Marine Toxins That Target Voltage-gated Sodium Channels

Hotchkiss Brain Institute and the Department of Physiology and Biophysics, University of Calgary, 3330 Hospital Drive N.W., Calgary, Alberta, Canada
*
Author to whom correspondence should be addressed.
Mar. Drugs 2006, 4(3), 157-192; https://doi.org/10.3390/md403157
Submission received: 4 January 2006
/
Accepted: 1 March 2006
/
Published: 6 April 2006
(This article belongs to the Special Issue Marine Drugs and Ion Channels)
Abstract
:Eukaryotic, voltage-gated sodium (NaV) channels are large membrane proteins which underlie generation and propagation of rapid electrical signals in nerve, muscle and heart. Nine different NaV receptor sites, for natural ligands and/or drugs, have been identified, based on functional analyses and site-directed mutagenesis. In the marine ecosystem, numerous toxins have evolved to disrupt NaV channel function, either by inhibition of current flow through the channels, or by modifying the activation and inactivation gating processes by which the channels open and close. These toxins function in their native environment as offensive or defensive weapons in prey capture or deterrence of predators. In composition, they range from organic molecules of varying size and complexity to peptides consisting of ~10–70 amino acids. We review the variety of known NaV-targeted marine toxins, outlining, where known, their sites of interaction with the channel protein and their functional effects. In a number of cases, these natural ligands have the potential applications as drugs in clinical settings, or as models for drug development.
1. Overview
In this chapter, we broadly sketch the range of marine toxins that interact with voltage-gated sodium (NaV) channels, outlining their functional effects and discussing their actions in the context of recognized ligand binding sites on the channel. Four other articles in this issue provide more specialized treatments of different groups of toxins: tetrodotoxin resistance and adaptive mutations in targeted species (Geffeny and Ruben), ciguatoxins (Nicholson and Lewis), and conotoxins (Layer and McIntosh). We organize our presentation in terms of functional effects of the toxins, such as pore blockers and gating modifiers, and within these functionally circumscribed groups, we consider both peptide and non-peptide agents. Generally, the non-peptides are smaller metabolites, which are concentrated in food chains that may span from bacteria to vertebrates, and become of particular concern for human health and economic reasons during the phytoplankton blooms commonly known as “red tides” [139]. In contrast, the peptides of interest generally are larger molecules, which are elaborated in several phyla of eukaryotes, in specialized structures such as nematocysts or venom glands. The identification and classification of the NaV channel receptors, through which both peptide and non-peptide toxins and modulators act, has progressed through definitions based strictly on functional interactions to, in some cases, identification of specific amino acid residues and atomic interactions involved. Where possible, we present our overview of functional groups of NaV channel targeted toxins in the context of the relevant, recognized receptors on the channel.
2. Structure of NaV channels
Eukaryotic NaV channels are integral membrane proteins consisting of a main structural α subunit of ~2000 amino acids [22–24], and one or more β subunits [70; 71]. NaV channels are members of a family of voltage-gated ion channels which also includes calcium (CaV) and potassium (KV) channels. The voltage-gated channels are members of a larger superfamily of P loop channels, which include the smaller inwardly rectifying K+ channels. The simplest members of the voltage-gated channel family are homotetrameric, and include a number of delayed rectifier KV channels and certain prokaryotic NaV channels [108; 118]. Homotetrameric channels are presumably the evolutionary predecessors of the 4-domain members of the voltage-gated channel family.
The α subunit of NaV channels (Figure 1) is made up of four homologous domains, I-IV, each of which possesses six transmembrane helices, S1–S6, with cytoplasmic N- and C-termini. The cytoplasmic, III-IV connecting loop forms the fast inactivation gate of the channel [122]. Each of the four major domains can be sub-divided into a voltage sensing domain (S1–S4) and a pore-forming P domain (S5–S6). Within each P domain, the S5–S6 connector loops into the membrane to form, in sequence from its N- to C- terminal end, a random-coiled “turret”, short supporting P helix, the lining outer pore including the narrowest part – the selectivity filter, and a P-S6 connector. The P domain forms part or all of the receptors for a number of pore-blocking toxins and drugs, including guanidinium toxins and μ-conotoxins. The voltage-sensor domain includes a conspicuous S4 segment, with Arg or Lys residues, spaced generally at every third position, and comprising most of the charges contributing to gating charge movement generated during voltage-sensitive channel gating. A complementary acidic residue in S2 also makes some contribution to gating current [128]. Channel opening following voltage activation appears to occur by a conformational change near the crossing of S6 helices towards the cytoplasmic end of the conducting pore. Inactivation occurs on several time scales: fast – gated by the III-IV linker latching to a receptor near the cytoplasmic end of the S6 segments, and a complex of slow inactivation processes which appear, at least, to involve conformational changes in the selectivity filter near the outer end of the pore [147; 149]. NaV channels typically show more than 10-fold selectivity for Na+ ions over other physiological ion species [63], based on a narrow selectivity filter [63], whose primary element is a ring of residues, DEKA [126], contributed by the DI-DIV respectively, arranged clockwise as viewed from the extracellular end of the conducting pore [37; 84].
3. Drug and toxin receptors on NaV channels
Beginning in the 1960s and 1970s with the pioneering studies of Narahashi, Moore, Catterall and colleagues, ligand sites on NaV channels have been identified and classified. Key pharmacological targets in NaV channels tend to be clustered on the functionally important structures, which are intimately associated with activation and inactivation gating, and with ion conduction and selectivity. Nine receptors are widely recognized, as listed in Table 1.
In Figure 2, we loosely map the nine receptor sites on to a cartoon derived from the recent crystal structure of the voltage-gated K+ channel, KV1.2, in Figure 2. The KV1.2 structure [90] is the only available, high resolution structure of a voltage-gated eukaryotic channel, and thus provides the most up to date clues as to the likely structure of the four-domain NaV and CaV channels. In the discourse which follows, we collect the different toxins into functional groups, each of which is cross-referenced to the receptor site with which it interacts, where this is known.
4. NaV channel blockers and other inhibitors
4.1. Receptor site 1
We will define as pore “blockers” toxins which inhibit ion transport by sterically and/or electrostatically occluding the ion conducting pore of the channel. The best known NaV pore blockers are tetrodotoxin (TTx), saxitoxin (STx) and the μ-conotoxins (see below). Receptor site 1 is located in the outer vestibule of the channel and is formed by the P loops from the four domains of the channel (see Figs. 1 and 2). Historically, the functional criterion for a toxin or drug to be considered to act at site 1 has been competition for binding with TTx or STx. The similar functional effects of chemically different blockers notwithstanding, the specific channel residues that make the most important contacts with different toxins and drugs will depend on the size and structure of the blocker. Thus, smaller heterocyclic toxins such as TTx and STx have less points of interaction with the pore than larger peptide molecules like the μ-conotoxins, which have a more complex set of contacts between ligand and receptor. The strongest known interactions of all of these toxins are in the vestibule-lining P loops within 3 or 4 residues on the C-terminal side of the residues making up the DEKA “inner ring” of the selectivity filter. Particularly important for binding of the GIII μ-conotoxins are the “outer ring” acidic residues located 3 (domains I, II, and IV) or 4 (domain III) residues C-terminal from the DEKA ring. In addition, extrapore residues have been shown to affect both affinity and selectivity among different channel isoforms [28; 85]. Identified interactions of μ-conotoxin GIIIA have been recently reviewed in detail by French and Terlau [52].
Studies on rat brain channels identified the following domain I residues as important for high affinity TTx block: D384, E387, Y385 [100; 143]. Here, we cite rNaV1.2 numbering, in which the inner DEKA ring is made up of D384, E942, K1422, and A1714. A somewhat different set of channel residues provided the most important interactions for STx. These included E942 and E945 (domain II), and K1422 and D1426 (domain III) [143]. A more recent, detailed analysis of interactions of TTx and STx with NaV1.4 has been provided by Dudley and colleagues [29; 30; 110; 111].
Different channel isoforms from brain, heart and muscle have a different sensitivity to TTx, STx, and to μ-conotoxins. In TTx sensitive (TTx-s) channels in brain and adult skeletal muscle, an aromatic residue appears essential for high affinity block by TTx. In skeletal muscle (rNaV1.4), this is Y401, corresponding to F385 in a rat brain isoform (rNaV1.2), while the homologous residue in the relatively TTx-resistant (TTx-r) heart channel is C374 (rNaV1.5), as reviewed by Denac and coworkers [35]. A similar amino acid, located in the P loop regions, is associated with TTx sensitivity in different neuronal channels. In dorsal root ganglia neurons, F385 is found in TTx-s NaV channels, while C or S is present in the homologous position in TTx-r channels, [104]. An earlier study found that mutation of E403Q in the rat skeletal muscle sodium channel, which essentially abolishes TTx and STx block, reduced sensitivity to μ-conotoxin GIIIA and GIIIB only about 4-fold [137].
Thus, different toxins may produce similar pharmacological effects by interacting with different subsets of residues within the same larger surface on the target channel molecule. The key to selective targeting of drugs may lie in a deeper understanding of the non-overlapping portions of larger receptor sites. Ecologically, NaV channel blockers may play either defensive or offensive roles [120].
4.2. Non-peptide NaV channel pore blockers and inhibitors
4.2.1. Tetrodotoxin
Tetrodotoxin is one of the best known marine toxins because of health concerns based on its involvement in fatal food poisoning, its unique chemical structure, and its specific action of blocking NaV channels of excitable membranes [27; 64; 164; 167]. The toxin, whose name was derived from the order of fish with which it is most commonly associated, the Tetraodontiformes. There are more than 40 species of tetraodon pufferfish, known in Japan as fugu [27]. Other marine organisms, including polyclad flatworms [120], and some terrestrial poisonous animals, have also been found to store TTx. However, bony fish from the family Tetraodontidae are especially well known to cause tetrodotoxin poisoning. These fish do not possess a venom apparatus, but TTx may be found in all organs of the body. The occurrence and distribution of TTx among different phylogenic lines of organisms gave evidence for speculation that TTx and its derivatives originated from symbiotic microorganisms. Indeed, a number of bacteria, including the genera Aeromonas and Alteromonas, Escherichia coli, Photobacterium phosphoreum, Plesiomonas shigelloides, Pseudomonas sp. and some Vibrio sp. have been shown to produce TTx (reviewed in [51; 79; 164]).
TTx is of amphoteric nature with an unusual tricyclic structure (Figure 3-A). It possesses both cationic (guanidine) and anionic (acid hydroxyl) groups. TTx binds with high affinity [dissociation constant (Kd) in the nM concentration range] to the external surface of NaV channels by binding to site 1 of these channels [27]. The positively charged guanidinium group interacts with a negatively charged carboxyl group at the mouth of the NaV channel on the extracellular side of plasma membranes of excitable membranes. When it does so, the inward flow of Na+ through the channel pore is blocked, leading to inhibition of action potential generation and propagation [64; 79]. Seven derivatives of TTx have been detected in puffer fish, newts and poisonous frogs. These TTx derivatives vary in their potencies [164]. The value of this particular toxin in electrophysiological research is profound. Recent models of the molecular structure of the NaV pore have been strongly based on interactions with this toxin, and NaV channels are often divided broadly into two pharmacological classes, TTx-s and TTx-r.
4.2.2. Saxitoxin
Paralytic shellfish poisoning (PSP) is a deadly affliction that results from the accidental consumption of some potent natural neurotoxins, typically via contaminated seafood [114]. Among all seafood poisoning, PSP poses the most serious threat to public health, and the economic damage caused by accumulation of toxins in shellfish is immeasurable (see [79] and references therein). Saxitoxin (STx) and its analogs are collectively called paralytic shellfish toxins (PST) and are considered the causative agents for PSP. The name STx was derived from the mollusc in which it was first identified, Saxidomus giganteus[79]. Toxins are present in dinoflagellates such as Alexandrium sp., Pyrodinium bahamense var. compressum and Gymnodinium catenatum [88; 135] during red tides, which causes STx to be concentrated by filter-feeding shellfish and subsequently, to be conveyed to humans when they consume the shellfish. Neurological symptoms occur shortly after the time of ingestion, and, in extreme cases, can lead into death [166]. Although it has been generally considered that PST are associated with dinoflagellates, there is evidence that heterotrophic bacteria are responsible for toxin synthesis in these organisms [31; 55; 92; 148].
The saxitoxins are a family of water-soluble neurotoxins and are among the most potent toxins known. Information on the toxicity of saxitoxins is mainly restricted to acute toxicity in mammals and humans (reviewed in [79]). For example, the LD50 values (lethal dose/concentration for 50% of population) of STx by intravenous, intraperitoneal and oral routs in mice are 2.4 μg/kg, 10 μg/kg and 263 μg/kg, respectively. Saxitoxins are tricyclic compounds (Figure 3-B) and their molecular skeletons are structurally related to TTx, but with two guanidinium moieties [131] in STx compared to the one guanidinium moiety in TTx. More than 20 analogs have been described [79]. Like the case of TTx, saxitoxins are well documented non-peptide neurotoxins that are highly selective blockers of NaV channels in excitable cells at site 1, thereby affecting nerve and muscle impulse generation in animals [27; 64; 131; 164; 167]. Recently a new, exceptionally potent derivative of saxitoxin has been described [165].
4.2.3. Meroditerpenoids
Brown algae (Phaeophyceae) produce a variety of secondary metabolites possessing many different structural varieties and biological activities ([50] and reviews therein). The genus Stypopodium is a tropical group of the Phaeophyceae well-known for its rich complement of polycyclic diterpenoids fused to oxidized aromatic rings (meroditerpenoids) [50]. These terpenoid metabolites are useful chemotaxonomic markers because of their structural complexity and genetic origins [56]. Meroditerpenoids display potent biological activities, with toxic and enzyme-modulating functions, which may be of biomedical and pharmacological utility [155]. Three new meroditerpenoids, 2β,3α-epitaondiol (Figure 3-C-1), flabellinol (Figure 3-C-2) and flabellinone (Figure 3-C-3), from Stypopodium flabelliforme were assayed to have neurotoxic and NaV channel modulation activity [123]. In this study, these compounds were cytotoxic to NCI-H460 cells with an LD50 of 24, 9, and 14 μM, respectively. In a NaV channel modulation assay using the neuro-2a neuroblastoma cell line, with STx as a control inhibitor, these compounds showed detectable NaV channel blocking activity at 0.7, 2, and 7 μM, respectively. Overall, these meroditerpenoid metabolites show a broad range of biological activities, including being toxic to fish and brine shrimp, and to some human cancer cells. Some of this toxicity may be due to a blocking of NaV channels [50; 123].
4.3. Peptide NaV channel pore blockers and other inhibitors
4.3.1. μ-Conotoxins
Conotoxins in general, and μ-conotoxins in particular, are found in the venom of predatory tropical marine cone snails, genus Conus. Venom is a primary weapon used by these molluscs to capture prey (small fish or invertebrates) and to defend themselves. Venoms of all cone snails are complex and an individual species may express 100–200 venom peptides. Toxic components of the venom are mostly small peptides [106], which are synthesized in a tubular venom duct. All cone snails have specialized radular teeth that penetrate the skin of prey and inject venom. These teeth work as a harpoon and sharp needles. After the venom injection, the prey becomes quickly paralyzed and is now available for consumption. Some fish-hunting species (e.g., Conus geographus) are dangerous to humans, and fatalities have been recorded in the medical literature [144].
μ-Conotoxins are highly basic, cysteine-rich peptides which have three disulphide bridges in the three loop framework. The amino acid sequences of seven μ-conotoxins showed in Figure 4, and some basic properties of these μ-conotoxins are displayed in Table 2.
The number of recognized μ-conotoxins is rapidly increasing, thanks to a combination of molecular cloning techniques to identify new sequences, and application of improved synthetic procedures to produce oxidized, active conformations of the folded toxins (see Table 3). NMR-derived 3D structures of GIIIA, PIIIA and SmIIIA are shown in Figure 5. Discovery of new toxins is revealing increased functional diversity in the μ-conotoxin family. In addition, recent work shows that there is a divergence of the disulfide framework generating structural diversity among the μ-conotoxins [32]. At present time, no μ-conotoxins are used directly in a clinical application. However, discovery of new isoform-specific μ-conotoxins from Conus venoms, or constructs not existing in nature, offers a promising way to neurologically active drugs [1]. Discoveries of μ-conotoxins SmIIIA, SIIIA and KIIIA open new opportunities for creation of specific drugs for pain relief. These toxins block the TTx-r NaV channels that are responsible for pain signal propagation in dorsal root ganglia neurons.
4.3.2. μO-Conotoxins
The μO- and δ-conotoxins are unusually hydrophobic peptides, which belong to the O-superfamily of conotoxins. The δ-conotoxins inhibit inactivation of NaV channels, while μO-conotoxins affect both Na+ and Ca2+ currents [48]. μO-conotoxins were first isolated from the cone snail, Conus marmoreus [48; 95]. C. marmoreus is a molluscivorous (snail hunting) cone snail which can be found in the waters of the Philippines [95]. To date, only two μO-conotoxins have been discovered, the MrVIA and MrVIB, both from C. marmoreus. Both of these toxins are 31 residues long, with three intramolecular disulphide bonds. They show high sequence homology with only two residues not conserved between the two toxins. The 3D solution structures of these toxins were first determined using simulated annealing procedures and based on NMR data [34]. The μO-conotoxins inhibit NaV channels as well as molluscan calcium channels [48].
The chemical structure of μO-conotoxins has similarities with δ-conotoxins, and the ω-conotoxins (which target calcium channels and not discussed further in this chapter) [34]. The block of NaV channels by the μO-conotoxins seems to be independent of site 1, at which TTx, STx, and the μ-conotoxins bind, and shows no effect on either activation or inactivation of the channel [146]. Competition studies showed that the μO-conotoxins have no effect on the binding of [3H]STx. Furthermore, targeting of μO-conotoxins differs significantly from the μ-conotoxins. As well as inhibiting TTx-s NaV channels from brain (NaV1.2) with IC50 (concentration producing 50% inhibition) values of 200 nM [146], they also block TTx-r NaV channels from the rat dorsal root ganglion (probably NaV1.8) with IC50 values of 82.8 nM. The equivalent TTx-r channels in humans are responsible for transmission of pain sensation, which makes μO-conotoxins a very promising therapeutic agents [34]. However, the molecular mechanisms of interaction of the μO-conotoxins with NaV and CaV channels are not yet well understood.
4.3.3. Unclassified conotoxins, GS and TVIIA
The unusual properties of some conotoxins do not allow them to be grouped within the known families [144]. For example, TVIIA from the venom of Conus tulipa, a 30-residue polypeptide, displays high sequence identity with conotoxin GS, from venom of Conus geographus. Although there are reports indicating that GS binds to the TTx receptor in rat skeletal muscle, no more detailed functional studies have appeared. Their overall structure suggests that GS and TVIIA represent a distinct subgroup of the family which includes the CaV-targeted ω-conotoxins [62].
4.4. Lipopeptide Nav channel inhibitors
4.4.1. Jamaicamides
Screening extracts from marine Dinoflagellate Lyngbya majuscule revealed a number of potently bioactive metabolites ([20] and reviews therein). Jamaicamides A-C (Figure 6-A) are recently characterized polyketide-peptide neurotoxins from L. majuscule[38]. The latter authors examined the neurotoxic activity of crude extracts as well as purified jamaicamide compounds. Neither jamaicamide A nor B showed significant brine shrimp toxicity, while jamaicamide C was only modestly active at 10 ppm (25% lethality).
In the goldfish toxicity assay, jamaicamide B was the most active (100% lethality at 5 ppm after 90 min), followed by jamaicamide C (100% lethality at 10 ppm after 90 min). Interestingly, jamaicamide A was the least active fish toxin (sublethal toxicity at 10 ppm after 90 min). Also, Jamaicamide compounds exhibited cytotoxicity to both the H-460 human lung and neuro-2a mouse neuroblastoma cell lines. Using a bioassay method, modified from Manger et al. [91], it has been suggested NaV channel modulation by jamaicamides A-C [38]. The LD50s from these bioassays were approximately 15 μM for all three compounds in both cell lines. All three compounds also inhibited NaV channel activity at 5 μM, producing approximately half the response of STx applied at 0.15 μM. None of the jamaicamides were shown to activate the tested NaV channels. Overall, jamaicamides show a spectrum of biological activities consistent with their functioning as defense metabolites, including Nav channel blocking activity, and arthropod and fish toxicity. Further work should be done to study the molecular interaction of jamaicamide compounds with NaV channels.
4.4.2. Kalkitoxin
Kalkitoxin (KTx) is a neurotoxic, thiazoline-containing lipopeptide (see Figure 6-B) from a Caribbean dinoflagellate, Lyngbya majuscule[160]. This toxin acts exclusively as a potent blocker of activated TTx-s NaV channels in mouse neuron-2a cells (50% block occurred at IC50 = 1 nM, compared to the STx IC50 = 8 nM; [160]). In another study, KTx produced N-methyl-D-aspartate (NMDA) receptor-dependent cytotoxicity in rat cerebellar granule neurons, with an LD50 of 3.86 ± 1.91 nM [9]. It is somewhat surprising that the agonist, antillatoxin (ATx, see below), also induced cytotoxicity that was prevented by non-competitive NMDA receptor antagonists. Nevertheless, the cytotoxicity responses to ATx and KTx displayed distinct temporal patterns [9], in that ATx induced an acute response, whereas KTx produced delayed cytotoxicity. Thus, cytotoxicity may be induced by different pathways in these two cases. Additionally, KTx is highly active in an inflammatory disease model which measures IL-1β-induced secreted phopholipase A2 secretion from HepG2 cells (IC50 = 27 nM; [160]). Recently, KTx was shown to interact with TTx-s NaV channels at site 7 in cerebellar granule neuron cultures [81]. Consistent with previous work, KTx may be a valuable tool in efforts to more clearly define the structure-function relationships of these NaV channels, and may provide possible leads for analgesic and neuroprotection drug discovery.
5. Gating modifiers
5.1. Non-peptide NaV channel modulators
5.1.1. Alkaloid inhibitors of Na+ channels
Marine organisms are known to produce compounds with novel chemical structures and interesting biological activities. Apart from algae, cone snails, sea anemones and many other organisms, sponges are the source of the greatest diversity of marine natural products (see references [15; 50] and previous reviews cited therein). Alkaloid compounds found in tropical Agelas sponges have been shown to be active on muscle and nerve membrane receptors and channels, including NaV channels, and are excellent pharmacological tools to study the physiology and biophysics of these macromolecules [119]. Morales and Rodriguez [96] reported that Caribbean sponges of the genus Agelas contain alkaloids including monomers of clathrodin (CLA, Figure 7-A) and dibromosceptrin (DBS, Figure 7- B). Both compounds have guanidine moieties linked to pyrrol groups with bromide substitutions ranging from none (CLA) to four (DBS). Use of the whole-cell configuration of the patch-clamp technique revealed that these compounds have neurotoxic activity [119]. Both compounds decreased the average maximum amplitude of inward Na+ current by 27–40% on cells isolated from sympathetic ganglia of chick embryo to test the CLA and DBS effect on provoked Na+ currents. In this study, neither compound altered the voltage dependence of current activation.
However, the voltage dependence of current inactivation was shifted toward more negative potentials by DBS. In contrast, CLA shifted this voltage dependence of inactivation toward more positive potentials. Time for current reactivation was not altered by CLA but was slightly prolonged by DBS. Similarly, DBS was more effective than CLA in prolonging the time course of current decay. Voltage clamp experiments on frog muscle showed that both CLA and DBS alter the voltage-dependent ionic currents, with CLA causing an early potentiation followed by total inhibition, and DBS exerting an immediate current blockade [119].
CLA and DBS effects on ionic channels were examined using CLA and DBS in radioligand binding studies, which showed that DBS inhibited nearly 90% of [3H]-STx binding to rat brain synaptosomes while CLA inhibited only 30% [119]. Thus, CLA and DBS are NaV channel neurotoxins which bind to a site that inhibits STx binding. Nevertheless, further investigation is needed to specify the molecular target of each toxin. Functionally, CLA and DBS modulate NaV channels by different mechanisms; DBS, a dimer with four bromide substitutions, acted on the voltage-dependence of inactivation, while CLA, the monomer without bromide substitutions, was directed towards the channel ionic conductance. Recently, DBS has been shown to disrupt the cellular calcium homeostasis, by targeting CaV channels (IC50 ~μM) in PC12 cells [11]. This potent effect of DBS on Ca2+ influx and Ca2+ levels in neurons and sensory cells is consistent with that the previously reported feeding deterrent activity of brominated pyrrole alkaloids in Agelas sponges against predatory reef fish (reviewed by [87]). Furthermore, chemoreception, sensing bromopyrrole alkaloids in sea water, was exhibited by sensory neurons in the rhinophore of the sea slug Aplysia punctata[11].
5.1.2. Brevetoxins
The brevetoxins, are part of a large family of dinoflagellate-derived polyketide toxins which pose a threat to human health through the consumption of tainted seafood. This family also includes ciguatoxin, okadaic acid and the related dinophysis toxins, pectenotoxins, yessotoxin and the azaspir acids [136]. Ingestion of bivalve molluscs contaminated with brevetoxins leads to neurotoxic shellfish poisoning (NSP), the symptoms of which include nausea, cramps, paresthesias of lips, face and extremities, weakness and difficulty in movement, paralysis, seizures and coma [4; 39; 94]. Inhalation of aerosolized brevetoxins from sea spray results mainly in respiratory irritation as well as dizziness, tunnel vision and skin rashes [4; 112].
Brevetoxins (PbTx-1 to PbTx-10) are potent lipid-soluble polyether neurotoxins produced by the marine dinoflagellate, Karenia brevis (formerly known as Gymnodium breve or Ptychodiscus breve; [2]). These organisms are associated with ‘red tide’ blooms in the Gulf of Mexico and in New Zealand [69]. Brevetoxins exert a variety of actions in vivo affecting both the PNS as well as the CNS, probably due to their high lipid solubility and ability to penetrate the blood brain barrier [78]. The brevetoxins are a suite of polyether ladder type compounds which have two parent backbone structures, brevetoxin A and brevetoxin B, each with several side chain variants [3] (Figure 8-A). Brevetoxin 1, PbTx-1 from group A, is the most active ichthyotoxic brevetoxin analog with IC50 values ranging between 3 and 4 nM [2].
Brevetoxins are known to interact specifically with neurotoxin receptor site 5 on the α subunit of NaV channels [2]. This interaction with the α subunit causes a shift in the voltage dependence of channel activation to more negative potentials and inhibits channel inactivation, thereby producing neuronal depolarization at rest [26; 113; 116; 117; 129]. In addition, brevetoxins augment Na+ influx through NaV channels because of the shift in activation, and by prolonging channel opening, resulting in a dose-dependent depolarization of excitable membranes [72]. Bottein Dechraoui and Ramsdell [16] showed that type A (PbTx-1) and type B (PbTx-3 and PbTx-2) brevetoxins, as well as ciguatoxins, target both cardiac and muscle channels. Type B brevetoxins show isoform selectivity, presenting a lower affinity for the cardiac (NaV1.5) than the skeletal muscle (NaV1.4) Nav channel isoforms expressed in Human Embryonic Kidney (HEK) cells. The lower affinity of type B brevetoxins for heart Nav channels may result from a more rigid backbone structure than that found in type A brevetoxins and in ciguatoxins. On the other hand, PbTx-2 augmented NMDA receptor function and, therefore, may exacerbate NMDA receptor-mediated pathologic sequelae associated with neurodegenerative diseases [36]. Furthermore, a series of naturally occurring fused-ring polyether compounds with fewer rings than brevetoxin, known as brevenals, were shown to exhibit antagonistic properties and counteract the effects of the brevetoxins in neuronal and pulmonary model systems [3]. This finding supports the idea that the toxicity of the red tide blooms depends on the amount of each toxin present as well as the spectrum of molecular activities elicited by each toxin.
5.1.3. Ciguatoxins
The term ciguatera originated in the Caribbean areas to designate intoxication induced by the ingestion of the marine snail Turbo pica (called cigua), first described by Cuban ichthyologists (reviewed by [109]). Today, the term is widely used to denote a fish-poisoning that result from ingestion of ciguatoxic fish found mainly in tropical and subtropical regions [83]. Human victims are the end link in a food chain cascade where ciguatoxins are accumulated in a variety of consumed coral reef fishes [82]. Ciguatoxins (CigTx) are lipid-soluble, heat-stable, highly oxygenated cyclic polyether marine toxins originally produced in coral reef areas by the benthic dinoflagellate Gambierdiscus toxicus. Ciguatoxins are responsible for a complex human food poisoning, characterized by gastrointestinal, neuronal and cardiovascular disturbances with no immunity and symptoms may persist for months or years [79]. The symptoms of ciguatera fish poisoning overlap those of the brevetoxins with temperature reversal as supposedly unique for ciguatoxins [112]. Moreover, epidemiology of the CigTx poisoning is complex and impossible to predict outbreaks with global health concern. Worldwide coral bleaching, together with man-made factors (such as tourism, industrial constructions, sewage and eutrophication) and environmental factors (such as earthquakes and hurricanes) create a more favorable environment for G. toxicus blooms and therefore trigger ciguatera outbreaks [79].
More than 20 CigTx analogs have been found, and grouped according to their origins and carbon backbone, as Pacific (P-CigTx; Figure 8-B) or Caribbean (C-CigTx; Figure 8-B) ciguatoxins [93; 109]. These polyether toxins are highly toxic to fish (LD50 1 μg/kg; [79]) and to mammals (LD50 for mouse near 1 μg/kg; [83]). Interestingly, the more oxidized form, P-CigTx, is up to 10-fold more toxic than the less oxidized form, C-CigTx [83; 164]. This increase in potency amplifies the effective level of more oxidized forms of CigTx accumulated by fish. Extensive experimental studies of Pacific ciguatoxins, using whole-cell patch clamp of rat dissociated dorsal root ganglion neurons, have shown that P-CigTx-1 causes TTx-s NaV channels to open at more negative membrane potentials than normal. By contrast, TTx-r NaV channels recover from inactivation more quickly, enabling an earlier transition to the open state [138]. These properties, combined with the high lipid solubility of the toxin explain, at least in part, the pathogenic nature of the neurological symptoms, including the pathogenesis of sensory neurological disturbances associated with ciguatoxic fish poisoning.
Pharmacological studies have revealed that ciguatoxins activate NaV channels in the nM to pM range [7; 12; 16; 83; 89]. Specifically, ciguatoxins cause a hyperpolarizing shift in the voltagedependence of the channel activation, increasing the open probability of NaV channels at normal resting membrane potentials [7; 12; 89]. These toxic effects lead to a TTx-s, Na+-dependent membrane depolarization, and repetitive or spontaneous action potentials, in various axons and skeletal muscle fibers [16; 83; 93]. Ciguatoxins in common with brevetoxins selectively target a common receptor site, receptor site 5, on NaV channel proteins [12; 89; 93]. Because of their potency, ciguatoxins can be useful pharmacological tools to study NaV channels, as are TTx, STx and PbTx. Moreover, an increase of Na+ influx through activation of these channels at the resting membrane potential, is not only able to enhance nerve excitability but also may cause water movements across the cell membrane [93]. These toxins may be useful tools to study the Na+-dependent mechanisms of volume changes and their homeostasis in nerve preparations.
5.1.4. Ostreotoxin-3
More than 20 species of marine dinoflagellates produce toxins that are among the most potent, nonpertinacious toxin known [20; 49; 159; 164]. These dinoflagellate toxins have been implicated in human intoxications resulting from consumption of poisonous fish and shellfish [159; 164]. The benthic dinoflagellate, Ostreopsis lenticularis, is the producer of this NaV channel activating toxin, Ostreotoxin-3 [OTx-3; [44]]. Toxicity of OTx-3 was first revealed when O. lenticularis cells treated with acetone before methanol extraction showed a 10-fold increase in mouse toxicity [44]. Experiments in chick embryo sympathetic neurons demonstrated that OTx-3 extracts were acting on TTx-s NaV currents [44]. The extract affected the voltage-dependence by shifting the activation and inactivation currents to more negative potentials but its major effect was on inactivation. In the presence of μ-conotoxin GIIIA, the remaining Na+ current was not altered by OTx-3 extracts. Meunier et al. [44] performed intracellular recording experiments which demonstrated that OTx-3-enriched extracts depolarized frog skeletal muscle and decreased the amplitude of endplate potentials without affecting the nerve impulse. This effect was partially blocked by 10 μM TTx and totally blocked by μ-conotoxin GIIIA. Thus, the toxic effect of O. lenticularis extracts may be due to the presence OTx-3 that recognized TTx-s NaV currents. OTx-3 is a unique dinoflagellate toxin that can discriminate between nerve and skeletal muscle.
5.2. Peptide NaV channel gating modifiers
5.2.1. Antillatoxin
Blooms of the dinoflagellate, Lyngbya majuscule, have been associated with various adverse effects on human health, including respiratory irritation, eye inflammation, and severe contact dermatitis in exposed individuals [21; 97]. These effects are associated with the production of toxic substances by L. majuscule such as lyngbyatoxins [21; 131; 164] and debromoaplysiatoxin [97; 131]. Other toxic secondary metabolites target NaV channels, (see Table 1); these include antillatoxin [107], kalkitoxin [9; 160] and jamaicamides A-C [38]. The diversity of these natural products gives them potential for importance as biochemical and pharmacological tools.
Antillatoxin (ATx) is a structurally unusual cyclic lipopeptide (Figure 9), which has been reported to be among the most ichthyotoxic metabolite isolated to date from a marine dinoflagellate. It is exceeded in potency only by the brevetoxins [107]. ATx ichthyotoxic activity with LD50 value of 0.05 μg/ml using fish toxicity bioassay [107]. Using cerebellar granule cells, ATx produced concentrationdependent cytotoxicity with LD50 value of 20.1 ± 6.4 nM [9]. In the latter study, the neurotoxic response of ATx was prevented by co-application with noncompetitive antagonists of the NMDA receptor such as MK-801 (or dizocilpine) and dextrorphan. This neurotoxic effect of ATx was antagonized by TTx, consistent with an interaction of ATx with NaV channels [86]. ATx-induced stimulation of Ca2+ influx in cerebellar granule cells was similarly prevented by TTx. Additional, more direct, evidence that the NaV channel serves as the molecular target for ATx was provided by the demonstration that ATx stimulation of [3H]batrachotoxin binding increases 22Na+ influx in cerebellar granule cells [86]. Together, these results suggest that ATx is a novel activator of NaV channels, most probably acting at site 4. The precise recognition site for ATx on the NaV channel, however, remains to be defined. It has been suggested that development of antagonists targeted at the ATx lipopeptide site may provide another means of inhibiting NaV channels, and thereby a new strategy for the discovery of anticonvulsant and neuroprotectant drugs [86].
Recently, a new N-methyl homophenylalanine analog of ATx was identified and denoted as antillatoxin B (ATx-B; [101], Figure 9). Both ATx and ATx-B are potent activators of the NaV channel in mouse neuro-2a neuroblastoma cells, with half-maximal activation at EC50 values of 0.18 and 1.77 μM, respectively. In addition, ATx B is strongly ichthyotoxic to goldfish, with an LD50 of 1.0 μM. It seems likely that substitution of a larger N-methyl homophenylalanine residue in ATx-B for an N-methyl valine residue in ATx accounts for the 10-fold decrease in ATx-B’s potency in compare to ATx [101].
5.3. Peptide gating modifiers
5.3.1. Sea anemone toxins
Sea anemones are a member of the phylum Cnidaria, which have stinging organelles known as nematocysts, housing tiny harpoons, on their tentacles. Sea anemones have adapted their venom apparatus for use in both protection and prey capture. They have been known to capture both small fish and crustaceans. Toxins produced by sea anemones encompass many targets including both KV and NaV channels, while some species have cytolysins. Sea anemones have a wide variety of NaV channel toxins that affect inactivation from the extracellular side of the channel. Sea anemone toxins were first isolated as cardiac stimulators and neurotoxins and these are the two actions that have been studied the most. Sea anemone toxins affecting NaV channels range in size from 3–5 kDa and have been isolated from many different species [102]. Sea anemone toxins bind to NaV channels of excitable tissue and preferentially act on the open state to inhibit inactivation of the channel [35]. Sea anemone toxins bind to site 3 on the NaV channel [27] in domain IV on the S3–S4 linker. The binding to this linker prevents the necessary movement of the voltage sensing S4 segment during fast inactivation. Sea anemone toxins bind to a receptor which partially overlaps that of scorpion toxins, as seen in binding studies of the interaction between the sea anemone toxin, AneTx II, and the scorpion toxin, LqTx [121].
Sea anemone toxins, because of their large variety, can be split into three distinct groups called Type-1, Type-2 and Type-3. These groups are distinguished based on both their size and the genus from which they were isolated. Type-1 and Type-2 sea anemone toxins usually range from about 46–49 amino acids in length. Type-1 toxins were isolated from the family Actiniidae, mostly from the genera Anemonia and Anthopleura. Both of these genera produce several neurotoxins that affect NaV channels. For example, AFT I, AFT II [141], Anthopleurin-A [142], Anthopleurin-B [115], Anthopleurin-Q [53], and APE 1 to APE 5 [17], are produced by Anthopleura spp., while AneTx I, AneTx II, AneTx V [125; 161; 162] are produced by members of the genus Anemonia. Type-2 toxins have been isolated from two genera Heteracti and Stichodacyla. Examples of type-2 toxins are RTX I to RTX V [168–171], Rp II [152], Rp III [44] and ShI [75]. Type-1 toxins, like Type-2 toxins, have 3 disulphide bonds [102]. Within each type, these two toxin subclasses show high sequence identity (>60%), but they show only about 30% identity between Type-1 and Type-2 toxins. Alignment of Type-1 and Type-2 toxins reveals a conserved Arg-14. It is believed that the loop of residues surrounding the Arg-14 is important in the binding of these toxins to the channel [127]. The C-terminus is more basic in the Type-2 toxins based on the addition of several lysines and a compensating truncation of the N-terminus [102]. In all, of the Type-1 and Type-2 toxins for which structures have been determined to date, all contain a four-stranded anti-parallel β-sheet in their secondary structure, with no apparent helical structure, as shown in Figure 10. Competition studies of these two types of toxins suggest that they bind to distinct yet close receptor sites, given that some toxin pairs show competition between the two groups, while others do not [102].
Type-3 toxins comprise a small group of short polypeptides produced by sea anemones. Some examples of Type-3 toxins are PaTx from Parasicyonis actinostoloides[99], AneTx III from Anemonia sulcata[8], as well as three others from Dofleinia armata (Da I and Da II) and Entacmaea ramsayi (Er I) [66]. Type-3 toxins are usually composed of 27–31 residues, all so far with high sequence homology, with AneTx III and PaTx showing the lowest amino acid identity of approximately 50% [66]. Similar to Type-1 and Type-2 toxins, Type-3 toxins seem to bind to a receptor which overlaps position on site 3, and cause a slowing of the inactivation of NaV channels.
5.3.2. Goniopora Toxin
Goniopora toxin (GPT) was first isolated from the marine coral, Goniopora spp. GPT is a polypeptide toxin of approximately 12,000 Da [60]. Electrophysiological experiments have shown GPT to prolong the action potential in heart, without significantly affecting its amplitude, or the resting membrane potential [54]. GPT inhibited inactivation of the cardiac NaV channels, thus causing an increased permeability to Na+. The resultant broadening of the action potential leads to cardiac arrhythmias. These physiological effects are similar to those of several other NaV channel toxins, including sea anemone toxins and α-scorpion toxins, even though there is little or no sequence homology among them [60].
In N18 neuroblastoma cells, GPT shifts the conductance-voltage relationship to the left, and thus causes activation of sodium currents in response to an unusually small depolarizing stimulus [58]. This result is similar to that produced by the Leiurus scorpion toxin. GPT also shifts the steady state inactivation curve of the NaV channels to the right without altering its shape, in contrast to sea anemone and scorpion toxins, which cause a reduction in the steepness of the inactivation curve while not shifting the midpoint appreciably [57]. Depolarization has a marked affect on the affinity of GPT. The voltage dependence of GPT action reflects an e-fold increase in Kd per 48.3 mV of depolarization [57]. Again, this result differs from that observed for sea anemone toxins, which shows much steeper voltage dependence. Thus, GPT may bind preferentially with different voltage-sensitive states, or alter different voltage-sensitive transitions of the channel.
GPT shows no competitive binding with the scorpion toxins which bind to site 3. Thus, it has been determined that GPT must bind to a distinct site (site 8) to that of the scorpion toxins, even though they both preferentially slow channel inactivation.
5.3.3. B-toxins
The B-toxins were isolated from the mucus secreted by the marine heteronemertine Cerebratulus lacteus[74]. C. lacteus is a ribbon worm found off the Atlantic coast of North America. B-toxins bind to NaV channels in crustaceans, but had no effect when they were injected into an insect (cockroach) or a mammal (mouse) [74]. They appear to be selective for crustacean neuronal action potentials. The B-toxins, with molecular masses of ~6,000 Da, are a group of neurotoxins comprised of 4 subtypes, B-I to B-IV of which the B-I toxin (49 residues) contains three intrachain disulfide bonds, while B-II, B-III and B-IV (53 or 54 residues) contain four intrachain disulphide bonds [74]. These B-toxins act by delaying the inactivation of neuronal NaV channels, as do the α-scorpion and sea anemone toxins that act at site 3 [27]. There is little sequence similarity between B-toxins and the α-scorpion and sea anemone toxins [13]. The B-toxin is composed of two anti-parallel α-helices cross-linked by disulphide bonds forming a hairpin structure, while α-scorpion and sea anemone toxins are mainly β secondary structures [6]. Of the four B-toxins, B-II shows the highest toxicity and is some 15-fold higher than that of B-IV, despite their high overall sequence similarity [74].
To date, the best studied of the B-toxins is B-IV, which is the most abundant form. The B-IV toxin was first sequenced by Blumenthal and Kem in 1976 [14] and it is reported to be the first marine invertebrate polypeptide neurotoxin sequenced. The NMR solution structure of the B-IV toxin has been solved by Norton and colleagues [6] and is shown in Figure 10. There are many charged residues, which could play a role in interacting with the NaV channel α subunit. Site-directed mutagenesis of the B-IV toxin has shown that neutral replacement of its arginine residues can almost abolish its biological activity [153]. Specifically, replacement of Arg-17 and Arg-25 [67], in the N-terminal helix and the hairpin region respectively, led to at least a 700-fold loss in activity. Furthermore, replacement of Trp-30 with a non-aromatic residue caused a greater than 40-fold decrease in toxicity, whereas aromatic substitutions caused less than 5-fold decrease in toxicity [154].
5.3.4. δ-Conotoxins
Cone snail δ-conotoxins [144], are a group of peptides consisting of about 30 amino acid residues, which exert a similar effect on NaV channels. This effect is the inhibition of the fast inactivation of Na+ currents, which causes a marked prolongation of action potentials [47; 61]. This results in a hyperexcited state of the affected cells, which can eventually lead to a massive electrical hyper-excitation in the complete organism. δ-Conotoxins belong to the O-superfamily and have the same cysteine framework as the μO- and ω-conotoxins, that target NaV and CaV channels, respectively [144]. This disulfide bonding pattern gives these peptides an inhibitory cysteine knot motif which is found in a variety of biological active peptides from different organisms [103].
Olivera and collaborators have coined the term “cabal” for a group of toxins, present in a venom, which acts together toward a specific physiological end. The set of venom peptides that almost instantaneously immobilize a fish by eliciting the seizure-like symptoms is referred to as the “lightning-strike cabal” [105]. δ-Conotoxins, which in effect cause a dramatic increase in electrical activity of axons, may well be a key venom component in the lightning-strike cabal [132; 145].
It has not yet been established how widely distributed δ-conotoxins are among cone snails, especially fish-hunting cones. Every Indo-Pacific fish-hunting cone snail species examined, that is known to use a “harpoon-and-line” strategy for prey capture, expressed δ-conotoxin in its venom duct [18]. Several δ-conotoxins from fish- and snail-hunting Conus species have been identified and the biological importance of these peptides for the capture of prey seems evident. Similar to other conotoxin families which interact with NaV channels, δ-conotoxins target specific isoforms of NaV channels with major differences among vertebrate and invertebrate systems (Table 5). δ-PVIA is a 29 amino acid peptide, from the fish-hunting cone snail C. purpurascens, which slows fast inactivation of NaV1.2-mediated currents expressed in the Xenopus expression system as well as of Na+ currents recorded from hippocampal neurons in culture [145]. More recently, it has been shown that the peptide also affects NaV1.4- and NaV1.6-mediated currents [124]. By using rat brain synaptosomes, it was demonstrated that δ-PVIA competes with δ-TxVIA for the same receptor site, even though δ-TxVIA is non-toxic in vertebrate systems. When δ-PVIA is administered to fish, it causes contraction of specific muscles resulting in a characteristic extension of the mouth, which was termed “lockjaw syndrome” [132]. Although δ-PVIA elicits excitatory symptoms in both mice and fish, this peptide is inactive in molluscs, even at doses 100-fold higher. During the envenomation of a prey fish, the excitatory effects of δ-PVIA act synergistically with the KV channel-blocking peptide κ-PVIIA, resulting in the almost immediate, titanic paralysis of the prey [145].
δ-TxVIA from C. textile was originally called the “King Kong” peptide because it causes a characteristic dominant behaviour in lobster upon injection [65]. This peptide prolongs inactivation of Na+ currents only in molluscan neuronal membranes, whereas in vertebrates, δ-TxVIA binds to NaV channels but without any toxic effects in vivo and in vitro [45; 46]. Thus, δ-TxVIA can be used as molecular probe to discriminate between molluscan and mammalian NaV subtypes on the basis of structural differences and activity but not binding [130]. δ-NgVIA, isolated from piscivorous snail C. nigropunctatus, competes with δ-TxVIA for binding to rat brain synaptosomes and molluscan central nervous system, and strongly inhibits Na+ current inactivation in snail neurons, as does TxVIA [47].
δ-GmVIA, isolated from C. gloriamaris, causes action potential broadening in Aplysia neurons [133]. This peptide also affects NaV1.2 and NaV1.4 at micromolar concentrations but does not affect NaV1.6-mediated Na+ currents. A 26-residue peptide (Am 2766) has been isolated and purified from the venom of the molluscivorous snail, Conus amadis[140]. Conotoxin Am 2766, reported by the same group, belongs to the δ-conotoxin family and inhibits the decay of the Na+ current in brain rNaV1.2 channels, stably expressed in Chinese Hamster Ovary (CHO) cells. The δ-conotoxin family member from C. striatus, δ-SVIE inhibited fast inactivation of NaV channels in dissociated frog sympathetic neurons, and prolonged action potentials on the frog neuromuscular junction [18]. In the same work, δ-SVIE was a potent excitotoxin, when injected into mice, and competed with δ-TxVIA and δ-PVIA for binding to rat brain membranes in autoradiographic experiments. δ-SVIE proved to be highly potent against rNaV1.4 channels expressed in mammalian cells with a Kd value in the nanomolar range [80]. Both δ-SVIE and δ-PVIA conotoxins slowed the time-course of inactivation of Na+ currents and shifted the voltage-dependence of activation and steady-state inactivation to more hyperpolarized potentials in frog sympathetic neurons [157].
δ-EVIA, a 32-residue polypeptide from C. ermineus, is the first conotoxin known to inhibit NaV channel inactivation in neuronal membranes from amphibian and mammals (subtypes rNaV1.2, rNaV1.3 and rNaV1.6) without affecting rat skeletal muscle (subtype rNaV1.4) and human cardiac muscle (subtype hNaV1.5) NaV channels [5]. The NMR structures of δ-EVIA and δ-TxVIA were solved by Volpon et al. [150] and Kohno et al. [77], respectively. Of particular interest in the structure of δ-conotoxins, is the relatively high number of conserved hydrophobic residues exposed on the surface of the toxins. There is some evidence that extracellular binding of these peptides is important to affect inactivation of the channels and that the hydrophobicity of these peptides is probably important for their action. Previous competition experiments measuring the binding of radioactively labeled toxins to membrane preparations containing NaV channels have suggested that their molecular receptor, site 6, is distinct from that of scorpion α-toxins [46; 47; 130]. The molecular target of δ-conotoxins action on NaV channels was recently identified. Leipold et al [80], showed that δ-conotoxin SVIE interacts with a conserved hydrophobic triad (YFV) in the domain IV voltage sensor of mammalian NaV channels. These findings support the hypothesis of hydrophobic interaction of δ-conotoxins with NaV channels [77; 150]. In NaV channels, the S3–S4 linker in domain IV is known to harbor parts of both receptor sites 3 and 6, where site 3 is located more toward S3 and site 6 toward S4 (see Figure 2). This channel section forms the major part of a “receptor site”, i.e., receptor site 3 for α-scorpion toxins and receptor site 6 for δ-conotoxins. With a second moiety, the toxins attack the DIV voltage sensor, whose movement is tightly coupled to the process of inactivation [80].
Earlier studies have shown that a δ-conotoxin from the venom of a mollusc-hunting cone snail, δ-TxVIA, does not compete for binding with site 3 toxins [46]. Additionally, a much larger peptide from the venom of Conus striatus, CsTx, with an activity similar to that of δ-SVIE, only partially competed with the site 3 α-scorpion toxin LqTx [59]. For these reasons, the receptor site for δ-conotoxins has been defined as site 6 [27]. In the same context, bulky δ-conotoxins, such as δ-SVIE bind in the S3– S4 linker in domain IV and may overlap between site 3 and site 6. Likewise, it has been shown for α-scorpion toxins that critical residues for site 3 binding are found in the S3–S4 linker in domain IV [121]. The α-scorpion toxin receptor site suggests a mechanism by which these toxins exert their effects; namely, binding of toxin to site 3 inhibits the movement of the S4 voltage sensor in domain IV [27; 121]. “Voltage-sensor trapping” appears to be a mechanism employed by a variety of peptide toxins that interact with NaV channels [25], and these results suggest that δ-conotoxins act in a similar fashion.
5.3.5. Conus striatus toxin
Conus striatus toxin (CsTx), a 25,000 Da polypeptide toxin from the marine cone snail Conus striatus, was first isolated by Kobayashi, Ohizumi and colleagues [76]. CsTx binds to NaV channels causing a slowing of NaV channel inactivation in both muscle and nerves [40; 43] as well as spontaneous activity in the cardiac musculature [41], while showing no effects on the activation of the channel [59]. CsTx induces a shift in the voltage dependence of steady state inactivation to more negative membrane potentials [59], which is opposite the effect of Goniopora toxin [57]. CsTx has been shown to have a positive inotropic effect in the heart [42] similar to that of Goniopora toxin. The work by Gonoi et al. [59] showed that CsTx appears to bind in a one-to-one fashion with the channel and its action seems to be voltage dependent with an e-fold increase in apparent Kd per 19 mV depolarization. CsTx has a similar mode of action to the δ-conotoxins, although it only partially competed with the site 3 scorpion toxin α-LqTx [59]. This suggests that there may in fact be a partial overlap between the δ-conotoxins and CsTx [47]. Similar to the α-scorpion toxins, CsTx enhances toxin binding to site 2 (e.g., batrachotoxin binding) through an allosteric mechanism, though to a lesser degree than do the α-scorpion toxins [59].
6. Concluding remarks
In this article, we have attempted to outline the remarkable variety of NaV channel-targeted toxins which occur naturally in the marine environment. In trying to make our coverage reasonably comprehensive, but still keep the article concise, we have often cited more detailed reviews in specific areas, rather than comprehensively citing original references. Regarding the choices made, we ask patience and understanding of the authors and colleagues who have produced the myriad of excellent, painstaking primary studies, which ultimately provided the basis for this chapter.
The marine ecosystem offers a source of chemically and functionally diverse toxins that interact with NaV channels. These have already offered considerable insight into the structure and function of the channel protein and provide a rich array of potential pharmacological agents for experimental studies, as well as inspiration for future drug design.
Acknowledgments
This work supported by operating grants from CIHR, and from the HSF of Alberta, NWT and Nunavut. R.J.F. is an AHFMR Medical Scientist.
Abbreviations
| AneTx | anemone toxin (often abbreviated ATX in other papers) |
| ATx | antillatoxin |
| B toxins | ribbon worm toxins |
| CaV, NaV, and KV | voltage-gated ion channels selective for calcium, sodium or potassium, respectively |
| NMDA | N-methyl-D-aspartate |
| CigTx | ciguatoxin |
| CLA | clathrodin |
| CNS | central nervous system |
| PNS | peripheral nervous system |
| DBS | dibromosceptin |
| EC50 | concentration producing 50% of the maximal effect |
| GPT | Goniopora (coral) toxin |
| KTx | kalkitoxin |
| IC50 | concentration producing 50% inhibition |
| NSP | neurotoxic shellfish poisoning |
| LD50 | lethal dose/concentration for 50% of population |
| Kd | dissociation contant |
| OTx | ostreotoxin |
| PbTx-n | brevetoxins where n = 1–10 |
| PSP/PST | paralytic shellfish poisoning/toxins |
| STx | saxitoxin |
| TTx | tetrodotoxin |
| TTx-s | TTx sensitive |
| TTx-r | TTx resistant |
- Samples Availability: Available from the authors.
References
- Alonso, D.; Khalil, Z.; Satkunanthan, N.; Livett, B.G. Drugs from the sea: conotoxins as drug leads for neuropathic pain and other neurological conditions. [Review] [37 refs]. Mini-Rev. in Med. Chem 2003, 3, 785–787. [Google Scholar]
- Baden, D.G. Brevetoxins: unique polyether dinoflagellate toxins. [Review] [56 refs]. FASEB J 1989, 3, 1807–1817. [Google Scholar]
- Baden, D.G.; Bourdelais, A.J.; Jacocks, H.; Michelliza, S.; Naar, J. Natural and derivative brevetoxins: historical background, multiplicity, and effects. [Review] [42 refs]. Environ. Health Persp 2005, 113, 621–625. [Google Scholar]
- Baden, D.G.; Mende, T.J. Toxicity of two toxins from the Florida red tide marine dinoflagellate, Ptychodiscus brevis. Toxicon 1982, 20, 457–461. [Google Scholar]
- Barbier, J.; Lamthanh, H.; Le Gall, F.; Favreau, P.; Benoit, E.; Chen, H.; Gilles, N.; Ilan, N.; Heinemann, S.H.; Gordon, D.; Menez, A.; Molgo, J. A delta-conotoxin from Conus ermineus venom inhibits inactivation in vertebrate neuronal Na+ channels but not in skeletal and cardiac muscles. J. Biol. Chem 2004, 279, 4680–4685. [Google Scholar]
- Barnham, K.J.; Dyke, T.R.; Kem, W.R.; Norton, R.S. Structure of neurotoxin B-IV from the marine worm Cerebratulus lacteus: a helical hairpin cross-linked by disulphide bonding. J. Mol. Biol 1997, 268, 886–902. [Google Scholar]
- Benoit, E.; Legrand, A.M.; Dubois, J.M. Effects of ciguatoxin on current and voltage clamped frog myelinated nerve fibre. Toxicon 1986, 24, 357–364. [Google Scholar]
- Beress, L.; Wunderer, G.; Wachter, E. Amino acid sequence of toxin III from Anemonia sulcata. Hoppe Seylers Z. Physiol. Chem 1977, 358, 985–988. [Google Scholar]
- Berman, F.W.; Gerwick, W.H.; Murray, T.F. Antillatoxin and kalkitoxin, ichthyotoxins from the tropical cyanobacterium Lyngbya majuscula, induce distinct temporal patterns of NMDA receptor-mediated neurotoxicity. Toxicon 1999, 37, 1645–1648. [Google Scholar]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res 2000, 28, 235–242. [Google Scholar]
- Bickmeyer, U.; Drechsler, C.; Kock, M.; Assmann, M. Brominated pyrrole alkaloids from marine Agelas sponges reduce depolarization-induced cellular calcium elevation. Toxicon 2004, 44, 45–51. [Google Scholar]
- Bidard, J.N.; Vijverberg, H.P.; Frelin, C.; Chungue, E.; Legrand, A.M.; Bagnis, R.; Lazdunski, M. Ciguatoxin is a novel type of Na+ channel toxin. J. Biol. Chem 1984, 259, 8353–8357. [Google Scholar]
- Blumenthal, K.M.; Keim, P.S.; Heinrikson, R.L.; Kem, W.R. Structure and action of heteronemertine polypeptide toxins. Amino acid sequence of Cerebratulus lacteus toxin B-II and revised structure of toxin B-IV. J. Biol. Chem 1981, 256, 9063–9067. [Google Scholar]
- Blumenthal, K.M.; Kem, W.R. Structure and action of heteronemertine polypeptide toxins. Primary structure of Cerebratulus lacteus toxin B-IV. J. Biol. Chem 1976, 251, 6025–6029. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep 2004, 21, 1–49. [Google Scholar]
- Bottein Dechraoui, M.Y.; Ramsdell, J.S. Type B brevetoxins show tissue selectivity for voltage-gated sodium channels: comparison of brain, skeletal muscle and cardiac sodium channels. Toxicon 2003, 41, 919–927. [Google Scholar]
- Bruhn, T.; Schaller, C.; Schulze, C.; Sanchez-Rodriguez, J.; Dannmeier, C.; Ravens, U.; Heubach, J.F.; Eckhardt, K.; Schmidtmayer, J.; Schmidt, H.; Aneiros, A.; Wachter, E.; Beress, L. Isolation and characterisation of five neurotoxic and cardiotoxic polypeptides from the sea anemone Anthopleura elegantissima. Toxicon 2001, 39, 693–702. [Google Scholar]
- Bulaj, G.; DeLaCruz, R.; zimi-Zonooz, A.; West, P.; Watkins, M.; Yoshikami, D.; Olivera, B.M. Delta-conotoxin structure/function through a cladistic analysis. Biochemistry 2001, 40, 13201–13208. [Google Scholar]
- Bulaj, G.; West, P.J.; Garrett, J.E.; Marsh, M.; Zhang, M.M.; Norton, R.S.; Smith, B.J.; Yoshikami, D.; Olivera, B.M. Novel conotoxins from Conus striatus and Conus kinoshitai selectively block TTX-resistant sodium channels. Biochemistry 2005, 44, 7259–7265. [Google Scholar]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Burgess, J.G.; Wright, P.C. Marine cyanobacteria - a profile source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar]
- Cardellina, J.H.; Marner, F.J.; Moore, R.E. Seaweed dermatitis: structure of lyngbyatoxin A. Science 1979, 204, 193–195. [Google Scholar]
- Catterall, W.A. Structure and function of voltage-sensitive ion channels. Science 1988, 242, 50–61. [Google Scholar]
- Catterall, W.A. Molecular properties of a superfamily of plasma-membrane cation channels. Curr. Opin. Cell Biol 1994, 6, 607–615. [Google Scholar]
- Catterall, W.A. From ionic currents to molecular mechanisms: the structure and function of voltage-gated sodium channels. Neuron 2000, 26, 13–25. [Google Scholar]
- Catterall, W.A. Molecular mechanisms of gating and drug block of sodium channels. Novartis Found. Symp 2002, 241, 206–218. [Google Scholar]
- Catterall, W.A.; Gainer, M. Interaction of brevetoxin A with a new receptor site on the sodium channel. Toxicon 1985, 23, 497–504. [Google Scholar]
- Cestele, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. [Review] [111 refs]. Biochimie 2000, 82, 883–892. [Google Scholar]
- Chahine, M.; Sirois, J.; Marcotte, P.; Chen, L.-Q.; Kallen, R.G. Extrapore residues of the S5–S6 loop of domain 2 of the voltage-gated skeletal muscle sodium channel (rSkM1) contribute to the μ-conotoxin GIIIA binding site. Biophys. J 1998, 75, 236–246. [Google Scholar]
- Choudhary, G.; Shang, L.; Li, X.; Dudley, S.C., Jr. Energetic localization of saxitoxin in its channel binding site. Biophys. J. 2002, 83, 912–919. [Google Scholar]
- Choudhary, G.; Yotsu-Yamashita, M.; Shang, L.; Yasumoto, T.; Dudley, S.C., Jr. Interactions of the C-11 hydroxyl of tetrodotoxin with the sodium channel outer vestibule. Biophys. J. 2003, 84, 287–294. [Google Scholar]
- Córdova, J.L.; Escudero, C.; Bustamante, J. Bloom inside the bloom: intracellular bacteria multiplication within toxic dinoflagellates. Revista de Biologia Marina y Oceanografia 2003, 38, 57–67. [Google Scholar]
- Corpuz, G.P.; Jacobsen, R.B.; Jimenez, E.C.; Watkins, M.; Walker, C.; Colledge, C.; Garrett, J.E.; McDougal, O.; Li, W.; Gray, W.R.; Hillyard, D.R.; Rivier, J.; McIntosh, J.M.; Cruz, L.J.; Olivera, B.M. Definition of the M-conotoxin superfamily: characterization of novel peptides from molluscivorous Conus venoms. Biochemistry 2005, 44, 8176–8186. [Google Scholar]
- Cruz, L.J.; Gray, W.R.; Olivera, B.M.; Zeikus, R.D.; Kerr, L.; Yoshikami, D.; Moczydlowski, E. Conus geographus toxins that discriminate between neuronal and muscle sodium channels. J. Biol. Chem 1985, 260, 9280–9288. [Google Scholar]
- Daly, N.L.; Ekberg, J.A.; Thomas, L.; Adams, D.J.; Lewis, R.J.; Craik, D.J. Structures of muO-conotoxins from Conus marmoreus. Inhibitors of tetrodotoxin (TTX)-sensitive and TTXresistant sodium channels in mammalian sensory neurons. J. Biol. Chem 2004, 279, 25774–25782. [Google Scholar]
- Denac, H.; Mevissen, M.; Scholtysik, G. Structure, function and pharmacology of voltagegated sodium channels. [Review] [288 refs]. Naunyn-Schmiedebergs Arch. Pharmacol 2000, 362, 453–479. [Google Scholar]
- Dravid, S.M.; Baden, D.G.; Murray, T.F. Brevetoxin augments NMDA receptor signaling in murine neocortical neurons. Brain Res 2005, 1031, 30–38. [Google Scholar]
- Dudley, S.C., Jr; Chang, N.; Hall, J.; Lipkind, G.; Fozzard, H.A.; French, R.J. μ-Conotoxin interactions with the voltage-gated Na+ channel predict a clockwise arrangement of domains. J. Gen. Physiol. 2000, 116, 679–689. [Google Scholar]
- Edwards, D.J.; Marquez, B.L.; Nogle, L.M.; McPhail, K.; Goeger, D.E.; Roberts, M.A.; Gerwick, W.H. Structure and biosynthesis of the jamaicamides, new mixed polyketide-peptide neurotoxins from the marine cyanobacterium Lyngbya majuscula.[see comment]. Chem. & Biol 2004, 11, 817–833. [Google Scholar]
- Ellis, S. Introduction to symposium - brevetoxins: chemistry and pharmacology of ‘red tide’ toxins from Ptychodiscus brevis (formerly Gymnodinium breve). Toxicon 1985, 23, 469–472. [Google Scholar]
- Endean, R.; Gyr, P.; Surridge, J. The pharmacological actions on guinea-pig ileum of crude venoms from the marine gastropods Conus striatus and Conus magus. Toxicon 1977, 15, 327–337. [Google Scholar]
- Endean, R.; Gyr, P.; Surridge, J. The effects of crude venoms of Conus magus and Conus striatus on the contractile response and electrical activity of guinea-pig cardiac musculature. Toxicon 1979, 17, 381–395. [Google Scholar]
- Endean, R.; Surridge, J.; Gyr, P. Some effects of crude venom from the cones Conus striatus and Conus magus on isolated guinea-pig atria. Toxicon 1977, 15, 369–374. [Google Scholar]
- Endean, R.; Williams, H.; Gyr, P.; Surridge, J. Some effects on muscle and nerve of crude venom from the gastropod Conus striatus. Toxicon 1976, 14, 267–274. [Google Scholar]
- Escalona de Motta, G.; Rentas, A.L.; Mercado, J.A.; Tosteson, T.R.; Gonzalez, I. Identification of a sodium channel toxin from a Caribbean benthic dinoflagellate. Toxicon 1996, 34, 308. [Google Scholar]
- Fainzilber, M.; Gordon, D.; Hasson, A.; Spira, M.E.; Zlotkin, E. Mollusc-specific toxins from the venom of Conus textile neovicarius. Eur. J. Biochem 1991, 202, 589–595. [Google Scholar]
- Fainzilber, M.; Kofman, O.; Zlotkin, E.; Gordon, D. A new neurotoxin receptor site on sodium channels is identified by a conotoxin that affects sodium channel inactivation in molluscs and acts as an antagonist in rat brain. J. Biol. Chem 1994, 269, 2574–2580. [Google Scholar]
- Fainzilber, M.; Lodder, J.C.; Kits, K.S.; Kofman, O.; Vinnitsky, I.; Van Rietschoten, J.; Zlotkin, E.; Gordon, D. A new conotoxin affecting sodium current inactivation interacts with the delta-conotoxin receptor site. J. Biol. Chem 1995, 270, 1123–1129. [Google Scholar]
- Fainzilber, M.; van der, S.R.; Lodder, J.C.; Li, K.W.; Geraerts, W.P.; Kits, K.S. New sodium channel-blocking conotoxins also affect calcium currents in Lymnaea neurons. Biochemistry 1995, 34, 5364–5371. [Google Scholar]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep 1999, 16, 155–198. [Google Scholar]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep 2002, 19, 1–48. [Google Scholar]
- Freitas, J.C.; Rangel, M.; Oliveira, J.S.; Zaharenko, A.J.; Rozas, E. An online of marine toxicology studies in the Brazilian coast. Comm. Toxicol 2003, 9, 137–159. [Google Scholar]
- French, R.J.; Terlau, H. Sodium channel toxins--receptor targeting and therapeutic potential. [Review] [85 refs]. Curr. Med. Chem 2004, 11, 3053–3064. [Google Scholar]
- Fu, L.Y.; Li, Y.; Cheng, L.; Zhou, H.Y.; Yao, W.X.; Xia, G.J.; Jiang, M.X. Effect of sea anemone toxin anthopleurin-Q on sodium current in guinea pig ventricular myocytes. Acta Pharmacol. Sinica 2001, 22, 1107–1112. [Google Scholar]
- Fujiwara, M.; Muramatsu, I.; Hidaka, H.; Ikushima, S.; Ashida, K. Effects of Goniopora toxin, a polypeptide isolated from coral, on electromechanical properties of rabbit myocardium. J. Pharmacol. Exp. Ther 1979, 210, 153–157. [Google Scholar]
- Gallacher, S.; Flynn, K.J.; Franco, J.M.; Brueggemann, E.E.; Hines, H.B. Evidence for production of paralytic shellfish toxins by bacteria associated with Alexandrium spp. (Dinophyta) in culture. Appl. Env. Microbiol 1997, 63, 239–245. [Google Scholar]
- Gerwick, W.H.; Fenical, W.; Norris, J.N. Chemical variation in the tropical seaweed Stypopodium zonale (dictyotaceae). Phytochemistry 1985, 24, 1279–1283. [Google Scholar]
- Gonoi, T.; Ashida, K.; Feller, D.; Schmidt, J.; Fujiwara, M.; Catterall, W.A. Mechanism of action of a polypeptide neurotoxin from the coral Goniopora on sodium channels in mouse neuroblastoma cells. Mol. Pharmacol 1986, 29, 347–354. [Google Scholar]
- Gonoi, T.; Hille, B. Gating of Na channels. Inactivation modifiers discriminate among models. J. Gen. Physiol 1987, 89, 253–274. [Google Scholar]
- Gonoi, T.; Ohizumi, Y.; Kobayashi, J.; Nakamura, H.; Catterall, W.A. Actions of a polypeptide toxin from the marine snail Conus striatus on voltage-sensitive sodium channels. Molecular Pharmacology 1987, 32, 691–698. [Google Scholar]
- Hashimoto, Y.; Ashida, K. Screening of toxic corals and isolation of a toxic polypeptide from Goniopora spp. Pub. the Seto Marine Biol. Lab 1973, 20, 703–711. [Google Scholar]
- Hasson, A.; Shon, K.J.; Olivera, B.M.; Spira, M.E. Alterations of voltage-activated sodium current by a novel conotoxin from the venom of Conus gloriamaris. J. Neurophysiol 1995, 73, 1295–1301. [Google Scholar]
- Hill, J.M.; Alewood, P.F.; Craik, D.J. Conotoxin TVIIA, a novel peptide from the venom of Conus tulipa 2. Three-dimensional solution structure. Eur. J. Biochem 2000, 267, 4649–4657. [Google Scholar]
- Hille, B. The permeability of the sodium channel to metal cations in myelinated nerve. J. Gen. Physiol 1972, 59, 637–658. [Google Scholar]
- Hille, B. Ion channels of excitable membranes; Sunderland; Sinauer Associates, Inc, 2001. [Google Scholar]
- Hillyard, D.R.; Olivera, B.M.; Woodward, S.; Corpuz, G.P.; Gray, W.R.; Ramilo, C.A.; Cruz, L.J. A molluscivorous Conus toxin: conserved frameworks in conotoxins. Biochemistry 1989, 28, 358–361. [Google Scholar]
- Honma, T.; Iso, T.; Ishida, M.; Nagashima, Y.; Shiomi, K. Occurrence of type 3 sodium channel peptide toxins in two species of sea anemones (Dofleinia armata and Entacmaea ramsayi). Toxicon 2003, 41, 637–639. [Google Scholar]
- Howell, M.L.; Blumenthal, K.M. Mutagenesis of Cerebratulus lacteus neurotoxin B-IV identifies NH2-terminal sequences important for biological activity. J. Biol. Chem 1991, 266, 12884–12888. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: visual molecular dynamics. J. Mol. Graphics 1996, 14, 33–38. [Google Scholar]
- Ishida, H.; Nozawa, A.; Totoribe, K.; Muramatsu, N.; Nukaya, H.; Tsuji, K.; Yamaguchi, K.; Yasumoto, T.; Kaspar, H.; Berkett, N.; Kosuge, T. Brevetoxin B1, a new polyether marine toxin from the New Zealand shellfish, Austrovenus stutchburyi. Tetrahedron Letters 1995, 36, 725–728. [Google Scholar]
- Isom, L.L. Sodium channel β subunits: anything but auxiliary. Neuroscientist 2001, 7(1), 42–54. [Google Scholar]
- Isom, L.L.; De Jongh, K.S.; Catterall, W.A. Auxiliary subunits of voltage-gated ion channels. Neuron 1994, 12, 1183–1194. [Google Scholar]
- Jeglitsch, G.; Rein, K.; Baden, D.G.; Adams, D.J. Brevetoxin-3 (PbTx-3) and its derivatives modulate single tetrodotoxin-sensitive sodium channels in rat sensory neurons. J. Pharmacol. Exp. Ther 1998, 284, 516–525. [Google Scholar]
- Keizer, D.W.; West, P.J.; Lee, E.F.; Yoshikami, D.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structural basis for tetrodotoxin-resistant sodium channel binding by mu-conotoxin SmIIIA. J. Biol. Chem 2003, 278, 46805–46813. [Google Scholar]
- Kem, W.R. Purification and characterization of a new family of polypeptide neurotoxins from the heteronemertine Cerebratulus lacteus (Leidy). J. Biol. Chem 1976, 251, 4184–4192. [Google Scholar]
- Kem, W.R.; Parten, B.; Pennington, M.W.; Price, D.A.; Dunn, B.M. Isolation, characterization, and amino acid sequence of a polypeptide neurotoxin occurring in the sea anemone Stichodactyla helianthus. Biochemistry 1989, 28, 3483–3489. [Google Scholar]
- Kobayashi, J.; Nakamura, H.; Hirata, Y.; Ohizumi, Y. Isolation of a cardiotonic glycoprotein, striatoxin, from the venom of the marine snail Conus Striatus. Biochem. Biophys. Res. Commun 1982, 105, 1389–1395. [Google Scholar]
- Kohno, T.; Sasaki, T.; Kobayashi, K.; Fainzilber, M.; Sato, K. Three-dimensional solution structure of the sodium channel agonist/antagonist delta-conotoxin TxVIA. J. Biol. Chem 2002, 277, 36387–36391. [Google Scholar]
- Kulagina, N.V.; O’shaughnessy, T.J.; Ma, W.; Ramsdell, J.S.; Pancrazio, J.J. Pharmacological effects of the marine toxins, brevetoxin and saxitoxin, on murine frontal cortex neuronal networks. Toxicon 2004, 44, 669–676. [Google Scholar]
- Lehane, L.; Lewis, R.J. Ciguatera: recent advances but the risk remains. Int. J. Food Microbiol 2000, 61, 91–125. [Google Scholar]
- Leipold, E.; Hansel, A.; Olivera, B.M.; Terlau, H.; Heinemann, S.H. Molecular interaction of delta-conotoxins with voltage-gated sodium channels. FEBS Lett 2005, 579, 3881–3884. [Google Scholar]
- LePage, K.T.; Goeger, D.; Yokokawa, F.; Asano, T.; Shioiri, T.; Gerwick, W.H.; Murray, T.F. The neurotoxic lipopeptide kalkitoxin interacts with voltage-sensitive sodium channels in cerebellar granule neurons. Toxicol. Lett 2005, 158, 133–139. [Google Scholar]
- Lewis, R.J. Ciguatoxins are potent ichthyotoxins. Toxicon 1992, 30, 207–211. [Google Scholar]
- Lewis, R.J. The changing face of ciguatera. [Review] [64 refs]. Toxicon 2001, 39, 97–106. [Google Scholar]
- Li, R.A.; Ennis, I.L.; French, R.J.; Dudley, S.C., Jr.; Tomaselli, G.F.; Marbán, E. Clockwise domain arrangement of the sodium channel revealed by μ−conotoxin (GIIIA) docking orientation. J. Biol. Chem. 2001, 276(14), 11072–11077. [Google Scholar]
- Li, R.A.; Ennis, I.L.; Xue, T.; Nguyen, H.M.; Tomaselli, G.F.; Goldin, A.L.; Marban, E. Molecular basis of isoform-specific μ−conotoxin block of cardiac, skeletal muscle, and brain Na+ channels. J. Biol. Chem 2003, 278, 8717–8724. [Google Scholar]
- Li, W.I.; Berman, F.W.; Okino, T.; Yokokawa, F.; Shioiri, T.; Gerwick, W.H.; Murray, T.F. Antillatoxin is a marine cyanobacterial toxin that potently activates voltage-gated sodium channels. Proc. Natl. Acad. Sci. U. S. A 2001, 98, 7599–7604. [Google Scholar]
- Lindel, T.; Hoffman, H.; Hochgurtel, M.; Pawlik, J.R. Structure-activity relationship of inhibition of fish feeding by sponge-derived and synthetic pyrrole-imidazole alkaloids. J. Chem. Ecol. 2000, 26. [Google Scholar]
- Llewellyn, L.; Negri, A.; Quilliam, M. High affinity for the rat brain sodium channel of newly discovered hydroxybenzoate saxitoxin analogues from the dinoflagellate Gymnodinium catenatum. Toxicon 2004, 43, 101–104. [Google Scholar]
- Lombet, A.; Bidard, J.N.; Lazdunski, M. Ciguatoxin and brevetoxins share a common receptor site on the neuronal voltage-dependent Na+ channel. FEBS Lett 1987, 219, 355–359. [Google Scholar]
- Long, S.B.; Campbell, E.B.; MacKinnon, R. Crystal structure of a mammalian voltagedependent Shaker family K+ channel. Science 2005, 309, 897–903. [Google Scholar]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of sodium channel toxins: directed cytotoxicity assays of purified ciguatoxins, brevetoxins, saxitoxins, and seafood extracts. J. AOAC Int 1995, 78, 521–527. [Google Scholar]
- Martins, C.A.; Alvito, P.; Tavares, M.J.; Pereira, P.; Doucette, G.; Franca, S. Reevaluation of production of paralytic shellfish toxin by bacteria associated with dinoflagellates of the Portuguese coast. Appl. Environ. Microbiol 2003, 69, 5693–5698. [Google Scholar]
- Mattei, C.; Dechraoui, M.Y.; Molgo, J.; Meunier, F.A.; Legrand, A.M.; Benoit, E. Neurotoxins targetting receptor site 5 of voltage-dependent sodium channels increase the nodal volume of myelinated axons. J. Neurosci. Res 1999, 55, 666–673. [Google Scholar]
- McFarren, E.F.; Silva, F.J.; Tanabe, H.; Wilson, W.B.; Campbell, J.E.; Lewis, K.H. The occurrence of a ciguatera-like poison in oysters, clams, and Gymnodinium breve cultures. Toxicon 1965, 3, 111–123. [Google Scholar]
- McIntosh, J.M.; Hasson, A.; Spira, M.E.; Gray, W.R.; Li, W.; Marsh, M.; Hillyard, D.R.; Olivera, B.M. A new family of conotoxins that blocks voltage-gated sodium channels. J. Biol. Chem 1995, 270, 16796–16802. [Google Scholar]
- Morales, J.J.; Rodriguez, A.D. The structure of clathrodin, a novel alkaloid isolated from the Caribbean Sea sponge Agelas clathrodes. J. Nat. Prod 1991, 54, 629–631. [Google Scholar]
- Mynderse, J.S.; Moore, R.E.; Kashiwagi, M.; Norton, T.R. Antileukemia activity in the Osillatoriaceae: isolation of Debromoaplysiatoxin from Lyngbya. Science 1977, 196, 538–540. [Google Scholar]
- Nielsen, K.J.; Watson, M.; Adams, D.J.; Hammarstrom, A.K.; Gage, P.W.; Hill, J.M.; Craik, D.J.; Thomas, L.; Adams, D.; Alewood, P.F.; Lewis, R.J. Solution structure of mu-conotoxin PIIIA, a preferential inhibitor of persistent tetrodotoxin-sensitive sodium channels. J. Biol. Chem 2002, 277, 27247–27255. [Google Scholar]
- Nishida, S.; Fujita, S.; Warashina, A.; Satake, M.; Tamiya, N. Amino acid sequence of a sea anemone toxin from Parasicyonis actinostoloides. Eur. J. Biochem 1985, 150, 171–173. [Google Scholar]
- Noda, M.; Suzuki, H.; Numa, S.; Stühmer, W. A single point mutation confers tetrodotoxin and saxitoxin insensitivity on the sodium channel II. FEBS Lett 1989, 259, 213–216. [Google Scholar]
- Nogle, L.M.; Okino, T.; Gerwick, W.H. Antillatoxin B, a neurotoxic lipopeptide from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod 2001, 64, 983–985. [Google Scholar]
- Norton, R.S. Structure and structure-function relationships of sea anemone proteins that interact with the sodium channel. [Review] [127 refs]. Toxicon 1991, 29, 1051–1084. [Google Scholar]
- Norton, R.S.; Pallaghy, P.K. The cystine knot structure of ion channel toxins and related polypeptides. Toxicon 1998, 36, 1573–1583. [Google Scholar]
- Ogata, N.; Ohishi, Y. Molecular diversity of structure and function of the voltage-gated Na+ channels. [Review] [82 refs]. Jap. J. Pharmacol 2002, 88, 365–377. [Google Scholar]
- Olivera, B.M. E.E. Just Lecture, 1996. Conus venom peptides, receptor and ion channel targets, and drug design: 50 million years of neuropharmacology. Mol. Biol. Cell 1997, 8, 2101–2109. [Google Scholar]
- Olivera, B.M. Conus venom peptides: reflections from the biology of clades and species. Annu. Rev. Ecol. Syst 2002, 33, 25–47. [Google Scholar]
- Orjala, J.; Nagle, D.G.; Hsu, V.L.; Gerwick, W.H. Antillatoxin: an exceptionally ichthyotoxic cyclic lipopeptide from the tropical cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc 1995, 117, 8281–8282. [Google Scholar]
- Pavlov, E; Bladen, C; Diao, C; French, RJ. Bacterial Na channels: progenitors, progeny, or parallel evolution? In Bacterial ion channels and eukaryotic homologues; Kubalski, A., Martinac, B., Eds.; American Society for Microbiology, 2005; pp. 191–207. [Google Scholar]
- Pearn, J. Neurology of ciguatera. J. Neurol. Neurosurg. Psych 2001, 70, 4–8. [Google Scholar]
- Penzotti, J.L.; Fozzard, H.A.; Lipkind, G.M.; Dudley, S.C., Jr. Differences in saxitoxin and tetrodotoxin binding revealed by mutagenesis of the Na+ channel outer vestibule. Biophys. J. 1998, 75, 2647–2657. [Google Scholar]
- Penzotti, J.L.; Lipkind, G.; Fozzard, H.A.; Dudley, S.C., Jr. Specific neosaxitoxin interactions with the Na+ channel outer vestibule determined by mutant cycle analysis. Biophys. J. 2001, 80, 698–706. [Google Scholar]
- Pierce, R.H. Red tide (Ptychodiscus brevis) toxin aerosols: a review. Toxicon 1986, 24, 955–965. [Google Scholar]
- Poli, M.A.; Mende, T.J.; Baden, D.G. Brevetoxins, unique activators of voltage-sensitive sodium channels, bind to specific sites in rat brain synaptosomes. Mol. Pharmacol 1986, 30, 129–135. [Google Scholar]
- Pomati, F.; Rossetti, C.; Calamari, D.; Neilan, B.A. Effects of saxitoxin (STx) and Veratridine on Bacterial Na+-K+ fluxes: a Prokaryote-based STx bioassay. Appl. Environ. Microbiol 2003, 69, 7371–7376. [Google Scholar]
- Reimer, N.S.; Yasunobu, C.L.; Yasunobu, K.T.; Norton, T.R. Amino acid sequence of the Anthopleura xanthogrammica heart stimulant, anthopleurin-B. J. Biol. Chem 1985, 260, 8690–8693. [Google Scholar]
- Rein, K.S.; Baden, D.G.; Gawley, R.E. Conformational analysis of the sodium channel modulator, brevetoxin A, comparison with brevetoxin B conformations, and a hypothesis about the common pharmacophore of the “site 5” toxins. J. Org. Chem 1994, 59, 2101–2106. [Google Scholar]
- Rein, K.S.; Lynn, B.; Gawley, R.E.; Baden, D.G. Brevetoxin B: chemical modifications, synaptosome binding, toxicity, and an unexpected conformational effect. J. Org. Chem 1994, 59, 2107–2113. [Google Scholar]
- Ren, D.; Navarro, B.; Xu, H.; Yue, L.; Shi, Q.; Clapham, D.E. A prokaryotic voltage-gated sodium channel. Science 2001, 294, 2372–2375. [Google Scholar]
- Rentas, A.L.; Rosa, R.; Rodriguez, A.D.; De Motta, G.E. Effect of alkaloid toxins from tropical marine sponges on membrane sodium currents. Toxicon 1995, 33, 491–497. [Google Scholar]
- Ritson-Williams, R.; Yotsu-Yamashita, M.; Paul, V.J. Ecological functions of tetrodotoxin in a deadly polyclad flatworm. Proc. Natl. Acad. Sci. U. S. A 2006, 103, 3176–3179. [Google Scholar]
- Rogers, J.C.; Qu, Y.; Tanada, T.N.; Scheuer, T.; Catterall, W.A. Molecular determinants of high affinity binding of alpha-scorpion toxin and sea anemone toxin in the S3–S4 extracellular loop in domain IV of the Na+ channel alpha subunit. J. Biol. Chem 1996, 271, 15950–15962. [Google Scholar]
- Rohl, C.A.; Boeckman, F.A.; Baker, C.; Scheuer, T.; Catterall, W.A.; Klevit, R.E. Solution structure of the sodium channel inactivation gate. Biochemistry 1999, 38, 855–861. [Google Scholar]
- Sabry, O.M.; Andrews, S.; McPhail, K.L.; Goeger, D.E.; Yokochi, A.; LePage, K.T.; Murray, T.F.; Gerwick, W.H. Neurotoxic meroditerpenoids from the tropical marine brown alga Stypopodium flabelliforme. J. Nat. Prod 2005, 68, 1022–1030. [Google Scholar]
- Safo, P.; Rosenbaum, T.; Shcherbatko, A.; Choi, D.Y.; Han, E.; Toledo-Aral, J.J.; Olivera, B.M.; Brehm, P.; Mandel, G. Distinction among neuronal subtypes of voltage-activated sodium channels by mu-conotoxin PIIIA. J. Neurosci 2000, 20, 76–80. [Google Scholar]
- Scheffler, J.J.; Tsugita, A.; Linden, G.; Schweitz, H.; Lazdunski, M. The amino acid sequence of toxin V from Anemonia sulcata. Biochem. Biophys. Res. Commun 1982, 107, 272–278. [Google Scholar]
- Schlief, T.; Schonherr, R.; Imoto, K.; Heinemann, S.H. Pore properties of rat brain II sodium channels mutated in the selectivity filter domain. Eur. Biophys. J 1996, 25, 75–91. [Google Scholar]
- Seibert, A.L.; Liu, J.; Hanck, D.A.; Blumenthal, K.M. Arg-14 loop of site 3 anemone toxins: effects of glycine replacement on toxin affinity. Biochemistry 2003, 42, 14515–14521. [Google Scholar]
- Seoh, S.A.; Sigg, D.; Papazian, D.M.; Bezanilla, F. Voltage-sensing residues in the S2 and S4 segments of the Shaker K+ channel. Neuron 1996, 16, 1159–1167. [Google Scholar]
- Sharkey, R.G.; Jover, E.; Couraud, F.; Baden, D.G.; Catterall, W.A. Allosteric modulation of neurotoxin binding to voltage-sensitive sodium channels by Ptychodiscus brevis toxin 2. Mol. Pharmacol 1987, 31, 273–278. [Google Scholar]
- Shichor, I.; Fainzilber, M.; Pelhate, M.; Malecot, C.O.; Zlotkin, E.; Gordon, D. Interactions of delta-conotoxins with alkaloid neurotoxins reveal differences between the silent and effective binding sites on voltage-sensitive sodium channels. J. Neurochem 1996, 67, 2451–2460. [Google Scholar]
- Shimizu, Y. Microalgal metabolites. Chem. Rev 1993, 93, 1685–1698. [Google Scholar]
- Shon, K.J.; Grilley, M.M.; Marsh, M.; Yoshikami, D.; Hall, A.R.; Kurz, B.; Gray, W.R.; Imperial, J.S.; Hillyard, D.R.; Olivera, B.M. Purification, characterization, synthesis, and cloning of the lockjaw peptide from Conus purpurascens venom. Biochemistry 1995, 34, 4913–4918. [Google Scholar]
- Shon, K.J.; Hasson, A.; Spira, M.E.; Cruz, L.J.; Gray, W.R.; Olivera, B.M. Delta-conotoxin GmVIA, a novel peptide from the venom of Conus gloriamaris. Biochemistry 1994, 33, 11420–11425. [Google Scholar]
- Shon, K.-J.; Olivera, B.M.; Watkins, M.; Jacobsen, R.B.; Gray, W.R.; Floresca, C.Z.; Cruz, L.J.; Hillyard, D.R.; Brink, A.; Terlau, H.; Yoshikami, D. μ-conotoxin PIIIA, a new peptide for discriminating among tetrodotoxin-sensitive Na channel subtypes. J. Neurosci. 1998, 18(12), 4473–4481. [Google Scholar]
- Smith, E.A.; Grant, F.; Ferguson, C.M.; Gallacher, S. Biotransformations of paralytic shellfish toxins by bacteria isolated from bivalve molluscs. Appl. Env. Microbiol 2001, 67, 2345–2353. [Google Scholar]
- Snyder, R.V.; Guerrero, M.A.; Sinigalliano, C.D.; Winshell, J.; Perez, R.; Lopez, J.V.; Rein, K.S. Localization of polyketide synthase encoding genes to the toxic dinoflagellate Karenia brevis. Phytochemistry 2005, 66, 1767–1780. [Google Scholar]
- Stephan, M.M.; Potts, J.F.; Agnew, W.S. The μI skeletal muscle sodium channel: mutation E403Q eliminates sensitivity to tetrodotoxin but not to μ-conotoxins GIIIA and GIIIB. J. Membrane Biol 1994, 137, 1–8. [Google Scholar]
- Strachan, L.C.; Lewis, R.J.; Nicholson, G.M. Differential actions of pacific ciguatoxin-1 on sodium channel subtypes in mammalian sensory neurons. J Pharmacol. Exp. Ther 1999, 288, 379–388. [Google Scholar]
- Suarez-Isla, B; Guzman-Mendez, L. Mareas rojas y toxinas marinas. Floraciones de algas nocivas; Santiago de Chile; Editorial Universitaria, 1998. [Google Scholar]
- Sudarslal, S.; Majumdar, S.; Ramasamy, P.; Dhawan, R.; Pal, P.P.; Ramaswami, M.; Lala, A.K.; Sikdar, S.K.; Sarma, S.P.; Krishnan, K.S.; Balaram, P. Sodium channel modulating activity in a delta-conotoxin from an Indian marine snail. FEBS Lett 2003, 553, 209–212. [Google Scholar]
- Sunahara, S.; Muramoto, K.; Tenma, K.; Kamiya, H. Amino acid sequence of two sea anemone toxins from Anthopleura fuscoviridis. Toxicon 1987, 25, 211–219. [Google Scholar]
- Tanaka, M.; Hainu, M.; Yasunobu, K.T.; Norton, T.R. Amino acid sequence of the Anthopleura xanthogrammica heart stimulant, anthopleurin A. Biochemistry 1977, 16, 204–208. [Google Scholar]
- Terlau, H.; Heinemann, S.H.; Stühmer, W.; Pusch, M.; Conti, F.; Imoto, K.; Numa, S. Mapping the site of block by tetrodotoxin and saxitoxin of sodium channel II. FEBS Lett 1991, 293, 93–96. [Google Scholar]
- Terlau, H.; Olivera, B.M. Conus venoms: a rich source of novel ion channel-targeted peptides. [Review] [196 refs]. Physiol. Rev 2004, 84, 41–68. [Google Scholar]
- Terlau, H.; Shon, K.J.; Grilley, M.; Stocker, M.; Stuhmer, W.; Olivera, B.M. Strategy for rapid immobilization of prey by a fish-hunting marine snail. Nature 1996, 381, 148–151. [Google Scholar]
- Terlau, H.; Stocker, M.; Shon, K.-J.; McIntosh, J.M.; Olivera, B.M. μO-conotoxin MrVIA inhibits mammalian sodium channels, but not through site I. J. Neurophysiol. 1996, 76(3), 1423–1429. [Google Scholar]
- Todt, H.; Dudley, S.C., Jr; Kyle, J.W.; French, R.J.; Fozzard, H.A. Ultra-slow inactivation in mu1 Na+ channels is produced by a structural rearrangement of the outer vestibule. Biophys. J. 1999, 76, 1335–1345. [Google Scholar]
- Vasquez, M.; Gruttner, C.; Moeller, B.; Moore, E.R. Limited selection of sodium channel blocking toxin-producing bacteria from paralytic shellfish toxin-contaminated mussels (Aulacomya ater). Res. Microbiol 2002, 153, 333–338. [Google Scholar]
- Vilin, Y.Y.; Ruben, P.C. Slow inactivation in voltage-gated sodium channels: molecular substrates and contributions to channelopathies. Cell Biochem. Biophys 2001, 35, 171–190. [Google Scholar]
- Volpon, L.; Lamthanh, H.; Barbier, J.; Gilles, N.; Molgo, J.; Menez, A.; Lancelin, J.M. NMR solution structures of delta-conotoxin EVIA from Conus ermineus that selectively acts on vertebrate neuronal Na+ channels. J. Biol. Chem 2004, 279, 21356–21366. [Google Scholar]
- Wang, S.Y.; Wang, G.K. Voltage-gated sodium channels as primary targets of diverse lipid-soluble neurotoxins. [Review] [68 refs]. Cell. Signal 2003, 15, 151–159. [Google Scholar]
- Wemmer, D.E.; Kumar, N.V.; Metrione, R.M.; Lazdunski, M.; Drobny, G.; Kallenbach, N.R. NMR analysis and sequence of toxin II from the sea anemone Radianthus paumotensis. Biochemistry 1986, 25, 6842–6849. [Google Scholar]
- Wen, P.H.; Blumenthal, K.M. Role of electrostatic interactions in defining the potency of neurotoxin B-IV from Cerebratulus lacteus. J. Biol. Chem 1996, 271, 29752–29758. [Google Scholar]
- Wen, P.H.; Blumenthal, K.M. Structure and function of Cerebratulus lacteus neurotoxin B-IV: tryptophan-30 is critical for function while lysines-18, -19, -29, and -33 are not required. Biochemistry 1997, 36, 13435–13440. [Google Scholar]
- Wessels, M.; Konig, G.M.; Wright, A.D. A new tyrosine kinase inhibitor from the marine brown alga Stypopodium zonale. J. Nat. Prod 1999, 62, 927–930. [Google Scholar]
- West, P.J.; Bulaj, G.; Garrett, J.E.; Olivera, B.M.; Yoshikami, D. Mu-conotoxin SmIIIA, a potent inhibitor of tetrodotoxin-resistant sodium channels in amphibian sympathetic and sensory neurons. Biochemistry 2002, 41, 15388–15393. [Google Scholar]
- West, P.J.; Bulaj, G.; Yoshikami, D. Effects of delta-conotoxins PVIA and SVIE on sodium channels in the amphibian sympathetic nervous system. J. Neurophysiol 2005, 94, 3916–3924. [Google Scholar]
- Widmer, H.; Billeter, M.; Wuthrich, K. Three-dimensional structure of the neurotoxin ATX Ia from Anemonia sulcata in aqueous solution determined by nuclear magnetic resonance spectroscopy. Proteins 1989, 6, 357–371. [Google Scholar]
- Wolfe, G.V. The chemical defense ecology of marine unicellular plankton: constraints, mechanisms, and impacts. [Review] [270 refs]. Biol. Bull 2000, 198, 225–244. [Google Scholar]
- Wu, M.; Okino, T.; Nogle, L.M.; Marquez, B.L.; Williamson, R.T.; Sitachitta, N.; Berman, F.W.; Murray, T.F.; McGough, K.; Jacobs, R.; Colsen, K.; Asano, T.; Yokokawa, F.; Shioiri, T.; Gerwick, W.H. Structure, synthesis, and biological properties of kalkitoxin, a novel neurotoxin from the marine cyanobacterium Lyngbya majuscule. J. Am. Chem. Soc 2000, 122, 1241–1242. [Google Scholar]
- Wunderer, G.; Eulitz, M. Amino-acid sequence of toxin I from Anemonia sulcata. Eur. J. Biochem 1978, 89, 11–17. [Google Scholar]
- Wunderer, G.; Fritz, H.; Wachter, E.; Machleidt, W. Amino-acid sequence of a coelenterate toxin: toxin II from Anemonia sulcata. Eur. J. Biochem 1976, 68, 193–198. [Google Scholar]
- Yanagawa, Y.; Abe, T.; Satake, M.; Odani, S.; Suzuki, J.; Ishikawa, K. A novel sodium channel inhibitor from Conus geographus: purification, structure, and pharmacological properties. Biochemistry 1988, 27, 6256–6262. [Google Scholar]
- Yasumoto, T.; murata, M. Marine toxins. Chem. Rev 1993, 93, 1897–1909. [Google Scholar]
- Yotsu-Yamashita, M.; Kim, Y.H.; Dudley, S.C., Jr; Choudhary, G.; Pfahnl, A.; Oshima, Y.; Daly, J.W. The structure of zetekitoxin AB, a saxitoxin analog from the Panamanian golden frog Atelopus zeteki: a potent sodium-channel blocker. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 4346–4351. [Google Scholar]
- Zingone, A.; Enevoldsen, H.O. The diversity of harmful algal blooms: a challenge for science and management. Ocean & Coastal Management 2000, 43, 725–748. [Google Scholar]
- Zlotkin, E. The insect voltage-gated sodium channel as target of insecticides. [Review] [73 refs]. Ann. Rev. Entomol 1999, 44, 429–455. [Google Scholar]
- Zykova, T.A.; Kozlovskaia, E.P. (Amino acid sequence of a neurotoxin from the anemone Radianthus macrodactylus). Bioorg. Khim 1989, 15, 1301–1306.
- Zykova, T.A.; Kozlovskaia, E.P. (Disulfide bonds in neurotoxin-III from the sea anenome Radianthus macrodactylus). Bioorg. Khim 1989, 15, 904–907.
- Zykova, T.A.; Kozlovskaia, E.P.; Eliakov, G.B. (Amino acid sequence of neurotoxin II from the sea anemone Radianthus macrodactylus). Bioorg. Khim 1988, 14, 878–882.
- Zykova, T.A.; Kozlovskaia, E.P.; Eliakov, G.B. (Amino acid sequence of neurotoxins IV and V from the sea anemone Radianthus macrodactylus). Bioorg. Khim. 1988, 14, 1489–1494.
Figure 1.
Schematic presentation of the membrane topology of the α subunit of an NaV channel. Homologous domains (DI-DIV), and, within each domain, transmembrane segments (S1–S6) are labeled. Each domain has a P loop (P) between S5–S6 segment and a cationic, voltage-sensing (+) S4 segment. The hydrophobic sequence (IFM) in the intracellular DIII-DIV connecting loop is required for fast inactivation, and serves as inactivation latch for the Na+ channel. Representative NaV channels have been cloned from different tissues including: rat brain (rNaV1.2), rat skeletal muscle (rNaV1.4), adult rat heart (rNaV1.5), and PNS (rNaV1.9). Residue numbering for each domain is indicated.
Figure 1.
Schematic presentation of the membrane topology of the α subunit of an NaV channel. Homologous domains (DI-DIV), and, within each domain, transmembrane segments (S1–S6) are labeled. Each domain has a P loop (P) between S5–S6 segment and a cationic, voltage-sensing (+) S4 segment. The hydrophobic sequence (IFM) in the intracellular DIII-DIV connecting loop is required for fast inactivation, and serves as inactivation latch for the Na+ channel. Representative NaV channels have been cloned from different tissues including: rat brain (rNaV1.2), rat skeletal muscle (rNaV1.4), adult rat heart (rNaV1.5), and PNS (rNaV1.9). Residue numbering for each domain is indicated.
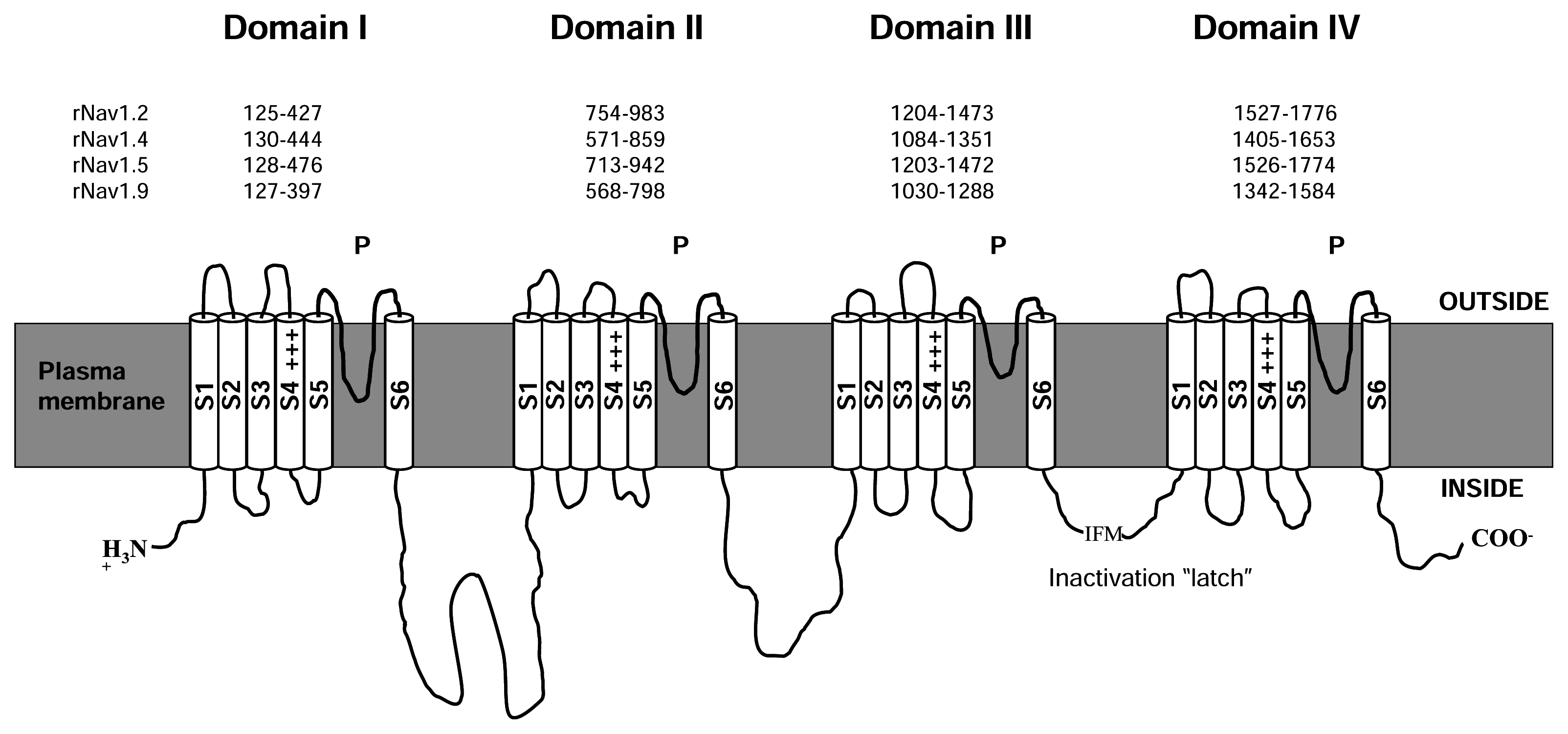
Figure 2.
NaV channel receptor sites mapped onto the structure of KV1.2. A. Side-view of KV1.2 with P loops modified to resemble more closely the likely structure of NaV channel P loops, which allow block by agents such as STx, TTx, and μ-conotoxin. Only DII and DIV shown; S1–S3 red, S4 blue and S5-P loop-S6 cyan. B. Top view of KV1.2 showing all four domains with domains DII and DIV colored as in A. The eye at the bottom shows the point of view for part A. The figures were produced using VMD software [68] and final images rendered with POV-ray ( http://www.povray.org/).
Figure 2.
NaV channel receptor sites mapped onto the structure of KV1.2. A. Side-view of KV1.2 with P loops modified to resemble more closely the likely structure of NaV channel P loops, which allow block by agents such as STx, TTx, and μ-conotoxin. Only DII and DIV shown; S1–S3 red, S4 blue and S5-P loop-S6 cyan. B. Top view of KV1.2 showing all four domains with domains DII and DIV colored as in A. The eye at the bottom shows the point of view for part A. The figures were produced using VMD software [68] and final images rendered with POV-ray ( http://www.povray.org/).
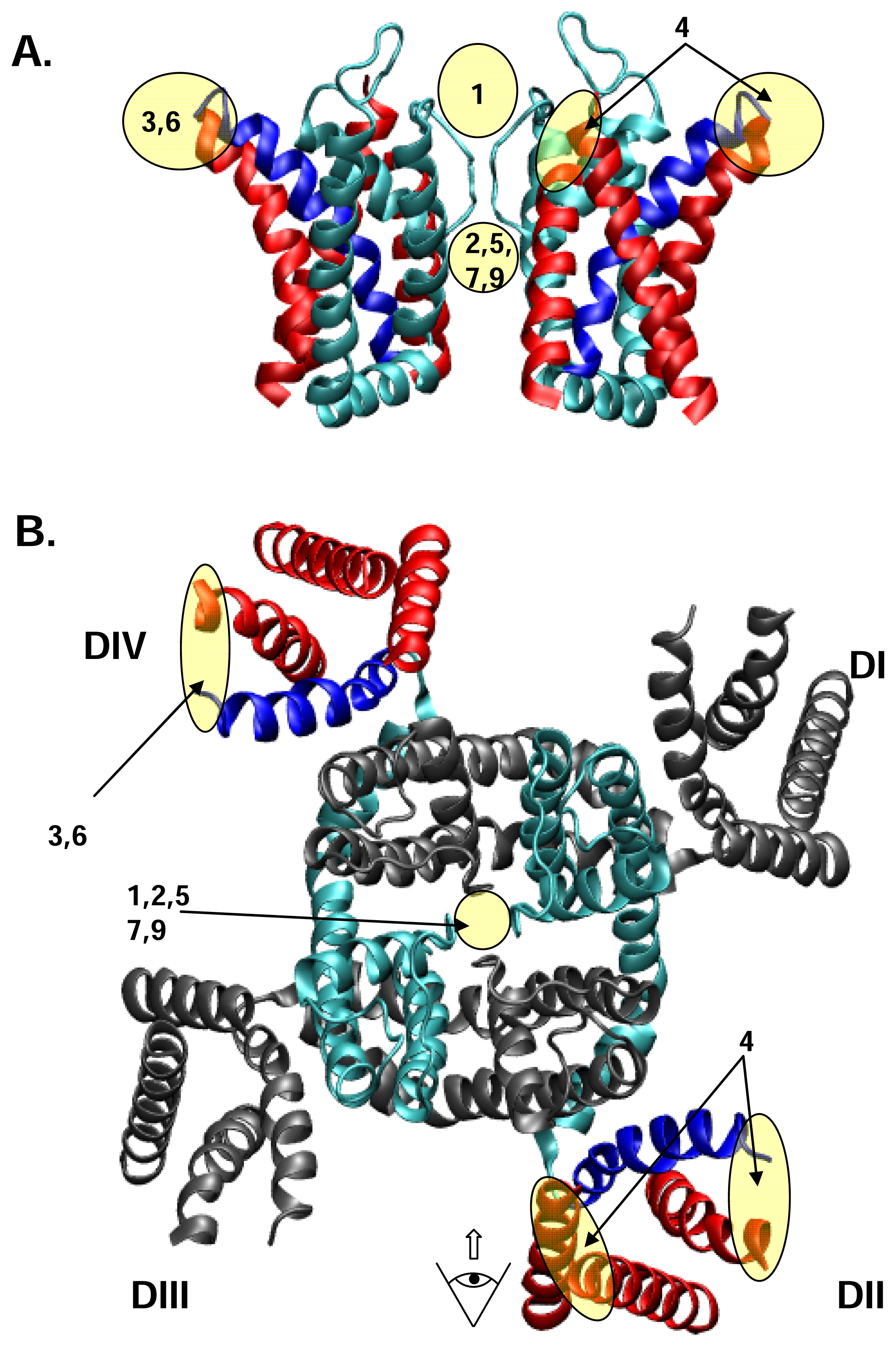
Figure 3.
Chemical structures of non-peptide NaV channels blockers. A. Tetrodotoxin, and B. Saxitoxin, both of which are tricyclic, guanidinium-derived marine toxins. C. Meroditerpenoids from the marine brown alga Stypopodium flabelliforme, which act as NaV channel modifiers: 1. 2β,3α-Epitaondiol; 2. Flabellinol; 3. Flabellinone. The structures of the meroditerpenoids were reproduced from Sabry et al. [123], with permission.
Figure 3.
Chemical structures of non-peptide NaV channels blockers. A. Tetrodotoxin, and B. Saxitoxin, both of which are tricyclic, guanidinium-derived marine toxins. C. Meroditerpenoids from the marine brown alga Stypopodium flabelliforme, which act as NaV channel modifiers: 1. 2β,3α-Epitaondiol; 2. Flabellinol; 3. Flabellinone. The structures of the meroditerpenoids were reproduced from Sabry et al. [123], with permission.
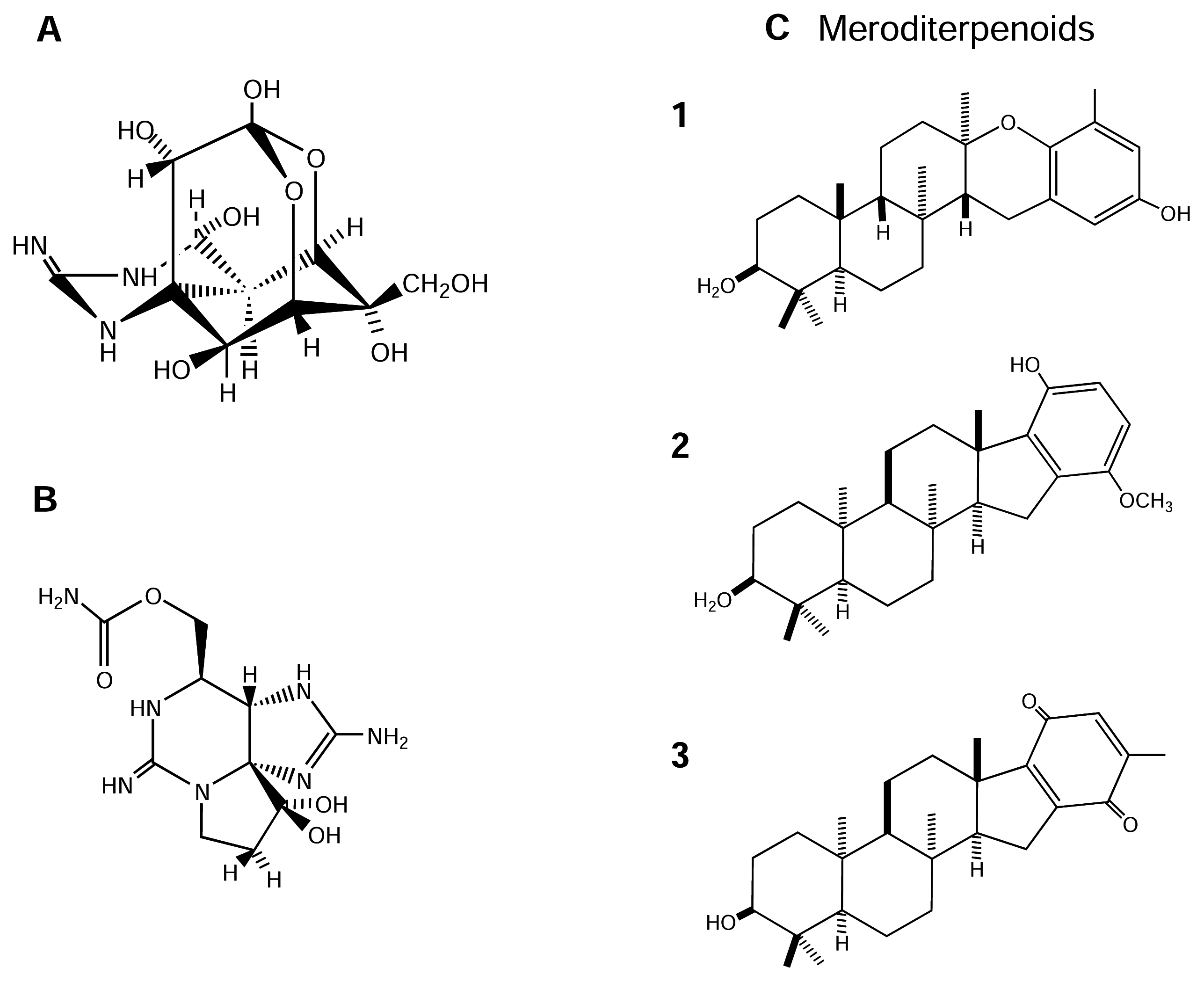
Figure 4.
Primary sequences of μ-conotoxins from five different species of Conus. The consensus numbering defined here is used in the following figure. Blue (positive), red (negative), green (cysteine), pE (pyroglutamate), O (hydroxyproline). Carboxy termini are amidated.
Figure 4.
Primary sequences of μ-conotoxins from five different species of Conus. The consensus numbering defined here is used in the following figure. Blue (positive), red (negative), green (cysteine), pE (pyroglutamate), O (hydroxyproline). Carboxy termini are amidated.
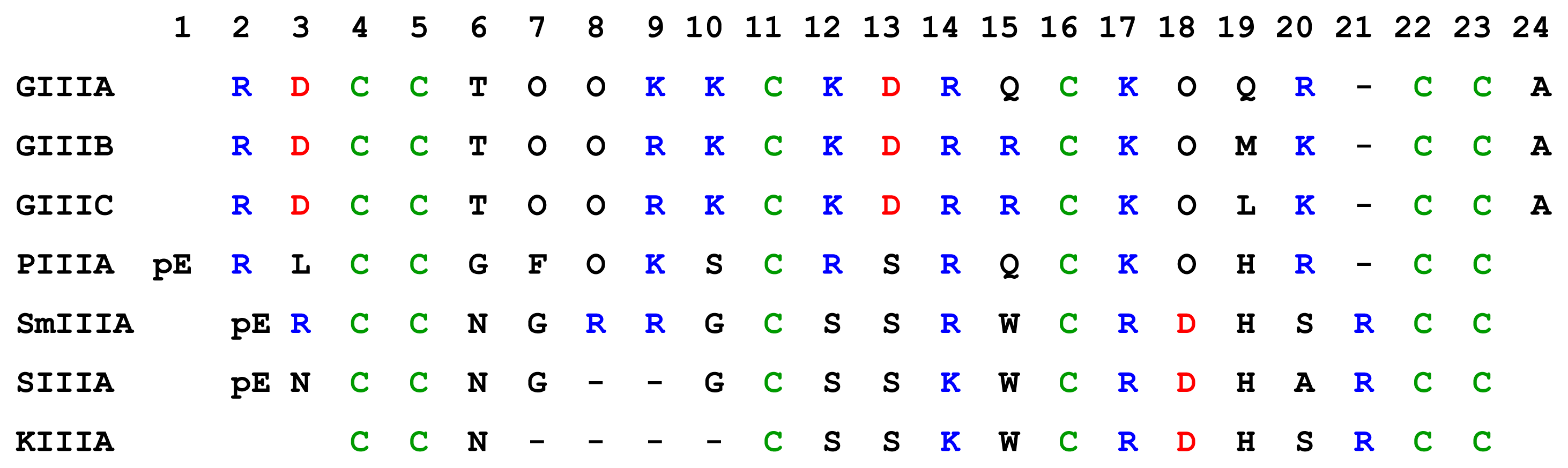
Figure 5.
NMR-derived structures for three μ-conotoxins. Atomic coordinates were taken from RCSB Protein Data Base [10]; coordinates used were as follows: GIIIA, 1TCG; PIIIA, 1RD1, structure #19; SmIIIA, 1Q2J, structure #19. Software for figure generation was as in Figure 2. The conotoxin molecules are oriented in similar directions with an axis along the R14 side-chain facing down. In the lower set, images are rotated 180 degrees around the R14 axis with respect to the upper set. Positively charged residues are shown in blue; negatively charged residues are shown in red. Residue numbering follows the consensus numbering from the alignment in Figure 4.
Figure 5.
NMR-derived structures for three μ-conotoxins. Atomic coordinates were taken from RCSB Protein Data Base [10]; coordinates used were as follows: GIIIA, 1TCG; PIIIA, 1RD1, structure #19; SmIIIA, 1Q2J, structure #19. Software for figure generation was as in Figure 2. The conotoxin molecules are oriented in similar directions with an axis along the R14 side-chain facing down. In the lower set, images are rotated 180 degrees around the R14 axis with respect to the upper set. Positively charged residues are shown in blue; negatively charged residues are shown in red. Residue numbering follows the consensus numbering from the alignment in Figure 4.

Figure 6.
Chemical structures of some NaV channel peptide blockers. A. Jamaicamides A-C: lipopeptides from dinoflagellate Lyngbya majuscula. B. Kalkitoxin: a thiazoline-containing lipopeptide toxin also from dinoflagellate L. majuscula. The structures of jamaicamides were reproduced from Edwards et al.[38], with permission. The structure of kalkitoxin was reproduced from LePage et al.[81], with permission.
Figure 6.
Chemical structures of some NaV channel peptide blockers. A. Jamaicamides A-C: lipopeptides from dinoflagellate Lyngbya majuscula. B. Kalkitoxin: a thiazoline-containing lipopeptide toxin also from dinoflagellate L. majuscula. The structures of jamaicamides were reproduced from Edwards et al.[38], with permission. The structure of kalkitoxin was reproduced from LePage et al.[81], with permission.
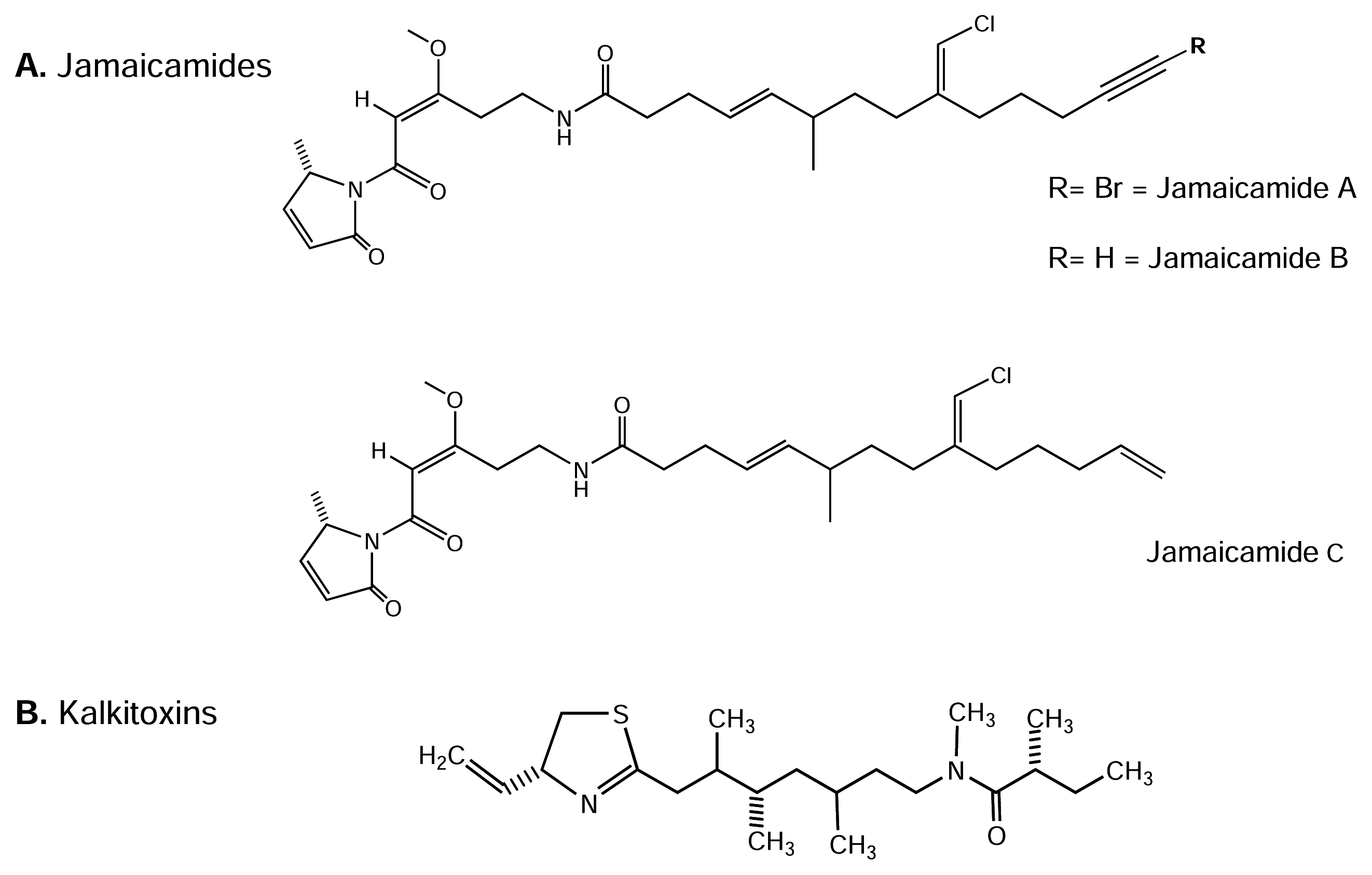
Figure 7.
Chemical structures of non-peptide gating modifiers of NaV channels. A. Clathrodin (CLA) and B. Dibromosceptrin (DBS) alkanoid toxins from Agelas sponge. The chemical structure of CLA was reproduced from Morales and Rodriguez [96], with permission; the structure of DBS was reproduced from Bickmeyer et al [11], with permission.
Figure 7.
Chemical structures of non-peptide gating modifiers of NaV channels. A. Clathrodin (CLA) and B. Dibromosceptrin (DBS) alkanoid toxins from Agelas sponge. The chemical structure of CLA was reproduced from Morales and Rodriguez [96], with permission; the structure of DBS was reproduced from Bickmeyer et al [11], with permission.
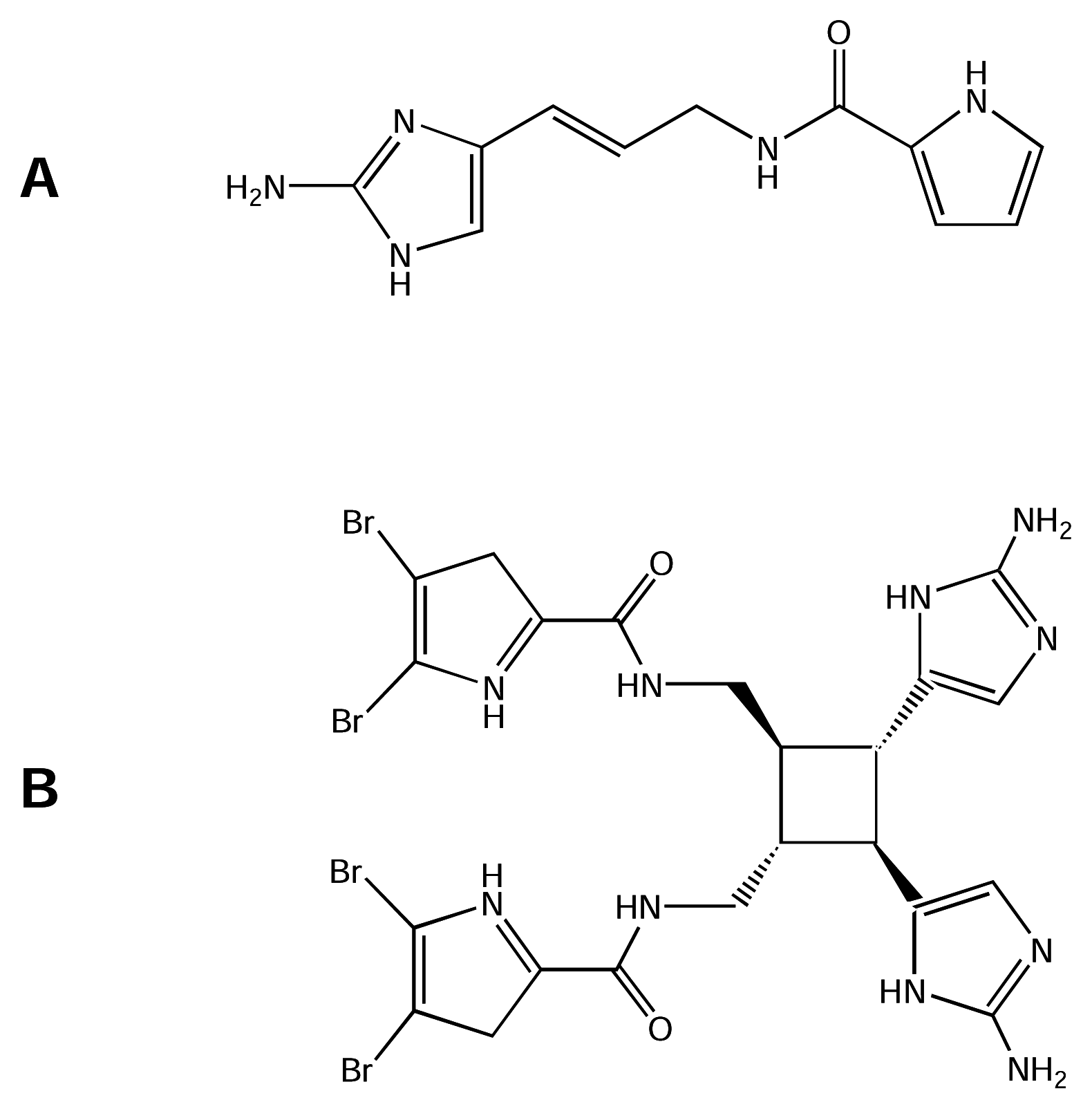
Figure 8.
Molecular structures of polyether ladder toxins which are NaV channel modifiers. A. Brevetoxins A and B are isolated from marine dinoflagellate, Karenia brevis.B. Ciguatoxins (Pacific; P-CigTx and Caribbean; C-CigTx) from marine dinoflagellate, Gambierdiscus toxicus. Structures of brevetoxins were reproduced from Baden et al. [3], with permission. Structures of ciguatoxins were reproduced from Pearn [109], with permission.
Figure 8.
Molecular structures of polyether ladder toxins which are NaV channel modifiers. A. Brevetoxins A and B are isolated from marine dinoflagellate, Karenia brevis.B. Ciguatoxins (Pacific; P-CigTx and Caribbean; C-CigTx) from marine dinoflagellate, Gambierdiscus toxicus. Structures of brevetoxins were reproduced from Baden et al. [3], with permission. Structures of ciguatoxins were reproduced from Pearn [109], with permission.
Figure 9.
The cyclic lipopeptide structures of antillatoxins, ATx and ATx-B, from Lyngbya majuscula. The structures of antillatoxins were reproduced from Li et al. [86], with permission.
Figure 9.
The cyclic lipopeptide structures of antillatoxins, ATx and ATx-B, from Lyngbya majuscula. The structures of antillatoxins were reproduced from Li et al. [86], with permission.
Figure 10.
Examples of peptide toxin backbone structures shown with disulphide bonds (green) created with VMD [68] of B-IV [6], AneTx Ia [158], μO-conotoxin MrVIB [34] and δ-conotoxin TxVIB [77].
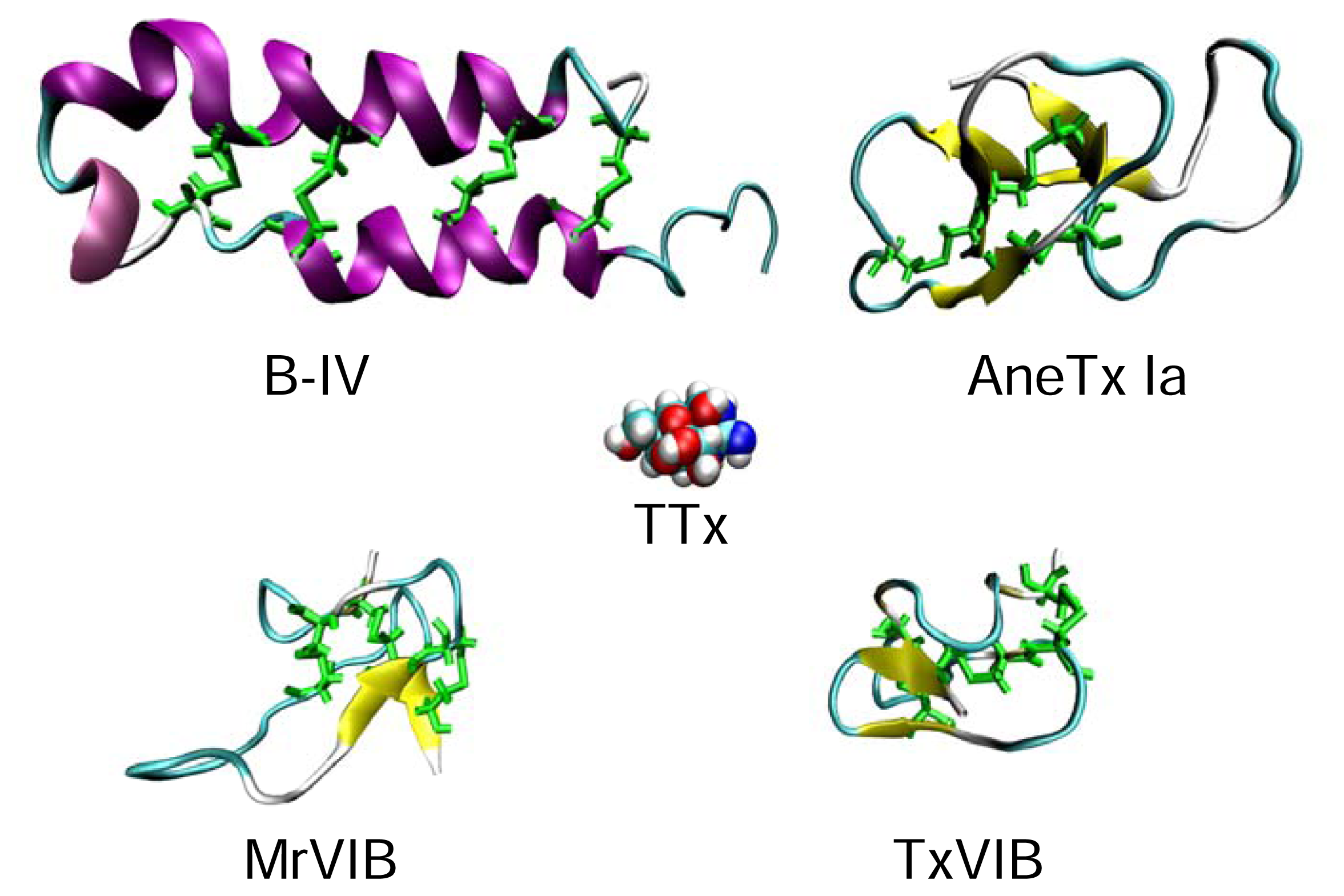

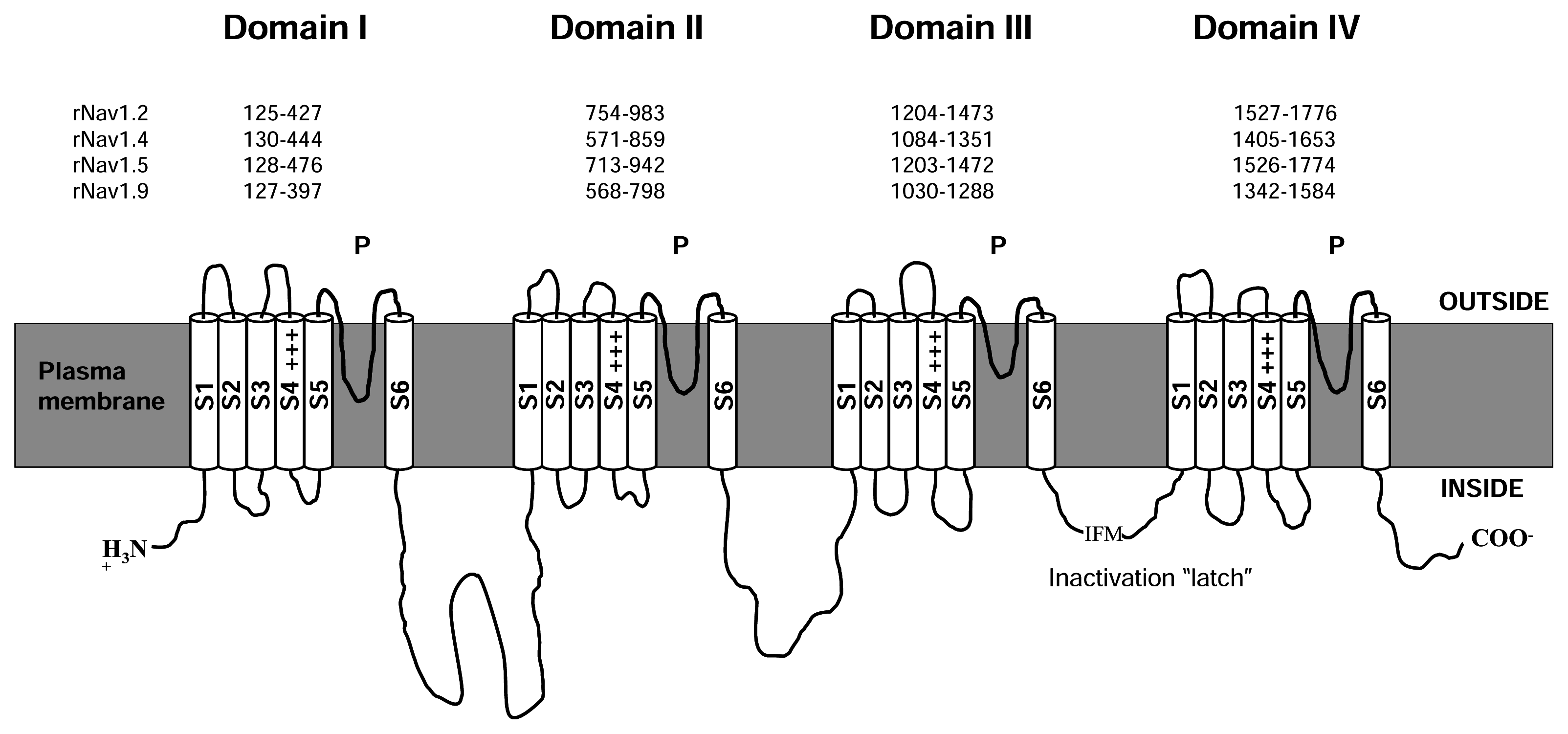{kind=link}
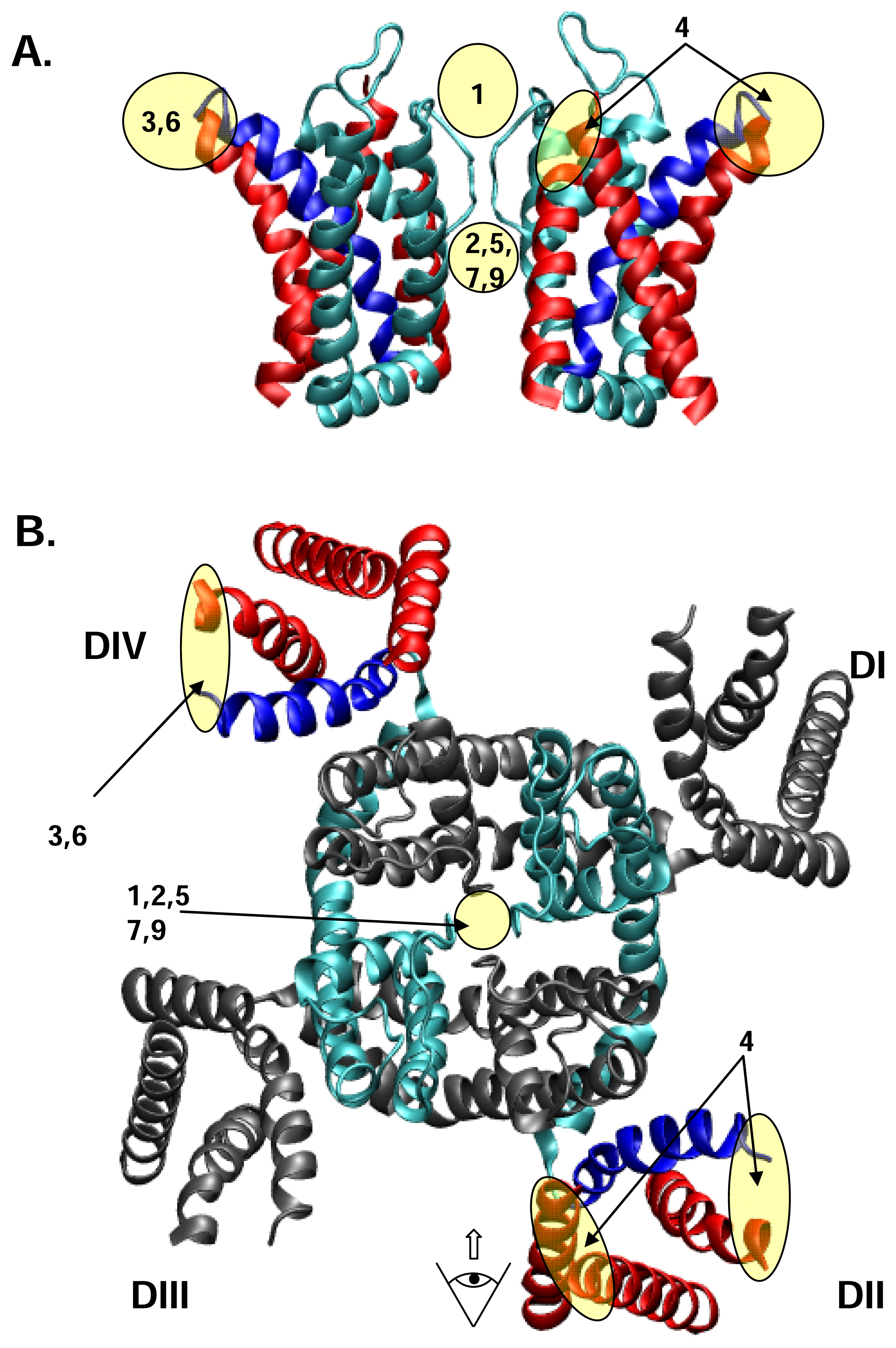{kind=link}
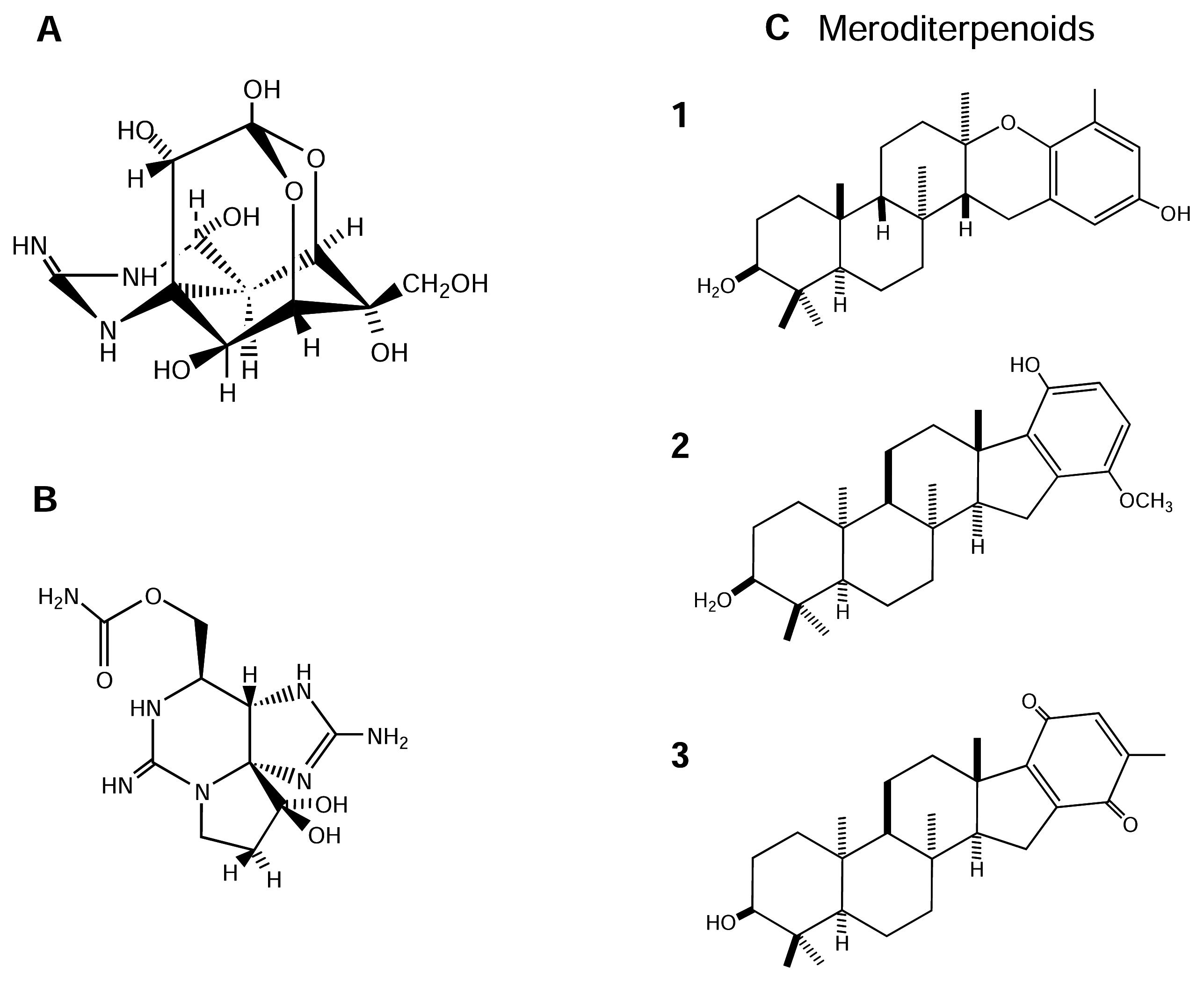{kind=link}
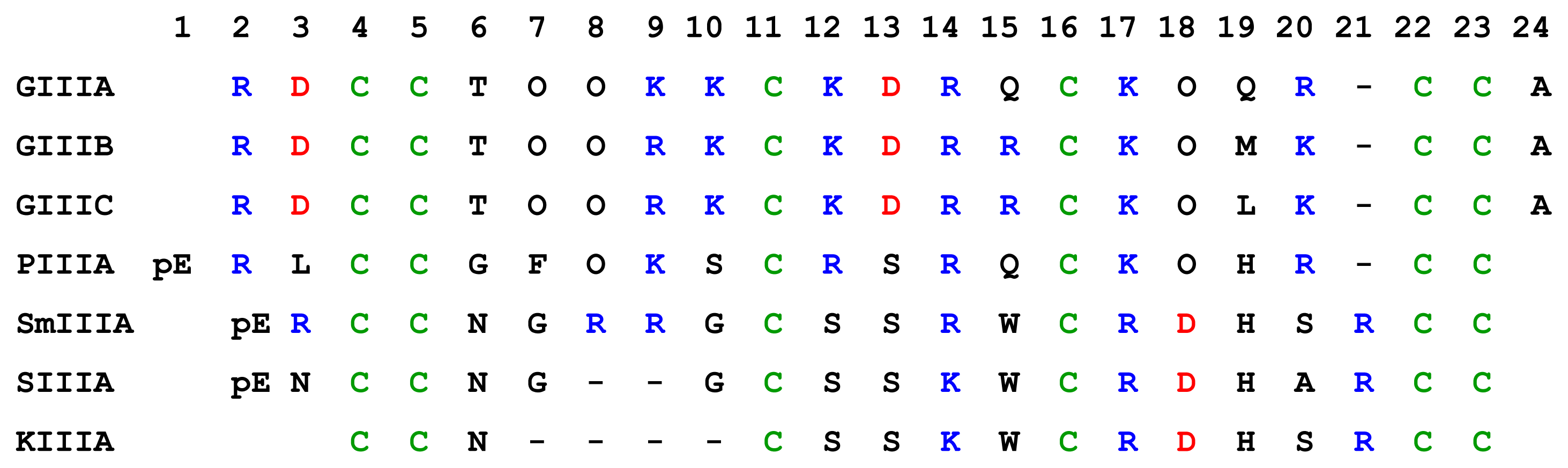{kind=link}
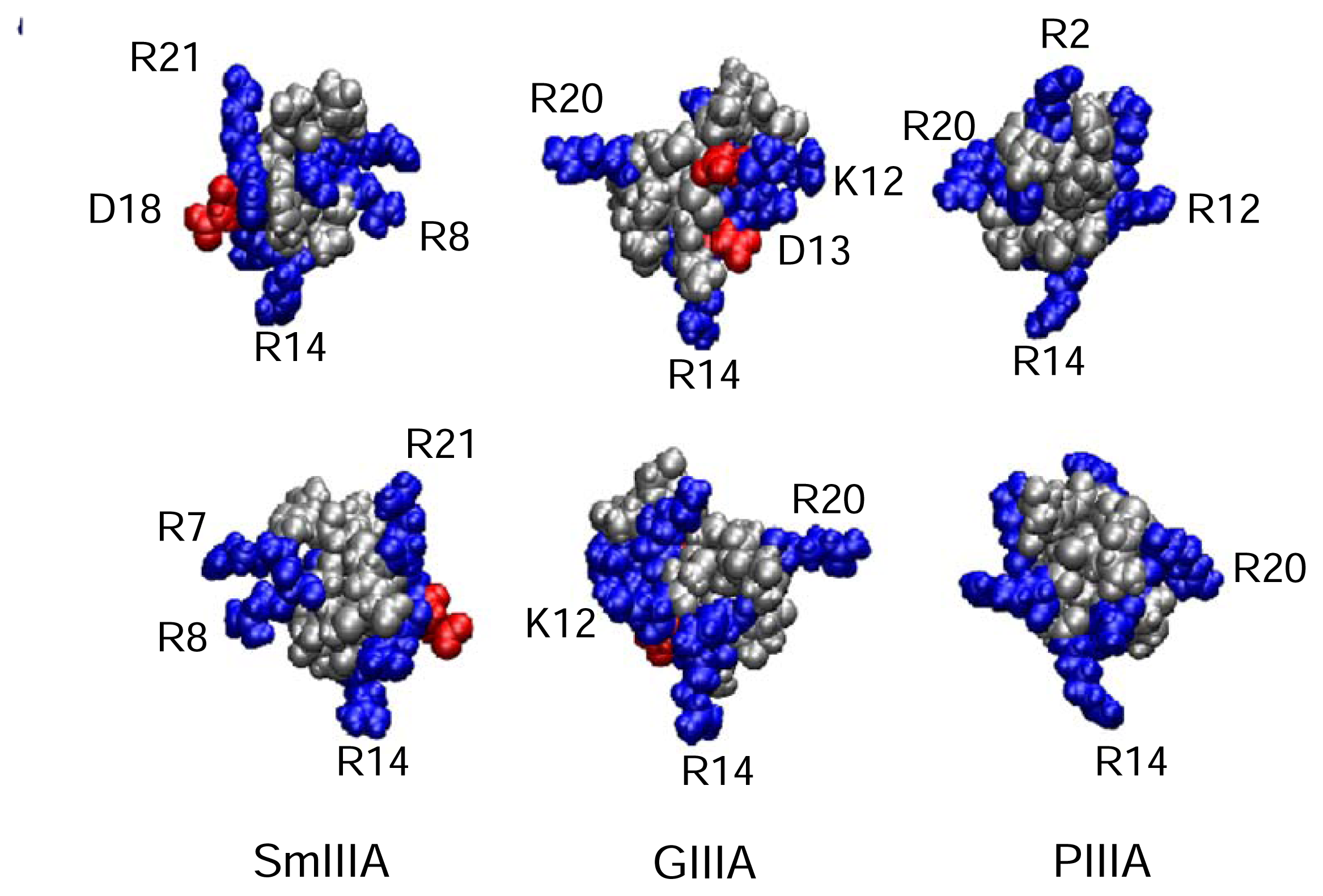{kind=link}
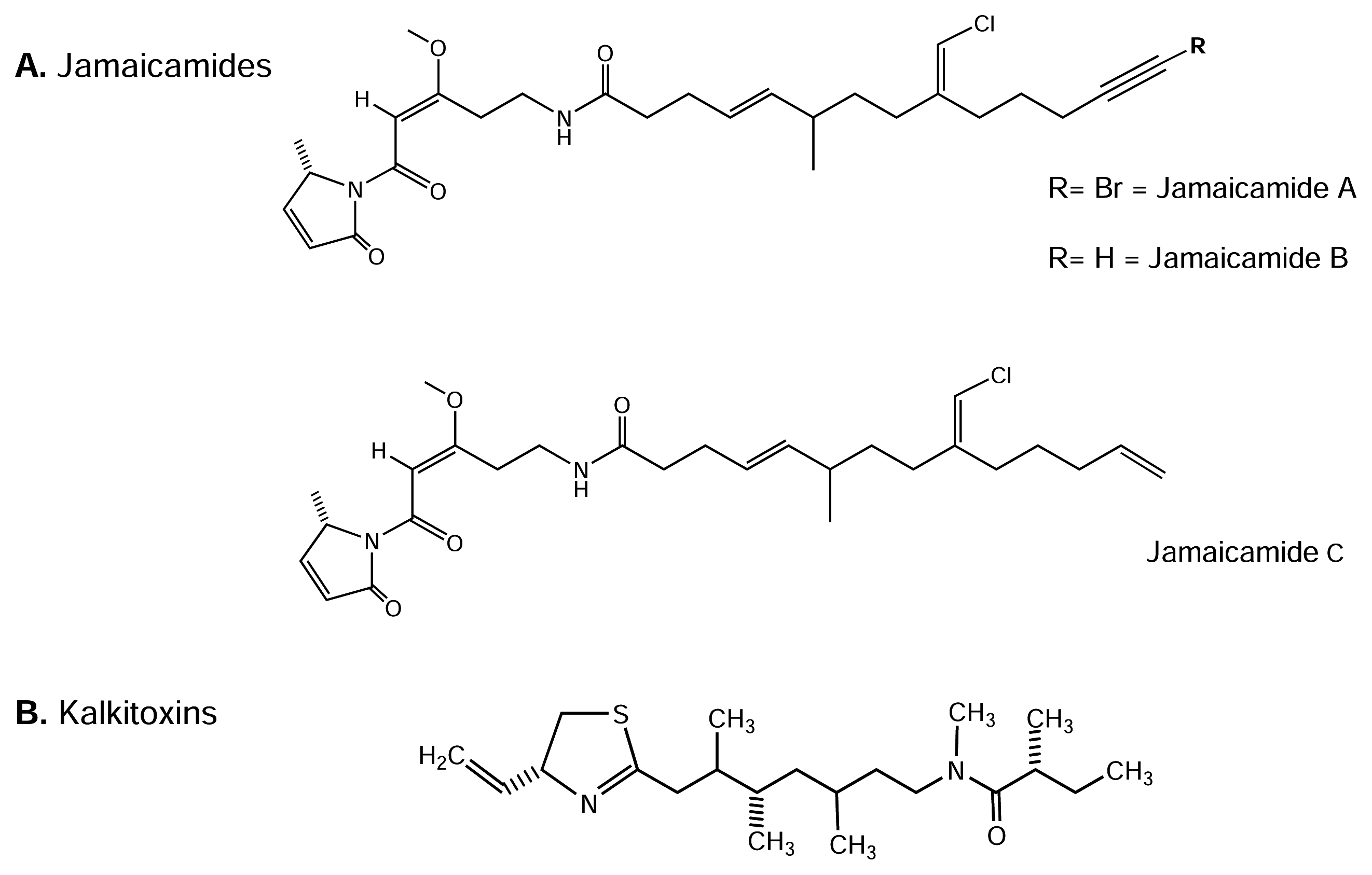{kind=link}
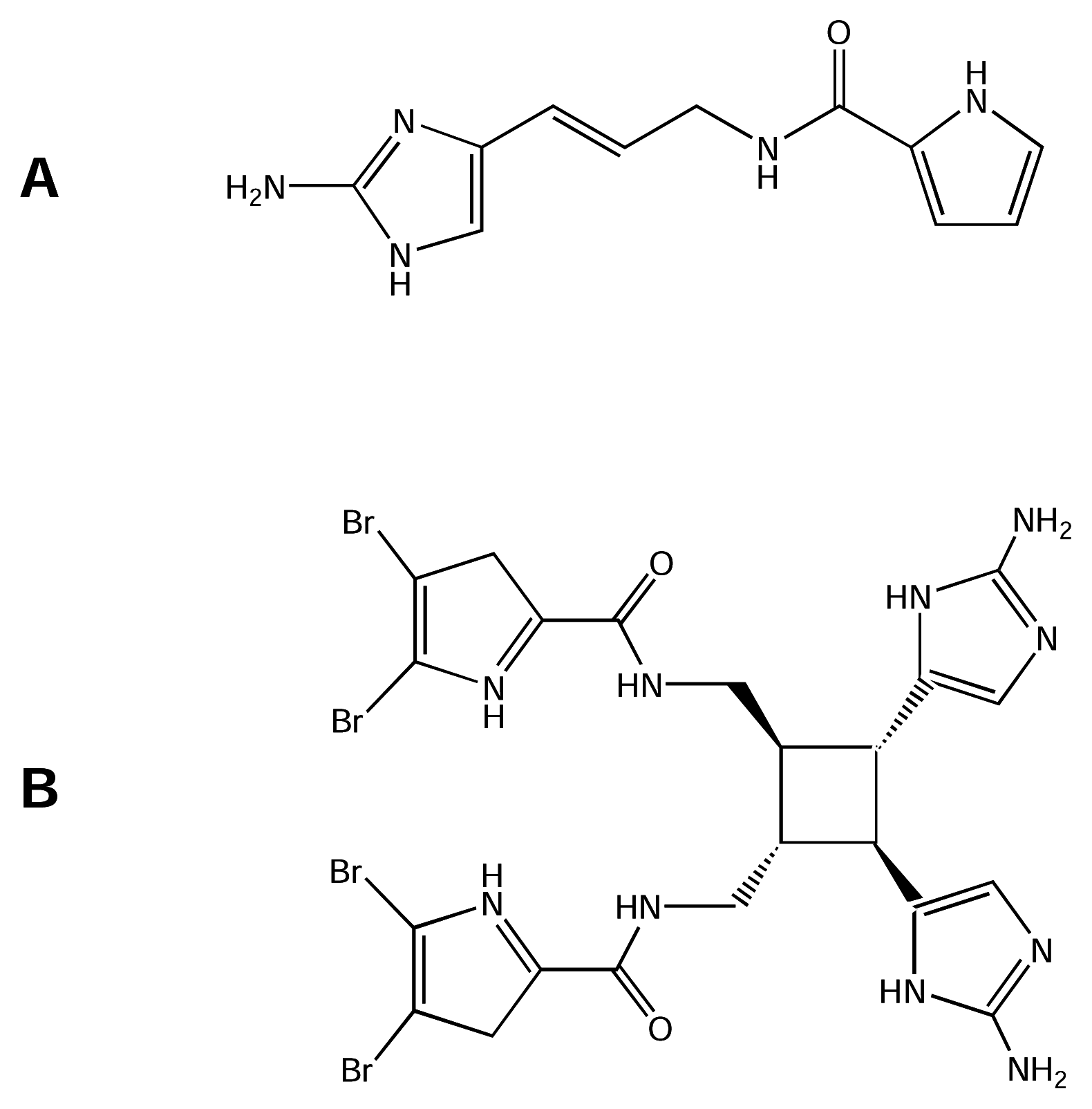{kind=link}
{kind=link}
{kind=link}
| Receptor Site | Physiological Effect | Putative location | Toxin | Organism |
|---|---|---|---|---|
| 1 | Pore blocker | P loop at DI, DII, DIII, DIV | Tetrodotoxins | Puffer fish (Tetraodontiformes), starfish, molluscs, tunicates, dinoflagellates, symbiotic bacteria |
| Saxitoxins | Shellfishes, Red tide dinoflagellates and symbiotic bacteria | |||
| μ-Conotoxins | Cone snails | |||
| 2 | Persistent activation gating; | DI-S6 | Batrachotoxin | [Dendrobatid “poison dart” frogs] |
| Depolarization of resting potentials | DII:S6 | Veratridine | [In the seed of Schoenocaulon officinale and in rhizome of Veratrum album] | |
| Repetitive firings | DIII:S6 | Aconitine | [In the root of Aconitum plants] | |
| DIV:S6 | Grayanotoxin | [In plants of the family Ericaceae;] | ||
| 3 | Prolonged Na+ channel opening | DIV:S3–S4 | α-Scorpion toxins | [Scorpion spp.] |
| DI:S3–S4 Loop | Sea anemone toxins | Sea anemones spp. | ||
| Inactivation gating modifier | DIV:S5–S6 Loop | δ-Atracotoxins | [Funnel-web spider Hadronyche versuta] | |
| 4 | Agonist | ? | Antillatoxin | Dinoflagellate Lyngbya majuscule |
| Shifts in activation gating; repetitive firings | DII:S3–S4 | Loop β-Scorpion toxins | [Scorpion spp,] | |
| 5 | Shift in activation gating | DI:S6 | Brevetoxins | Red tide dinoflagellate Karenia brevis |
| DIV:S5 | Ciguatoxins | Red tide dinoflagellate Gambierdiscus toxicus and tropical coral fishes | ||
| 6 | Prolonged Na+ channel opening | DIV:S3–S4 | δ-Conotoxins | Cone snails |
| 7 | Inhibitor, not a pore blocker | ? | Kalkitoxin | Dinoflagellate Lyngbya majuscule |
| Persistent activation gating; depolarization of resting potentials; repetitive firings | DI-S6 DII-S6 DIII-S6 | DDT and analogs, Pyrethroids | [Insecticide chemicals] | |
| 8 | Inactivation gating modifier | ? | Goniopora | genus Goniopora |
| Prolonged Na+ channel opening | ? | Conus striatus toxin | Conus striatus | |
| 9 | Inhibition of Na+ permeability | DI-S6 DIII-S6 DIV-S6 | Local anesthetics, Anticonvulsants, Antidepressants | [Chemicals from various sources] |
| N.I. | Inhibitor, not a pore blocker | ? | μO-Conotoxins | Cone snails |
| N.I. | Inactivation gating modifier | ? | B-Toxins | Ribbon worm (Cerebratulus lacteus) |
| N.I. | Inhibitor, not a pore blocker | ? | Meroditerpenoids | Brown algae |
| N.I. | Inactivation gating modifier | ? | Clathrodin | Sponge (genus Agelas) |
| N.I. | Inactivation gating modifier | ? | Dibromosceptrin | Sponge (genus Agelas) |
| N.I. | Antagonist | ? | Jamaicamides A-C | Dinoflagellate Lyngbya majuscule |
| N.I. | Inactivation gating modifier | ? | Ostreotoxin-3 | Dinoflagellate Ostreopsis lenticularis |
| μ-Conotoxins | Conus species | # of residues | Nominal net charge (using R, K=1, H=+0.5, D=−1) | Major tissue or channel subtype(s) targeted | References |
|---|---|---|---|---|---|
| GIIIA | C. geographus | 22 | 6 | Rat muscle | [98] |
| GIIIB | C. geographus | 22 | 7 | Rat muscle, rat brain | [98] |
| GIIIC | C. geographus | 22 | 7 | Rat muscle | [33; 98] |
| PIIIA | C. purpurascens | 22 | 6.5 | Rat muscle, rat brain, human brain | [98; 134] |
| SmIIIA | C. stercusmuscarum | 22 | 5.5 | TTx-r (amphibian) skeletal muscle, sympathetic and dorsal root ganglia (DRG) neurons | [19; 73; 156] |
| SIIIA | C. striatus | 20 | 2.5 | TTx-r (amphibian) sympathetic and DRG neurons | [19] |
| KIIIA | C. kinoshitai | 16 | 3.5 | TTx-r (amphibian) sympathetic and DRG neurons | [19] |
| μO-Conotoxins | Conus species | Number of residues | Subtype Targeted | Reference |
|---|---|---|---|---|
| MrVIA | C. marmoreus | 31 | Inhibits TTx-r NaV channels rat dorsal root ganglia, TTx-s NaV channels, molluscan Ca2+ channels | [34; 95] |
| MrVIB | C. marmoreus | 31 | Inhibits TTx-r NaV channels rat dorsal root ganglia, TTx-s NaV channels, molluscan Ca2+ channels | [34; 95] |
| Unclassified Conotoxins | Conus species | Number of residues | Subtype Targeted | Reference |
|---|---|---|---|---|
| TVIIA | C. tulipa | 30 | Not established | [62] |
| GS | C. geographus | 34 | Muscle Na+ blocker receptor site 1 | [163] |
| δ-Conotoxin | Conus species | Natural prey | No. of residues | Subtype Targeted | Reference |
|---|---|---|---|---|---|
| PVIA | C. purpurascens | Fish | 29 | Vertebrate preparations only (including fish, frog and mammalian NaV1.2; NaV1.4 and NaV1.6). | [132; 157] [124; 145] |
| EVIA | C. ermineus | Fish | 32 | Amphibians and mammalian preparations (rNaV1.2, rNaV1.3 and rNaV1.6 but not muscular rNaV1.4 and cardiac hNaV1.5) | [5; 150] |
| SVIE | C. striatus | Fish | 31 | Diverse (Molluscan and Amphibian sympathic neurons) | [18; 157] |
| Am2766 | C. amadis | Molluscs | 26 | Diverse [molluscan and mammalian preparations (rNaV1.2)]. | [140] |
| TxVIA | C. textile | Molluscs | 27 | Molluscan subtype and rat brain neuronal membranes but not toxic to vertebrate system | [45–47; 65; 77] |
| GmVIA | C. gloriamaris | Molluscs | 29 | Diverse (molluscan subtype and mammalian NaV1.2 and NaV1.4 but not NaV1.6-mediated Na+ currents). | [61; 133] |
| NgVIA | C. nigropunctatus | Molluscs | 31 | Diverse (molluscan and more potent to mammalian preparations) | [47] |
Share and Cite
MDPI and ACS Style
Al-Sabi, A.; McArthur, J.; Ostroumov, V.; French, R.J. Marine Toxins That Target Voltage-gated Sodium Channels. Mar. Drugs 2006, 4, 157-192. https://doi.org/10.3390/md403157
AMA Style
Al-Sabi A, McArthur J, Ostroumov V, French RJ. Marine Toxins That Target Voltage-gated Sodium Channels. Marine Drugs. 2006; 4(3):157-192. https://doi.org/10.3390/md403157
Chicago/Turabian StyleAl-Sabi, Ahmed, Jeff McArthur, Vitaly Ostroumov, and Robert J. French. 2006. "Marine Toxins That Target Voltage-gated Sodium Channels" Marine Drugs 4, no. 3: 157-192. https://doi.org/10.3390/md403157