Characterization of a Novel PolyM-Preferred Alginate Lyase from Marine Vibrio splendidus OU02
by
 ,
,
Jingjing Zhuang
1, 
Keke Zhang
2,
Xiaohua Liu
2,
Weizhi Liu
2,
Qianqian Lyu
2,* and
Aiguo Ji
1,3,* 1
Marine College, Shandong University, Weihai 264209, China
2
College of Marine Life Sciences, Ocean University of China, Qingdao 266003, China
3
School of Pharmaceutical Sciences, Shandong University, Jinan 250012, China
*
Authors to whom correspondence should be addressed.
Mar. Drugs 2018, 16(9), 295; https://doi.org/10.3390/md16090295
Submission received: 14 July 2018
/
Revised: 9 August 2018
/
Accepted: 21 August 2018
/
Published: 22 August 2018
(This article belongs to the Collection Marine Polysaccharides)
Abstract
:Alginate lyases are enzymes that degrade alginate into oligosaccharides which possess a variety of biological activities. Discovering and characterizing novel alginate lyases has great significance for industrial and medical applications. In this study, we reported a novel alginate lyase, AlyA-OU02, derived from the marine Vibrio splendidus OU02. The BLASTP searches showed that AlyA-OU02 belonged to polysaccharide lyase family 7 (PL7) and contained two consecutive PL7 domains, which was rare among the alginate lyases in PL7 family. Both the two domains, AlyAa and AlyAb, had lyase activities, while AlyAa exhibited polyM preference, and AlyAb was polyG-preferred. In addition, the enzyme activity of AlyAa was much higher than AlyAb at 25 °C. The full-length enzyme of AlyA-OU02 showed polyM preference, which was the same as AlyAa. AlyAa degraded alginate into di-, tri-, and tetra-alginate oligosaccharides, while AlyAb degraded alginate into tri-, tetra-, and penta-alginate oligosaccharides. The degraded products of AlyA-OU02 were similar to AlyAa. Our work provided a potential candidate in the application of alginate oligosaccharide production and the characterization of the two domains might provide insights into the use of alginate of this organism.
1. Introduction
Alginate is a linear polysaccharide consisting of β-D-mannuronate (M) and α-L-guluronate (G) residues, arranged in varying sequences of poly β-D-mannuronate (polyM), poly α-L-guluronate (polyG), and the heteropolymer (polyMG) [1]. It has been widely used in biomedical science and engineering, due to its low toxicity, good biocompatibility, low cost, and mild gelation [2]. Alginate now has attracted increasing attention, since it is the most abundant polysaccharide of brown algae, which is considered as the third-generation biofuel [3]. Moreover, studies have showed that alginate oligosaccharides (AOS) possess a variety of biological activities, such as antioxidant, neuroprotective, antibacterial, and antitumor [4,5,6,7]. Therefore, to research the depolymerization of alginate has great importance.
Alginate lyases are enzymes that degrade alginate into unsaturated oligosaccharides by cleaving glycosidic bonds via a β-elimination mechanism [8]. To date, a number of alginate lyases have been identified from various organisms (e.g., algae, marine mollusks, bacteria, virus, and fungi) [2,8,9] and part of their structural relationships have been elucidated [10,11,12,13]. Alginate lyases are divided into polyM-specific, polyG-specific, and polyMG lyases, according to their substrate specificity [14]. According to the primary structures, alginate lyases are classified into seven polysaccharide lyase (PL) families, namely PL-5,6,7,14,15,17, and 18 [15]. Among the seven PL families, PL7 is the most prevalent as reported in the Carbohydrate-Active enZYmes database (http://www.cazy.org/). Most of the PL7 alginate lyases belong to polyG-specific lyases, and few belong to polyM-specific lyases [12]. The substrate specificity is related with the amino acids in the highly conserved regions [16,17]. Commonly, the polyM-specific, polyG-specific, and polyMG alginate lyases contain QVH, QIH, and QIH in the conserved regions, respectively [17].
In this study, we reported a new polyM-preferred alginate lyase (AlyA-OU02) from the marine Vibrio splendidus OU02 which possessed two consecutive PL7 domains (Figure 1A). We cloned, expressed, and characterized AlyA-OU02 and the two PL7 domains (AlyAa and AlyAb), respectively. Both PL7 domains contained a highly conserved QIH sequence which was thought to indicate ployG specificity. Interestingly, it turned out that AlyA-OU02 was polyM-preferred in accordance with AlyAa. Our work suggested the potential applications in AOS production of AlyA-OU02, and provided insights into the use of alginate of this organism.
2. Results and Discussions
2.1. Sequence Analysis of AlyA-OU02
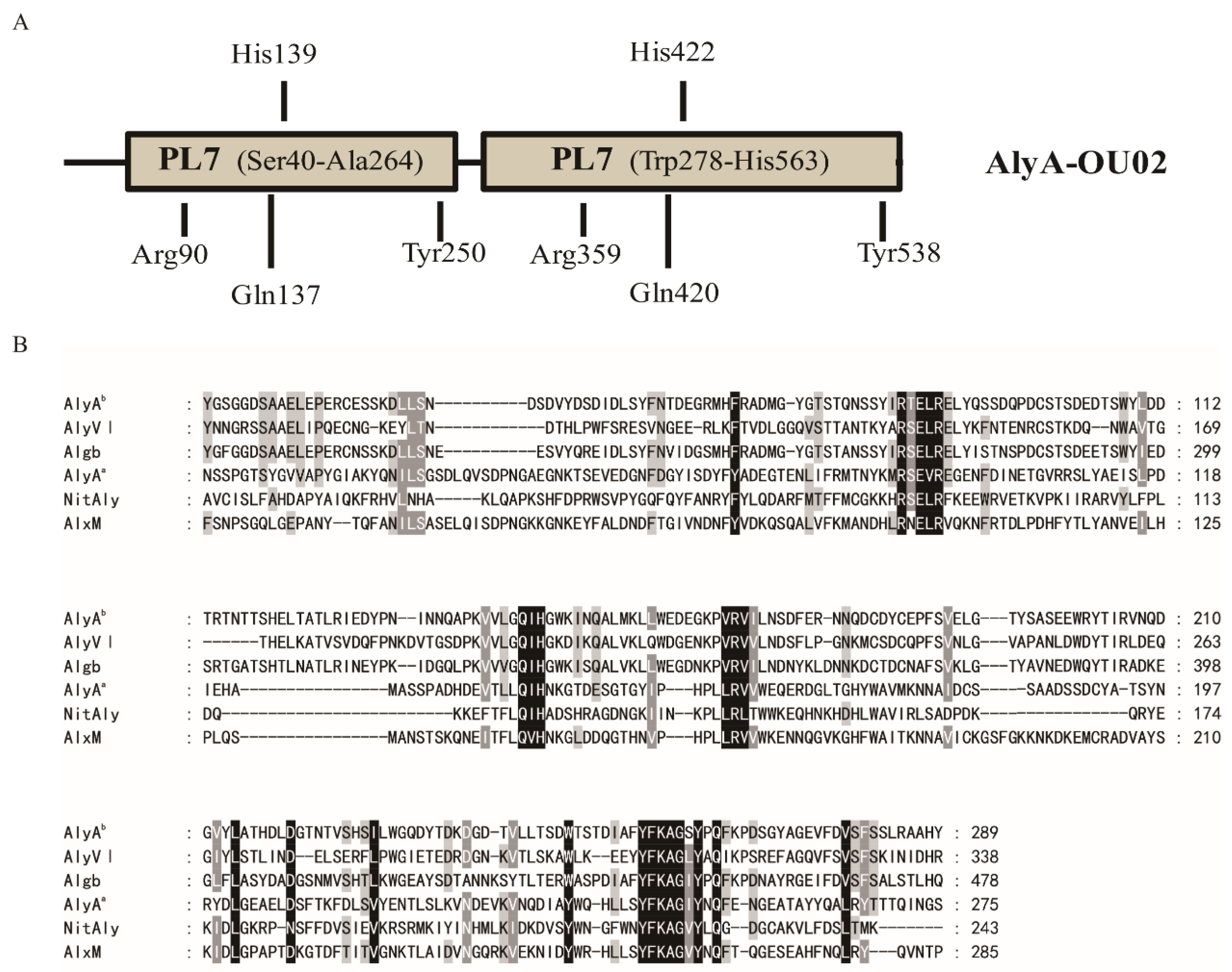
The alginate lyase gene was cloned from marine Vibrio splendidus OU02, and was designated as AlyA-OU02. It consists of 1695 bp encoding 564 amino acids. The calculated molecular mass was ~64 kDa, and the isoelectric point (PI) was 4.27. The BLASTP searches showed that AlyA-OU02 contained two alginate_lyase2 superfamily modules, and belonged to PL7 family (Figure 1A), which was similar to AlyA from Vibrio splendidus 12B01 [18]. Figure 1B showed the multiple sequence alignments of AlyAa, AlyAb, and related alginate lyases of PL7 family [19,20,21,22]. Both domains contained the highly conserved regions R(S/T)E(V/L)R, QIH, and YFKAG(I/S)Y, which were necessary for substrate recognition and catalysis [23]. The putative residues involved in the catalytic reaction were pointed out in Figure 1A.
2.2. Cloning, Expression, and Purification of Different Versions of AlyA-OU02
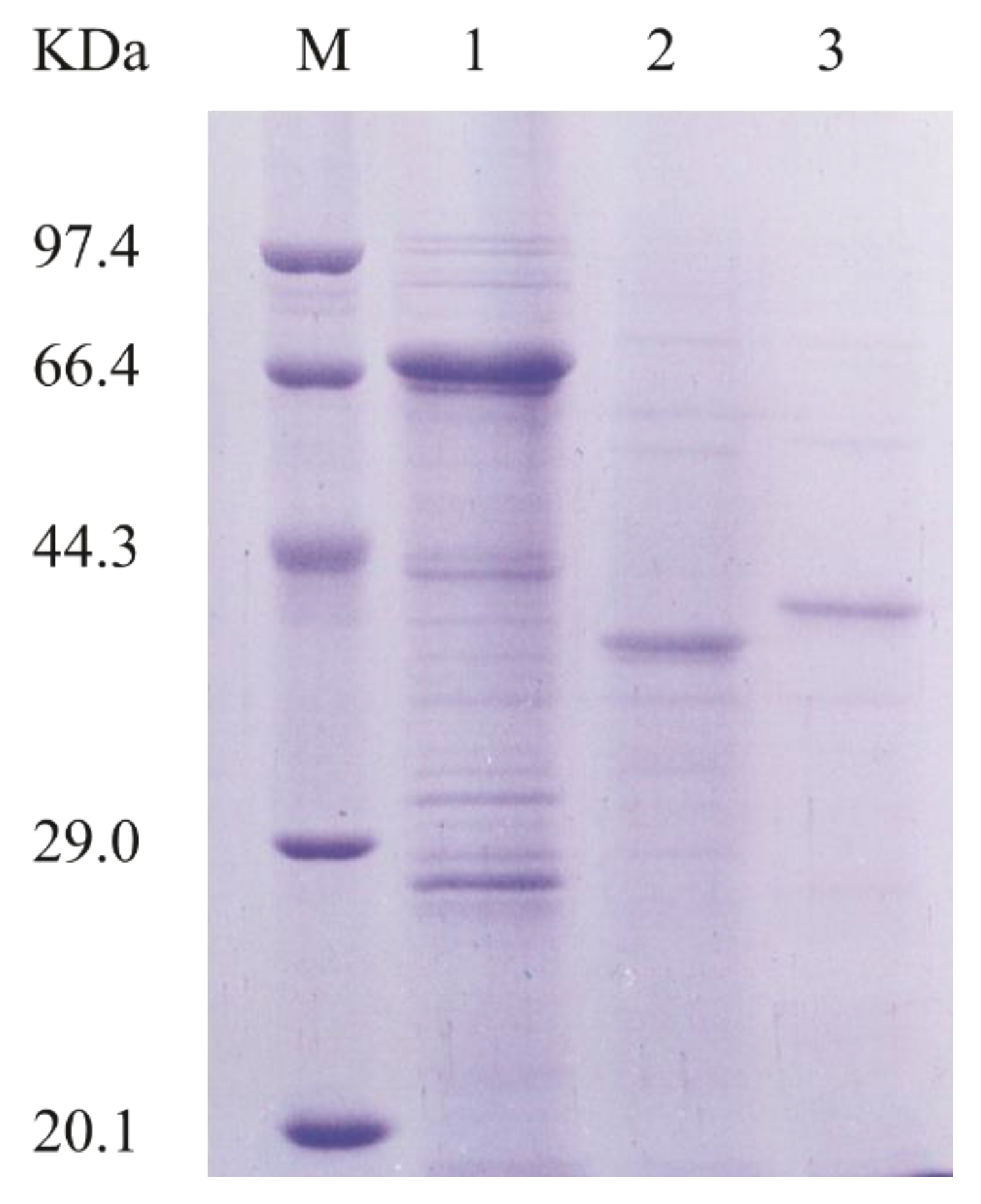
To better characterize AlyA-OU02, we cloned, expressed, and purified AlyA-OU02 and its two domains, respectively. The AlyA-OU02, AlyAa, and AlyAb genes were cloned into pET-32a-PreScission vector (modified by our laboratory) respectively, and expressed in the E. coli BL21 (DE3)-pET-32a-PreScission system. The enzymes were then purified by Ni-nitrilotriacetic acid agarose (Ni-NTA) affinity chromatography and analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Figure 2 showed that the molecular weights of AlyA-OU02, AlyAa, and AlyAb were approximately 65 kDa, 31 kDa, and 33 kDa, which were consistent with the calculated molecular weights.
2.3. Enzymatic Activity Measurements for AlyA-OU02, AlyAa, and AlyAb
Alginate lyases could degrade alginate into unsaturated oligosaccharides with the formation of a double bond between C4 and C5 at the non-reducing terminus. The enzyme activity was measured by monitoring the increased absorbance at 235 nm. Both domains showed alginate lyase activities, and the enzyme activity of AlyAa was much higher than AlyAb at 25 °C (Figure 3). The enzyme activity of AlyA-OU02 was similar to AlyAa. The km values of AlyA-OU02, AlyAa, and AlyAb toward alginate were 3.55, 4.80, and 0.34 mg/mL respectively. The kcat values of AlyA-OU02, AlyAa, and AlyAb toward alginate were 630, 1052, and 42 min−1, respectively. The enzymatic kinetics assay was consistent with the enzyme activity assay.
AlyA from Vibrio splendidus 12B01 also contained two PL7 domains, and the sequence similarity between AlyA-OU02 and AlyA was 89%, however, domain 1 of AlyA had no lyase activity [18]. We speculated the potential reason was that AlyA was purified by renaturation from inclusion bodies, and the length of AlyA (53–286) was different from AlyAa [18].
Crystal structure of FlAlyA showed that His124 and Tyr239 in the highly conserved regions participated in the catalytic reaction [12]. Furthermore, the catalytic activities of the H124N/Y239F mutants were much lower than the wild type. We constructed H139N/Y250F mutants of AlyAa and measured the lyase activities in order to identify the catalytic residues. The purified H139N/Y250F of AlyAa were analyzed by SDS-PAGE (Figure S1). The alginate lyase activities of the mutants were undetectable, suggesting that His139 and Tyr250 were necessary for catalytic reaction of AlyAa (Figure S2).
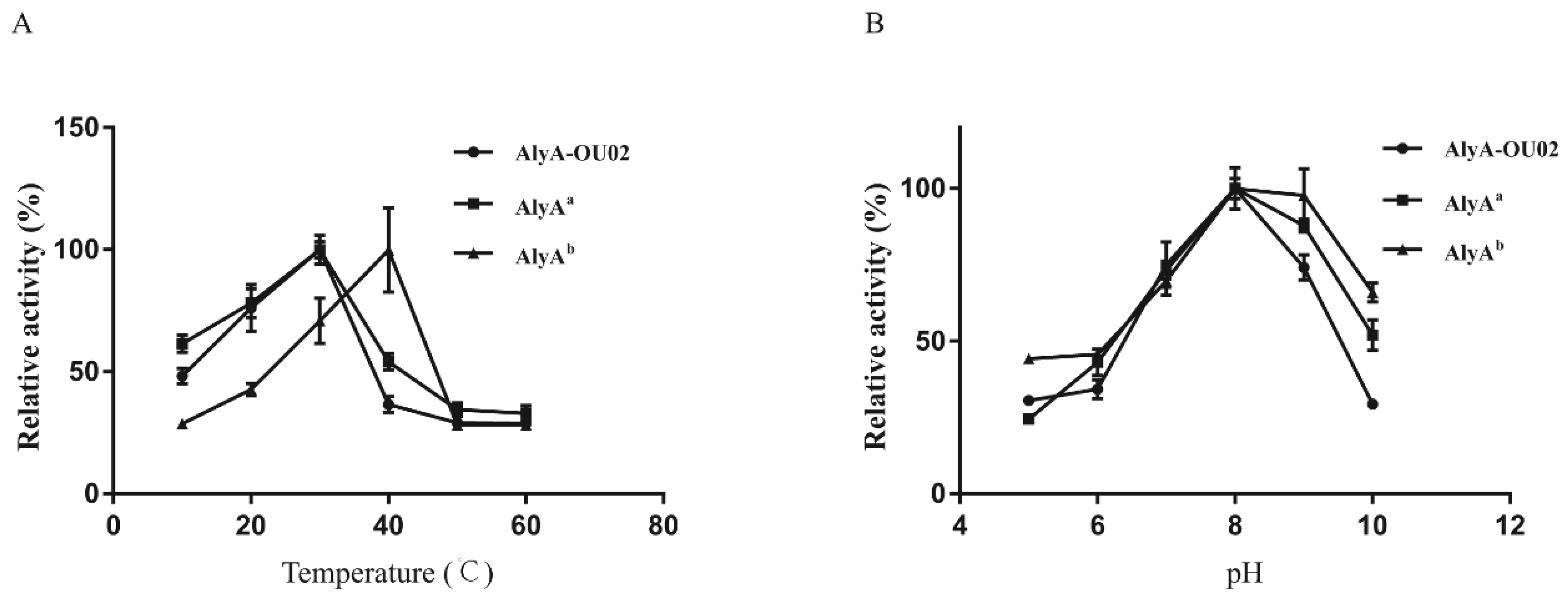
Alginate lyases with different sequences exhibited different optimal temperatures. Most of the PL7 alginate lyases in Table 1 had maximum activity at 30–45 °C. AlyAa had maximum activity at 30 °C, while AlyAb had maximum activity at 40 °C (Figure 4A). The different optimal temperature of the two domains might help the bacteria to better adapt to the changing environment. The alginate lyases in Table 1 had maximum activity in a pH range of 6.0 to 9.0. Both AlyAa and AlyAb exhibited maximum activity at pH 8.0, which corresponded with the range (Figure 4B). The optimal temperature/pH for AlyA-OU02 was 30 °C/8.0, the same as for AlyAa. Moreover, the optimal temperature/pH for Algb [20] and ALG-5 [24] in Table 1 was also 30 °C/8.0. Considering that the ion types in the buffer might impact the enzyme activity, we also investigated the pH effect in sodium phosphate buffer (pH 6.0–8.0) and Tris-HCl buffer (pH 7.0–10.0). Our results showed that sodium phosphate buffer decreased enzyme activities of AlyA-OU02, AlyAa, and AlyAb, compared with Tris-HCl buffer (Figure S3).
To assess the thermostabilities of AlyA-OU02 and its two domains, the enzymes were incubated in Tris-HCl buffer at different temperatures for 1 h. The enzyme activities were assayed using alginate as the substrate. As shown in Figure 5, the activity of AlyAa remained unchanged, while AlyAb retained 40% activity after incubation at 30 °C for 1 h. The stability of AlyA-OU02 decreased with increasing temperature, and retained 50% activity after incubation at 30 °C for 1 h. All the three enzymes were inactive after incubation at 50 °C. Our findings suggested that AlyAa exhibited more thermostability compared to AlyAb and AlyA-OU02.
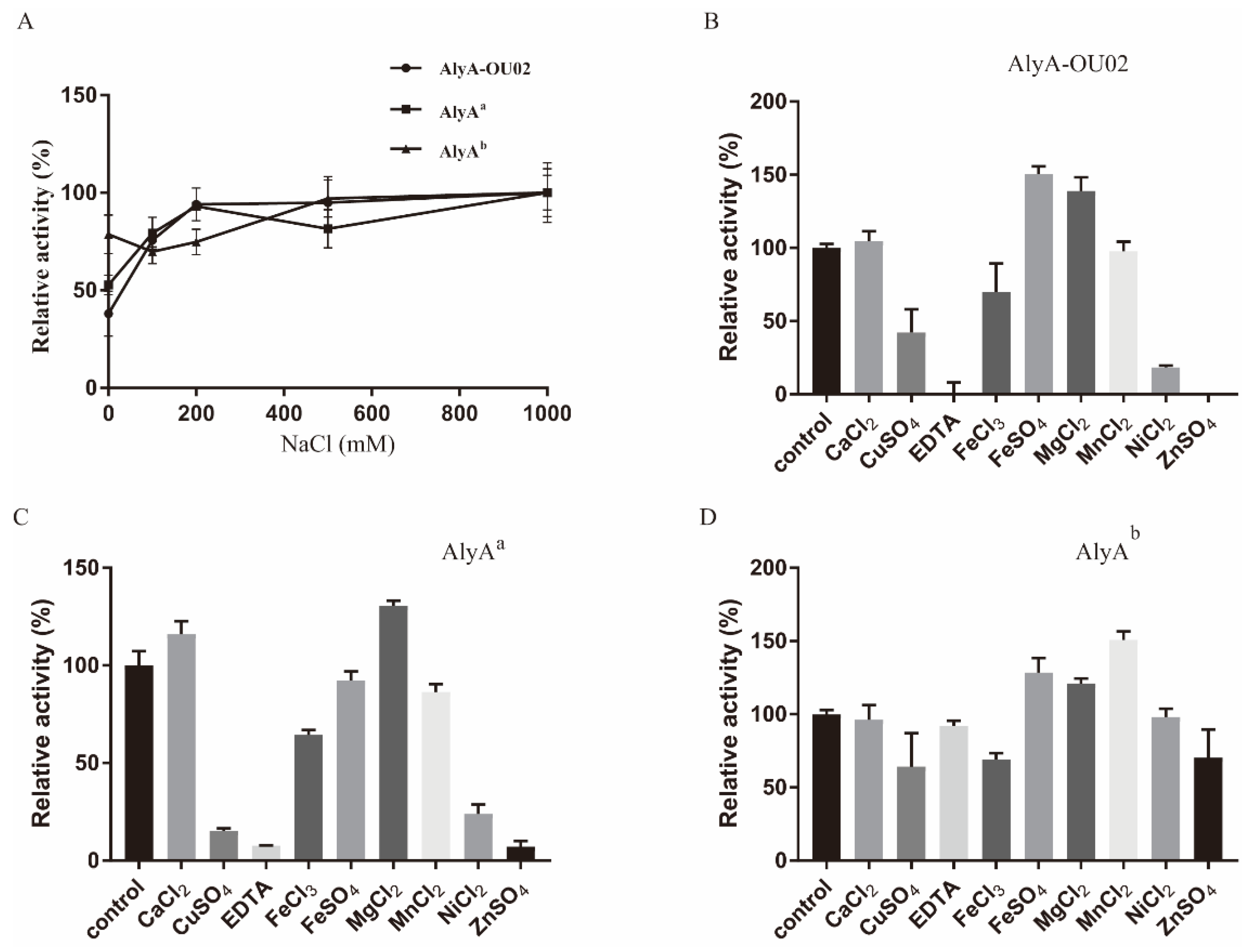
The activities of alginate lyases might be affected by metal ions, since various organisms survived and evolved in various environments. We first examined the effects of NaCl on the enzymatic activities. As shown in Figure 6A, 200 mM NaCl could effectively enhance the activities of AlyA-OU02 and AlyAa, while moderately affecting AlyAb. Certain alginate lyases could be activated by NaCl, but were not resistant to high concentrations of NaCl, such as AlyH1 [36] and AlgA [39]. AlyA-OU02, AlyAa, and AlyAb all exhibited high salt tolerance, which was analogous to AlgM4 from marine bacterium Vibrio weizhoudaoensis M0101 [32]. The effects of chelating agent and other metal ions on AlyA-OU02, AlyAa, and AlyAb activities were studied in the absence of NaCl. As shown in Figure 6B–D, Ca2+ showed little effect on the enzymatic activities. Similarly to rAlgSV1-PL7 [35], AlyA-OU02, AlyAa, and AlyAb could be activated by Mg2+. Besides, enzymatic activities of AlyA-OU02 and AlyAa were inhibited by the chelating agent ethylenediaminetetraacetic acid (EDTA), indicating that AlyA-OU02 and AlyAa might be ion-dependent alginate lyases.
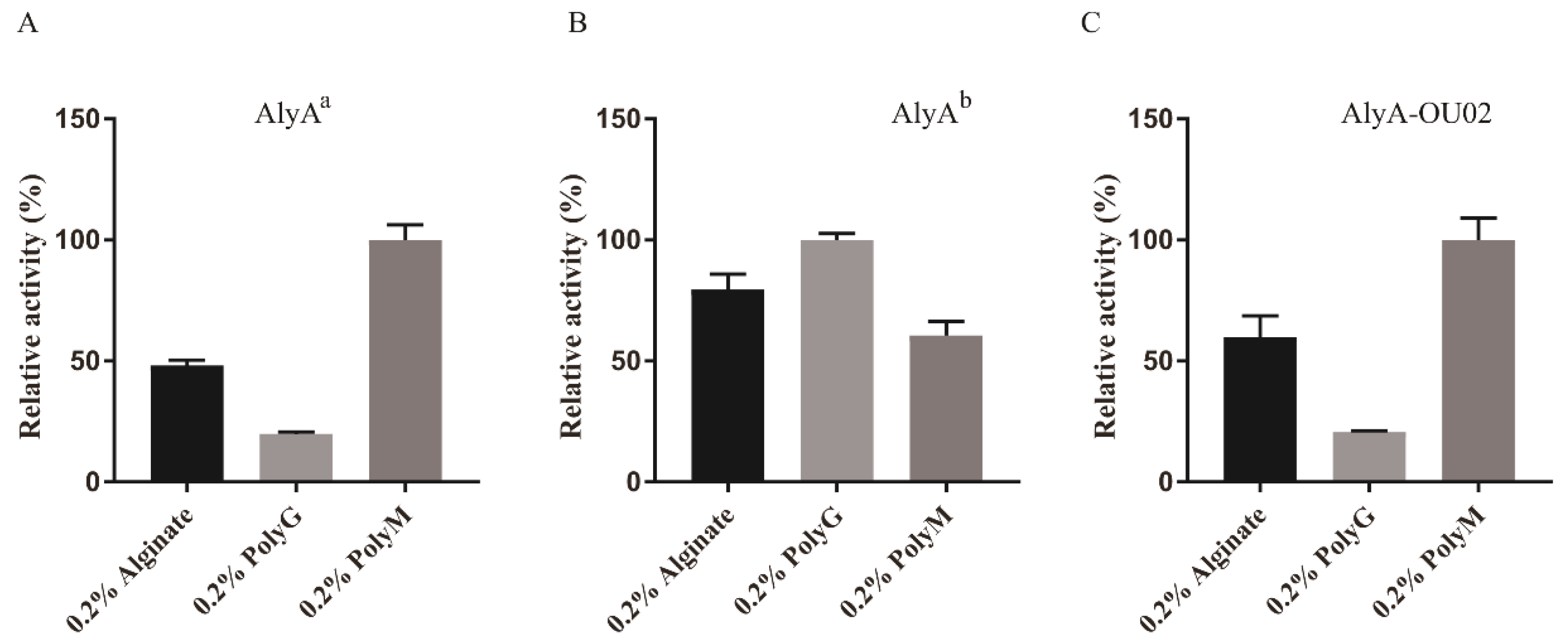
To determine the substrate specificity, 0.2% alginate, polyG, and polyM were used as the substrates to study the enzyme activities. The relative activities of AlyAa toward alginate, polyG, and polyM were 48.1% ± 2.2%, 20.0% ± 2.2%, and 100.0% ± 6.2%, respectively, indicating that AlyAa was a polyM-preferred alginate lyase (Figure 7A). AlyAb showed slight polyG preference (Figure 7B). The relative activities of AlyA-OU02 toward alginate, polyG, and polyM, were 60.0% ± 8.7%, 20.7% ± 0.4%, and 100.0% ± 9.1%, respectively (Figure 7C), which was similar to AlyAa. The specific activities of AlyA-OU02, AlyAa, and AlyAb toward alginate, polyG, and polyM are shown in Table 2. The substrate specificities of PL7 alginate lyases were related with the protein sequences in the conserved regions [17]. Crystal structures and sequence analyses showed that the three conserved regions in PL7 family formed the cavity composed of a jelly roll β-sandwich structure, which was assumed to bind to a suitable substrate [24]. The conserved amino acids are thought to be pivotal in catalytic activity or folding of the structure. Recent studies have revealed that the polyM-specific, polyG-specific, and polyMG alginate lyases contained QVH, QIH, and QIH in the conserved regions, respectively [17]. As shown in Table 1, the alginate lyases that showed activities toward polyG and polyM contained QIH in the conserved region, with the exception of rAlgSV1-PL7 [35]. A9mT and PyAly contained QVH and were polyM-specific [24,38]. The polyG-specific enzymes, such as Alg2A, ALG-5, and Aly2, all contained QIH sequence [26,30,37]. Contrary to this accepted rule, AlyAa was active toward polyM, containing QIH sequence which was similar to FlAlyA and AlyDW11 [27,31].
2.4. Thin-Layer Chromatography Analysis of the Degradation Products
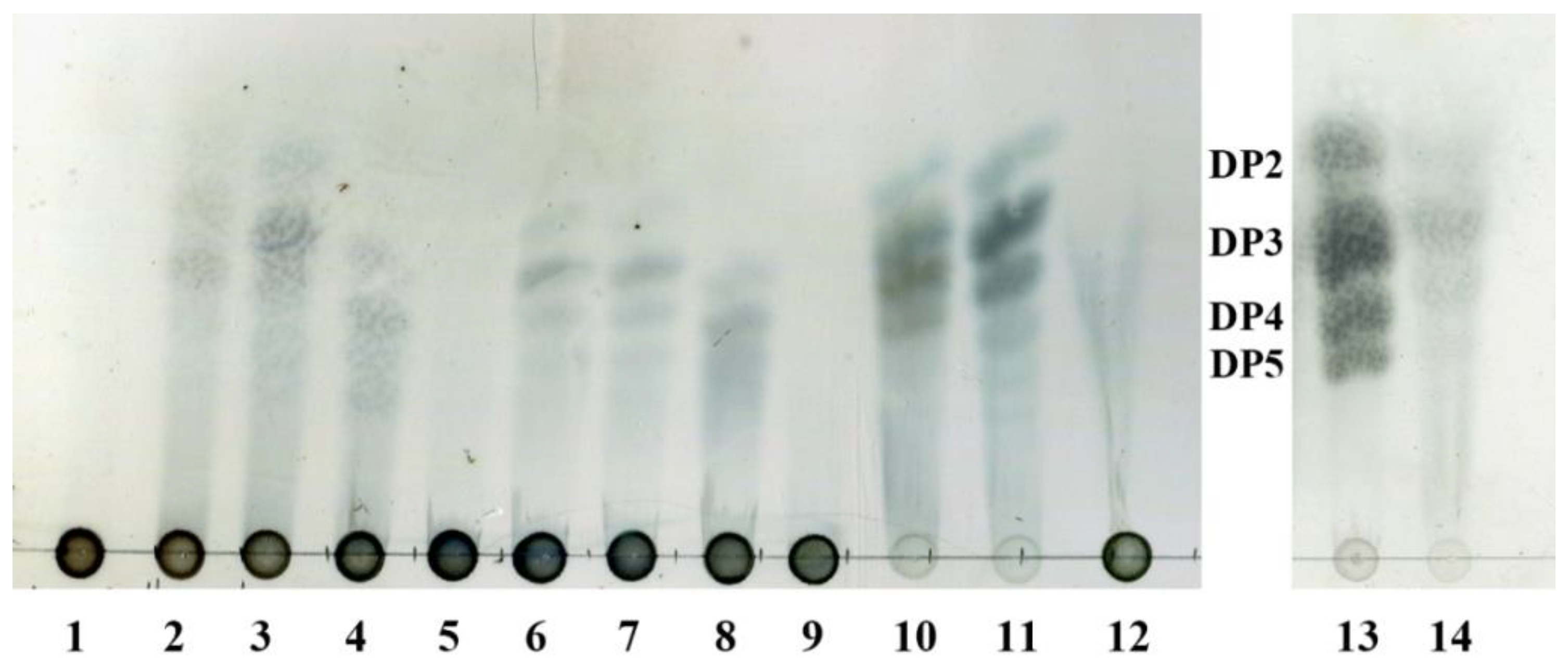
To investigate the action modes of AlyA-OU02, AlyAa, and AlyAb, the degradation products of alginate, polyG, and polyM after 1 h were analyzed by thin-layer chromatography (TLC). As shown in Figure 8, AlyAa degraded alginate and polyM into di-, tri-, and tetra-oligosaccharides, and trisaccharide was the main product. The products patterns of AlyA-OU02 were similar to AlyAa. For AlyAb, tri-, tetra-, and penta-oligosaccharides were the main hydrolysis products, and AlyAb had limited activity toward polyM. The results were consistent with the enzymatic assays. We also analyzed the overnight products of alginate, polyG, and polyM, and the product patterns remained the same (Figure S4). This indicated that AlyA-OU02 and AlyAa might be potential tools for preparation of lower molecular weight polyM products which have wide pharmaceutical applications.
3. Materials and Methods
3.1. Materials
Alginate was purchased from Shanghai Yuanye co., Ltd. The alginate viscosity is 4500 cP at 2% (w/v) concentration at 25 °C. The M/G ratio of alginate is 0.6 as determined by NMR analysis. PolyG and polyM were prepared according to the reference [10]. Other chemicals and reagents used in this study were of analytical grade.
3.2. Sequence Analysis of AlyA-OU02
For functional annotation, the BLAST algorithm on the National Center for Biotechnology Information server (http://www.ncbi.nlm.nih.gov) was used to analyze the similarity of the amino acid sequences. Molecular weights of the enzymes were estimated by the peptide mass tool on the ExPASY server of the Swiss Institute of Bioinformatics (http://swissmodel.expasy.org/). Multiple sequence alignments were performed by MEGA version 5.05.
3.3. Construction of Expression Vectors
The gene encoding AlyA-OU02 was amplified from genomic Vibrio splendidus OU02 DNA (unpublished data) using the forward primer (5’-CGGAATTCAGTAGTTCAAATAGCTCGACTG-3′) and reverse primer (5′-CCGCTCGAGTCAATAGTGTGCCGCTCTAAGAG-3′). The AlyAa gene was amplified using the forward primer (5’-GGAATTCAGTAGTTCAAATAGCTCGACTG-3′) and reverse primer (5′-CCGCTCGAGTTATGACCCATTGATTTG-3′). The AlyAb gene was amplified using the forward primer (5’-GGAATTCAACGATTGGGACATTAATGATTG-3′) and reverse primer (5′-CCGCTCGAGTTAATAGTGTGCCGCTCTAAG-3′). DNA amplification products were cloned into the expression vector pET-32a-PreScission. The recombinant plasmids were transformed into E. coli BL21 (DE3) cells. DNA sequencing was used to confirm the integrities of the nucleotide sequences of newly constructed plasmids.
Site-directed mutagenesis was conducted using the QuikChange Site-Directed Mutagenesis Kit (Aglient Technologies, CA, USA). For generation of the H139N mutant, the primers used in this study were forward primer (5′-GTGACGTTGCTGCAGATAAACAATAAGGGGACTGATG-3′) and reverse primer (5′-CATCAGTCCCCTTATTGTTTATCTGCAGCAACGTCAC-3′). For the Y250F mutant, the primers used in this study were forward primer (5′-GCTACTTCAAAGCGGGTATCTTTAACCA``ATTTGAGAATGGTG-3′) and reverse primer (5′-CACCATTCTCAAATTGGTTAAAGATACCCGCTTTGAAGTAGC-3′).
3.4. Expression and Purification of AlyA-OU02, AlyAa and AlyAb
E. coli BL21 (DE3) cells harboring the plasmid were initially cultured in LB broth containing ampicillin at 37 °C. When the cell density reached an A600 value of 0.6, 0.1 mM (final concentration) isopropyl β-d-1-thiogalactopyranoside was used to induce enzyme expression for 8 h at 16 °C. The cells were harvested by centrifugation (1500× g, 30 min, 4 °C) and sonicated in 20 mM Tris, 500 mM NaCl (pH 7.5). The lysates were clarified by centrifugation (20,000× g, 30 min, 4 °C). The supernatant containing soluble proteins was loaded onto a Ni-NTA column. The samples were eluted with a linear gradient of 10–500 mM imidazole. The molecular mass and the purities of the enzymes were analyzed by 12% SDS-PAGE.
3.5. Enzymatic Activity Assay
To determine the activities of the alginate lyases, 20 μL diluted enzyme (5 μM) was added to 180 μL substrate solution containing 0.2% (w/v) substrate, 50 mM Tris-HCl, and 200 mM NaCl (pH 7.5). After incubation at 25 °C for 10 min, alginate lyase activity was determined by measuring the increased absorbance at 235 nm of the reaction buffer. One unit was defined as the amount of enzyme required to increase the absorbance at 235 nm by 0.1 per min [40].
For kinetic analysis, the substrate concentrations varied from 0.33 mg/mL to 4 mg/mL. The reactions (200 μL for each assay) were carried out in 50 mM Tris-HCl (pH 7.5) with 200 mM NaCl at 25 °C for 10 min. After incubation, 200 μL of dinitrosalicylic acid was added to the reaction mixture, followed by heating at 100 °C for 5 min and centrifugation. The absorbance at 520 nm of the resulting supernatant was measured.
To determine the effects of temperature on enzyme activities, the reactions (200 μL for each assay) were carried out in 50 mM Tris-HCl (pH 7.5) with 200 mM NaCl at different temperatures (10–60 °C) for 10 min, with alginate as substrate. After incubation, 200 μL of dinitrosalicylic acid was added to the reaction mixture, followed by heating at 100 °C for 5 min and centrifugation. The absorbance at 520 nm of the resulting supernatant was measured. To determine the effects of pH on enzyme activities, the reactions (200 μL for each assay) were carried out in buffers with different pH values, including NaAC-HAC buffer (pH 5.0–6.0) and Tris-HCl buffer (pH 7.0–10.0) at 25 °C for 10 min, with alginate as substrate. The increased absorbance at 520 nm was measured.
To determine the thermostabilities, the enzyme solutions were incubated at various temperatures for 1 h. The reactions (200 μL for each assay) were carried out in 50 mM Tris-HCl (pH 7.5) with 200 mM NaCl at 25 °C for 10 min with 0.2% (w/v) alginate as substrate. After incubation, 200 μL of dinitrosalicylic acid was added to the reaction mixture, followed by heating at 100 °C for 5 min and centrifugation. The absorbance at 520 nm of the resulting supernatant was measured.
The effects of metal ions on enzyme activities were carried out by incubating enzymes at 4 °C for 1 h in the presence of various metal ions (1 mM) or EDTA (5 mM) in 50 mM Tris-HCl buffer (pH 7.5). The reactions (200 μL for each assay) were carried out in 50 mM Tris-HCl (pH 7.5) at 25 °C for 10 min with alginate as substrate. The increased absorbance at 235 nm was measured.
To study the substrate specificities, the reactions (200 μL for each assay) were carried out in 50 mM Tris-HCl (pH 7.5) with 200 mM NaCl at 25 °C for 10 min, using 0.2% (w/v) alginate, polyM, and polyG separately as substrates. The increased absorbance at 235 nm was measured.
3.6. TLC Analysis of the Degradation Products
To analyze the oligosaccharides produced by the enzymes, 20 μL diluted enzyme (5 μM) was added to 180 μL substrate solution containing 0.2% (w/v) substrate, 50 mM Tris-HCl, and 200 mM NaCl (pH 7.5). After incubation at 25 °C for 1 h or 12 h, the reaction buffer was boiled for 5 min, and analyzed by TLC with the solvent system (1-butanol/formic acid/water 4:6:1). The TLC plate was sprayed using 10% (v/v) sulfuric acid in ethanol, and then heated at 130 °C for 5 min.
4. Conclusions
In this study, we reported a new alginate lyase derived from the marine Vibrio splendidus OU02. AlyA-OU02 was characterized as a novel PL7 alginate lyase containing two PL7 domains. Surprisingly, both of the two domains showed alginate lyase activities. Although both domains contained QIH sequence in the conserved regions, which was thought to be polyG-specific, AlyA-OU02 turned out to be polyM-preferred. The two domains exhibited different enzymatic properties. The optimal temperature, pH, metal ions effects, and degradation patterns of AlyA-OU02 were similar to AlyAa. The putative reason was that the enzyme activity of AlyAa was much higher than AlyAb at 25 °C. Further work will be focused on obtaining crystals of the enzymes to elucidate the catalytic mechanism of AlyA-OU02.
Supplementary Materials
The following are available online at https://www.mdpi.com/1660-3397/16/9/295/s1, Figure S1: SDS-PAGE analysis of purified H139N/Y250F mutants of AlyAa. Lane 1, H139 mutant of AlyAa; Lane 2, Y250F mutants of AlyAa; Lane M, molecular weight markers, Figure S2: Enzyme activities of H139 and Y250F mutants of AlyAa. The relative enzyme activity of AlyAa was designated as 100%, Figure S3: Effects of pH on enzyme activities of AlyA-OU02, AlyAa, and AlyAb. pH effects on enzyme activities of AlyA-OU02 (A), AlyAa (B), and AlyAb (C) in sodium phosphate buffer (pH 6.0–8.0) and Tris-HCl buffer (pH 7.0–10.0), Figure S4: TLC analysis of the hydrolytic products of AlyA-OU02, AlyAa, and AlyAb. Analysis of the hydrolytic products of AlyA-OU02 (lane 1–3), AlyAa (lane 4–6), and AlyAb (lane 7–9) for 12 h with either alginate, polyG, and polyM as substrates. Lane 10, DP2, DP3, DP4, and DP5 mean the alginate disaccharide, trisaccharide, tetrasaccharide, and pentasaccharide, respectively.
Author Contributions
Q.L. and J.Z. conceived and designed the experiments; J.Z. and K.Z. performed the experiments; K.Z. and X.L. analyzed the data; W.L. contributed reagents/materials/analysis tools; J.Z. wrote the paper; W.L. and A.J. edited the paper.
Funding
This work was supported by the Shandong Provincial Natural Science Foundation, China (ZR2017PH053), the Shandong Provincial Natural Science Foundation, China (ZR2016DB17), the China Postdoctoral Science Foundation (2017M612297), and the National Natural Science Foundation of China (41706151).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pawar, S.N.; Edgar, K.J. Alginate derivatization: A review of chemistry, properties and applications. Biomaterials 2012, 33, 3279–3305. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wargacki, A.J.; Leonard, E.; Win, M.N.; Regitsky, D.D.; Santos, C.N.; Kim, P.B.; Cooper, S.R.; Raisner, R.M.; Herman, A.; Sivitz, A.B.; et al. An engineered microbial platform for direct biofuel production from brown macroalgae. Science 2012, 335, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Falkeborg, M.; Cheong, L.Z.; Gianfico, C.; Sztukiel, K.M.; Kristensen, K.; Glasius, M.; Xu, X.; Guo, Z. Alginate oligosaccharides: Enzymatic preparation and antioxidant property evaluation. Food Chem. 2014, 164, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Tusi, S.K.; Khalaj, L.; Ashabi, G.; Kiaei, M.; Khodagholi, F. Alginate oligosaccharide protects against endoplasmic reticulum- and mitochondrial-mediated apoptotic cell death and oxidative stress. Biomaterials 2011, 32, 5438–5458. [Google Scholar] [CrossRef] [PubMed]
- An, Q.D.; Zhang, G.L.; Wu, H.T.; Zhang, Z.C.; Zheng, G.S.; Luan, L.; Murata, Y.; Li, X. Alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. J. Appl. Microbiol. 2009, 106, 161–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Hu, Y.; Zhang, L.; Wang, Y.; Wang, S.; Zhang, Y.; Guo, H.; Ji, D.; Wang, Y. Alginate Oligosaccharide DP5 Exhibits Antitumor Effects in Osteosarcoma Patients following Surgery. Front. Pharmacol. 2017, 8, 623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertesvag, H. Alginate-modifying enzymes: Biological roles and biotechnological uses. Front. Microbiol. 2015, 6, 523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Suzuki, K.; Inoue, A.; Ojima, T. A novel oligoalginate lyase from abalone, Haliotis discus hannai, that releases disaccharide from alginate polymer in an exolytic manner. Carbohydr. Res. 2006, 341, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Zhang, K.; Zhu, Q.; Li, Z.; Liu, Y.; Fitzek, E.; Yohe, T.; Zhao, L.; Li, W.; Liu, T.; et al. Structural and biochemical characterization of a multidomain alginate lyase reveals a novel role of CBM32 in CAZymes. Biochim. Biophys. Acta 2018, 1862, 1862–1869. [Google Scholar] [CrossRef] [PubMed]
- Ogura, K.; Yamasaki, M.; Mikami, B.; Hashimoto, W.; Murata, K. Substrate recognition by family 7 alginate lyase from Sphingomonas sp. A1. J. Mol. Biol. 2008, 380, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.M.; Miyakawa, T.; Inoue, A.; Nishiyama, R.; Nakamura, A.; Asano, A.; Ojima, T.; Tanokura, M. Structural basis for controlling the enzymatic properties of polymannuronate preferred alginate lyase FlAlyA from the PL-7 family. Chem. Commun. 2018, 54, 555–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, Y.; Kawada, R.; Iwasaki, K.; Kimura, Y.; Oda, T.; Muramatsu, T. Cloning and sequence analysis of a gene (aly PG) encoding poly (alpha-L-guluronate) lyase from Corynebacterium sp. strain ALY-1. J. Biosci. Bioeng. 2000, 89, 199–202. [Google Scholar] [CrossRef]
- Wong, T.Y.; Preston, L.A.; Schiller, N.L. Alginate lyaseL: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu. Rev. Microbiol. 2000, 54, 289–340. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Bernard, T.; Rancurel, C.; Brumer, H.; Coutinho, P.M.; Henrissat, B. A hierarchical classification of polysaccharide lyases for glycogenomics. Biochem. J. 2010, 432, 437–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osawa, T.; Matsubara, Y.; Muramatsu, T.; Kimura, M.; Kakuta, Y. Crystal structure of the alginate (poly alpha-l-guluronate) lyase from Corynebacterium sp. at 1.2 A resolution. J. Mol. Biol. 2005, 345, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Yin, H. Alginate lyase: Review of major sources and classification, properties, structure-function analysis and applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Badur, A.H.; Jagtap, S.S.; Yalamanchili, G.; Lee, J.K.; Zhao, H.; Rao, C.V. Alginate lyases from alginate-degrading Vibrio splendidus 12B01 are endolytic. Appl. Environ. Microbiol. 2015, 81, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Gong, Q.H.; Song, K.; Li, J.B.; Yu, W.G. Cloning, sequence analysis and expression of gene alyVI encoding alginate lyase from marine bacterium Vibrio sp. QY101. DNA Seq. 2004, 15, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Tan, H.; Qin, Y.; Xu, Q.; Du, Y.; Yin, H. Characterization of a new endo-type alginate lyase from Vibrio sp. W13. Int. J. Biol. Macromol. 2015, 75, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a Novel Alginate Lyase from Nitratiruptor sp. SB155-2 Thriving at Deep-sea Hydrothermal Vents and Identification of the Residues Responsible for Its Heat Stability. J. Biol. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef] [PubMed]
- Chavagnat, F.; Heyraud, A.; Colin-Morel, P.; Guinand, M.; Wallach, J. Catalytic properties and specificity of a recombinant, overexpressed D-mannuronate lyase. Carbohydr. Res. 1998, 308, 409–415. [Google Scholar] [CrossRef]
- Yamasaki, M.; Moriwaki, S.; Miyake, O.; Hashimoto, W.; Murata, K.; Mikami, B. Structure and function of a hypothetical Pseudomonas aeruginosa protein PA1167 classified into family PL-7: A novel alginate lyase with a beta-sandwich fold. J. Biol. Chem. 2004, 279, 31863–31872. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, X.; Zhang, L.; Yu, W.; Han, F. Cloning, Expression, and Characterization of a Cold-Adapted and Surfactant-Stable Alginate Lyase from Marine Bacterium Agarivorans sp. L11. J. Microbiol. Biotechnol. 2015, 25, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhou, J.; Li, X.; Peng, Q.; Lu, H.; Du, Y. Characterization of a new alginate lyase from newly isolated Flavobacterium sp. S20. J. Ind. Microbiol. Biotechnol. 2013, 40, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Takadono, K.; Nishiyama, R.; Tajima, K.; Kobayashi, T.; Ojima, T. Characterization of an alginate lyase, FlAlyA, from Flavobacterium sp. strain UMI-01 and its expression in Escherichia coli. Mar. Drugs 2014, 12, 4693–4712. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.; Han, F.; Yu, W. Cloning, sequence analysis, and expression of gene alyPI encoding an alginate lyase from marine bacterium Pseudoalteromonas sp. CY24. Can. J. Microbiol. 2009, 55, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Miyake, O.; Ochiai, A.; Hashimoto, W.; Murata, K. Origin and diversity of alginate lyases of families PL-5 and -7 in Sphingomonas sp. strain A1. J. Bacteriol. 2004, 186, 2891–2896. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Lee, E.Y.; Kim, H.S. Cloning and characterization of alginate lyase from a marine bacterium Streptomyces sp. ALG-5. Mar. Biotechnol. 2009, 11, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.J.; Baik, K.S.; Park, S.C.; Choe, H.N.; Seong, C.N.; Shin, T.S.; Woo, H.C.; Cho, J.Y.; Kim, D. Characterization of alginate lyase gene using a metagenomic library constructed from the gut microflora of abalone. J. Ind. Microbiol. Biotechnol. 2012, 39, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Wang, Q.; Lu, M.; Xu, C.; Li, F.; Zhang, R.; Liao, W.; Huang, S. AlgM4: A New Salt-Activated Alginate Lyase of the PL7 Family with Endolytic Activity. Mar. Drugs 2018, 16, 120. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Sun, Y.; Ni, F.; Ning, L.; Yao, Z. Characterization of a new endo-type alginate lyase from Vibrio sp. NJU-03. Int. J. Biol. Macromol. 2018, 108, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Hu, F.; Yuan, H.; Sun, Y.; Yao, Z. Biochemical Characterization and Degradation Pattern of a Unique pH-Stable PolyM-Specific Alginate Lyase from Newly Isolated Serratia marcescens NJ-07. Mar. Drugs 2018, 16, 129. [Google Scholar] [CrossRef] [PubMed]
- Yagi, H.; Isobe, N.; Itabashi, N.; Fujise, A.; Ohshiro, T. Characterization of a Long-Lived Alginate Lyase Derived from Shewanella Species YH1. Mar. Drugs 2017, 16, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, X.; Shi, H.; Zhou, J.; Tan, Z.; Yuan, M.; Yao, P.; Liu, X. Characterization of a Novel Alginate Lyase from Marine Bacterium Vibrio furnissii H1. Mar. Drugs 2018, 16, 30. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, Q.; Lu, D.; Han, W.; Li, F. A Novel Bifunctional Endolytic Alginate Lyase with Variable Alginate-Degrading Modes and Versatile Monosaccharide-Producing Properties. Front. Microbiol. 2018, 9, 167. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Mashino, C.; Uji, T.; Saga, N.; Mikami, K.; Ojima, T. Characterization of an Eukaryotic PL-7 Alginate Lyase in the Marine Red Alga Pyropia yezoensis. Curr. Biotechnol. 2015, 4, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhu, Y.; Men, Y.; Zeng, Y.; Sun, Y. Purification and Characterization of a Novel Alginate Lyase from the Marine Bacterium Bacillus sp. Alg07. Mar. Drugs 2018, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Gu, J.; Cheng, Y.; Liu, H.; Li, Y.; Li, F. Novel Alginate Lyase (Aly5) from a Polysaccharide-Degrading Marine Bacterium, Flammeovirga sp. Strain MY04: Effects of Module Truncation on Biochemical Characteristics, Alginate Degradation Patterns, and Oligosaccharide-Yielding Properties. Appl. Environ. Microbiol. 2016, 82, 364–374. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sequence properties of the alginate lyase AlyA-OU02 from marine Vibrio splendidus OU02. (A) Module organization of AlyA-OU02. The two alginate_lyase 2 modules were putative catalytic domains (Ser40 to Ala264, Trp278 to His563). The full-length protein and the two alginate_lyase 2 modules were expressed to yield the recombinant protein AlyA-OU02 (Ser1 to Tyr564), AlyAa (Ser1 to Ser275), and AlyAb (Asn276 to Tyr564). The indicated amino acid residues were hypothesized catalytic sites. (B) Comparison of the partial amino acid sequences of AlyAa and AlyAb with PL7 alginate lyases AlyVI from Vibrio sp. QY101 (AAP45155.1), Algb from Vibrio sp. W13 (AIY22661.1), NitAly from Nitratiruptor sp. SB155-2 (BAF69299.1), AlxM from Photobacterium sp. ATCC 43367 (CAA49630.1).
Figure 1.
Sequence properties of the alginate lyase AlyA-OU02 from marine Vibrio splendidus OU02. (A) Module organization of AlyA-OU02. The two alginate_lyase 2 modules were putative catalytic domains (Ser40 to Ala264, Trp278 to His563). The full-length protein and the two alginate_lyase 2 modules were expressed to yield the recombinant protein AlyA-OU02 (Ser1 to Tyr564), AlyAa (Ser1 to Ser275), and AlyAb (Asn276 to Tyr564). The indicated amino acid residues were hypothesized catalytic sites. (B) Comparison of the partial amino acid sequences of AlyAa and AlyAb with PL7 alginate lyases AlyVI from Vibrio sp. QY101 (AAP45155.1), Algb from Vibrio sp. W13 (AIY22661.1), NitAly from Nitratiruptor sp. SB155-2 (BAF69299.1), AlxM from Photobacterium sp. ATCC 43367 (CAA49630.1).
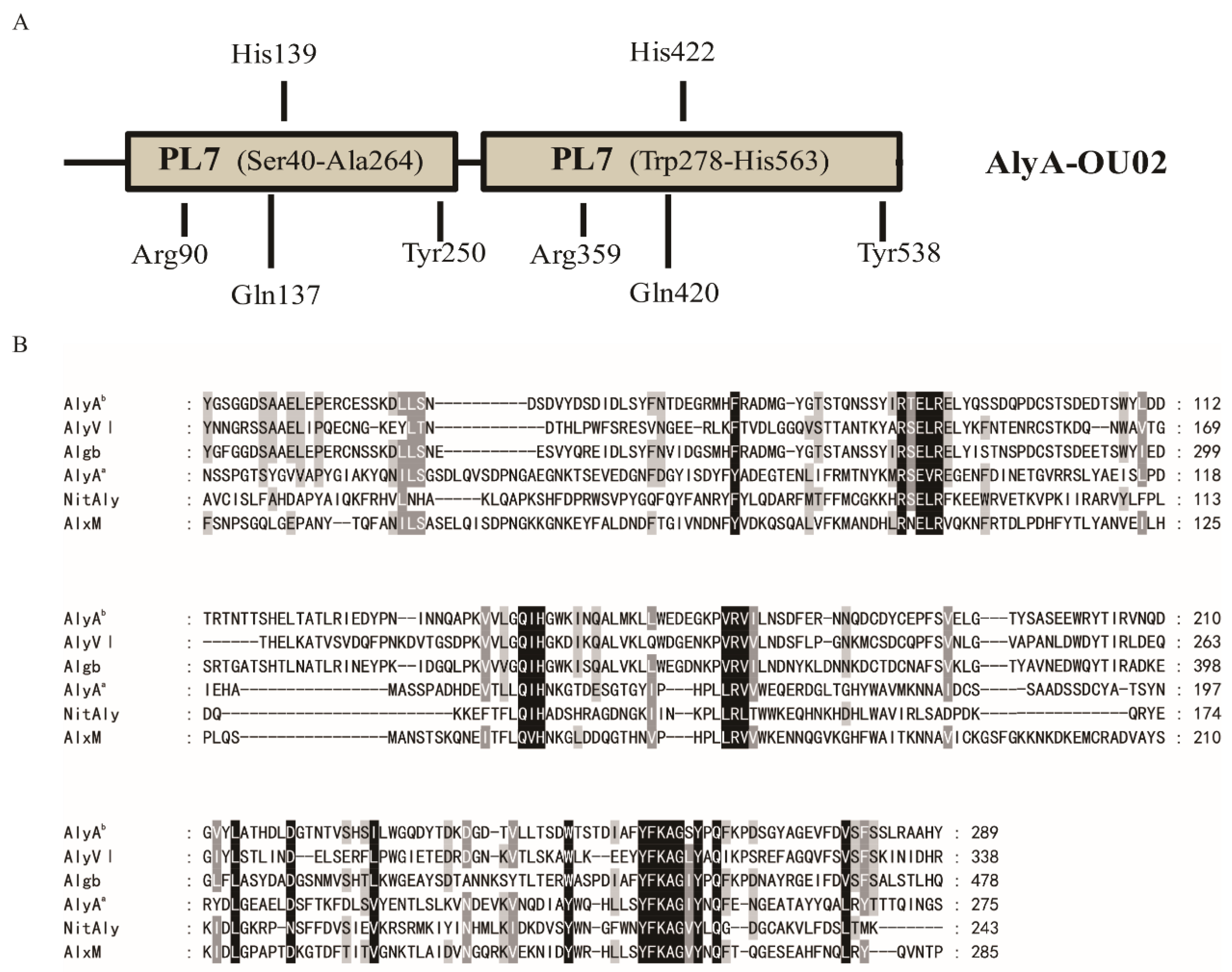
Figure 2.
SDS-PAGE analysis of purified AlyA-OU02, AlyAa, and AlyAb. Lane M, molecular weight markers; Lane 1, purified AlyA-OU02; Lane 2, purified AlyAa; Lane 3, purified AlyAb.
Figure 2.
SDS-PAGE analysis of purified AlyA-OU02, AlyAa, and AlyAb. Lane M, molecular weight markers; Lane 1, purified AlyA-OU02; Lane 2, purified AlyAa; Lane 3, purified AlyAb.
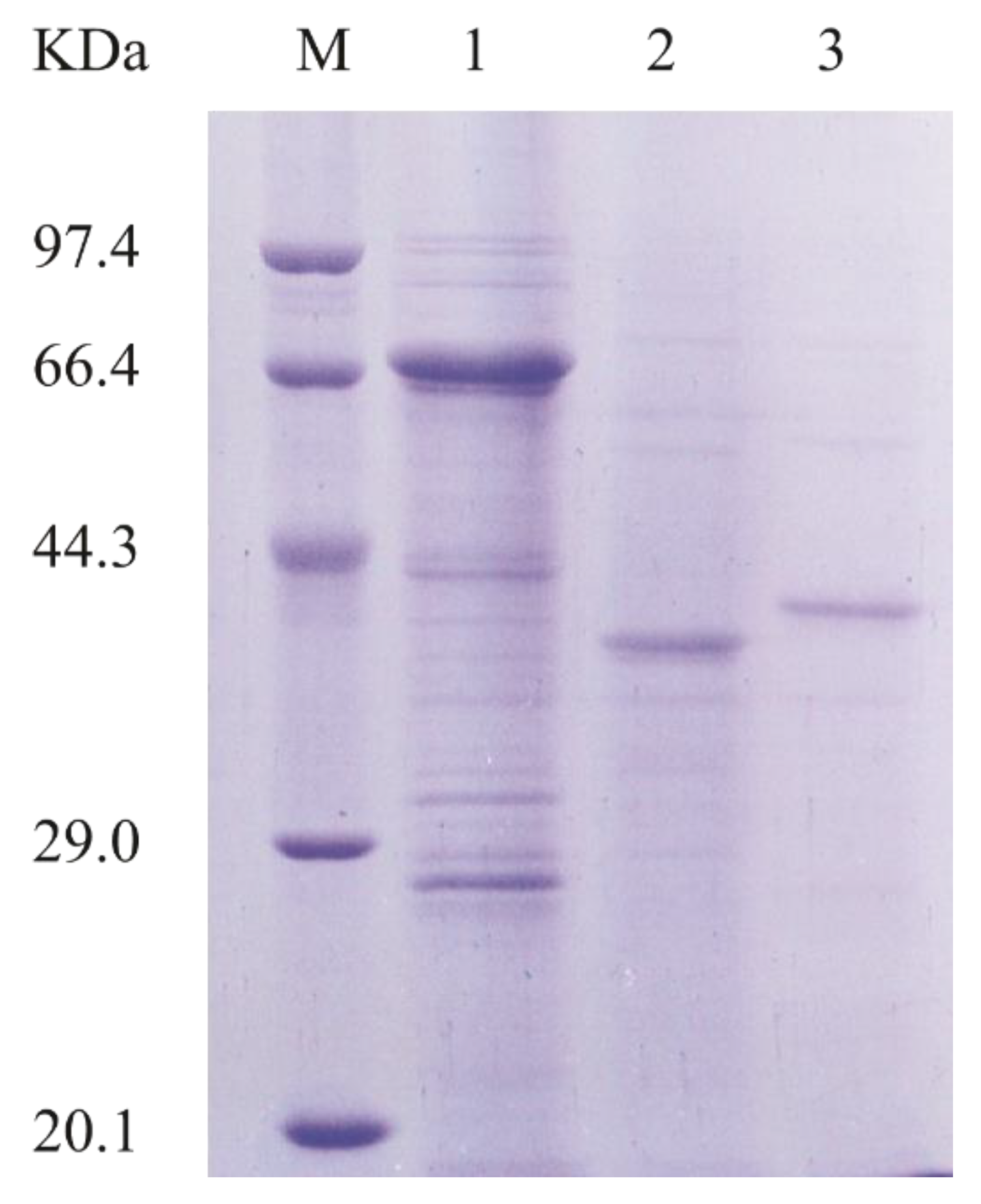
Figure 3.
Enzyme activity assay of AlyA-OU02, AlyAa and AlyAb toward alginate. The measurement was carried out in Tris-HCl buffer with 200 mM NaCl (pH 7.5) using 0.2% alginate as substrate.
Figure 3.
Enzyme activity assay of AlyA-OU02, AlyAa and AlyAb toward alginate. The measurement was carried out in Tris-HCl buffer with 200 mM NaCl (pH 7.5) using 0.2% alginate as substrate.
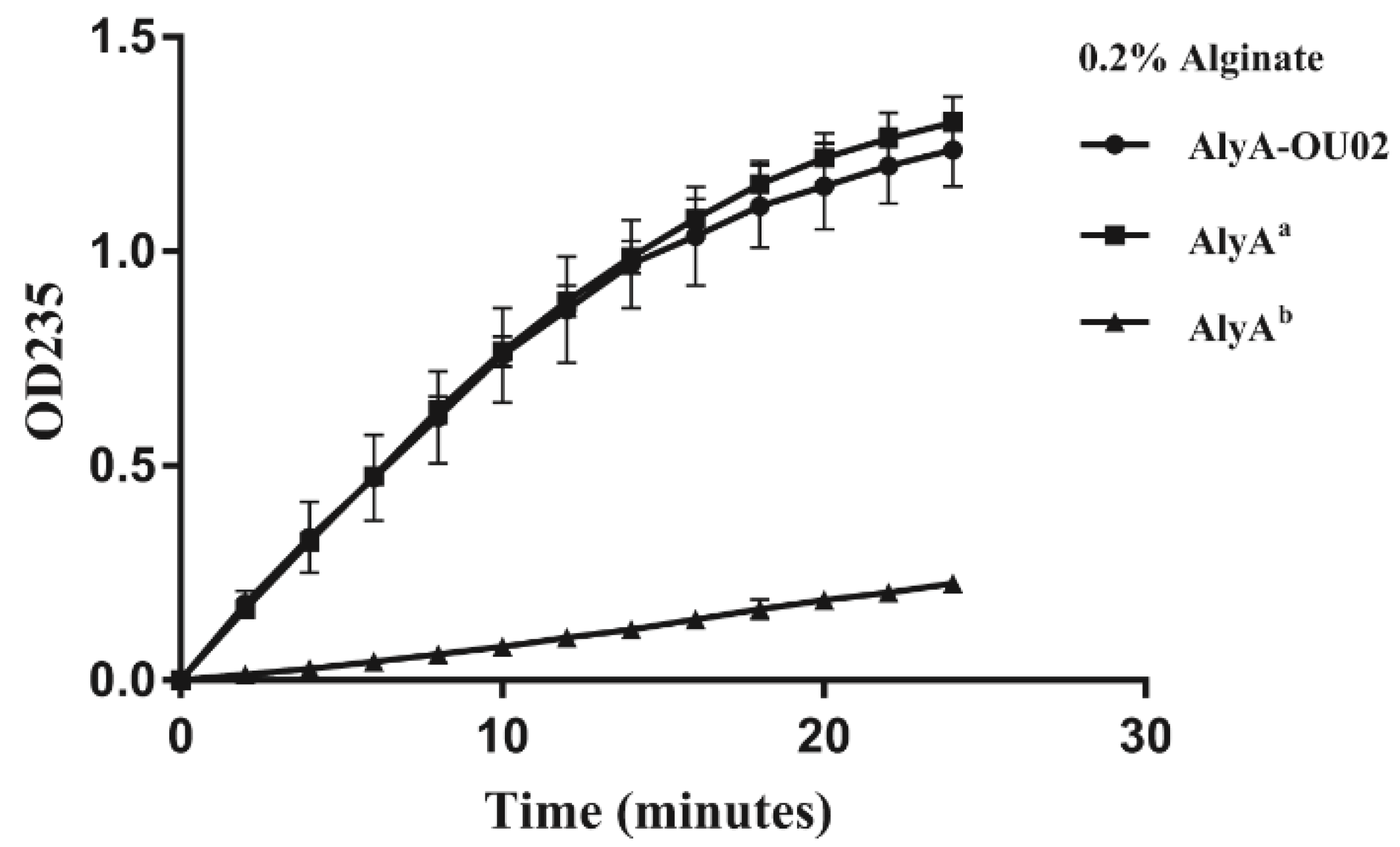
Figure 4.
Effects of temperature and pH on enzyme activities of AlyA-OU02, AlyAa, and AlyAb. (A) Optimal temperatures of the enzymes were determined in Tris-HCl buffer with 200 mM NaCl (pH 7.5) at different temperatures. (B) Optimal pH of the enzymes was determined at 25 °C in NaAc-HAc buffer (pH 5–6) and Tris-HCl buffer (pH 7–10).
Figure 4.
Effects of temperature and pH on enzyme activities of AlyA-OU02, AlyAa, and AlyAb. (A) Optimal temperatures of the enzymes were determined in Tris-HCl buffer with 200 mM NaCl (pH 7.5) at different temperatures. (B) Optimal pH of the enzymes was determined at 25 °C in NaAc-HAc buffer (pH 5–6) and Tris-HCl buffer (pH 7–10).

Figure 5.
Thermostabilities of AlyA-OU02, AlyAa, and AlyAb.
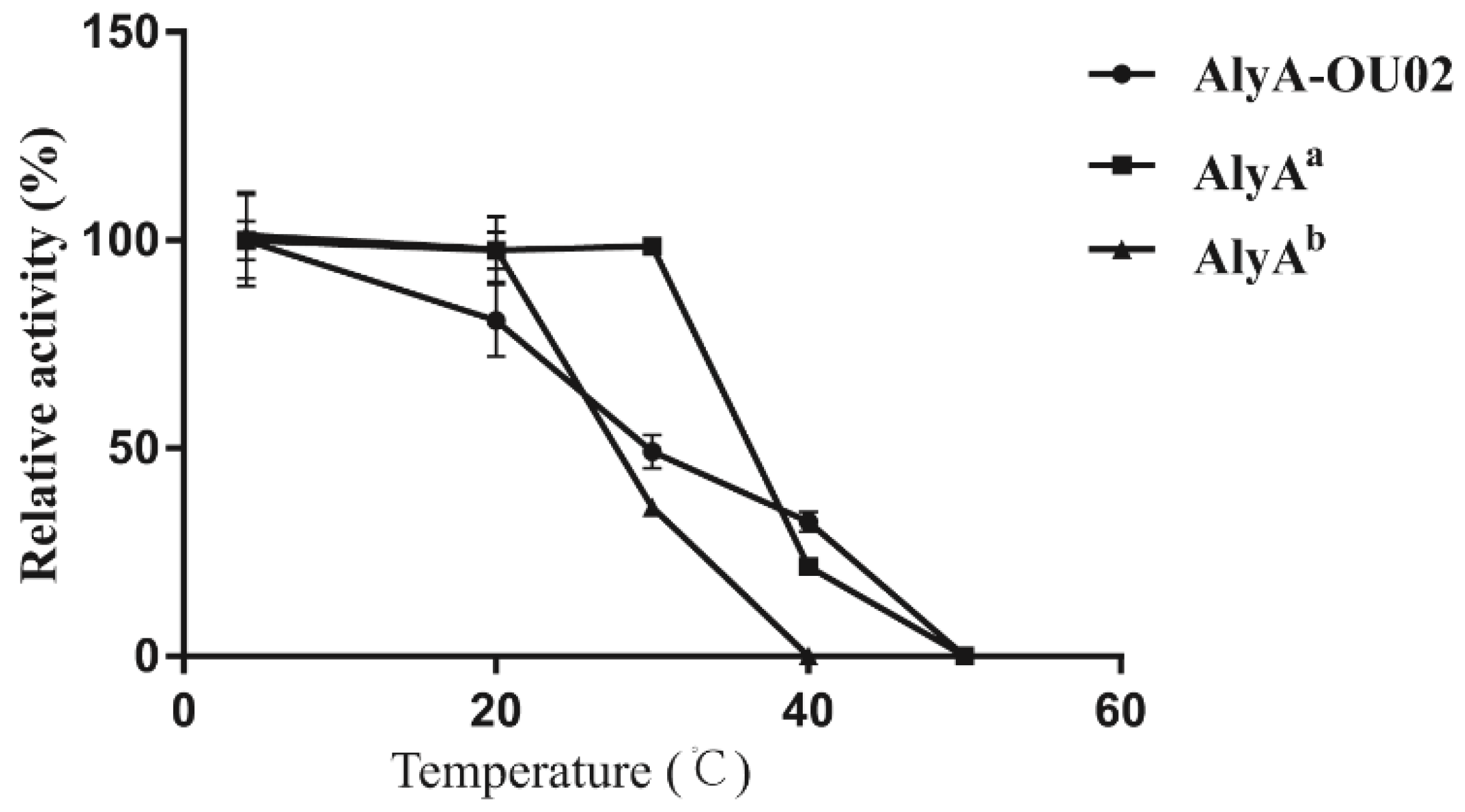
Figure 6.
Effects of NaCl and metal ions on enzymatic activities of AlyA-OU02, AlyAa, and AlyAb. (A) Influence of the concentration of NaCl. (B) Influence of the metal ions on AlyA-OU02. (C) Influence of the metal ions on AlyAa. (D) Influence of the metal ions on AlyAb. The enzymatic activity without metal ions served as the control, and the enzymatic activity was designated as 100%.
Figure 6.
Effects of NaCl and metal ions on enzymatic activities of AlyA-OU02, AlyAa, and AlyAb. (A) Influence of the concentration of NaCl. (B) Influence of the metal ions on AlyA-OU02. (C) Influence of the metal ions on AlyAa. (D) Influence of the metal ions on AlyAb. The enzymatic activity without metal ions served as the control, and the enzymatic activity was designated as 100%.

Figure 7.
Substrate specificities of AlyA-OU02, AlyAa, and AlyAb. (A) Relative enzyme activities of AlyAa toward alginate, polyM, and polyG. (B) Relative enzyme activities of AlyAb toward alginate, polyM, and polyG. (C) Relative enzyme activities of AlyA-OU02 toward alginate, polyM, and polyG.
Figure 7.
Substrate specificities of AlyA-OU02, AlyAa, and AlyAb. (A) Relative enzyme activities of AlyAa toward alginate, polyM, and polyG. (B) Relative enzyme activities of AlyAb toward alginate, polyM, and polyG. (C) Relative enzyme activities of AlyA-OU02 toward alginate, polyM, and polyG.
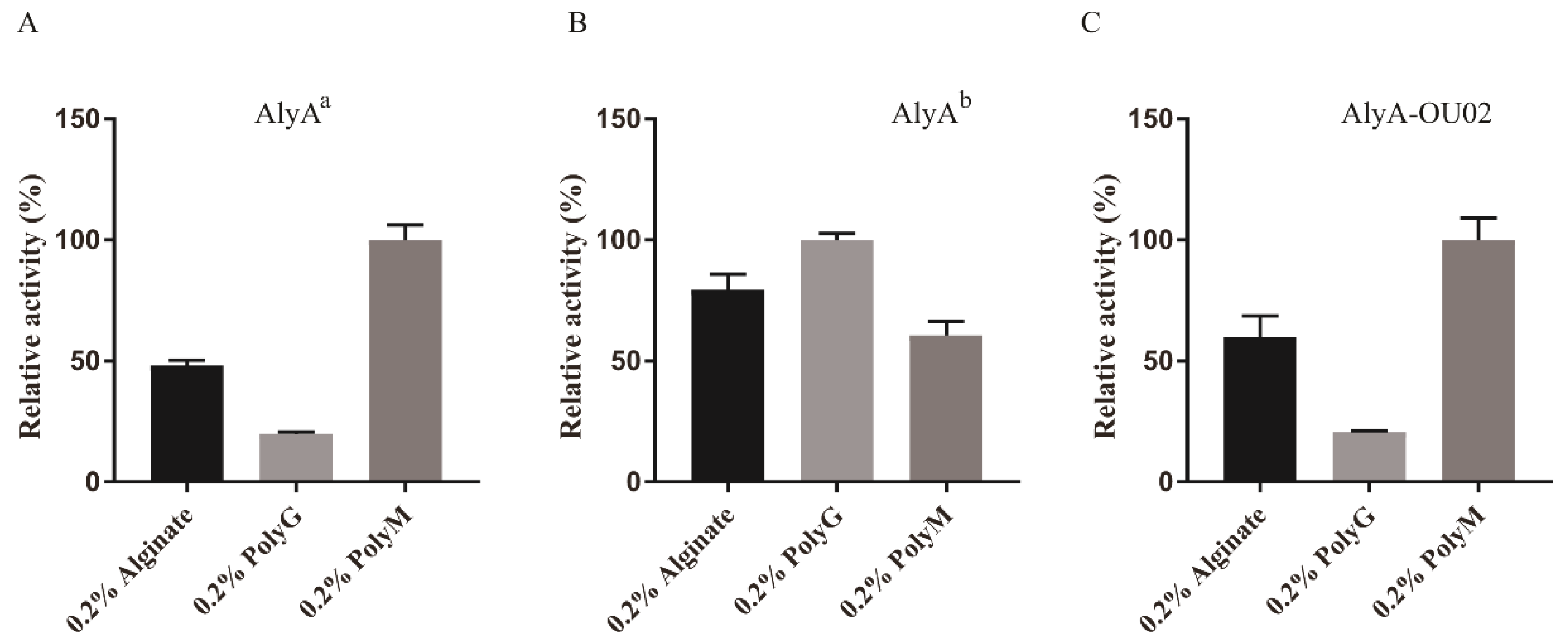
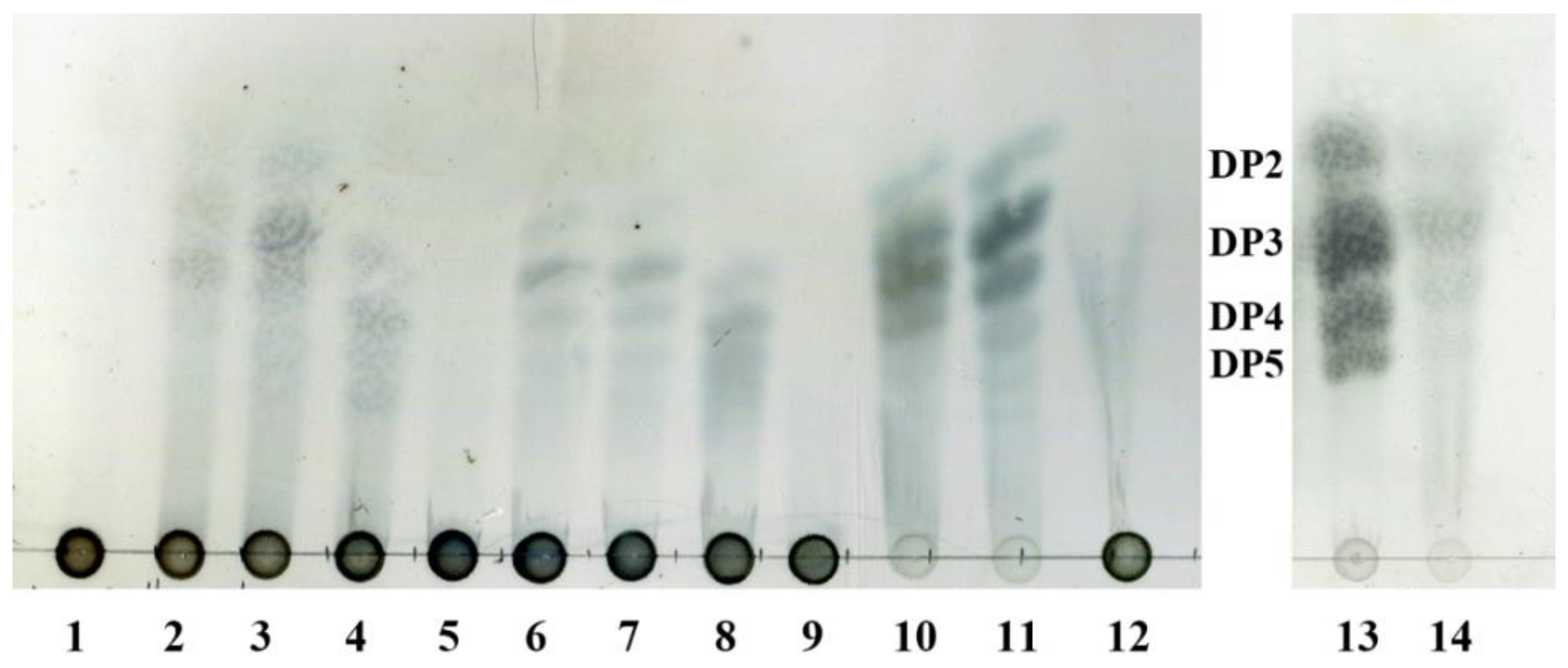
Figure 8.
TLC analysis of the hydrolytic products of AlyA-OU02, AlyAa, and AlyAb. Solutions of 0.2% (w/v) alginate, polyG, and polyM were each incubated with 5 uM AlyA-OU02, AlyAa, and AlyAb, respectively, at 25 °C for 1 h. Lane 1: alginate; lane 2–4: hydrolytic products of AlyA-OU02, AlyAa, and AlyAb toward alginate; lane 5: polyG; lane 6–8: hydrolytic products of AlyA-OU02, AlyAa, and AlyAb toward polyG; lane 9: polyM; lane 10–12: hydrolytic products of AlyA-OU02, AlyAa, and AlyAb toward polyM; lane 13: DP2, DP3, DP4, and DP5 mean the alginate disaccharide, trisaccharide, tetrasaccharide, and pentasaccharide, respectively; lane 14: hydrolytic products of AlyAa toward polyM.
Figure 8.
TLC analysis of the hydrolytic products of AlyA-OU02, AlyAa, and AlyAb. Solutions of 0.2% (w/v) alginate, polyG, and polyM were each incubated with 5 uM AlyA-OU02, AlyAa, and AlyAb, respectively, at 25 °C for 1 h. Lane 1: alginate; lane 2–4: hydrolytic products of AlyA-OU02, AlyAa, and AlyAb toward alginate; lane 5: polyG; lane 6–8: hydrolytic products of AlyA-OU02, AlyAa, and AlyAb toward polyG; lane 9: polyM; lane 10–12: hydrolytic products of AlyA-OU02, AlyAa, and AlyAb toward polyM; lane 13: DP2, DP3, DP4, and DP5 mean the alginate disaccharide, trisaccharide, tetrasaccharide, and pentasaccharide, respectively; lane 14: hydrolytic products of AlyAa toward polyM.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characterization of alginate lyases in PL7 family.
| Protein Name | Optimal pH/Temperature (°C) | Conserved Region QIH/QVH | Substrate Specificity | Reference |
|---|---|---|---|---|
| AlyL1 | 8.6/40 | QIH | PM, PG | [25] |
| Alg2A | 8.5/45 | QIH | PG | [26] |
| FlAlyA | 7.8/55 | QIH | PM | [27] |
| AlyPI | 7.0/40 | QIH | PM, PG | [28] |
| AI-II′ | 7.5/40 | QIH | PM, PG, PMG | [29] |
| ALG-5 | 8.0/30 | QIH | PG | [30] |
| AlyDW11 | 7.0/45 | QIH | PM | [31] |
| A9MT | 7.5/30 | QVH | PM | [24] |
| AlgM4 | 8.5/30 | QIH | PM, PG | [32] |
| AlgNJU-03 | 7.0/30 | QIH | PM, PG | [33] |
| AlgNJ-07 | 9.0/40 | - | PM | [34] |
| rAlgSV1-PL7 | 8.0/45 | QVH | PM, PG, PMG | [35] |
| AlyH1 | 7.5/40 | QIH | PM, PG | [36] |
| Aly2 | 6.0/40 | QIH | PG | [37] |
| Algb | 8.0/30 | QIH | PM, PG | [20] |
| PyAly | 8.0/35 | QVH | PM | [38] |
| AlyVI | 7.5/40 | QIH | PM, PG | [19] |
| NitAly | 6.0/70 | QIH | PM | [21] |
| AlxM | -/- | QVH | PM | [22] |
Table 2.
Specific activities of AlyA-OU02, AlyAa, and AlyAb toward alginate, polyG, and polyM.
| Specific Activity (U/mg) | Alginate | PolyG | PolyM |
|---|---|---|---|
| AlyA-OU02 | 119 | 41 | 198 |
| AlyAa | 252 | 104.5 | 520 |
| AlyAb | 24 | 33 | 15 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhuang, J.; Zhang, K.; Liu, X.; Liu, W.; Lyu, Q.; Ji, A. Characterization of a Novel PolyM-Preferred Alginate Lyase from Marine Vibrio splendidus OU02. Mar. Drugs 2018, 16, 295. https://doi.org/10.3390/md16090295
AMA Style
Zhuang J, Zhang K, Liu X, Liu W, Lyu Q, Ji A. Characterization of a Novel PolyM-Preferred Alginate Lyase from Marine Vibrio splendidus OU02. Marine Drugs. 2018; 16(9):295. https://doi.org/10.3390/md16090295
Chicago/Turabian StyleZhuang, Jingjing, Keke Zhang, Xiaohua Liu, Weizhi Liu, Qianqian Lyu, and Aiguo Ji. 2018. "Characterization of a Novel PolyM-Preferred Alginate Lyase from Marine Vibrio splendidus OU02" Marine Drugs 16, no. 9: 295. https://doi.org/10.3390/md16090295
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.