In Vitro Antithrombotic Properties of Salmon (Salmo salar) Phospholipids in a Novel Food-Grade Extract

 ,
,  ,
,  , and
, and 
Abstract
:
1. Introduction
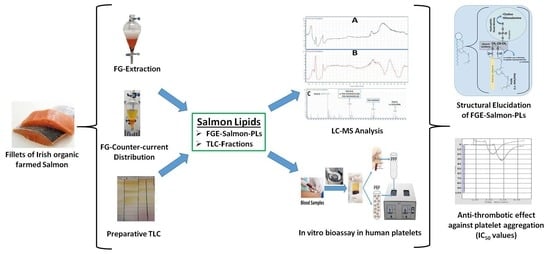
2. Results
2.1. Yield of FGE-salmon-PLs
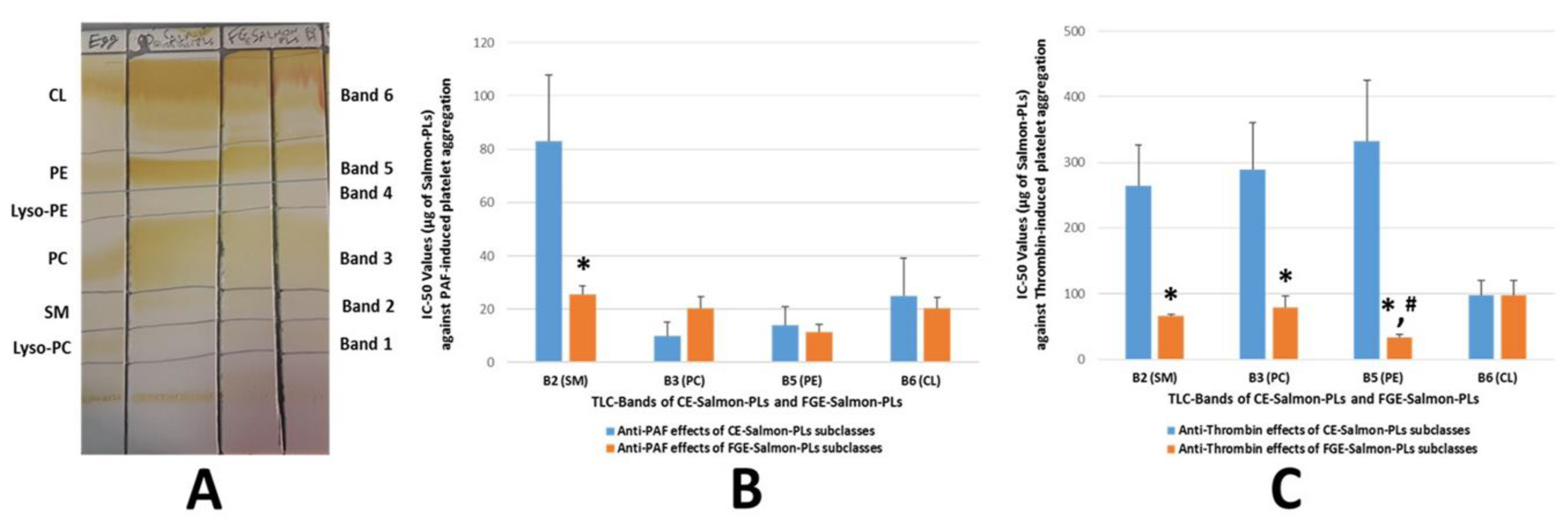
2.2. TLC Analysis of FGE-Salmon-PLs
2.3. Antithrombotic Effects of FGE-Salmon-PLs and Their Lipid Subclasses against Human Platelet Aggregation
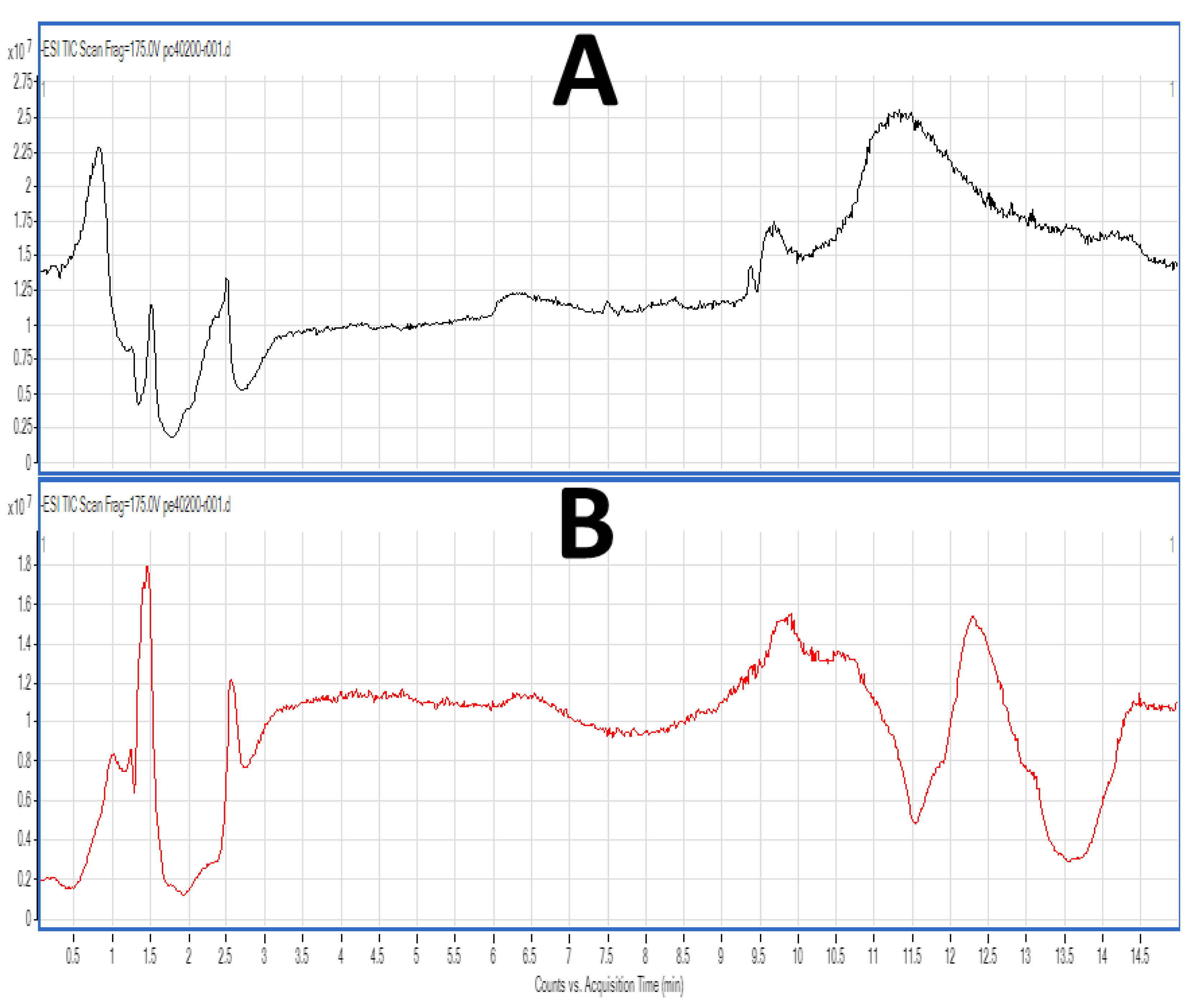
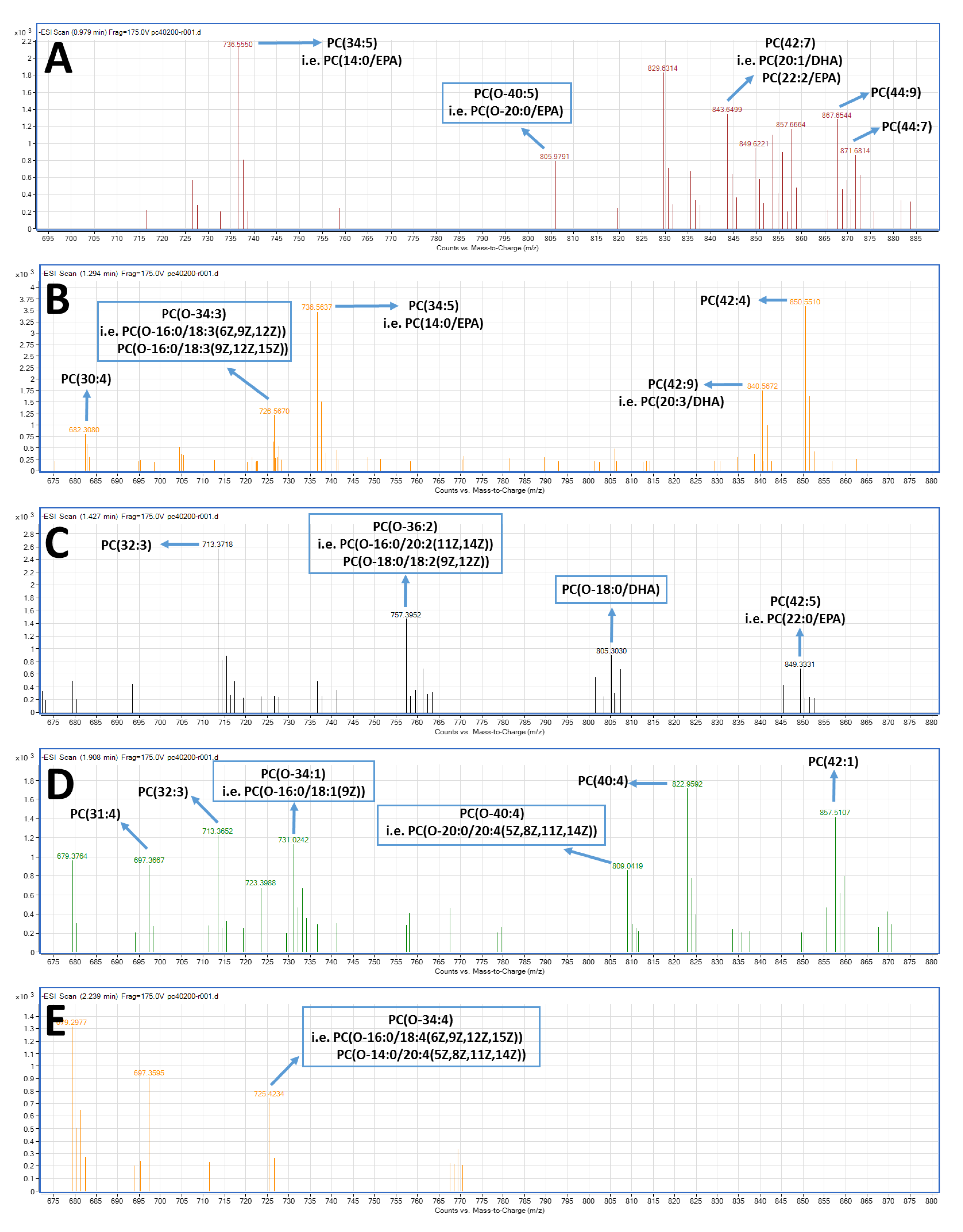
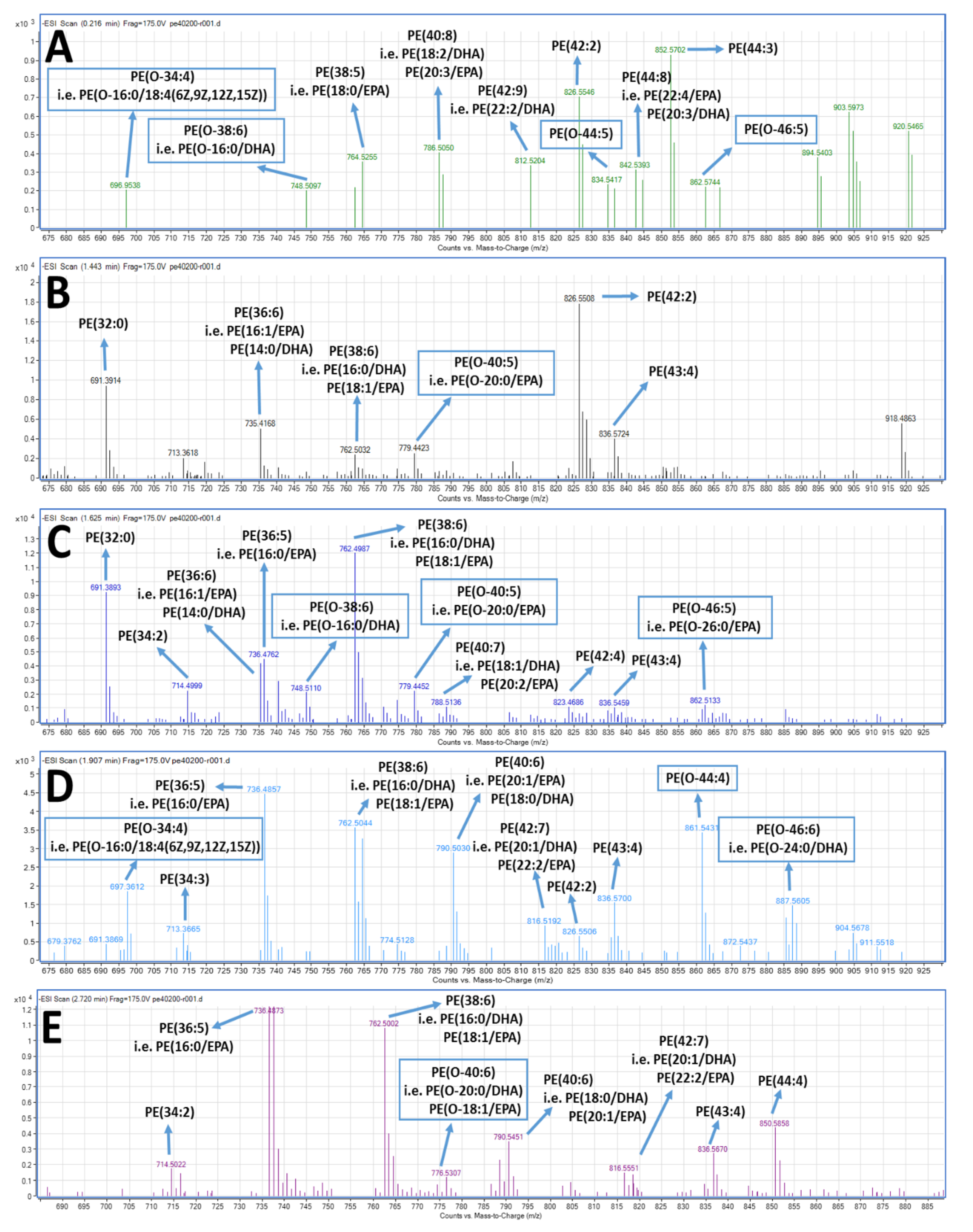
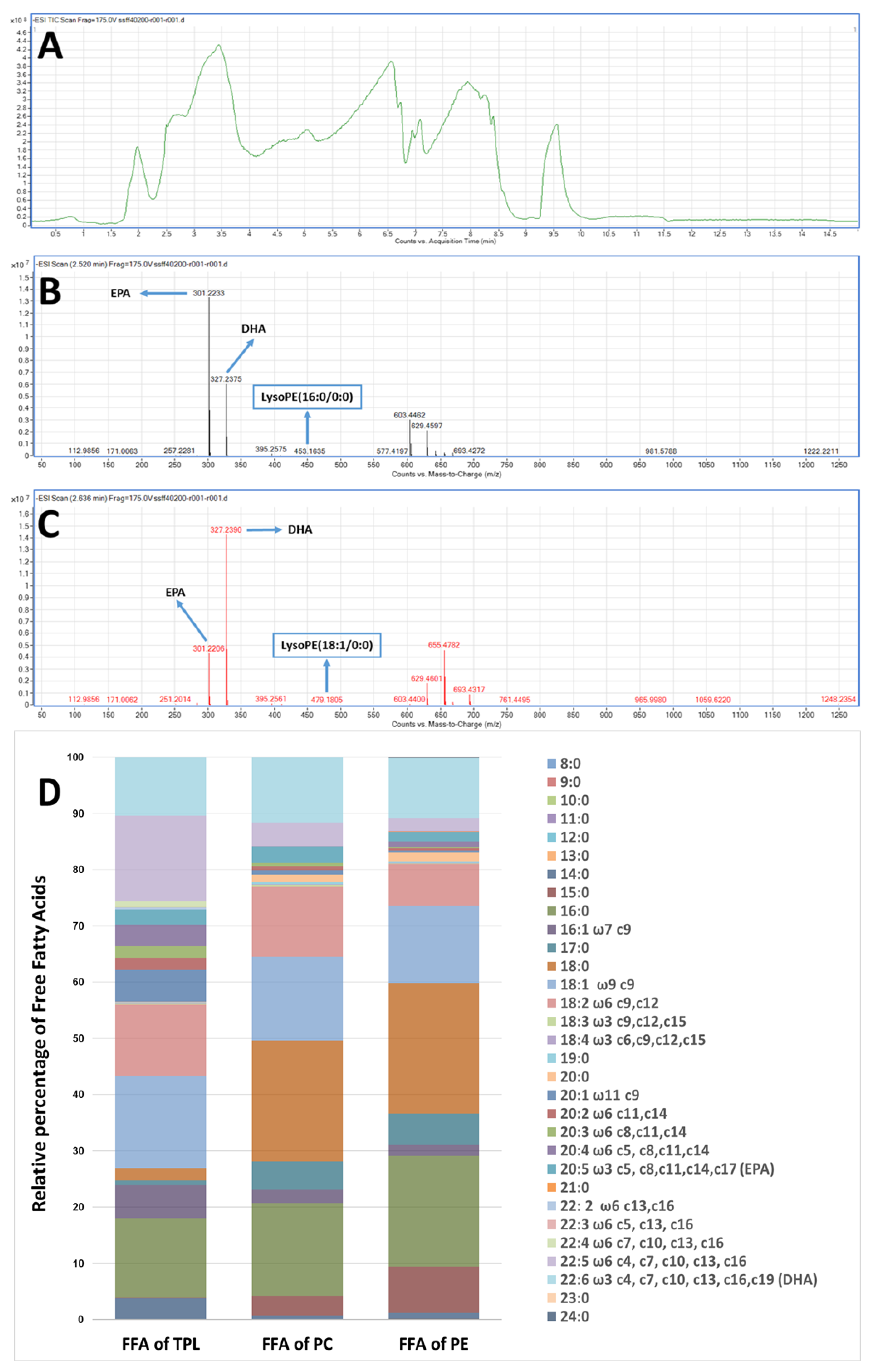
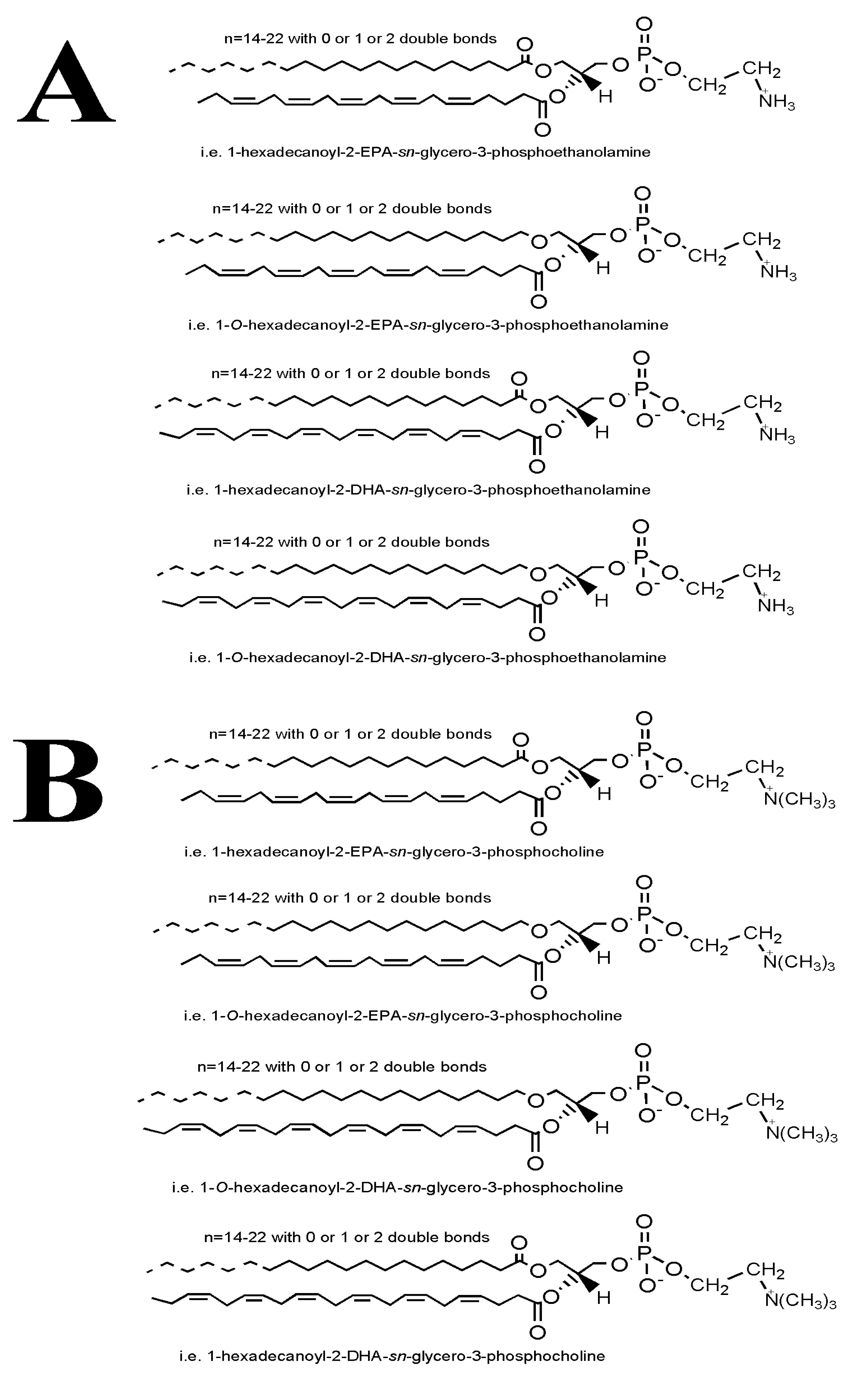
2.4. LC-MS Analysis of FGE-salmon-PLs and Structure–Activity Relationships
3. Discussion
4. Materials and Methods
4.1. Materials and Instrumentation
4.2. Salmon Samples Assessed
4.3. Isolation of FGE-Salmon-PLs from Salmon Fillets
4.4. Fractionation of FGE-Salmon-PLs to Subclasses by Preparative TLC
4.5. Human Platelet-Rich Plasma (hPRP) Aggregation Studies of FGE-Salmon-PLs
4.6. LC-MS Analysis of FGE-Salmon-PLs
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Küllenberg, D.; Taylor, L.A.; Schneider, M.; Massing, U. Health effects of dietary phospholipids. Lipids Health Dis. 2012, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Burri, L.; Hoem, N.; Banni, S.; Berge, K. Marine omega-3 phospholipids: Metabolism and biological activities. Int. J. Mol. Sci. 2012, 13, 15401–15419. [Google Scholar] [CrossRef] [PubMed]
- Murru, E.; Banni, S.; Carta, G. Nutritional properties of dietary omega-3-enriched phospholipids. BioMed Res. Int. 2013, 2013, 965417. [Google Scholar] [CrossRef]
- Lordan, R.; Tsoupras, A.; Zabetakis, I. Phospholipids of animal and marine origin: Structure, function, and anti-inflammatory properties. Molecules 2017, 22, 1964. [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation, not cholesterol, is a cause of chronic disease. Nutrients 2018, 10, 604. [Google Scholar] [CrossRef]
- Demopoulos, C.A.; Karantonis, H.C.; Antonopoulou, S. Platelet-activating factor—A molecular link between atherosclerosis theories. Eur. J. Lipid Sci. Technol. 2003, 105, 705–716. [Google Scholar] [CrossRef]
- Li, Z.; Delaney, M.K.; O’Brien, K.A.; Du, X. Signaling during platelet adhesion and activation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2341–2349. [Google Scholar] [CrossRef] [PubMed]
- Tsopanoglou, N.E.; Maragoudakis, M.E. Thrombin’s central role in angiogenesis and pathophysiological processes. Eur. Cytokine Netw. 2009, 20, 171–179. [Google Scholar] [PubMed]
- Keating, F.K.; Schneider, D.J. The influence of platelet activating factor on the effects of platelet agonists and antiplatelet agents in vitro. J. Thromb. Thrombolysis 2009, 28, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Melnikova, V.O.; Villares, G.J.; Bar-Eli, M. Emerging roles of PAR-1 and PAFR in melanoma metastasis. Cancer Microenviron. 2008, 1, 103–111. [Google Scholar] [CrossRef] [PubMed]
- White, M.; McHowat, J. The therapeutic potential of phospholipase A2 inhibitors in cardiovascular disease. Cardiovasc. Hematol. Agents Med. Chem. 2007, 5, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, G.A.; Mcintyre, T.M.; Prescott, S.M. Production of platelet-activating factor by human vascular endothelial cells: Evidence for a requirement for specific agonists and modulation by prostacyclin. Circulation 1985, 72, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Panayiotou, A.; Samartzis, D.; Nomikos, T.; Fragopoulou, E.; Karantonis, H.C.; Demopoulos, C.A.; Zabetakis, I. Lipid fractions with aggregatory and antiaggregatory activity toward platelets in fresh and fried cod (Gadus morhua): Correlation with platelet-activating factor and atherogenesis. J. Agric. Food Chem. 2000, 48, 6372–6379. [Google Scholar] [CrossRef] [PubMed]
- Nasopoulou, C.; Nomikos, T.; Demopoulos, C.; Zabetakis, I. Comparison of antiatherogenic properties of lipids obtained from wild and cultured sea bass (Dicentrarchus labrax) and gilthead sea bream (Sparus aurata). Food Chem. 2007, 100, 560–567. [Google Scholar] [CrossRef]
- Nasopoulou, C.; Psani, E.; Sioriki, E.; Demopoulos, C.A.; Zabetakis, I. Evaluation of sensory and in vitro cardio protective properties of sardine (Sardina pilchardus): The effect of grilling and brining. Food Nutr. Sci. 2013, 4, 940–949. [Google Scholar]
- Tsoupras, A.; Lordan, R.; Demuru, M.; Shiels, K.; Saha, S.K.; Nasopoulou, C.; Zabetakis, I. Structural elucidation of irish organic farmed salmon (Salmo salar) polar lipids with antithrombotic activities. Mar. Drugs 2018, 16, 176. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.B.; Fragopoulou, E.; Nomikos, T.; Iatrou, C.; Antonopoulou, S.; Demopoulos, C.A. Characterization of the de novo biosynthetic enzyme of platelet activating factor, DDT-insensitive cholinephosphotransferase, of human mesangial cells. Mediat. Inflamm. 2007, 2007, 27683. [Google Scholar] [CrossRef]
- Nasopoulou, C.; Tsoupras, A.B.; Karantonis, H.C.; Demopoulos, C.A.; Zabetakis, I. Fish polar lipids retard atherosclerosis in rabbits by down-regulating PAF biosynthesis and up-regulating paf catabolism. Lipids Health Dis. 2011, 10, 213. [Google Scholar] [CrossRef]
- Nasopoulou, C.; Karantonis, H.C.; Perrea, D.N.; Theocharis, S.E.; Iliopoulos, D.G.; Demopoulos, C.A.; Zabetakis, I. In vivo anti-atherogenic properties of cultured gilthead sea bream (Sparus aurata) polar lipid extracts in hypercholesterolaemic rabbits. Food Chem. 2010, 120, 831–836. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Galanos, D.S.; Kapoulas, V.M. Isolation of polar lipids from triglyceride mixtures. J. Lipid Res. 1962, 3, 134–136. [Google Scholar]
- Kalogeropoulos, N.; Nomikos, T.; Chiou, A.; Fragopoulou, E.; Antonopoulou, S. Chemical composition of greek avgotaracho prepared from mullet (Mugil cephalus): Nutritional and health benefits. J. Agric. Food Chem. 2008, 56, 5916–5925. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulou, S.; Karantonis, H.C.; Nomikos, T.; Oikonomou, A.; Fragopoulou, E.; Pantazidou, A. Bioactive polar lipids from Chroococcidiopsis sp. (cyanobacteria). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 142, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Lordan, R.; Nasopoulou, C.; Tsoupras, A.; Zabetakis, I. The anti-inflammatory properties of food polar lipids. In Bioactive Molecules in Food; Mérillon, J.M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–34. [Google Scholar]
- Stokes, K.Y.; Granger, D.N. Platelets: A critical link between inflammation and microvascular dysfunction. J. Physiol. 2012, 590, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Knapp, H.R. Dietary fatty acids in human thrombosis and hemostasis. Am. J. Clin. Nutr. 1997, 65, 1687S–1698S. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Beilin, L.J.; Burke, V.; Morris, J.; Ritchie, J. Interactions between dietary fat, fish, and fish oils and their effects on platelet function in men at risk of cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Axelrod, L.; Camuso, J.; Williams, E.; Kleinman, K.; Briones, E.; Schoenfeld, D. Effects of a small quantity of ω-3 fatty acids on cardiovascular risk factors in NIDDM: A randomized, prospective, double-blind, controlled study. Diabetes Care 1994, 17, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Chin, J.P.F.; Dart, A.M. How do fish oils affect vascular function? Clin. Exp. Pharmacol. Physiol. 1995, 22, 71–81. [Google Scholar] [CrossRef] [PubMed]
- von Schacky, C.; Weber, P.C. Metabolism and effects on platelet function of the purified eicosapentaenoic and docosahexaenoic acids in humans. J. Clin. Investig. 1985, 76, 2446–2450. [Google Scholar] [CrossRef]
- Hirai, A.; Terano, T.; Takenaga, M.; Kobayashi, S.; Makuta, H.; Ozawa, A.; Fujita, T.; Tamura, Y.; Kitagawa, H.; Kumagai, A. Effect of supplementation of highly purified eicosapentaenoic acid and docosahexaenoic acid on hemostatic function in healthy subjects. Adv. Prostaglandin Thromboxane Leukot. Res. 1987, 17, 838–845. [Google Scholar]
- Woodman, R.J.; Mori, T.A.; Burke, V.; Puddey, I.B.; Barden, A.; Watts, G.F.; Beilin, L.J. Effects of purified eicosapentaenoic acid and docosahexaenoic acid on platelet, fibrinolytic and vascular function in hypertensive type 2 diabetic patients. Atherosclerosis 2003, 166, 85–93. [Google Scholar] [CrossRef]
- Larson, M.K.; Tormoen, G.W.; Weaver, L.J.; Luepke, K.J.; Patel, I.A.; Hjelmen, C.E.; Ensz, N.M.; McComas, L.S.; McCarty, O.J.T. Exogenous modification of platelet membranes with the omega-3 fatty acids EPA and DHA reduces platelet procoagulant activity and thrombus formation. Am. J. Physiol. Cell Physiol. 2013, 304, C273–C279. [Google Scholar] [CrossRef] [PubMed]
- Malle, E.; Sattler, W.; Prenner, E.; Leis, H.J.; Hermetter, A.; Gries, A.; Kostner, G.M. Effects of dietary fish oil supplementation on platelet aggregability and platelet membrane fluidity in normolipemic subjects with and without high plasma Lp(a) concentrations. Atherosclerosis 1991, 88, 193–201. [Google Scholar] [CrossRef]
- Sperling, R.I.; Robin, J.L.; Kylander, K.A.; Lee, T.H.; Lewis, R.A.; Austen, K.F. The effects of n-3 polyunsaturated fatty acids on the generation of platelet-activating factor-acether by human monocytes. J. Immunol. 1987, 139, 4186–4191. [Google Scholar] [PubMed]
- Enns, J.E.; Yeganeh, A.; Zarychanski, R.; Abou-Setta, A.M.; Friesen, C.; Zahradka, P.; Taylor, C.G. The impact of omega-3 polyunsaturated fatty acid supplementation on the incidence of cardiovascular events and complications in peripheral arterial disease: A systematic review and meta-analysis. BMC Cardiovasc. Disord. 2014, 14, 70. [Google Scholar] [CrossRef] [PubMed]
- Walz, C.P.; Barry, A.R.; Koshman, S.L. Omega-3 polyunsaturated fatty acid supplementation in the prevention of cardiovascular disease. Can. Pharm. J. Revue des Pharmaciens du Can. 2016, 149, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Manson, J.E.; Cook, N.R.; Lee, I.-M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Albert, C.M.; Gordon, D.; Copeland, T.; et al. Marine n−3 fatty acids and prevention of cardiovascular disease and cancer. N. Engl. J. Med. 2018, 380, 23–32. [Google Scholar] [CrossRef]
- Galli, C.; Sirtori, C.R.; Mosconi, C.; Medini, L.; Gianfranceschi, G.; Vaccarino, V.; Scolastico, C. Prolonged retention of doubly labeled phosphatidylcholine in human plasma and erythrocytes after oral administration. Lipids 1992, 27, 1005–1012. [Google Scholar] [CrossRef]
- Ikuo, I.; Katsumi, I.; Michihiro, S. Absorption and transport of base moieties of phosphatidylcholine and phosphatidylethanolamine in rats. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1987, 921, 245–253. [Google Scholar] [CrossRef]
- Grima, E.M.; Medina, A.R.; Giménez, A.G.; Sánchez Pérez, J.A.; Camacho, F.G.; García Sánchez, J.L. Comparison between extraction of lipids and fatty acids from microalgal biomass. J. Am. Oil Chem. Soc. 1994, 71, 955–959. [Google Scholar] [CrossRef]
- Granafei, S.; Liebisch, G.; Palmisano, F.; Carlucci, R.; Lionetti, A.; Longobardi, F.; Bianco, G.; Cataldi, T.R.I. Effect of storage and extraction protocols on the lipid and fatty acid profiles of Dicentrarchus labrax brain. Food Anal. Methods 2017, 10, 4003–4012. [Google Scholar] [CrossRef]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Evaluation of food grade solvents for lipid extraction and impact of storage temperature on fatty acid composition of edible seaweeds Laminaria digitata (phaeophyceae) and Palmaria palmata (rhodophyta). Food Chem. 2016, 208, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lopez, N.; Stubhaug, I.; Ipharraguerre, I.; Rimbach, G.; Menoyo, D. Positional distribution of fatty acids in triacylglycerols and phospholipids from fillets of atlantic salmon (Salmo salar) fed vegetable and fish oil blends. Mar. Drugs 2015, 13, 4255–4269. [Google Scholar] [CrossRef] [PubMed]
- Beppu, F.; Yasuda, K.; Okada, A.; Hirosaki, Y.; Okazaki, M.; Gotoh, N. Comparison of the distribution of unsaturated fatty acids at the sn-2 position of phospholipids and triacylglycerols in marine fishes and mammals. J. Oleo Sci. 2017, 66, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Takama, K.; Suzuki, T.; Yoshida, K.; Arai, H.; Anma, H. Lipid content and fatty acid composition of phospholipids in white-flesh fish species. Fish. Sci. 1994, 60, 177–184. [Google Scholar] [CrossRef]
- Le Grandois, J.; Marchioni, E.; Zhao, M.; Giuffrida, F.; Ennahar, S.; Bindler, F. Investigation of natural phosphatidylcholine sources: Separation and identification by liquid chromatography−electrospray ionization−tandem mass spectrometry (LC−ESI−MS2) of molecular species. J. Agric. Food Chem. 2009, 57, 6014–6020. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhao, M.; Ennahar, S.; Bindler, F.; Marchioni, E. Determination of phosphatidylethanolamine molecular species in various food matrices by liquid chromatography–electrospray ionization–tandem mass spectrometry (LC–ESI–MS2). Anal. Bioanal. Chem. 2012, 403, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Andrioli, G.; Carletto, A.; Guarini, P.; Galvani, S.; Biasi, D.; Bellavite, P.; Corrocher, R. Differential effects of dietary supplementation with fish oil or soy lecithin on human platelet adhesion. Thromb. Haemost. 1999, 81, 1522–1527. [Google Scholar]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Warensjö, E.; Jansson, J.-H.; Berglund, L.; Boman, K.; Ahren, B.; Weinehall, L.; Lindahl, B.; Hallmans, G.; Vessby, B. Estimated intake of milk fat is negatively associated with cardiovascular risk factors and does not increase the risk of a first acute myocardial infarction. A prospective case–control study. Br. J. Nutr. 2004, 91, 635–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaw, K.-T.; Friesen, M.D.; Riboli, E.; Luben, R.; Wareham, N. Plasma phospholipid fatty acid concentration and incident coronary heart disease in men and women: The EPIC-Norfolk prospective study. PLoS Med. 2012, 9, e1001255. [Google Scholar] [CrossRef]
- Forouhi, N.G.; Koulman, A.; Sharp, S.J.; Imamura, F.; Kröger, J.; Schulze, M.B.; Crowe, F.L.; Huerta, J.M.; Guevara, M.; Beulens, J.W.J.; et al. Differences in the prospective association between individual plasma phospholipid saturated fatty acids and incident type 2 diabetes: The EPIC-interact case-cohort study. Lancet Diabetes Endocrinol. 2014, 2, 810–818. [Google Scholar] [CrossRef]
- Kurotani, K.; Sato, M.; Yasuda, K.; Kashima, K.; Tanaka, S.; Hayashi, T.; Shirouchi, B.; Akter, S.; Kashino, I.; Hayabuchi, H.; et al. Even- and odd-chain saturated fatty acids in serum phospholipids are differentially associated with adipokines. PLoS ONE 2017, 12, e0178192. [Google Scholar] [CrossRef]
- Zhou, L.; Le Grandois, J.; Marchioni, E.; Zhao, M.; Ennahar, S.; Bindler, F. Improvement of total lipid and glycerophospholipid recoveries from various food matrices using pressurized liquid extraction. J. Agric. Food Chem. 2010, 58, 9912–9917. [Google Scholar] [CrossRef] [PubMed]
- Zonouzi, A.; Auli, M.; Dakheli, M.J.; Hejazi, M. Oil extraction from microalgae Dunalliela sp. By polar and non-polar solvents. Int. J. Biol. Biomol. Agric. Food Biotechnol. Eng. 2016, 10, 634–637. [Google Scholar]
- Castro-Gómez, P.; Rodríguez-Alcalá, L.M.; Monteiro, K.M.; Ruiz, A.L.T.G.; Carvalho, J.E.; Fontecha, J. Antiproliferative activity of buttermilk lipid fractions isolated using food grade and non-food grade solvents on human cancer cell lines. Food Chem. 2016, 212, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Diagne, A.; Fauvel, J.; Record, M.; Chap, H.; Douste-Blazy, L. Studies on ether phospholipids: II. Comparative composition of various tissues from human, rat and guinea pig. Biochim. Biophys. Acta Lipids Lipid Metab. 1984, 793, 221–231. [Google Scholar] [CrossRef]
- Paltauf, F. Ether lipids in biomembranes. Chem. Phys. Lipids 1994, 74, 101–139. [Google Scholar] [CrossRef]
- Chen, S.; Subbaiah, P.V. Phospholipid and fatty acid specificity of endothelial lipase: Potential role of the enzyme in the delivery of docosahexaenoic acid (DHA) to tissues. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2007, 1771, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Gauster, M.; Rechberger, G.; Sovic, A.; Hörl, G.; Steyrer, E.; Sattler, W.; Frank, S. Endothelial lipase releases saturated and unsaturated fatty acids of high density lipoprotein phosphatidylcholine. J. Lipid Res. 2005, 46, 1517–1525. [Google Scholar] [CrossRef] [Green Version]
- Shamburek, R.D.; Zech, L.A.; Cooper, P.S.; Vandenbroek, J.M.; Schwartz, C.C. Disappearance of two major phosphatidylcholines from plasma is predominantly via LCAT and hepatic lipase. Am. J. Phys. 1996, 271, E1073–E1082. [Google Scholar] [CrossRef] [PubMed]
- Scagnelli, G.; Cooper, P.; VandenBroek, J.; Berman, W.; Schwartz, C. Plasma 1-palmitoyl-2-linoleoyl phosphatidylcholine. Evidence for extensive phospholipase A1 hydrolysis and hepatic metabolism of the products. J. Biol. Chem. 1991, 266, 18002–18011. [Google Scholar] [PubMed]
- Tsuda, T.; Yoshimura, H.; Hamasaki, N. Effect of phosphatidylcholine, phosphatidylethanolamine and lysophosphatidylcholine on the activated factor X–prothrombin system. Blood Coagul. Fibrinolysis 2006, 17, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Bakirova, D.R.; Faizullin, D.A.; Valiullina, Y.A.; Salnikov, V.V.; Zuev, Y.F. Effect of lipid surface composition on the formation and structure of fibrin clots. Bull. Exp. Biol. Med. 2017, 163, 722–725. [Google Scholar] [CrossRef] [PubMed]
- Skeaff, C.M.; Holub, B.J. Altered phospholipid composition of plasma membranes from thrombin-stimulated human platelets. Biochim. Biophys. Acta Lipids Lipid Metab. 1985, 834, 164–171. [Google Scholar] [CrossRef]
- Zhixin, L.; Wells, C.W.; North, P.E.; Kumar, S.; Duris, C.B.; McIntyre, J.A.; Ming, Z. Phosphatidylethanolamine at the luminal endothelial surface—Implications for hemostasis and thrombotic autoimmunity. Clin. Appl. Thromb. Hemost. 2009, 17, 158–163. [Google Scholar] [CrossRef]
- Horie, S.; Ishii, H.; Hara, H.; Kazama, M. Enhancement of thrombin-thrombomodulin-catalysed protein c activation by phosphatidylethanolamine containing unsaturated fatty acids: Possible physiological significance of phosphatidylethanolamine in anticoagulant activity of thrombomodulin. Biochem. J. 1994, 301, 683–691. [Google Scholar] [CrossRef]
- Jamin, S.P.; Crabos, M.; Catheline, M.; Martin-Chouly, C.; Legrand, A.B.; Saïag, B. Eicosapentaenoic acid reduces thrombin-evoked release of endothelin-1 in cultured bovine endothelial cells. Res. Commun. Mol. Pathol. Pharmacol. 1999, 105, 271–281. [Google Scholar]
- Ahmed, A.A.; Holub, B.J. Alteration and recovery of bleeding times, platelet aggregation and fatty acid composition of individual phospholipids in platelets of human subjects receiving a supplement of cod-liver oil. Lipids 1984, 19, 617–624. [Google Scholar] [CrossRef]
- van den Besselaar, A. Phosphatidylethanolamine and phosphatidylserine synergistically promote heparin’s anticoagulant effect. Blood Coagul. Fibrinolysis 1995, 6, 239–244. [Google Scholar] [CrossRef]
- Deguchi, H.; Yegneswaran, S.; Griffin, J.H. Sphingolipids as bioactive regulators of thrombin generation. J. Biol. Chem. 2004, 279, 12036–12042. [Google Scholar] [CrossRef] [PubMed]
- Raatz, S.K.; Rosenberger, T.A.; Johnson, L.K.; Wolters, W.W.; Burr, G.S.; Picklo, M.J. Dose-dependent consumption of farmed atlantic salmon (Salmo salar) increases plasma phospholipid n-3 fatty acids differentially. J. Acad. Nutr. Diet. 2013, 113, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Küllenberg de Gaudry, D.; Taylor, L.A.; Kluth, J.; Hübschle, T.; Fritzsche, J.; Hildenbrand, B.; Pletschen, L.; Schilli, K.; Hodina, A.; Griffith, L.S. Effects of marine phospholipids extract on the lipid levels of metastatic and nonmetastatic prostate cancer patients. Int. Sch. Res. Not. 2014, 2014, 249204. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yield & Bioactivity of Salmon-PLs | CE-Salmon-PLs & | FGE-Salmon-PLs |
|---|---|---|
| Yield of PLs $ (g) ± SD | 0.86 ± 0.36 | 0.61 ± 0.21 |
| IC50 † (µg) ± SD against PAF–induced platelet aggregation | 45 ± 22 | 86 ± 18 * |
| IC50 † (µg) ± SD against thrombin-induced platelet aggregation | 382 ± 39 | 102 ± 29 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsoupras, A.; Lordan, R.; Shiels, K.; Saha, S.K.; Nasopoulou, C.; Zabetakis, I. In Vitro Antithrombotic Properties of Salmon (Salmo salar) Phospholipids in a Novel Food-Grade Extract. Mar. Drugs 2019, 17, 62. https://doi.org/10.3390/md17010062
Tsoupras A, Lordan R, Shiels K, Saha SK, Nasopoulou C, Zabetakis I. In Vitro Antithrombotic Properties of Salmon (Salmo salar) Phospholipids in a Novel Food-Grade Extract. Marine Drugs. 2019; 17(1):62. https://doi.org/10.3390/md17010062
Chicago/Turabian StyleTsoupras, Alexandros, Ronan Lordan, Katie Shiels, Sushanta Kumar Saha, Constantina Nasopoulou, and Ioannis Zabetakis. 2019. "In Vitro Antithrombotic Properties of Salmon (Salmo salar) Phospholipids in a Novel Food-Grade Extract" Marine Drugs 17, no. 1: 62. https://doi.org/10.3390/md17010062