Preliminary Results on the Evaluation of the Occurrence of Tetrodotoxin Associated to Marine Vibrio spp. in Bivalves from the Galician Rias (Northwest of Spain)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Microbiological Results
2.2. Cell Assay (N2a) Results
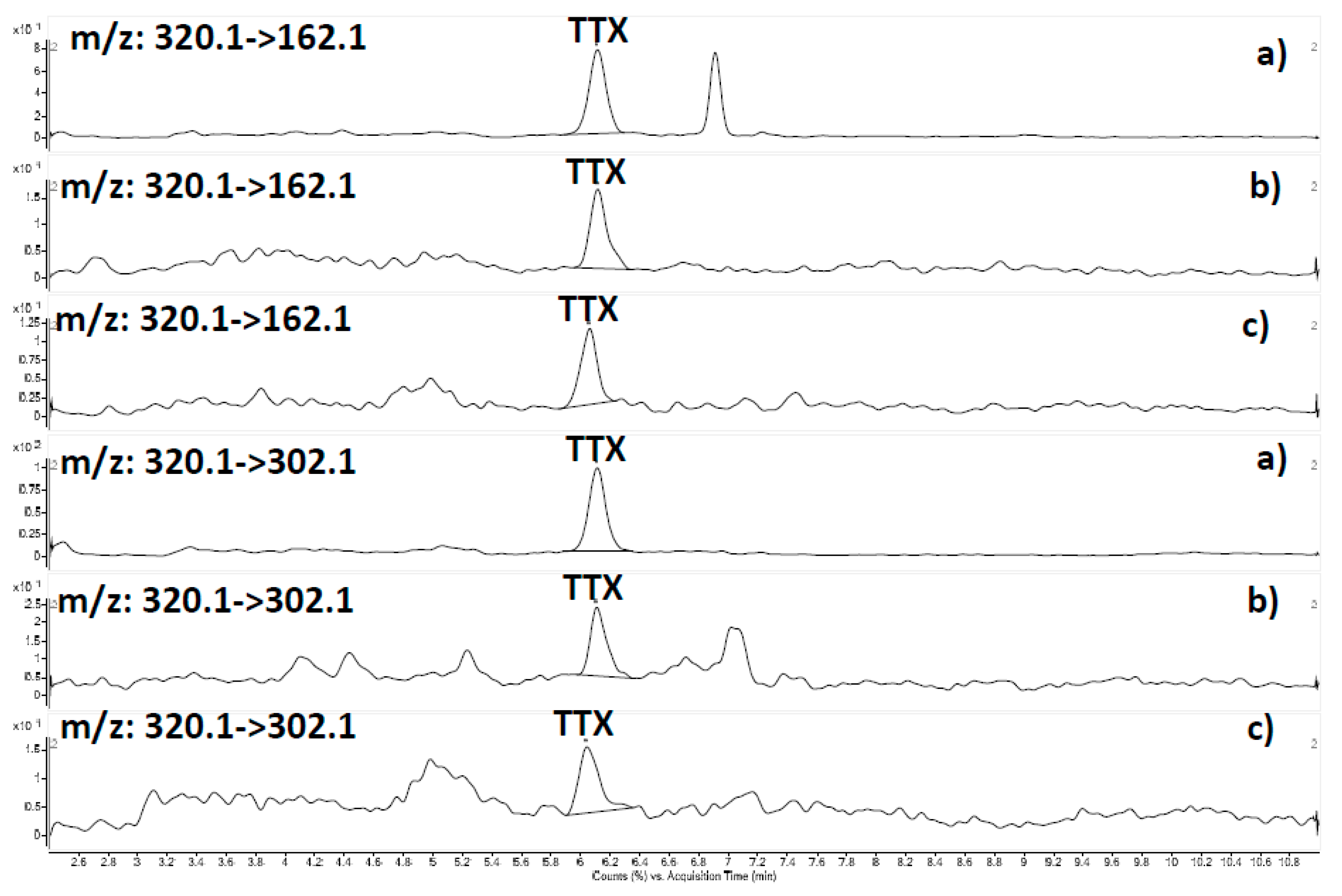
2.3. HILIC-MS/MS Results
3. Discussion
4. Materials and Methods
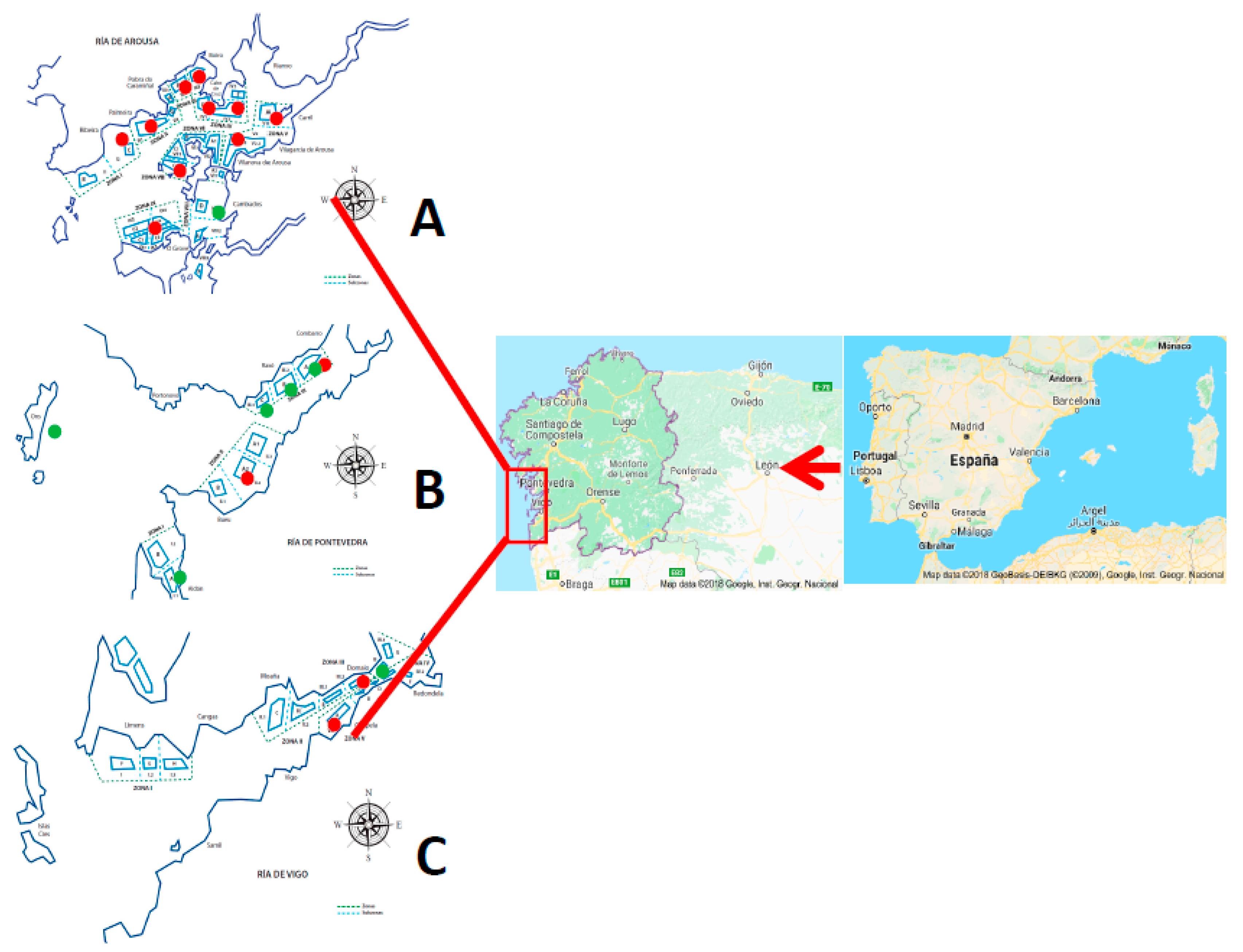
4.1. Sampling Plan
4.2. Microbiological Analysis
4.3. Investigation of PKS-NRPS Gene Fragments
4.4. N2a Assay
4.5. LC-MS/MS Analysis
4.5.1. Extraction
4.5.2. SPE-ENVI-Carb Clean-Up
4.5.3. HILIC-LC-MS/MS Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Isbister, G.K.; Kiernan, M.C. Neurotoxic marine poisoning. Lancet Neurol. 2005, 4, 219–228. [Google Scholar] [CrossRef]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origin and biosynthesis of tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Kogure, K.; Tamplin, M.L.; Simidu, U.; Colwell, R.R. A tissue culture assay for tetrodotoxin, saxitoxin and related toxins. Toxicon 1988, 26, 191–197. [Google Scholar] [CrossRef]
- Lipkind, G.M.; Fozzard, H.A. A Structural model of the tetrodotoxin and saxitoxin binding site of the Na+ channel. Biophys. J. 1994, 66, 1–13. [Google Scholar] [CrossRef]
- Lipkind, G.M.; Fozzard, H.A. KcsA Crystal Structure as Framework for a Molecular Model of the Na+ Channel Pore. Biochemistry 2000, 39, 8161–8170. [Google Scholar] [CrossRef] [PubMed]
- Tikhonov, D.B.; Zhorov, B.S. Sodium channel activators: Model of binding inside the pore and a possible mechanism of action. FEBS Lett. 2005, 579, 4207–4212. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain. Scientific opinion on the risks for public health related to the presence of tetrodotoxin (TTX) and TTX analogues in marine bivalves and gastropods. EFSA J. 2017, 15, 4752. [Google Scholar] [CrossRef]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves. Euro Surveill. 2015, 20, 21009. [Google Scholar] [CrossRef] [PubMed]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L. First detection of tetrodotoxin in Greek Shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [PubMed]
- Gerssen, A.; RIKILT Wageningen University and Research, Wageningen, The Netherlands. Personal communication, 2016.
- Pratheepa, V.; Alex, A.; Silva, M.; Vasconcelos, V. Bacterial diversity and tetrodotoxin analysis in the viscera of the gastropods from Portuguese coast. Toxicon 2016, 19, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.; Alfonso, A.; Alonso, E.; Rubiolo, J.; Roel, M.; Vlamis, A.; Katikou, P.; Jackson, S.; Lekha Menon, M.; Dobson, A.; et al. The association of bacterial C9-based TTX-like compounds with Prorocentrum minimum opens new uncertainties about shellfish seafood safety. Sci. Rep. 2017, 7, 40880. [Google Scholar] [CrossRef] [PubMed]
- Kotaki, Y.; Shimizu, Y. 1-Hydroxy-55, 11-dideoxytetrodotoxin, the first N-hydroxy and ring-deoxy derivative of tetrodotoxin found in the newt Taricha granulose. J. Am. Chem. Soc. 1993, 115, 827–830. [Google Scholar] [CrossRef]
- Liu, J.; Wei, F.; Lu, Y.; Ma, T.; Zhao, J.; Gong, X. Production level of tetradotoxin in Aeromonas is associated with the copy number of a plasmid. Toxicon 2015, 101, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.C.; Liub, C. Vibrio parahaemolyticus: A concern of seafood safety. Food Microbiol. 2007, 24, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Leon, A.; Torres, J.; Osorio, C.R.; Martinez-Urtaza, J. Identification of tdh-positive Vibrio parahaemolyticus from an outbreak associated with raw oyster consumption in Spain. FEMS Microbiol. Lett. 2003, 226, 281–284. [Google Scholar] [CrossRef]
- Martinez-Urtaza, J.; Lozano-Leon, A.; Varela-Pet, J.; Trinanes, J.; Pazos, Y.; García-Martín, O. Environmental Determinants of the Occurrence and Distribution of Vibrio parahaemolyticus in the Rias of Galicia, Spain. Appl. Environ. Microbiol. 2008, 74, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Leon, A. Estudio de Vibrio parahaemolyticus en Galicia. Aspectos Clínicos, Moleculares y Ecológicos. Ph.D. Thesis, Universidad de Santiago de Compostela, La Coruña, Spain, 2004. [Google Scholar]
- Martínez-Urtaza, J.; Blanco-Abad, V.; Rodriguez-Castro, A.; Ansede-Bermejo, J.; Miranda, A.; Rodriguez-Alvarez, M.X. Ecological determinants of the ocurrence and dynamics of Vibrio parahaemolyticus in offshore áreas. ISME J. 2012, 6, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of sodium channel toxins: Directed cytotoxicity assays for purified ciguatoxins, brevetoxins, saxitoxins and seafood extracts. J. AOAC Int. 1995, 78, 521–527. [Google Scholar] [PubMed]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: Semiautomated assay for Saxitoxins, Brevetoxins and Ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.Y.; Jang, J.H.; Cho, Y.; Konoki, K. Optimization of simultaneous analysis of tetrodotoxin, 4-epitetrodotoxin, 4,9-anhidrotetrodotoxin, and 5,6,11-trideoxytetrodotoxin by hydrophilic interaction liquid chromatography-tandem mass spectrometry. Forensic Toxicol. 2010, 29, 61–64. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Abe, Y.; Kudo, Y.; Ritson-Williams, R.; Paul, V.J.; Konoki, K.; Cho, Y.; Adachi, M.; Imazu, T.; Nishikawa, T.; et al. First Identification of 5,11-Dideoxytetrodotoxin in Marine Animals, and Characterization of Major Fragment Ions of tetrodotoxin and Its Analogs by High Resolution ESI-MS/MS. Mar. Drugs 2013, 11, 2799–2813. [Google Scholar] [CrossRef] [PubMed]
- EURLMB TTX SOP. Determination of Tetrodotoxin by HILIC-MS/MS; European Union Reference Laboratory for Marine Biotoxins: Vigo, Spain, 2017; Available online: http://www.eurlmb.org (accessed on 29 June 2017).
- Turner, A.D.; Dhanji-Rapkova, M.; Coates, L.; Bickerstaff, L.; Milligam, S.; O’Neill, A.; Faulkner, D.; McEneny, H.; Baker-Austin, C.; Lees, D.N.; et al. Detection of Tetrodotoxin Shellfish Poisoning (TSP) Toxins and Causative Factors in Bivalve Molluscs from the UK. Mar. Drugs 2017, 15, 277. [Google Scholar] [CrossRef]
- Tambadou, F.; Lanneluc, I.; Sablé, S.; Klein, G.L.; Doghri, I.; Sopéna, V.; Didelot, S.; Barthélémy, C.; Thiéry, V.; Chevrot, R. Novel nonribosomal peptide synthetase (NRPS) genes sequenced from intertidal mudflat bacteria. FEMS Microbiol. Lett. 2014, 357, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, M.C.; Neilan, B.A. On the presence of peptide synthetase and polyketide synthase genes in the cyanobacterial genus Nodularia. FEMS Microbiol. Lett. 2001, 196, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.; McNabb, P.; Harwood, T.; Selwood, A.; Boundy, M. Single-Laboratory Validation of a Multitoxin Ultra-Performance LC-Hydrophilic Interaction LC-MS/MS Method for Quantitation of Paralytic Shellfish Toxins in Bivalve Shellfish. J. AOAC Int. 2015, 98, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Boundy, M.J.; Selwood, A.I.; Harwood, D.T.; McNabb, P.S.; Turner, A.D. Development of a sensitive and selective liquid chromatography-mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitized carbon solid phase extraction. J. Chromatogr. A 2015, 1387, 1–12. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Rias | Samples | |||
|---|---|---|---|---|
| Number Tested | With Vibrio spp. (%) | Pelagic * | Infaunal ** | |
| Arosa | 866 | 20 | 171 | 2 |
| Pontevedra | 109 | 37 | 17 | 23 |
| Vigo | 304 | 24 | 70 | 3 |
| Time (min) | A (%) | B (%) | Flow Rate (mL/min) |
|---|---|---|---|
| 0.0 | 2.0 | 98.0 | 0.4 |
| 5.0 | 2.0 | 98.0 | 0.4 |
| 7.5 | 50.0 | 50.0 | 0.4 |
| 9.0 | 50.0 | 50.0 | 0.5 |
| 9.5 | 5.0 | 95.0 | 0.5 |
| 9.8 | 2.0 | 98.0 | 0.8 |
| 10.6 | 2.0 | 98.0 | 0.8 |
| Compound | Precursor Ion | Product Ion | Collision Energy (V) |
|---|---|---|---|
| TTX/4-epi-TTX | 320.1 | 302.1 | 28 |
| TTX/4-epi-TTX | 320.1 | 162.1 | 44 |
| 11-deoxy-TTX/5-deoxy-TTX | 304.1 | 286.1 | 28 |
| 11-deoxy-TTX/5-deoxy-TTX | 304.1 | 162.1 | 44 |
| 4,9-Anhydro TTX | 302.1 | 284.1 | 28 |
| 4,9-Anhydro TTX | 302.1 | 162.1 | 44 |
| 6,11-dideoxy-TTX | 290.1 | 272.1 | 28 |
| 6,11-dideoxy-TTX | 290.1 | 162.1 | 44 |
| 5,6,11-trideoxy-TTX | 272.1 | 254.1 | 28 |
| 5,6,11-trideoxy-TTX | 272.1 | 162.1 | 44 |
| Source Parameters | |||
|---|---|---|---|
| Gas temp (°C) | 150 | Polarity | Positive |
| Gas flow (L/min) | 12 | Fragmentor (V) | 380 |
| Nebulizer (psi) | 45 | Cell accelerator (V) | 5 |
| Sheath gas heater (°C) | 400 | Dwell (ms) | 20 |
| Sheath gas flow (L/min) | 12 | Delta EMV (V) | 400 |
| Capillary (V) | 4000 | ||
| V charging (V) | 300 | ||
| Ion Funnel Parameters (V) | |||
| Pos high pressure RF | 150 | Neg high pressure RF | 90 |
| Pos low pressure RF | 60 | Neg low pressure RF | 60 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leão, J.M.; Lozano-Leon, A.; Giráldez, J.; Vilariño, Ó.; Gago-Martínez, A. Preliminary Results on the Evaluation of the Occurrence of Tetrodotoxin Associated to Marine Vibrio spp. in Bivalves from the Galician Rias (Northwest of Spain). Mar. Drugs 2018, 16, 81. https://doi.org/10.3390/md16030081
Leão JM, Lozano-Leon A, Giráldez J, Vilariño Ó, Gago-Martínez A. Preliminary Results on the Evaluation of the Occurrence of Tetrodotoxin Associated to Marine Vibrio spp. in Bivalves from the Galician Rias (Northwest of Spain). Marine Drugs. 2018; 16(3):81. https://doi.org/10.3390/md16030081
Chicago/Turabian StyleLeão, Jose Manuel, Antonio Lozano-Leon, Jorge Giráldez, Óscar Vilariño, and Ana Gago-Martínez. 2018. "Preliminary Results on the Evaluation of the Occurrence of Tetrodotoxin Associated to Marine Vibrio spp. in Bivalves from the Galician Rias (Northwest of Spain)" Marine Drugs 16, no. 3: 81. https://doi.org/10.3390/md16030081