ulber, R. Fed-batch cultivation of the marine bacterium Sulfitobacter pontiacus using immobilized substrate and purification of sulfite oxidase by application of membrane adsorber technology. Biotechnol Bioeng 2008, 99, 870–875. [Google Scholar]
- Beshay, U; Moreira, A. Production of alkaline protease with Teredinobacter turnirae in controlled fed-batch fermentation. Biotechnol Lett 2005, 27, 1457–1460. [Google Scholar]
- Wicher, KB; Holst, OP; Hachem, MYA; Karlsson, EMN; Hreggvidsson, GO. Thermostable cellulase. US Patent No 6,812,018, 2004. [Google Scholar]
- Hreggvidsson, GO; Kaiste, E; Holst, O; Eggertsson, G; Palsdottir, A; Kristjansson, JK. An extremely thermostable cellulase from the thermophilic eubacterium Rhodothermus marinus. Appl Environ Microbiol 1996, 62, 3047–3049. [Google Scholar]
- Karlsson, EN; Holst, O; Tocaj, A. Efficient production of truncated thermostable xylanases from Rhodothermus marinus in Escherichia coli fed-batch cultures. J Biosci Bioeng 1999, 87, 598–606. [Google Scholar]
- Terashima, C; Uchida, K; Matsukawa, H; Oka, O; Fujita, T; Imanaka, T. Thermostable glutamate dehydrogenase and process for producing the same. US Patent No 6,162,626, 2000. [Google Scholar]
- Munshi, C; Hon, CL. High-level expression of recombinant aplysia ADP-ribosyl cyclase in Pichia pastoris by fermentation. Protein Expres Purif 1997, 11, 104–110. [Google Scholar]
- Kao, PM; Huang, SC; Chang, YC; Liu, YC. Development of continuous chitinase production process in a membrane bioreactor by Paenibacillus sp. CHE-N1. Process Biochem 2007, 42, 606–611. [Google Scholar]
- Kelly, RM; Brown, SH; Costantino, HR. Saccharification enzymes from hyperthermophilic bacteria and processes for their production. US Patent No 6,355,467, 2002. [Google Scholar]
- Sabu, A; Sukumaran, RK; Muthusamy, C. Continuous production of extracellular L-glutaminase by Ca-alginate-immobilized marine Beauveria bassiana BTMF S-10 in packed-bed reactor. Appl Biochem Biotechnol 2002, 71, 71–79. [Google Scholar]
- Kumar, SR; Chandrasekaran, M. Continuous production of L-glutaminase by an immobilized marine Pseudomonas sp. BTMS-51 in a packed bed reactor. Process Biochem 2003, 38, 1431–1436. [Google Scholar]
- Makowski, K; Białkowska, A; Szczesna-Antczak, M; Kalinowska, H; Kur, J; Cieśliński, H; Turkiewicz, M. Immobilized preparation of cold-adapted and halotolerant Antarctic β-galactosidase as a highly stable catalyst in lactose hydrolysis. FEMS Microbiol Ecol 2007, 59, 535–542. [Google Scholar]
- Kennedy, MJ; Reader, SL; Davies, RJ; Rhoades, DA; Silby, HW. The scale up of mycelial shake flask fermentations: A case study gamma linolenic acid production by Mucor hiemalis IRL 51. J Ind Microbiol 1994, 13, 212–216. [Google Scholar]
- Yang, JK; Shih, IL; Tzeng, YM; Wang, SL. Production and purification of protease from a Bacillus subtilis that can deproteinize crustacean wastes. Enzyme Microb Tech 2000, 26, 406–413. [Google Scholar]
- Young, DM. Thermostable proteolytic enzymes and uses thereof in peptide and protein synthesis. US Patent No 7,192,730, 2007. [Google Scholar]
- Chi, Z; Ma, C; Wang, P; Li, HF. Optimization of medium and cultivation conditions for alkaline protease production by the marine yeast Aureobasidium pullulans. Bioresour Technol 2007, 98, 534–538. [Google Scholar]
- Abusham, RA; Rahman, RNZRA; Salleh, A; Basri, M. Optimization of physical factors affecting the production of thermo-stable organic solvent-tolerant protease from a newly isolated halotolerant Bacillus subtilis strain Rand. Microb Cell Fact 2009, 8. [Google Scholar] [CrossRef]
- Vázquez, JA; Docasal, SF; Mirón, J; González, MP; Murado, MA. Proteases production by two Vibrio species on residuals marine media. J Ind Microbiol Biot 2006, 33, 661–668. [Google Scholar]
- Fujishima, S; Yamano, N; Maruyama, A; Higashihara, T. Chitinase and method for preparing the same. US Patent No 6,352,850, 2002. [Google Scholar]
- Gohel, V; Chaudhary, T; Vyas, P; Chhatpar, HS. Statistical screenings of medium components for the production of chitinase by the marine isolate Pantoea dispersa. Biochem Eng J 2006, 28, 50–56. [Google Scholar]
- Han, Y; Li, Z; Miao, X; Zhang, F. Statistical optimization of medium components to improve the chitinase activity of Streptomyces sp. Da11 associated with the South China Sea sponge Craniella australiensis. Process Biochem 2008, 43, 1088–1093. [Google Scholar]
- Patel, B; Gohel, V; Raol, B. Statistical optimisation of medium components for chitinase production by Paenibacillus sabina strain JD2. Ann Microbiol 2007, 57, 589–597. [Google Scholar]
- Aytekin, O; Elibol, M. Cocultivation of Lactococcus lactis and Teredinobacter turnirae for biological chitin extraction from prawn waste. Bioprocess Biosyst Eng 2010, 33, 393–399. [Google Scholar]
- D’Souza, DT; Tiwari, R; Sah, AK; Raghukumar, C. Enhanced production of laccase by a marine fungus during treatment of colored effluents and synthetic dyes. Enzyme Microb Technol 2006, 38, 504–511. [Google Scholar]
- D’Souza-Ticlo, D; Verma, AK; Mathew, M; Raghukumar, C. Effect of nutrient nitrogen on laccase production, its isozyme pattern and effluent decolorization by the fungus NIOCC #2a, isolated from mangrove wood. Indian J Mar Sci 2006, 35, 364–372. [Google Scholar]
- Raghukumar, C; Rivonkar, G. Decolorization of molasses spent wash by the white-rot fungus Flavodon flavus, isolated from a marine habitat. Appl Microbiol Biotechnol 2001, 55, 510–514. [Google Scholar]
- Bonugli-Santos, RC; Durrant, LR; Silva, MD; Sette, LD. Production of laccase, manganese peroxidase and lignin peroxidase by Brazilian marine-derived fungi. Enzyme Microb Technol 2010, 46, 32–37. [Google Scholar]
- Raghukumar, C; D’ Souza, TM; Thorn, RG; Reddy, CA. White rot-lignin-modifying fungus Flavodon flavus and a process for removing dye from dye containing water or soil using the fungus. US Patent No 6,395,534, 2002. [Google Scholar]
- Ping, R; Sun, M; Liu, J; Wang, Y; Hao, J; Zhang, S. Optimization of fermentation conditions for marine Bacillus licheniformis MP-2 esterase by response surface methodology. Chin J Appl Environ Biol 2008, 14, 548–552. [Google Scholar]
- Robertson, DE; Murphy, D; Reid, J; Maffia, AM; Link, S; Swanson, RV; Warren, PV; Kosmotka, A. Esterases. US Patent No 5,942,430, 1999. [Google Scholar]
- Kiran, GS; Shanmughapriya, S; Jayalakshmi, J; Selvin, J; Gandhimathi, R; Sivaramakrishnan, S; Arunkumar, M; Thangavelu, T; Natarajaseenivasan, K. Optimization of extracellular psychrophilic alkaline lipase produced by marine Pseudomonas sp. (MSI057). Bioprocess Biosyst Eng 2008, 31, 483–492. [Google Scholar]
- Shanmughapriya, S; Seghal, KG; Selvin, J; Gandhimathi, R; Bastin, BT; Manilal, A; Sujith, S. Optimization, production, and partial characterization of an alkalophilic amylase produced by sponge associated marine bacterium Halobacterium salinarum MMD047. Biotechnol Bioprocess Eng 2009, 14, 67–75. [Google Scholar]
- Ahuja, SK; Ferreira, GM; Moreira, AR. Production of an endoglucanase by the shipworm bacterium Teredinobacter turnirae. J Ind Microbiol Biotechnol 2004, 31, 41–47. [Google Scholar]
- Shanmughapriya, S; Kiran, GS; Selvin, J; Thomas, TA; Rani, C. Optimization, purification, and characterization of extracellular mesophilic alkaline cellulase from sponge-associated Marinobacter sp. MSI032. Appl Biochem Biotechnol 2009. [Google Scholar] [CrossRef]
- Wicher, KB; Holst, OP; Hachem, MYA; Karlsson, EMN; Hreggvidsson, GO. Thermostable cellulases. US Patent Appl No 20020102699, 2002. [Google Scholar]
- Xiaoting, FU; Hong, L; Moo, KS. Optimization of culturing condition and medium composition for the production of alginate lyase by a marine Vibrio sp. YKW-34. J Ocean Univ Chin 2008, 7, 97–102. [Google Scholar]
- Iyer, PV; Singhal, RS. Screening and selection of marine isolate for L-glutaminase production and media optimization using response surface methodology. Appl Biochem Biotechnol 2009, 159, 233–250. [Google Scholar]
- Nandakumar, R; Wakayama, M; Nagano, Y; Kawamura, T; Sakai, K; Moriguchi, M. Overexpression of salt-tolerant glutaminase from Micrococcus luteus K-3 in Escherichia coli and its purification. Protein Expr Purif 1999, 15, 155–161. [Google Scholar]
- Stougaard, P; Hansen, OC. Recombinant hexose oxidase, a method of producing same and use of such enzyme. US Patent No 7,544,795, 2009. [Google Scholar]
- Li, XY; Liu, ZQ; Chi, ZM. Production of phytase by a marine yeast Kodamaea ohmeri BG3 in an oats medium: Optimization by response surface methodology. Bioresour Technol 2008, 99, 6386–6390. [Google Scholar]
- Metz, JG; Weaver, CA; Barclay, WR; Flatt, JH. PUFA polyketide synthase systems and uses thereof. US Patent No 7,662,597, 2010. [Google Scholar]
- Roessler, PG; Matthews, TD; Ramseier, TM; Metz, JG. Product and process for transformation of Thraustochytriales microorganisms. US Patent No 7,001,772, 2006. [Google Scholar]
- Manyak, DM; Weiner, RM; Carlson, PS; Quintero, EJ. Preparation and use of biofilm-degrading, multiple-specificity, hydrolytic enzyme mixtures. US Patent No 6,759,040, 2004. [Google Scholar]
- Michelson, AM. Superoxide dismutase and process for production. US Patent No 3,997,402, 1976. [Google Scholar]
- Muller, WEG; Schroder, H; Krasko, A. Decomposition and modification of silicate and silicone by silase and use of the reversible enzyme. US Patent No 7,229,807, 2007. [Google Scholar]
- Mathur, EJ; Marsh, EJ; Schoettlin, WE. Purified thermostable Pyrococcus furiousus DNA ligase. US Patent No 5,700,672, 1997. [Google Scholar]
- Mandenius, CF; Brundin, A. Bioprocess optimization using design-of-experiments methodology. Biotechnol Prog 2008, 24, 1191–1203. [Google Scholar]
- Wright, PC; Stevenson, C; McEvoy, E; Burgess, JG. Opportunities for marine bioprocess intensification using novel bioreactor design: frequency of barotolerance in microorganisms obtained from surface waters. J Biotechnol 1999, 70, 343–349. [Google Scholar]
- Yan, L; Boyd, KG; Adams, DR; Burgess, JG. Biofilm specific cross-species induction of antimicrobial compounds in. Bacilli Appl Environ Microbiol 2003, 69, 3719–3727. [Google Scholar]
- Sarkar, S; Saha, M; Roy, D; Jaisankar, P; Das, S; Roy, LG; Gachhui, R; Sen, T; Mukherjee, J. Enhanced production of antimicrobial compounds by three salt-tolerant actinobacterial strains isolated from the Sundarbans in a niche-mimic bioreactor. Mar Biotechnol 2008, 10, 518–526. [Google Scholar]
- Sarkar, S; Mukherjee, J; Roy, D. Antibiotic production by a marine isolate (MS310) in an ultra-low-speed rotating disk bioreactor. Biotechnol Bioprocess Eng 2009, 14, 775–780. [Google Scholar]
- Sarkar, S; Roy, D; Mukherjee, J. Production of a potentially novel antimicrobial compound by a biofilm-forming marine Streptomyces sp. in a niche-mimic rotating disk bioreactor. Bioprocess Biosyst Eng 2010, 33, 207–217. [Google Scholar]
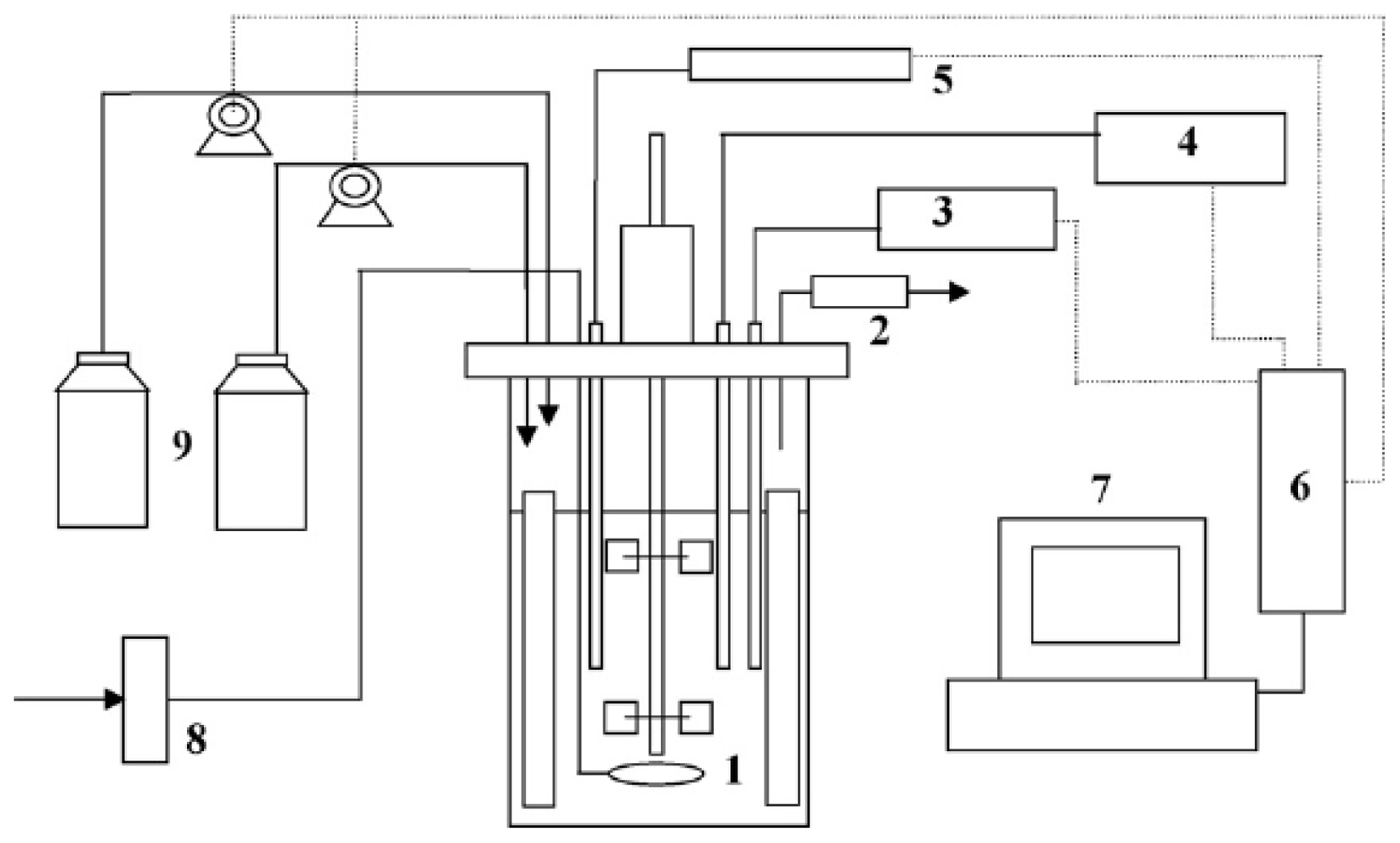



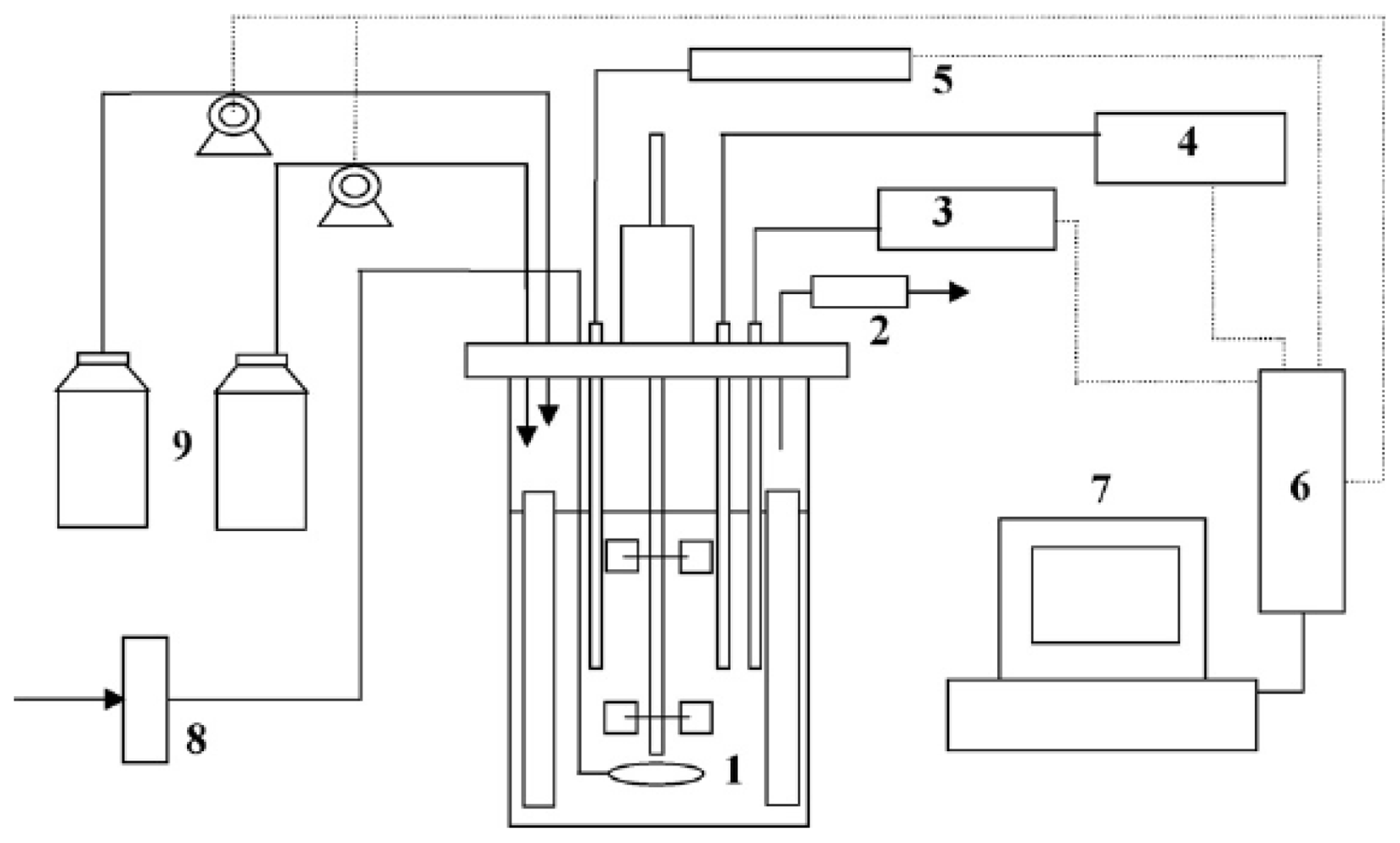{kind=link}
{kind=link}
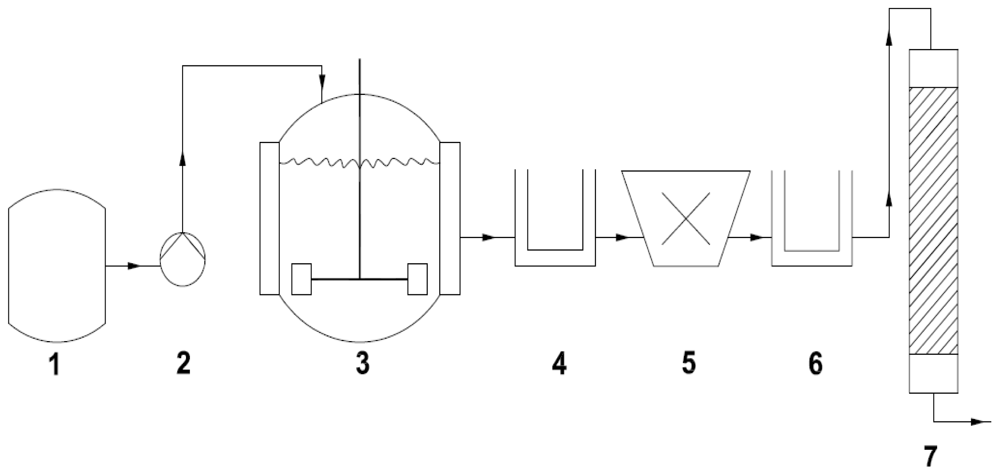{kind=link}
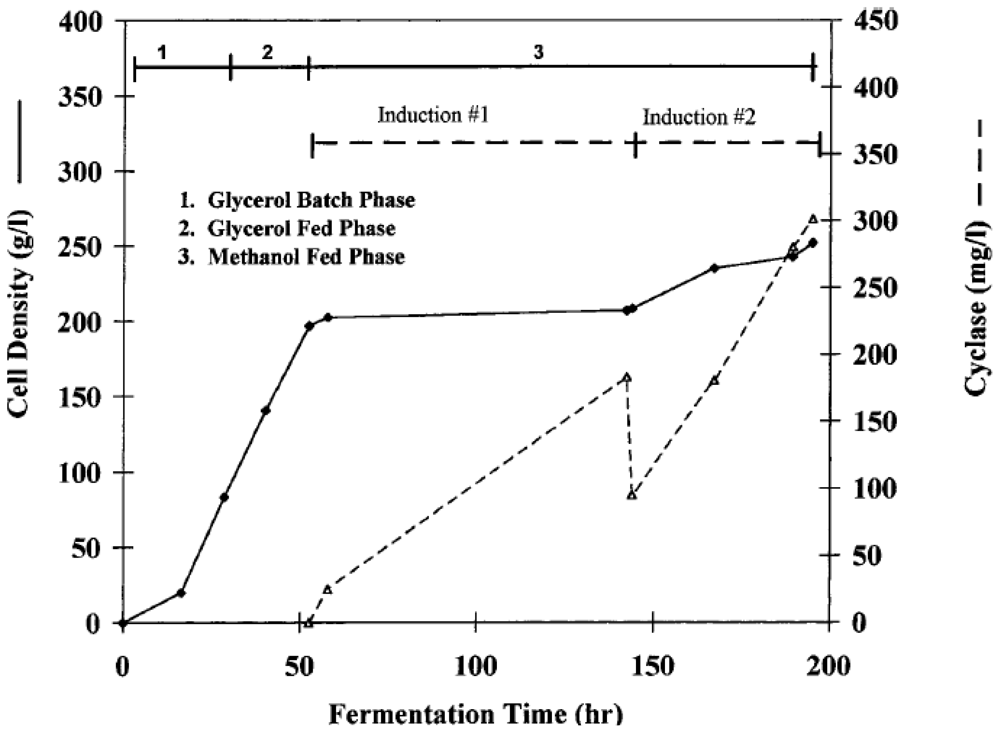{kind=link}
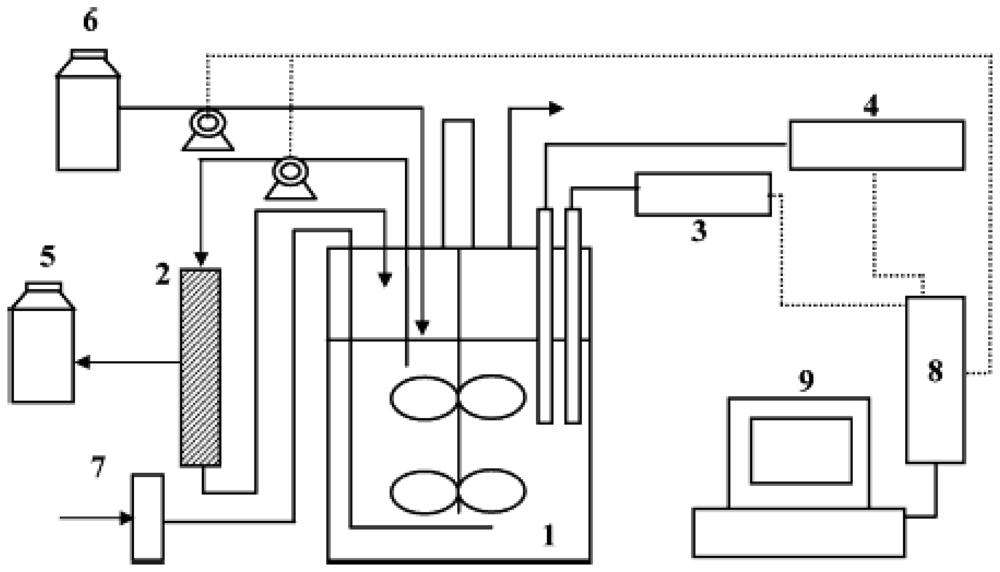{kind=link}
| Enzyme and source | Operational scale and working volume | Process parameters | Carbon source(s) | Enzyme activity | Reference | |||
|---|---|---|---|---|---|---|---|---|
| pH | Temperatu re (°C) | Aeration rate (L/L/min) and agitation rate (rpm) | Time | |||||
| Batch submerged processes | ||||||||
| Extracellular protease (Vibrio harveyi) | Applikon, (Holland) 1.5 L (reactor volume), 1.0 L (working volume) | 7.2 | 30.0 | 0.5, 700 | 9 h | Peptone 5 g/L, yeast extract 1 g/L supplemented with skim milk | 4.28 U/mg protein | [7] |
| Alkaline protease (Bacillus clausii No. 58) | Model KF5L, Kobiotech, Korea 5.0 L (reactor volume, 3.0 L (working volume) | 9.6 | 42.0 | 1.5, 400 | 40 h | Casein, corn starch, 0.5% (w/v) | 15,300 U/mL | [8] |
| Subtilisin, alkaline protease (Bacillus Strain TA39) | Model LH2000 10 L (working volume) | pH 7.6 | 4.0 and 25.0 | NR | 212.5 h | Bactopeptone 5 g/L, yeast extract 1 g/L | 6.85 U/mL | [9] |
| Alkaline metalloproteases (Hyphomonas jannaschiana) | NR | 7.6 | 37.0 | NR | 17 h | Carbohydrate based medium | NR | [10] |
| Chitinase (Penicillium janthineflum P9) | ADI 1020, Applikon (Holland) 3.0 L (reactor volume), 2.0 L (working volume) | 4.0 | 28.0 | 1.5, 500 | 7 d | Colloidal chitin 15 g/L, corn steep liquor 0.5 g/L | 686 U/L | [11] |
| Chitinase (Paenibacillus sp. CHE-N1) | BTF-A5L, Bio-Top Inc., Taiwan 5.0 L (reactor volume), 3.0 L (working volume), 10% preculture | 7.0 | 34.3 | 3.0, 200 | 56 h | Colloidal chitin 5 g/L, peptone 1 g/L | 11.8 U/mL | [12] |
| Chitinase (Verticillium lecanii F091) | 5.0 L (reactor volume, STR), 3.0 L (working volume), 10% preculture and 30.0 L (reactor volume, airlift bioreactor), 15.0 L (working volume) | 4.0 | 24.0 | 0.6, 150 and 0.9 | 6 d | Maltose 4.52%, marine peptone extract 1.79%, shrimp powder 0.4%, isolated soy protein 0.3% | 18.2 mU/mL and 19.9 mU/mL | [13] |
| Thermostable DNA polymerase (Thermotoga maritime) | 12.5 L (working volume) | 6.8 | 30.0 then shift to 35.0 | 2.0 | 25.5 h | Glucose10 g/L | 29.1 U/mg protein | [14] |
| Thermostable DNA polymerases (Thermotoga neapolitana) | Anaerobic process; 10.0 L (working volume) and 6.0 L (working volume) | NR | 75.0 and 37.0 | NR | 28 h and 5 h | Yeast extract, 10% and Luria broth (LB) containing 10 μg/mL tetracycline | 17 U/mL crude lysate | [16] |
| Agarase (Vibrio sp. Strain JT0107) | Marubishi Eng. Co., Tokyo 5.0 L (reactor volume), 3.0 L (working volume) | 8.0 | 25.0 | 0.5, 350 | 10 h | Polypeptone 20 g/L, yeast extract 4 g/L, agar 4 g/L | 625 U/L | [17] |
| Arylsulfatase (Sphingomonas sp. AS6330) | 10.0 L (reactor volume) | 7.0 | 30.0 | 1.0, 250 | 48 h | Sucrose 20 g/L | 1620 (U/mL) | [18] |
| Bromoperoxidase (Ochtodes secundiramea) | Planar bubble column photobioreactor 5.0 L (reactor volume), 3.0 L (working volume) | 8.0–8.5 | 22.0 | 0.71 | 70 d | Photoautotrophic | 1.9 μmol MCD/g DCW/min | [19] |
| Cephalexin haloperoxidase (Rathayibacter biopuresis) | 10.0 L (working volume, seed culture), inoculum size 1%–2% and 550.0 L (working volume, bioreactor) | 8.5 | 28.0 | NR | 20 h and 25 h | L-malic acid 0.05% | NR | [21–23] |
| Glucoamylase (Aureobasidium pullulans) | B. Braun, Biostat B2 (Germany) 2.0 L (reactor volume), 1.8 L (working volume) | 4.0 | 28 | 6.0, 250 | 56 | Soluble starch 1 g/100 mL sea water | 10 U/mL | [24] |
| Mycolytic enzymes (chitinases, proteases and glucanase) (Pantoea dispersa) | 5.0 L (reactor volume, 2.0 L (working volume) | 7.2 | 30.0 | 0.5, 300 | 72 h | Acid-swollen chitin, 5.0 g/L | 10.04 ± 0.19 U/mL | [25] |
| Alginases (Saccharophagus degradans) | Microferm New Brunswick Scientific (USA) 14.0 L (reactor volume), 8.0 L (working volume), 2% inoculum | 7.6 | 25.0 | 6.0, 400 | 36 | Glucose, alginate, xylose | 1690 alginase units | [26] |
| Superoxide dismutase (Debaryomyces hansenii Strain C- 11) | Microferm New Brunswick Scientific (USA) 2.0 L (reactor volume), 1.0 L (working volume), 1% inoculum | 5–7 | 40.0 | 5.0–7.0 | 24.5 | Glucose 2% | 400 U/mg protein | [27] |
| T th pyrophosphatase (Thermus thermophilus 111) | CHEMAP Ltd. (Switzerland) 450.0 L (working volume) | NR | 70.0 | 8.0, 4 | 18 | Disodium succinic acid 5.0 g/L, calcium succinic acid 0.5 g/L | 1760 U/mg protein | [28] |
| Esterase (Plexaura homomalla) | Extraction process, 30.0 L (working volume) | NR | 20.0–30.0 | Not required | 1 | Not required | NR | [29] |
| Silicatein (Suberites domuncula) | Aquarium volume 130.0 L | NR | 16 | NR | 7 weeks | Sea animal feeds | NR | [30] |
| Alcohol dehydrogenase, pyruvate decarboxylase (Ulvales) | Open pond 1.0 hectare | NR | NR | Not required | 15–20 d | Photoautotrophic | NR | [31] |
| Quinol oxidase (Shewanella sp. Strain DB-172F) | Pressurized vessel at 60 MPa | NR | 4 | NR | 6 d | Peptone 5g/L, Yeast extract 1g/L | 25.4 μmol/min/mg | [32] |
| Batch immobilized processes | ||||||||
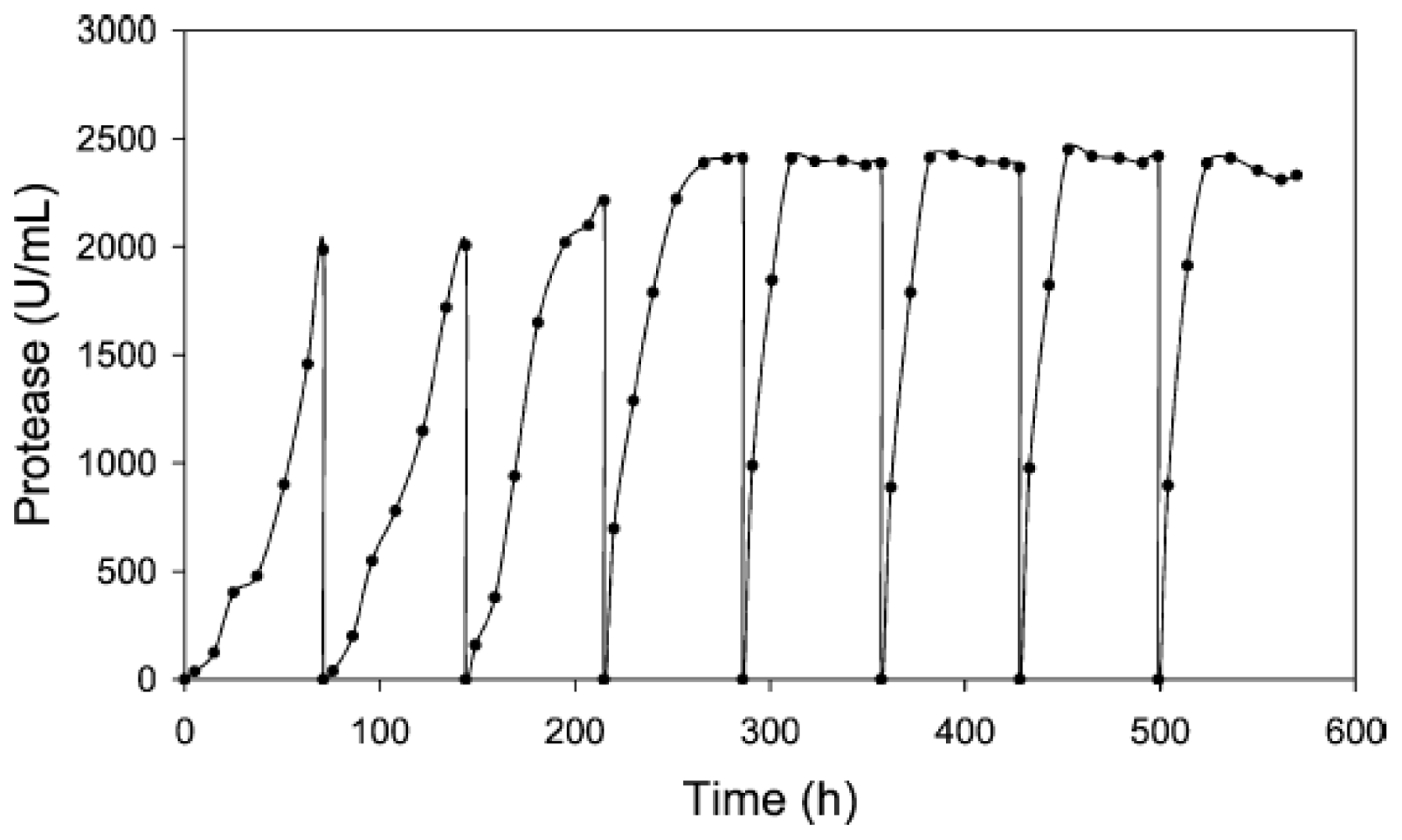
| Alkaline protease (Teredinobacter turnirae) | 250 mL Erlenmeyer flask | 8.0 | 30.0 | 0, 120 | 100 h (SB) 500 h (RB) | Sucrose 5 g/L | 2410 U/mL | [33] |
| Alkaline protease (Teredinobacter turnirae) | Immobilized process with glass carriers; 250 mL Erlenmeyer flask | 8.0 | 30.0 | 0, 120 | 50 h (SB) 16 d (RB) | Sucrose 5 g/L | 6605 U/mL | [34] |
| Lignin peroxidase (Caldariomyces fumago) | Immobilized process in silicone tubing; 2.5 L (reactor volume), 1.5 L (working volume) | NR | 37.0 | 0. 25, 40 | 3 d | Dextrose | 230 U/L | [36] |
| Batch solid-state processes | ||||||||
| L-glutaminase (Vibrio costicola) | 250 mL Erlenmeyer flask | 7.0 | 35.0 | NR | 24 h | L-glutamine | NR | [39] |
| Chitinase (Beauveria bassiana) | 250 mL Erlenmeyer flask | 9.2 | 28.0 | NR | 60 h | Chitin | 246.6 U/g IDS | [40] |
| Chitinase (Beauveria bassiana BTMF S10) | Petri plates (86 mm diameter and 17 mm height) | 9.5 | 27.0 | NR | 5 d | Prawn waste (23.08% chitin) | 248.0 U/g IDS | [41] |
| Protease (Engyodontium album BTMFS10) | 250 mL Erlenmeyer flask | 5.0 and 10.0 | 25.0 | NR | 5 d | Sucrose 0.1 M | 15, 912 U/g IDS | [42] |
| Alkaline protease (Teredinocabcter turnirae) | 250 mL Erlenmeyer flask | 7.34 | 30.0 | 0, 120 | 63 h | Soybean 1% | 1950 U/mL | [43] |
| Inulinase (Cryptococcus aureus G7a) | 250 mL Erlenmeyer flask | 5.5 | 29.0 | NR | 5 d | Inulin 20.0 g/L | 420.9 U/g IDS | [44] |
| L-glutaminase (Vibrio costicola) | Solid-state process on polystyrene beads, 250 mL Erlenmeyer flasks | 7.0 | 37.0 | NR | 36 h | Glucose | 88.0 U/g IDS | [45] |
| L-glutaminase (Vibrio costicola) | Solid-state process on polystyrene beads, 250 mL Erlenmeyer flasks | 7.0 | 35.0 | 0, 150 | 24 h | L-glutamine 3 % w/v, maltose 1% w/v | 196 U/g IDS | [46] |
| L-glutaminase (Beauveria sp.) | Solid-state process on polystyrene beads, 500 mL Erlenmeyer flasks | 9 | 27.0 | NR | 96 h | L-glutamine 0.25 % w/v, D-glucose 0.5% w/v | 49.89 U/mL | [47] |
| Enzyme and source | Bioreactor scale and working volume | Process parameters | Carbon source(s) | Enzyme activity | Reference | |||
|---|---|---|---|---|---|---|---|---|
| pH | Temperat ure (°C) | Aeration rate (L/L/min) and agitation rate (rpm) | Time | |||||
| Sulfite oxidase (Sulfitobacter pontiacus) | 10.0 L (working volume) | 7.3 | 26.0 | 0.4, 600 | 22 h | Peptone 5 g/L, yeast extract 1 g/L | 9.2 U/mg protein | [50] |
| Alkaline protease (Teredinobacter turnirae) | 2.0 L (working volume) | 8.0 | 30.0 | NR | 50 h | Sucrose 8 g/L | 8266 U/mL | [51] |
| Xylanase (Escherichia coli strain BL21DE3) | Chemoferm FLC- B-3 (Sweden) 3.0 L (reactor volume), 2.0 L (working volume) | 7.0 | 37.0 | DO maintained at 30% saturation | 14 h | Glucose 10.8 g/L | 245000 U/CDW | [54] |
| Glutamate dehydrogenase (Pyrococcos sp. KODI) | New Brunswick Scientific SF-116 (USA) 16.0 L (working volume) | 7.4 | 37.0 | 15.0, 800 | 10 h | Glucose | 300 U/mg protein | [55] |
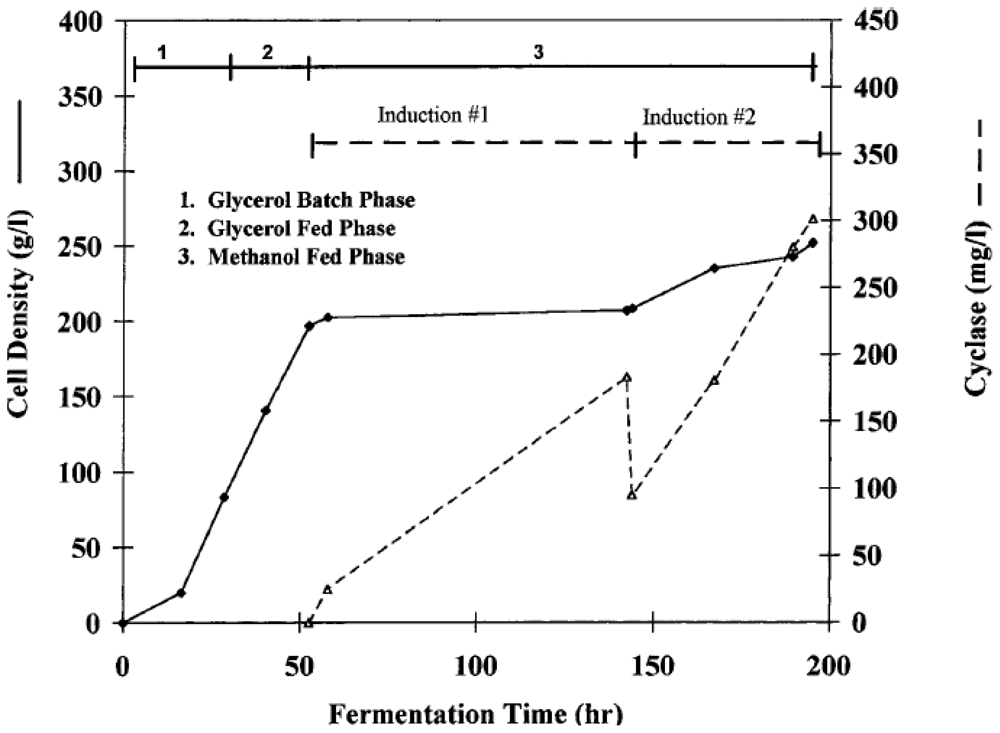
| ADP ribosyl cyclase (Aplysia californica) | Bioflo III 5.0 L (reactor volume) | NR | 28.0 | 1.14 | 200 h | Glycerol | 300 mg/L | [56] |
| Enzyme and source | Bioreactor scale and working volume | Process parameters | Carbon source(s) | Enzyme activity | Reference | |||
|---|---|---|---|---|---|---|---|---|
| pH | Temperature (°C) | Aeration rate (L/L/min) and agitation rate (rpm) | Time | |||||
| Chitinase (Paenibacillus sp. CHE-N1) | Biotop BTF-A5L (Taiwan) 5.0 L (reactor volume), 2.0 L (working volume) | 8.5 | 34.3 | 3, 200 | 132 h | Crab shell chitin powder 55.4 g/L | 42,800 mU/mL | [57] |
| α-glucosidase (Pyrococcus furiosus DSM 3638) | Anaerobic, 5.0 L (reactor volume), 2.0 L (working volume) | 6.8 | 98.0 | NR | NR | Yeast extract 0.1% | NR | [58] |
| L-glutaminase (Beauveria bassiana BTMF S-10) | Immobilized process; PBR, glass column 2.3 cm radius, 20.0 cm height | 9.0 | 27.0 | No aeration | 18 h | L-glutamine 0.25% w/v, D-glucose 0.5% w/v | 4.048 U/mL/h | [59] |
| L-glutaminase (Pseudomonas sp. BTMS-51) | Immobilized process; PBR, ID 3.6 cm, height 45.0 cm | 6.0 | 30.0 | NR | 120 h | L-glutamine 20 g/L, D-glucose 10 g/L | 13.49 U/mL/h | [60] |
| β-galactosidase (Pseudoalteromonas sp. 22b) | Immobilized process; PBR with substrate recycling | 7.6 | 15.0 and 4.0 | NR | 40 d | Lactose in milk | 3.7 U/mg protein | [61] |
| Enzyme | Source | Objective of study | Results of study | Reference |
|---|---|---|---|---|
| Protease | Bacillus subtilis | Optimize conditions for deproteinization of crustacean wastes in the preparation of chitin | The metal chelator sensitive neutral protease activity was highest (20.2 U/mL) at pH 8.0 and 50 °C with casein as substrate. | [63] |
| Proteases | Pyrococcus furiosus | New thermostable enzymes and the use of these enzymes both in proteolysis as well as protein and polypeptide synthesis | The serine proteases retained enzymatic activity at 100 °C. It facilitated highly specific and efficient peptide synthesis even at high temperatures with high yields. | [64] |
| Alkaline protease | Aureobasidium pullulans | Optimization of medium and cultivation conditions | First report of alkaline protease production by marine yeast. Maximum production of the enzyme (623.1 U/mg protein; 7.2 U/mL) was obtained. It had the highest activity at pH 9.0 and 45 °C. | [65] |
| Protease | Bacillus subtilis | Optimization of physical factors affecting the production | The thermostable, organic solvent-tolerant protease was high (4042.4 U/mg) after optimization. | [66] |
| Protease | Vibrio sp. | Mathematical modeling leading to scale-up to industrial level | On industrial scale, potential of protease production by Vibrio anguillarum was highest with rainbow trout and squid peptones. | [67] |
| Chitinase | Vibrio sp. (FERM BP-5769) | Effect of medium constituents on enzyme production | The marine psychrophilic strain was induced by chitin and derivatives such as chitin glycol singly or in combination. | [68] |
| Chitinase | Pantoea dispersa | Effect of nineteen medium components on chitinase production optimized by Plackett–Burman design. | 4.21-fold increase in chitinase production was observed in 22nd medium of Plackett–Burman experimental design. | [69] |
| Chitinase | Streptomyces sp. DA11 | Statistical Plackett–Burman design and Box–Henken response surface methodology to optimize medium components for enhancing chitinase activity | The chitinase activity and the maximum cell dry weight were 39.2-fold and 2.6-fold higher than that of the basic medium. | [70] |
| Chitinase | Paenibacillus sabina | Statistical optimisation of medium components | Biomass and pH played an important role in increasing chitinase production. 2.74 fold increase in chitinase production was achieved. | [71] |
| Enzymatic method for chitin extraction | Co-cultivation of Lactococcus lactis and Teredinobacter turnirae | Biological treatment of prawn waste for chitin production | Highest chitinase yield 95.5% was obtained when T. turnirae was first inoculated followed by L. lactis. This provided an environmentally friendly alternative. | [72] |
| Laccase | Unidentified basidiomycetous fungi | Decolourization of paper and pulp mills, textile, dye-making industries and alcohol distillery effluents | Isolate NIOCC #2a was efficient in decolorization of various colored effluents by producing laccase, which was active at pH 3.0, 6.0 and 60 °C in the presence of seawater. | [73] |
| Laccase, lignin peroxidase and manganese peroxidase | Unidentified basidiomycetous fungi | Effect of carbon and nitrogen sources on the production of lignin- degrading enzymes | The lignin–degrading enzymes and decolorization of effluent depended on the type of the nitrogen sources used. | [74] |
| Manganese- dependent peroxidase (MNP) and laccase | Flavodon flavus | Decolorization of molasses spent wash | Colour of molasses spent wash (MSW) was reduced by 80% and total phenolics, chemical oxygen demand were reduced by 50% with the strain. There was no role for manganese-dependent peroxidase in MSW. | [75] |
| Laccase, manganese peroxidase and lignin peroxidase | Aspergillus sclerotiorum CBMAI 849, Cladosporium cladosporioides CBMAI 857 and Mucor racemosus CBMAI 847 | Enzyme production with different carbon sources and salinity conditions by using statistical experimental design. | First report of lignolytic enzymes from zygomycetes of Mucor genus. Highest values for lignin peroxidase, manganese peroxidase and laccase were obtained with the fungus M. racemosus CBMAI 847. | [76] |
| Manganese- dependent peroxidase, lignin peroxidase and Laccase | White-rot fungus Flavodon flavus (K1) | A process for removal of dyes using the lignin-modifying white-rot fungus | The salt tolerant fungi is better suited for treatment of industrial wastes as it can grow in half strength seawater. | [77] |
| Esterase | Bacillus licheniformis MP-2 | Optimization of medium composition and cultural conditions by Plackett-Burman and Box-Henken design | The marine MP-2 esterase activity was improved from 258.8 U/mL to 318.2 U/mL and approached almost about 95% of the predicted value. | [78] |
| Esterase | Staphylothermus, Pyrodictium, Archaeoglobus, Aquifex, M11TL, Thermococcus, Teredinibacter and Sulfolobus | Enzyme production from native or recombinant host cells | Novel enzymes, active peptide fragments, analogs and derivatives were reported. | [79] |
| Lipase | Pseudomonas sp. (MSI057) | Optimization of culture conditions for psychrophilic alkaline lipase production | The optimum temperature and pH for the enzyme production was 30 °C and 9.0, respectively. The activity of purified enzyme was optimum at 37 °C and showed 80% activity at 20 °C though activity was decreased above 50 °C. | [80] |
| Amylase | Halobacterium salinarum MMD047 | Optimization of medium composition and cultural conditions | Production was highest in minimal medium supplemented with 1% sucrose. | [81] |
| Endoglucanase | Teredinobacter turnirae | Effect of various carbon and nitrogen sources on Teredinobacter turnirae | Sucrose, ammonium phosphates and Triton X-100 enhanced of endoglucanase production. Combination of the components improved the production by 3.6 fold. | [82] |
| Cellulase | Marinobacter sp. (MSI032) | Optimization of medium composition and cultural conditions | 1% maltose, 1% peptone and casein supported maximal production at 27 °C and pH 9.0. | [83] |
| Cellulase | Rhodothermus marinus | Methods for producing thermostable cellulases | Polypeptides from Rhodothermus marinus were expressed in E. coli. | [84] |
| Alginate Lyase | Vibrio sp. YKW-34 | Optimization of culturing conditions and medium composition | After inducing, the activity of alginate lyase, reached 5 U/mL. Alginate, Laminaria powder acted as inducer whereas fucoidan, cellulose and glucose had negative effect on the alginate lyase production. | [85] |
| Glutaminase | Providencia sp. | Screening of about 400 marine isolates, biochemical identification tests, 16S rRNA sequencing and media optimization studies | Applying response surface methodology, glutaminase activity and specific activity 119 ± 0.12 U/L and 0.63 U/mg protein respectively were obtained from one Providencia sp. | [86] |
| Glutaminase | Recombinant Escherichia coli | Overexpression of salt tolerant L- glutaminase from Micrococcus luteus | Escherichia coli JM109 transformed with pKSGHE3-1 exhibited more than 190-fold higher glutaminase activity than M. luteus K-3 under optimal culture conditions. | [87] |
| Recombinant hexose oxidase | Chondrus crispus, Iridophycus flaccidum and Euthora cristata | Expression of recombinant hexose oxidase in Pichia pastoris | An isolated DNA fragment having hexose oxidsae activity was expressed in E. coli or S. cerevisae or P. pastoris | [88] |
| Phytase | Marine yeast Kodamaea ohmeri | Medium optimization by response surface methodology | 9-fold enhancement in phytase activity (from 62.0 to 575.5 U/mL) was attained after optimization. | [89] |
| PUFA polyketide synthase | Schizochytrium (a Thraustochytrid marine microorganism) | Production of PUFA polyketide synthase | Methods of making and using the non-bacterial PUFA PKS systems were described. | [90,91] |
| Biofilm degrading enzymes | Microbulbifer sp. | Isolation of biofilm degrading enzymes | Method for preparing biofilm degrading, multiple specificity, hydrolytic enzyme mixtures which are specifically tailored to remove targeted biofilms. | [92] |
| Superoxide dismutase | Photobacterium phosphoreum, Photobacterium leiognathi and Photobacterium sepia | Production of superoxide dismutase containing iron. | Process for the production of superoxide dismutase extracted from marine bacterial strains was described. | [93] |
| Silicatein | Suberites domuncula | Use of highly-expressed and highly active recombinant silicatein | A method for the synthesis of amorphous silicone dioxide, silicones or other silicon (IV) or metal (IV) compounds or mixed polymers of these compounds by contacting a silicon substrate with a polypeptide or a metal complex of a polypeptide comprising a carbonic anhydrase domain was described. | [94] |
| DNA ligase | Pyrococcus furiosus | Isolation of thermostable DNA ligase. | DNA ligase that retained its activity from 85 °C to 100 °C was described. | [95] |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sarkar, S.; Pramanik, A.; Mitra, A.; Mukherjee, J. Bioprocessing Data for the Production of Marine Enzymes. Mar. Drugs 2010, 8, 1323-1372. https://doi.org/10.3390/md8041323
Sarkar S, Pramanik A, Mitra A, Mukherjee J. Bioprocessing Data for the Production of Marine Enzymes. Marine Drugs. 2010; 8(4):1323-1372. https://doi.org/10.3390/md8041323
Chicago/Turabian StyleSarkar, Sreyashi, Arnab Pramanik, Anindita Mitra, and Joydeep Mukherjee. 2010. "Bioprocessing Data for the Production of Marine Enzymes" Marine Drugs 8, no. 4: 1323-1372. https://doi.org/10.3390/md8041323