Expression, Purification and Characterization of Chondroitinase AC II from Marine Bacterium Arthrobacter sp. CS01

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
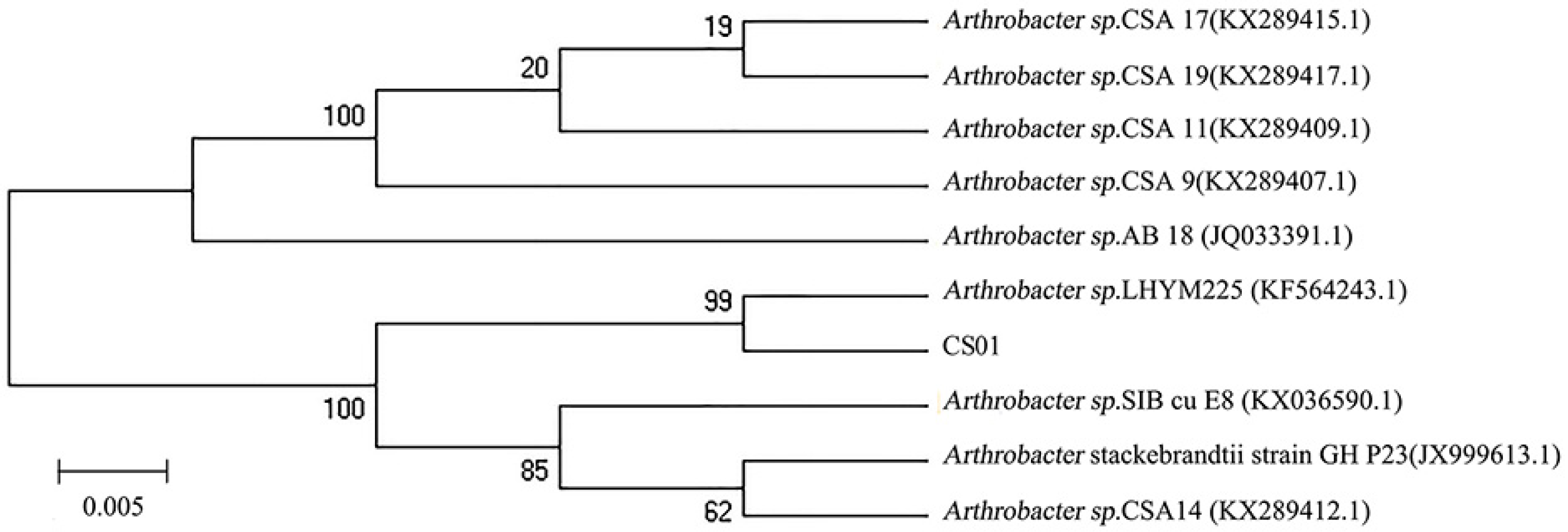
2.1. Isolation and Identification of Strain CS01
2.2. Cloning and Expression of Recombinant ChSase AC II
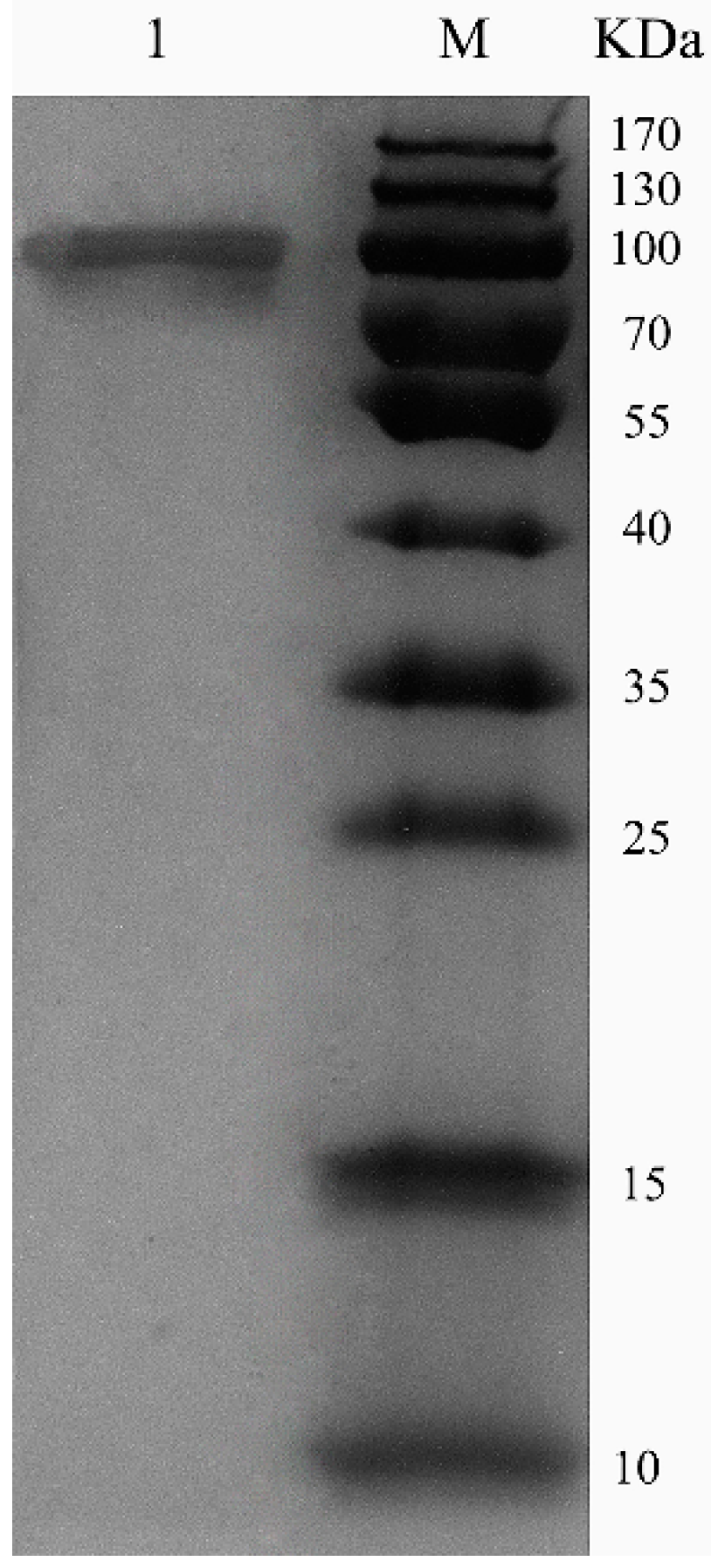
2.3. Purification of Recombinant ChSase AC II
2.4. Substrate Specificity and Kinetic Parameters of Recombinant ChSase AC II
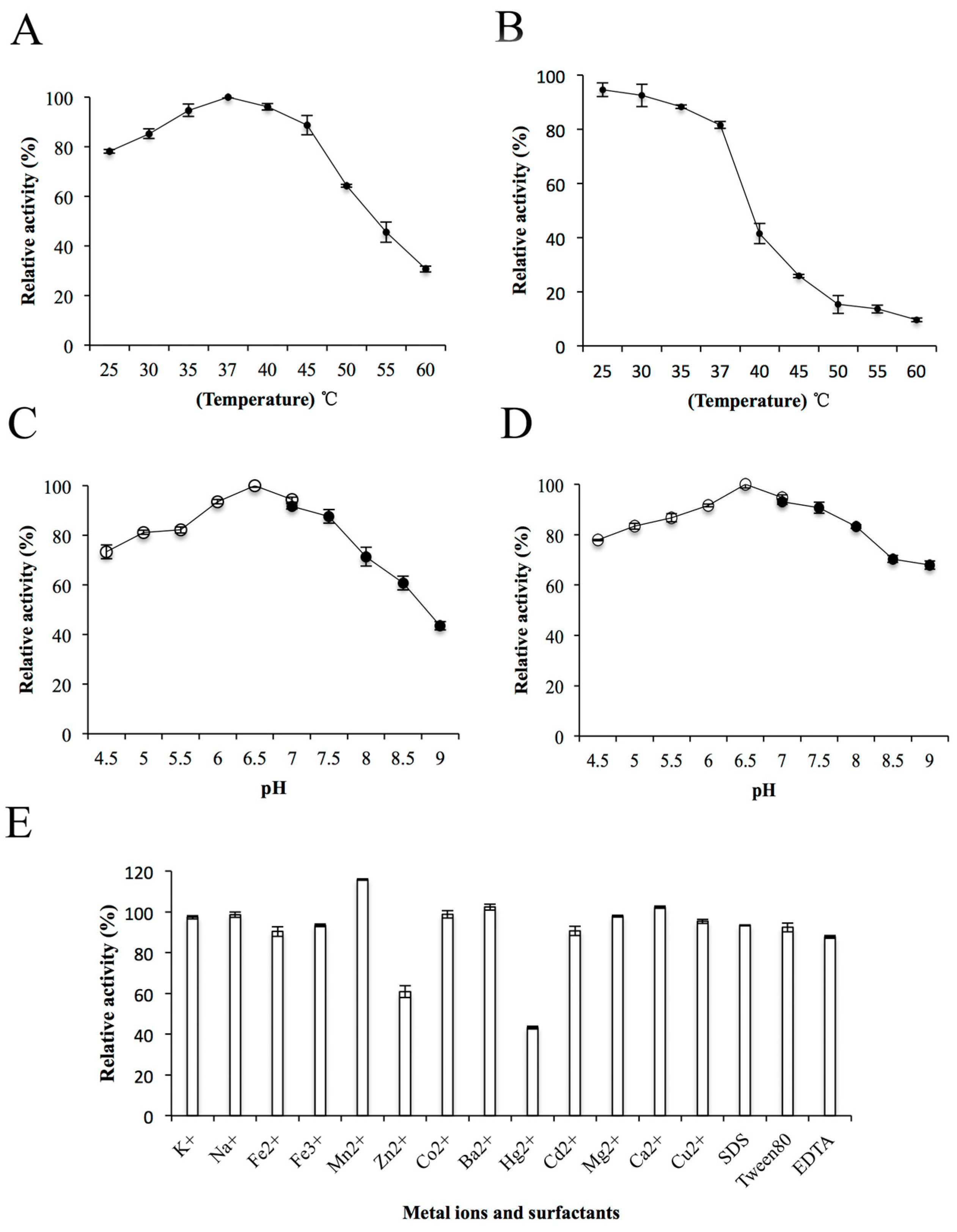
2.5. Biochemical Properties of Recombinant ChSase AC II
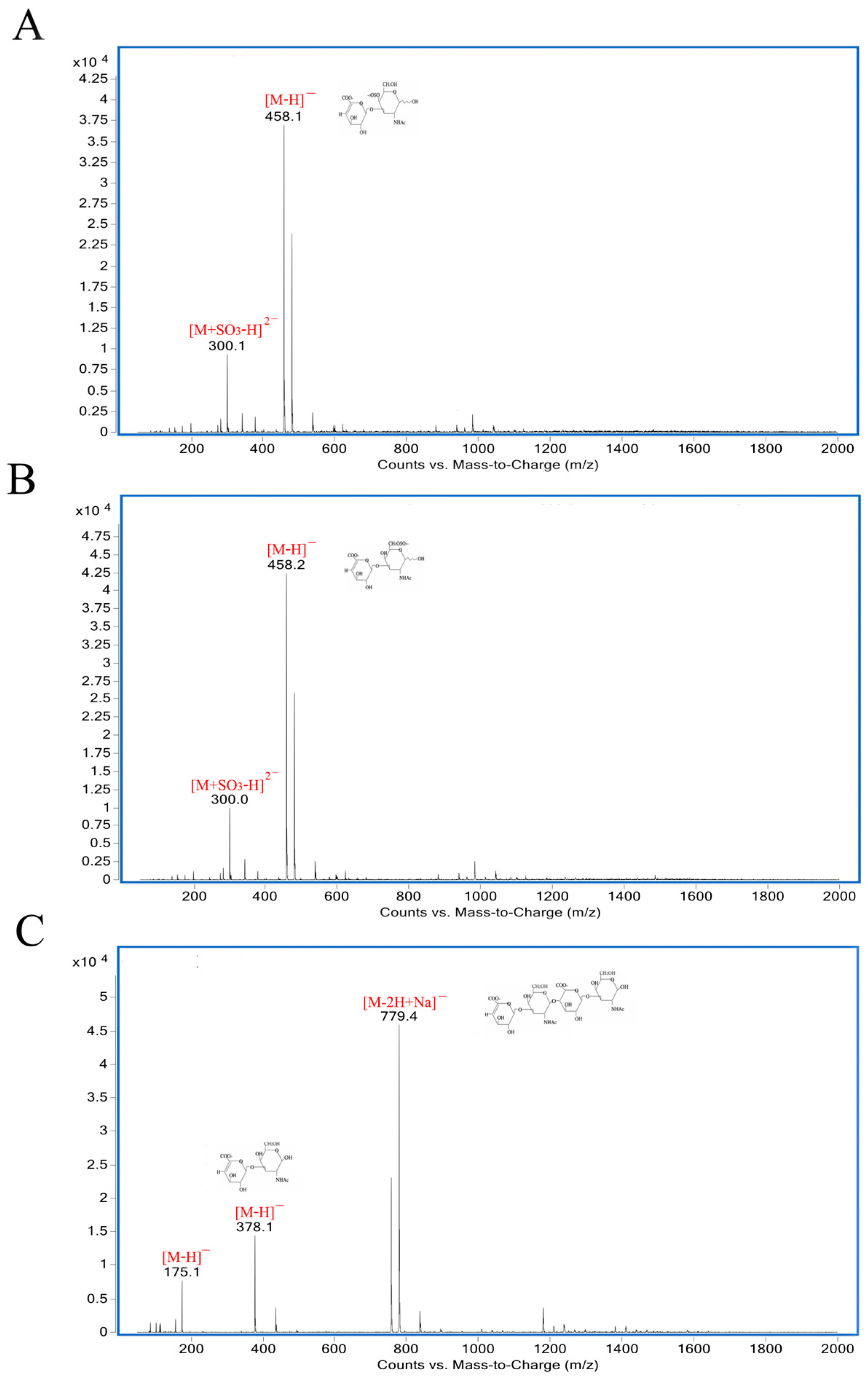
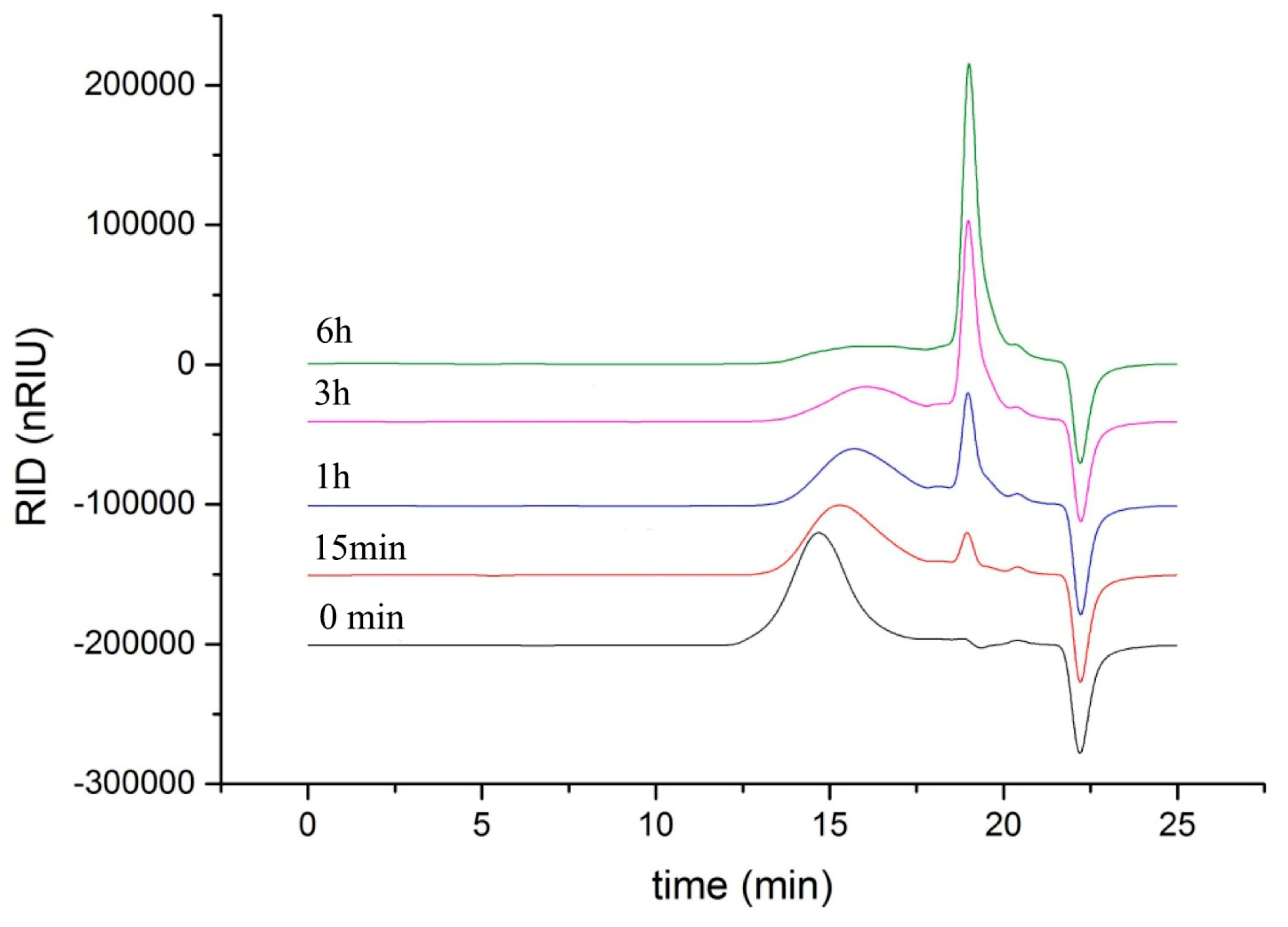
2.6. Analysis of Reaction Mode and Products
3. Materials and Methods
3.1. Strains, Plasmids, Reagents and Medium
3.2. Isolation and Identification of Strain CS01
3.3. Cloning and Expression of Recombinant ChSase AC II
3.4. Purification of Recombinant ChSase AC II
3.5. Assay of Enzyme Activity
3.6. Kinetic Parameters of the Recombinant ChSase AC II
3.7. Biochemical Properties of ChSase AC II
3.8. Analysis of Reaction Mode and Products
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shim, K.; Kim, D. Cloning and expression of chondroitinase AC from Bacteroides stercoris HJ-15. Protein Expr. Purif. 2008, 58, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Song, F.; Xu, N.; Li, D.; Linhardt, R.J.; Zhang, Z. New insights into the action of bacterial chondroitinase AC I and hyaluronidase on hyaluronic acid. Carbohydr. Polym. 2017, 158, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Schiraldi, C.; Cimini, D.; Rosa, M. Production of chondroitin sulfate and chondroitin. Appl. Microbiol. Biotechnol. 2010, 87, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Jiang, Z.; Chang, J.N.; Han, B.; Liu, W.; Peng, Y. Purification, characterization of Chondroitinase ABC from Sphingomonas paucimobilis and in vitro cardiocytoprotection of the enzymatically degraded CS-A. Int. J. Biol. Macromol. 2018, 115, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Monfort, J.R.; Martel-Pelletier, J.; Pelletier, J. Chondroitin sulphate for symptomatic osteoarthritis: Critical appraisal of meta-analyses. Curr. Med. Res. Opin. 2008, 24, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Volpi, N. Quality of different chondroitin sulfate preparations in relation to their therapeutic activity. J. Pharm. Pharmacol. 2009, 61, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Baggio, B.; Gambaro, G.; Marzaro, G.; Marchini, F.; Borsatti, A.; Crepaldi, G. Effects of the oral administration of glycosaminoglycans on cellular abnormalities associated with idiopathic calcium oxalate nephrolithiasis. Eur. J. Clin. Pharmacol. 1991, 40, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Zhang, Q.; Wang, Y.; Lee, B.T.; Betageri, G.V.; Chow, M.S.; Huang, M.; Zuo, Z. Bioavailability enhancement of glucosamine hydrochloride by chitosan. Int. J. Pharm. 2013, 455, 365–373. [Google Scholar] [CrossRef]
- Fraser, P.E.; Darabie, A.A.; McLaurin, J. Amyloid-beta interactions with chondroitin sulfate-derived monosaccharides and disaccharides. implications for drug development. J. Biol. Chem. 2001, 276, 6412–6419. [Google Scholar] [CrossRef]
- Wang, H.; Betti, M. Sulfated glycosaminoglycan-derived oligosaccharides produced from chicken connective tissue promote iron uptake in a human intestinal Caco-2 cell line. Food Chem. 2017, 220, 460–469. [Google Scholar] [CrossRef]
- Xiao, Y.; Li, P.; Cheng, Y.; Zhang, X.; Sheng, J.; Wang, D.; Li, J.J.; Zhang, Q.; Zhong, C.; Cao, R.; et al. Enhancing the intestinal absorption of low molecular weight chondroitin sulfate by conjugation with α-linolenic acid and the transport mechanism of the conjugates. Int. J. Pharm. 2014, 465, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Ariga, T.; Miyatake, T.; Yu, R.K. Role of proteoglycans and glycosaminoglycans in the pathogenesis of Alzheimer’s disease and related disorders: Amyloidogenesis and therapeutic strategies—A review. J. Neurosci. Res. 2010, 88, 2303–2315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liu, C.; Song, C.; Li, P.; Yin, F.; Xiao, Y.; Li, J.; Jiang, W.; Zong, A.; Zhang, X.; et al. Protective effects of low molecular weight chondroitin sulfate on amyloid beta (Aβ)-induced damage in vitro and in vivo. Neuroscience 2015, 305, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Dudás, B.; Hejna, M.J.; Cornelli, U.; Lee, J.C.; Lorens, S.A.; Mervis, R.F.; Hanin, I.; Fareed, J. The blood-brain barrier accessibility of a heparin-derived oligosaccharides C3. Thromb. Res. 2002, 105, 447–453. [Google Scholar] [CrossRef]
- Linhardt, R.J.; Avci, F.Y.; Toida, T.; Kim, Y.S.; Cygler, M. Cs lyases: Structure, activity, and applications in analysis and the treatment of diseases. Adv. Pharmacol. 2006, 53, 187–215. [Google Scholar] [PubMed]
- Jandik, K.A.; Gu, K.; Linhardt, R.J. Action pattern of polysaccharide lyases on glycosaminoglycans. Glycobiology 1994, 4, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Kunttu, H.M.; Jokinen, E.I.; Valtonen, E.T.; Sundberg, L. Virulent and nonvirulent Flavobacterium columnare colony morphologies: Characterization of chondroitin AC lyase activity and adhesion to polystyrene. J. Appl. Microbiol. 2011, 111, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; He, W.; Cress, B.F.; Liu, X.; Alexandria, J.; Yoshizawa, H.; Nishimura, K.; Toida, T.; Koffas, M.A.; Linhardt, R.J. Cloning and Expression of Recombinant Chondroitinase AC II and Its Comparison to the Arthrobacter aurescens Enzyme. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Du, Y.; Yang, F.; Wang, X.; Wang, Y.; Li, H.; Li, R.; Tang, W.; Li, X. Identification of an active-site residue in invertase SUC2 by mass spectrometry-based proteomics and site-directed mutagenesis. Int. J. Mass Spectrom. 2016, 409, 9–15. [Google Scholar] [CrossRef]
- Linhardt, R.J.; Laliberte, M.; Gu, K.; Zimmermann, J. Purification, characterization and specificity of chondroitin lyases and glycuronidase from Flavobacterium heparinum. Biochem. J. 1995, 312, 569–577. [Google Scholar]
- Shaya, D.; Hahn, B.; Park, N.Y.; Sim, J.; Kim, Y.S.; Cygler, M. Characterization of chondroitin sulfate lyase ABC from Bacteroides thetaiotaomicron WAL2926. Biochemistry 2008, 47, 6650–6661. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Kim, B.; Shin, H.M.; Kim, W.; Lee, K.; Kim, Y.; Kim, D. Purification and characterization of novel chondroitin ABC and AC lyases from Bacteroides stercoris HJ-15, a human intestinal anaerobic bacterium. Eur. J. Biochem. 2002, 269, 2934–2940. [Google Scholar] [CrossRef] [PubMed]
- Linn, S.; Chan, T.; Lipeski, L.E.; Salyers, A.A. Isolation and characterization of two chondroitin lyases from Bacteroides thetaiotaomicron. J. Bacteriol. 1983, 156, 859–866. [Google Scholar] [PubMed]
- Zhu, C.; Zhang, J.; Zhang, J.; Jiang, Y.; Shen, Z.; Guan, H.; Jiang, X. Purification and characterization of chondroitinase ABC from Acinetobacter sp. C26. Int. J. Biol. Macromol. 2017, 95, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Daneshjou, S.; Khodaverdian, S.; Dabirmanesh, B.; Rahimi, F.; Daneshjoo, S.; Ghazi, F.; Khajeh, K. Improvement of chondroitinases ABC I stability in natural deep eutectic solvents. J. Mol. Liq. 2017, 227, 21–25. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Total | Total | Specific Activity | Fold | Yield |
|---|---|---|---|---|---|
| Protein (mg) | Activity (U) | (U/mg Protein) | Purification | (%) | |
| Fermentation media | 889.9 | 2723 | 3.06 | 1.0 | 100.0 |
| Ultrafiltration | 480.2 | 1969 | 4.10 | 1.34 | 72.3 |
| HiTrap QFF column | 77.8 | 1454 | 18.7 | 6.11 | 53.4 |
| Substrate | Relative Activity | Vmax | Km | Kcat | Kcat/Km |
|---|---|---|---|---|---|
| (%) | (µM/min) | (µM) | (min−1) | (min−1/µM) | |
| CS-A | 115.64 | 186 | 2.44 | 232 | 95.1 |
| CS-C | 100 | 168 | 1.68 | 210 | 125 |
| HA | 294.93 | 472 | 0.165 | 590 | 3575 |
| Media | Composition |
|---|---|
| Selective | 0.5% CS, 0.5% yeast extract (YE), 0.5% NaCl, 0.2% MgSO4, 0.05% KH2PO4, pH 7.0 |
| Luria-Bertani (LB) | 0.5% YE, 1% NaCl, 1% peptone, pH 7.0 |
| Low Salt LB (LLB) | 0.5% YE, 0.5% NaCl, 1% peptone, pH 7.0 |
| Yeast extract peptone dextrose medium (YPD) | 1% YE, 2% peptone, 2% glucose, pH 7.0 |
| YPD+sorbitol (YPDS) | 1% YE, 2% peptone, 2% glucose, 1 M sorbitol, pH 7.0 |
| Minimal dextrase medium (MD) | 2% glucose, 1.34% yeast nitrogen base without amino acids and ammonium sulfate (YNB), 4 × 10−5% biotin, pH 7.0 |
| Buffered glycerol-complex medium (BMGY) | 1% Glycerol, 1% YE, 2% peptone, 0.34% YNB, 1% (NH4)2SO4 (w/v) and 4 × 10−5% biotin in pH 6.0 100 mM potassium phosphate buffer |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Y.; Yang, S.; Fu, X.; Xie, W.; Li, L.; Liu, Z.; Mou, H.; Zhu, C. Expression, Purification and Characterization of Chondroitinase AC II from Marine Bacterium Arthrobacter sp. CS01. Mar. Drugs 2019, 17, 185. https://doi.org/10.3390/md17030185
Fang Y, Yang S, Fu X, Xie W, Li L, Liu Z, Mou H, Zhu C. Expression, Purification and Characterization of Chondroitinase AC II from Marine Bacterium Arthrobacter sp. CS01. Marine Drugs. 2019; 17(3):185. https://doi.org/10.3390/md17030185
Chicago/Turabian StyleFang, Yangtao, Suxiao Yang, Xiaodan Fu, Wancui Xie, Li Li, Zhemin Liu, Haijin Mou, and Changliang Zhu. 2019. "Expression, Purification and Characterization of Chondroitinase AC II from Marine Bacterium Arthrobacter sp. CS01" Marine Drugs 17, no. 3: 185. https://doi.org/10.3390/md17030185