Anti-Inflammatory Effects of a Mytilus coruscus α-d-Glucan (MP-A) in Activated Macrophage Cells via TLR4/NF-κB/MAPK Pathway Inhibition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. THP-1 Cell Culture
2.3. Cell Viability Assay
2.4. Determination of Phagocytic Uptake of FITC-Labeled Dextran
2.5. ELISA for Cytokine Estimation
2.6. Nitric Oxide (NO) Production
2.7. Western Blot Analyses
2.8. Detection of NF-κB Activation and Nuclear Translocation
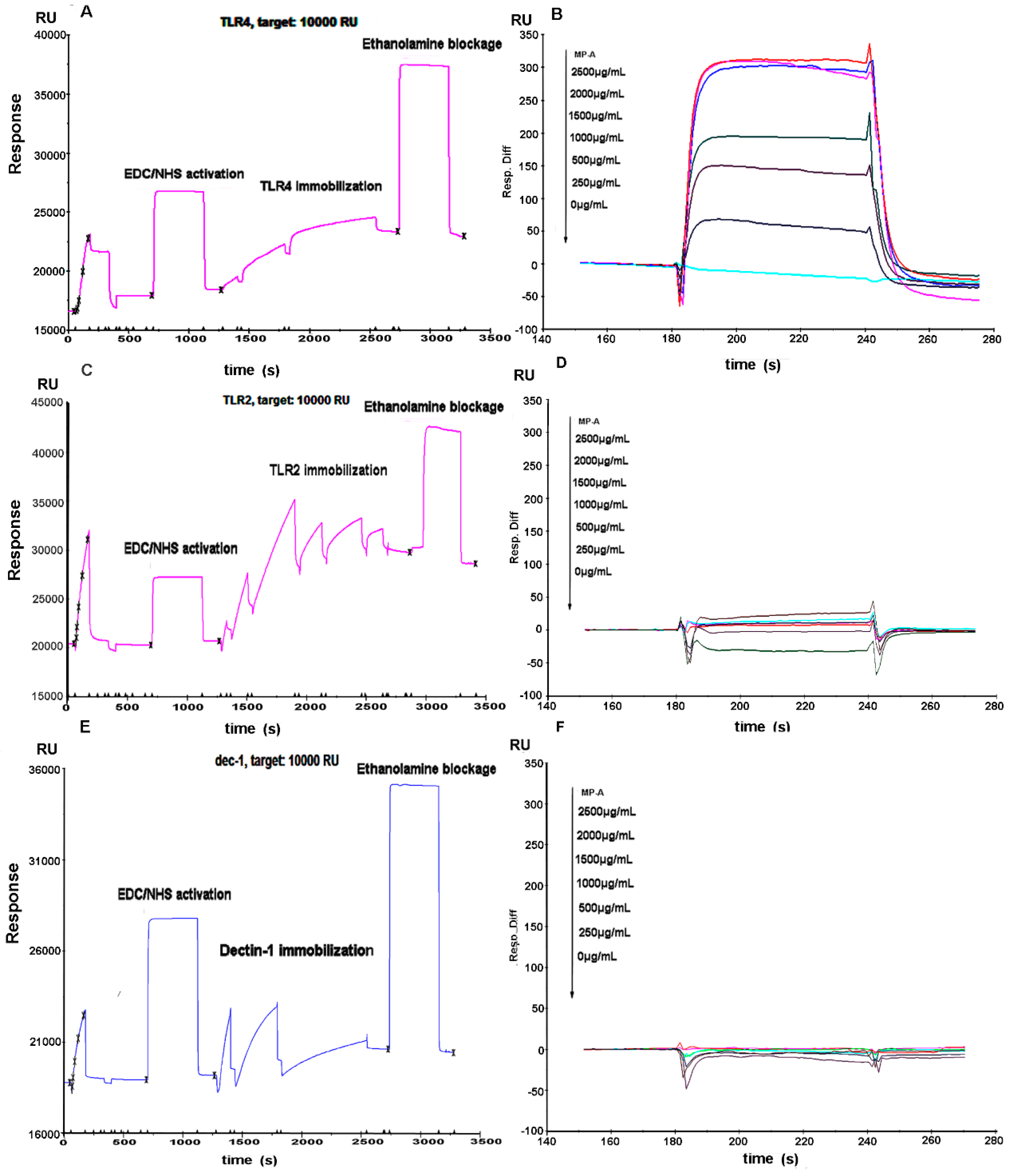
2.9. Affinity Assay for MP-A with TLR4, TLR2 and Dectin-1
2.10. Statistical Analyses
3. Results
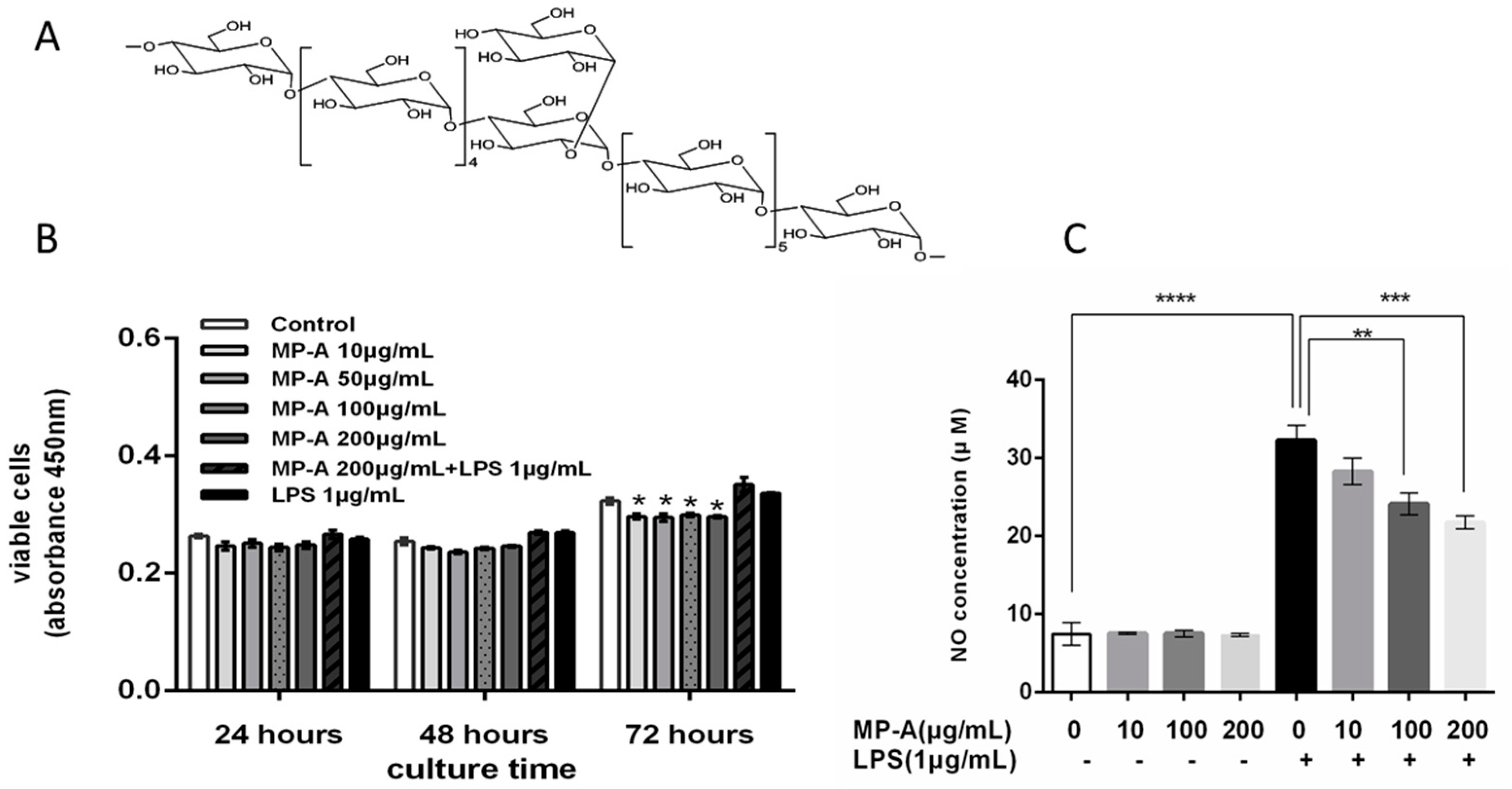
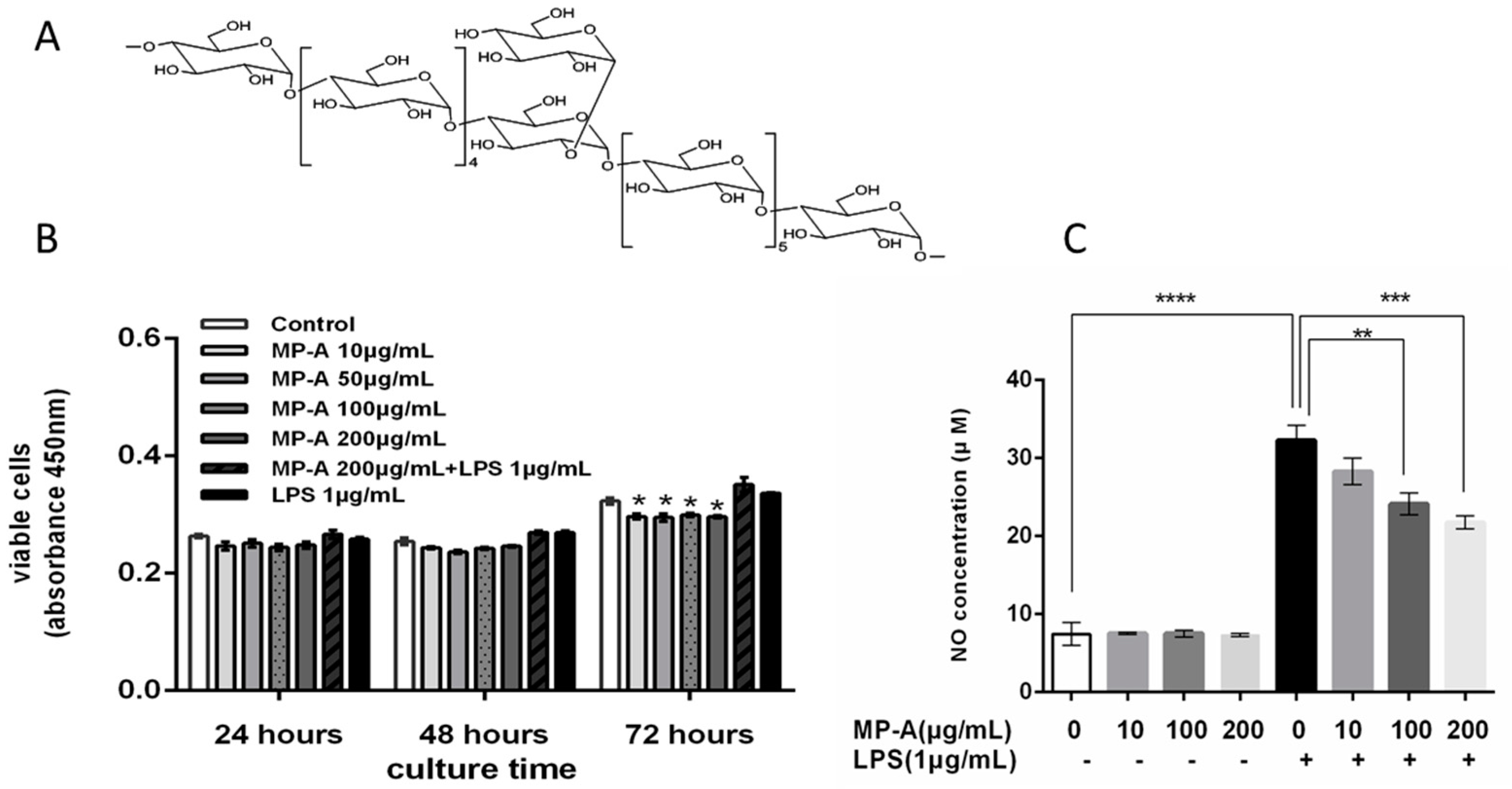
3.1. Effects of MP-A on THP-1 Cell Viability
3.2. Effect of MP-A on NO Release by THP-1 Cells
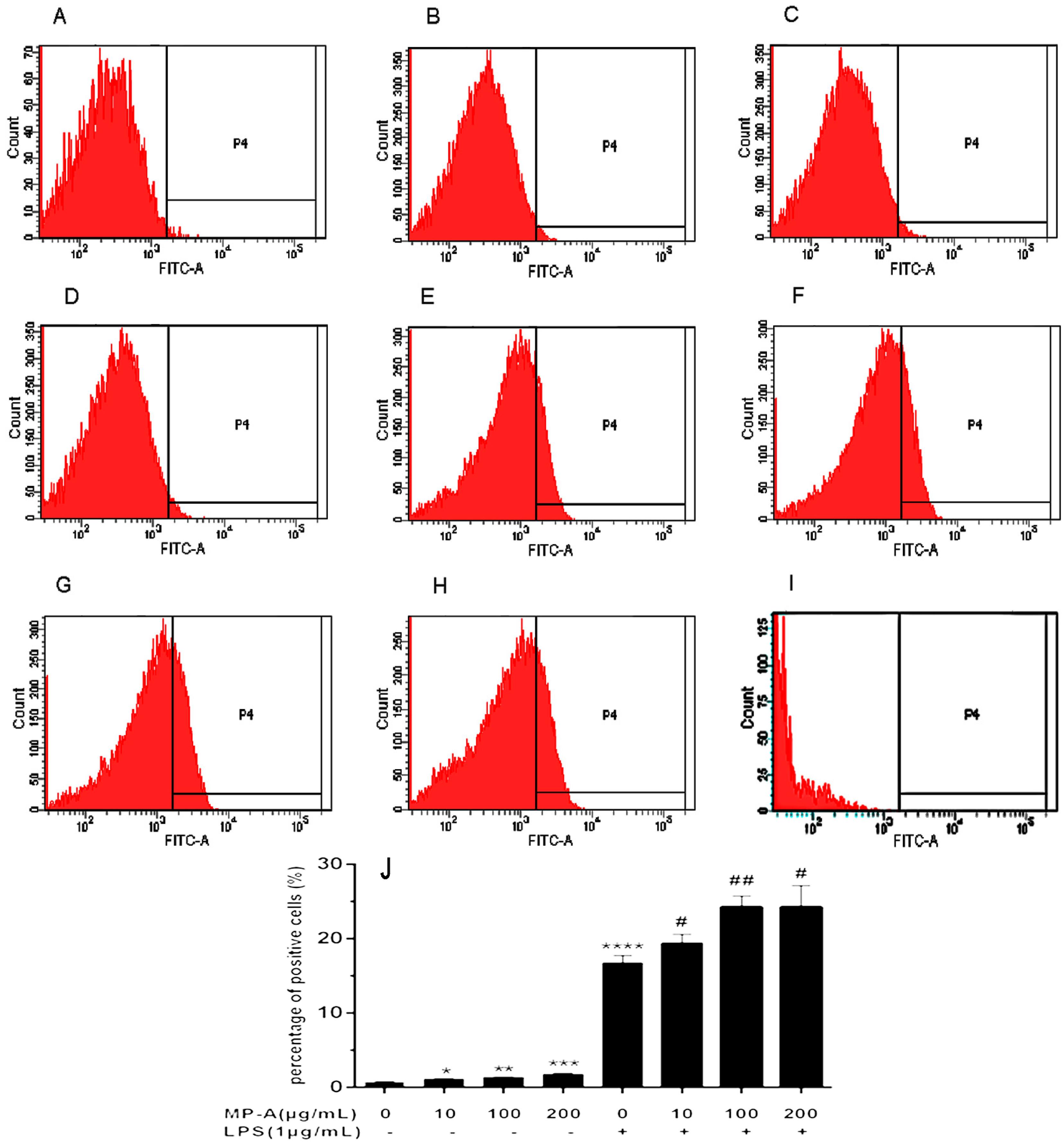
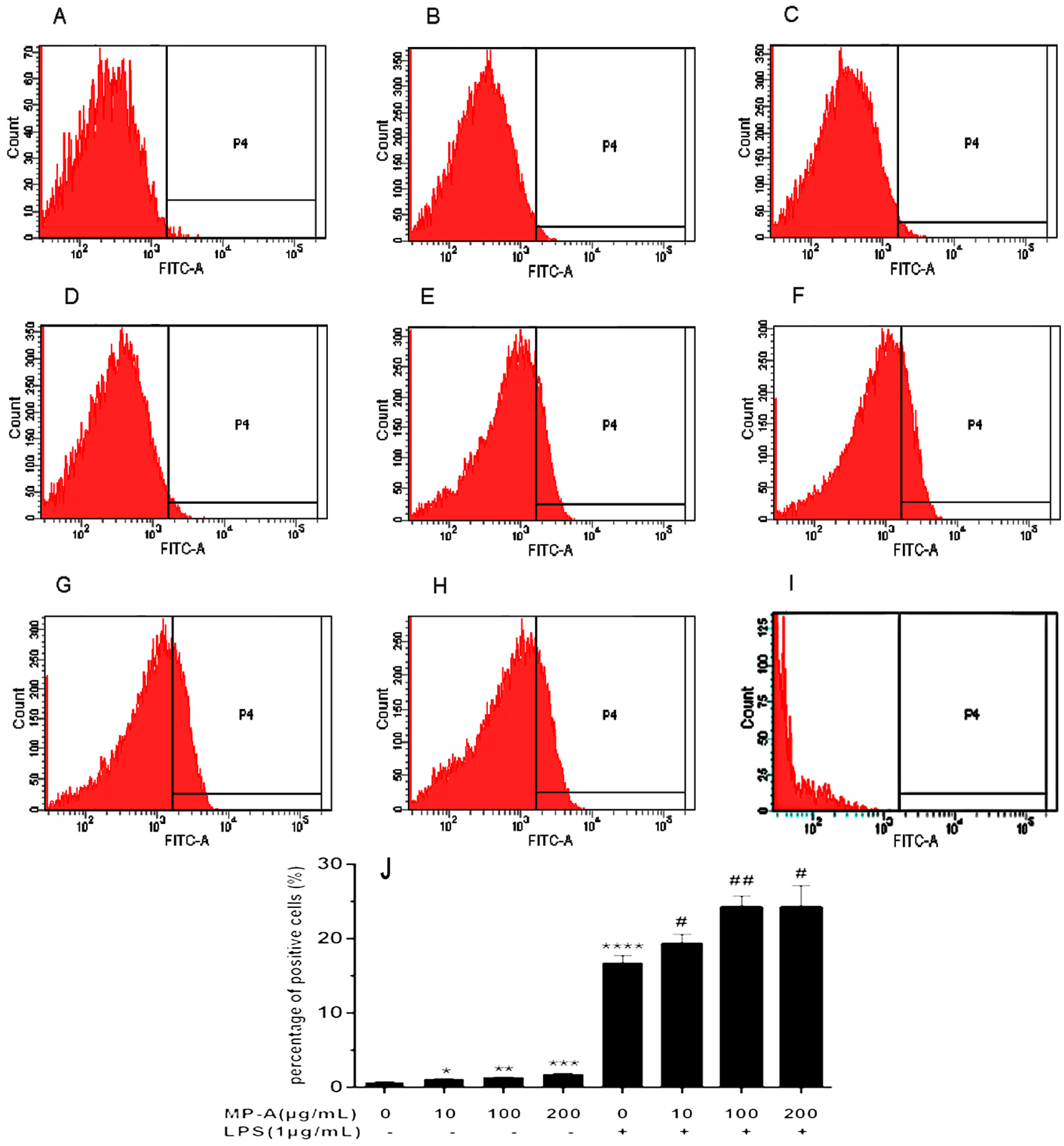
3.3. Effect of MP-A on Phagocytic Uptake of FITC-Labeled Dextran by THP-1 Cells
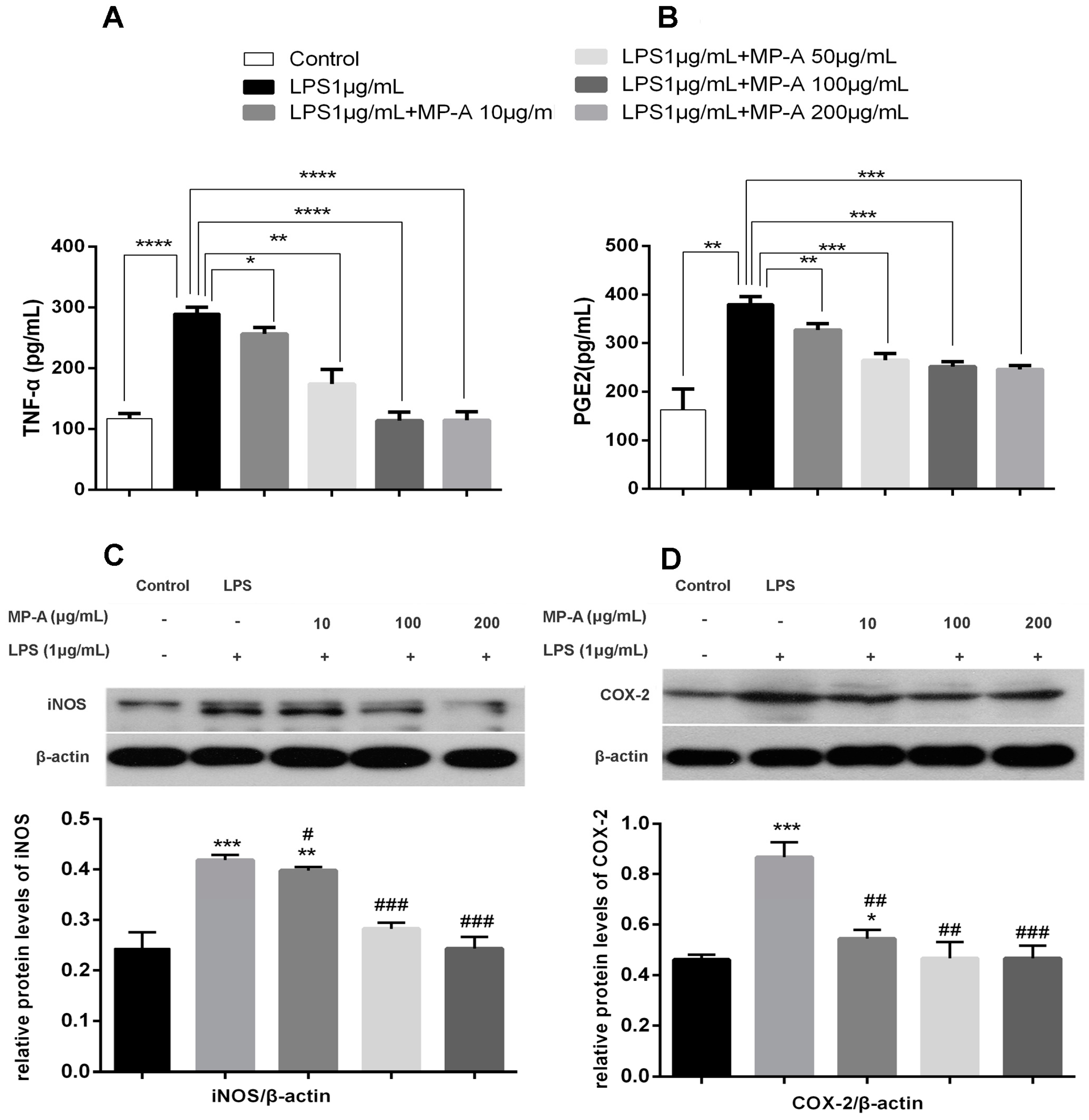
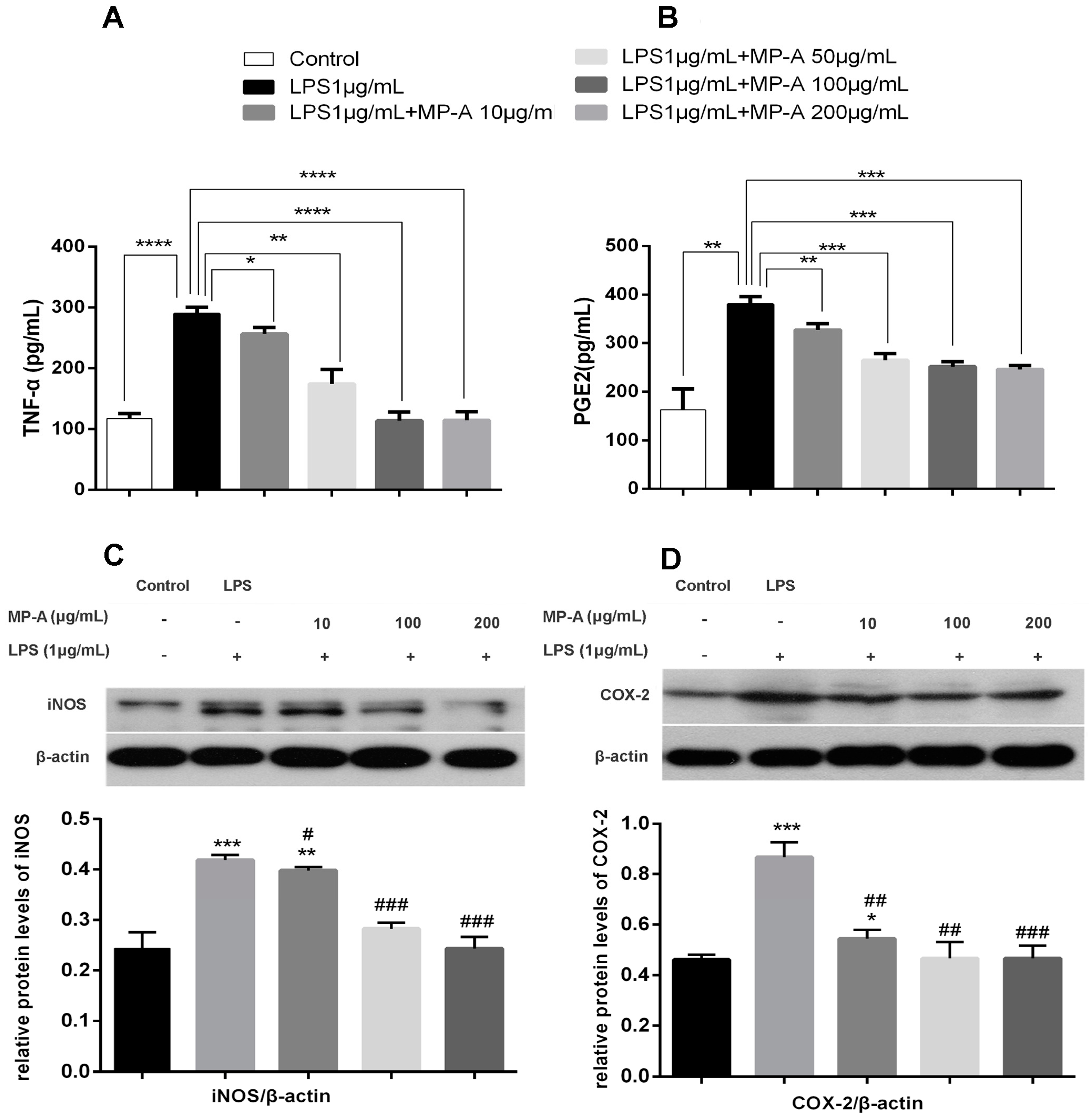
3.4. MP-A Inhibition of TNF-α and PGE2 Release by LPS-Stimulated THP-1 Cells
3.5. MP-A Inhibits iNOS and COX-2 Expression in LPS-Stimulated THP-1 Cells
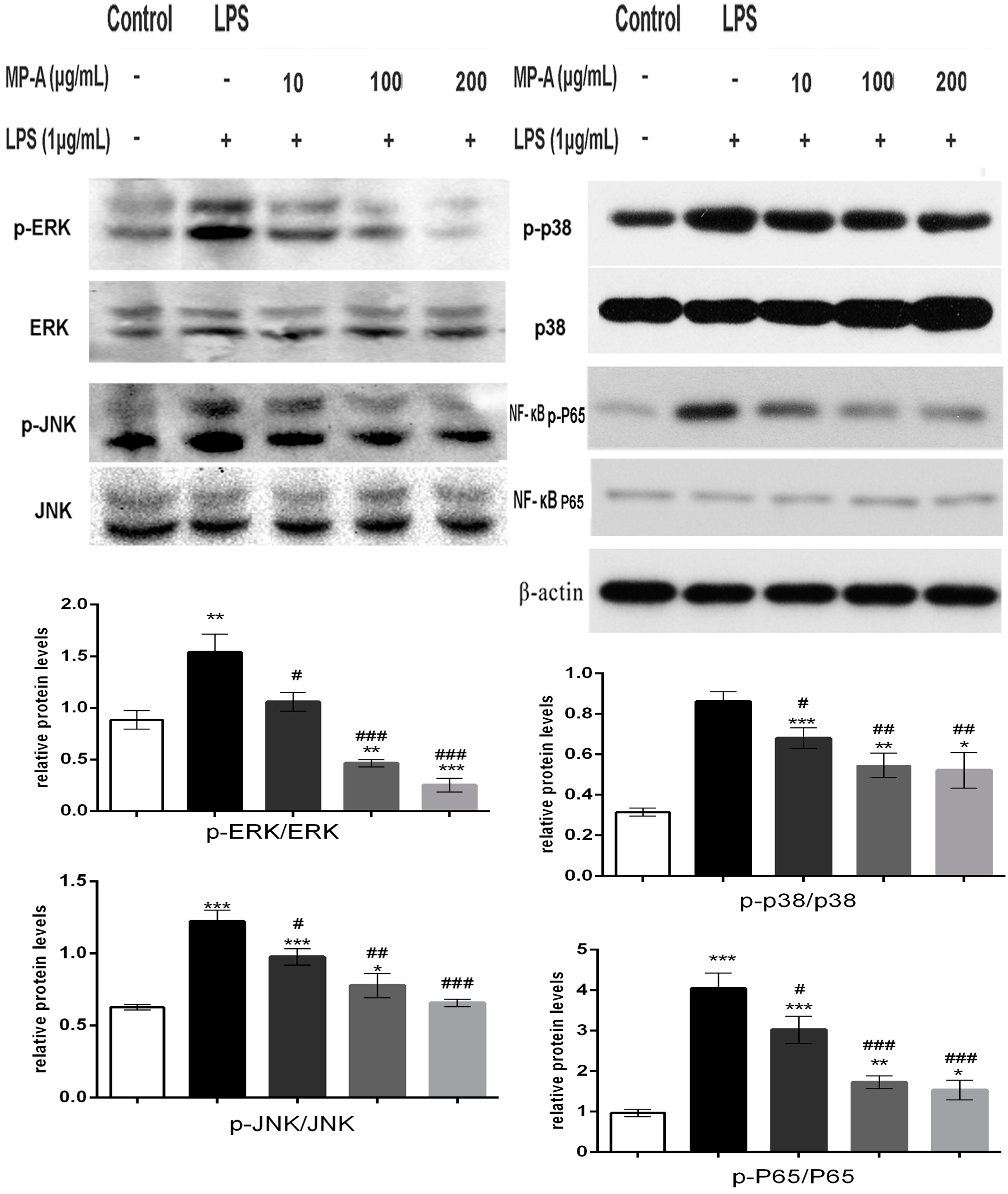
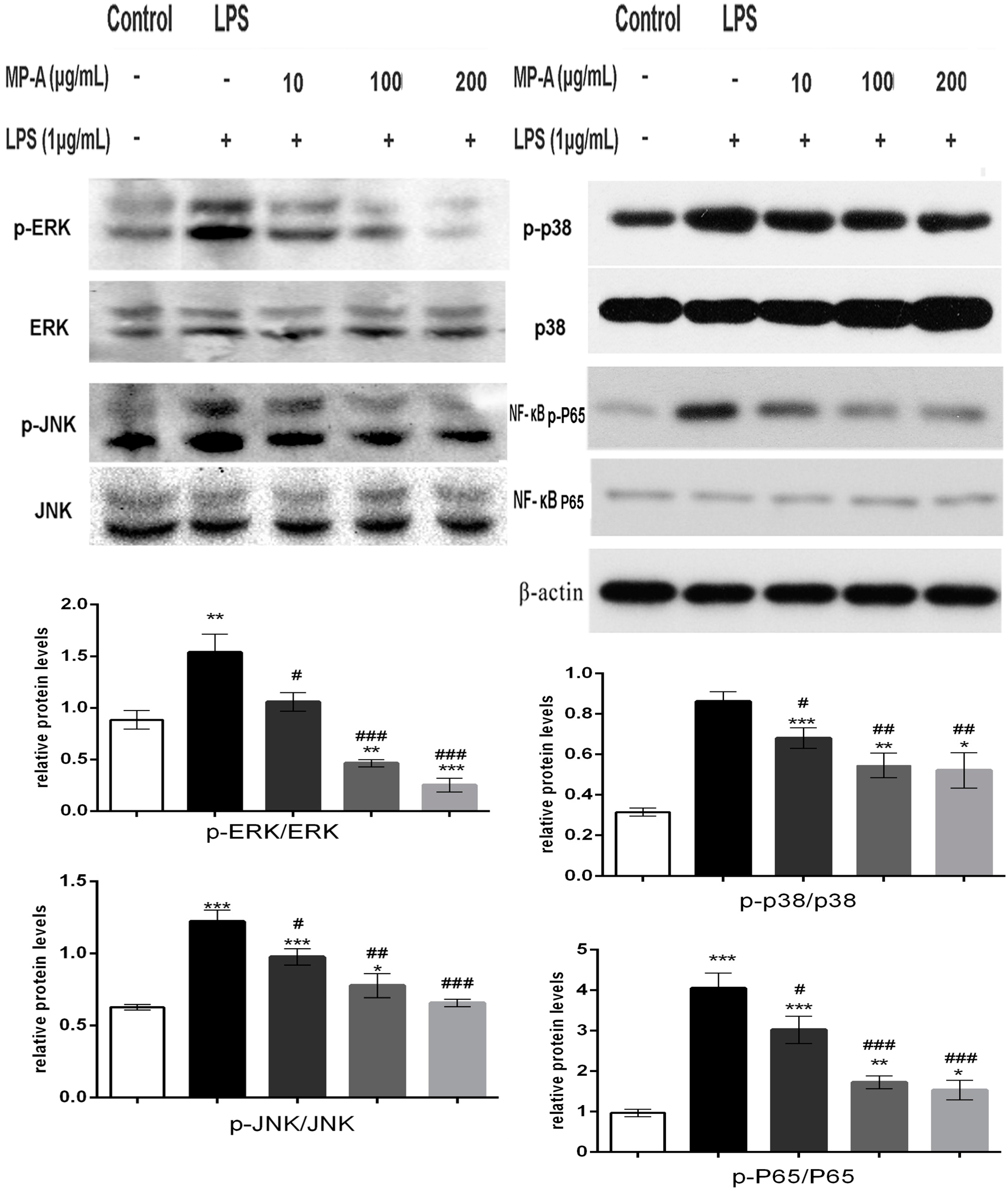
3.6. MP-A Treatment Inhibits LPS-Induced Phosphorylation of MAPKs and NF-κB in THP-1 Cells
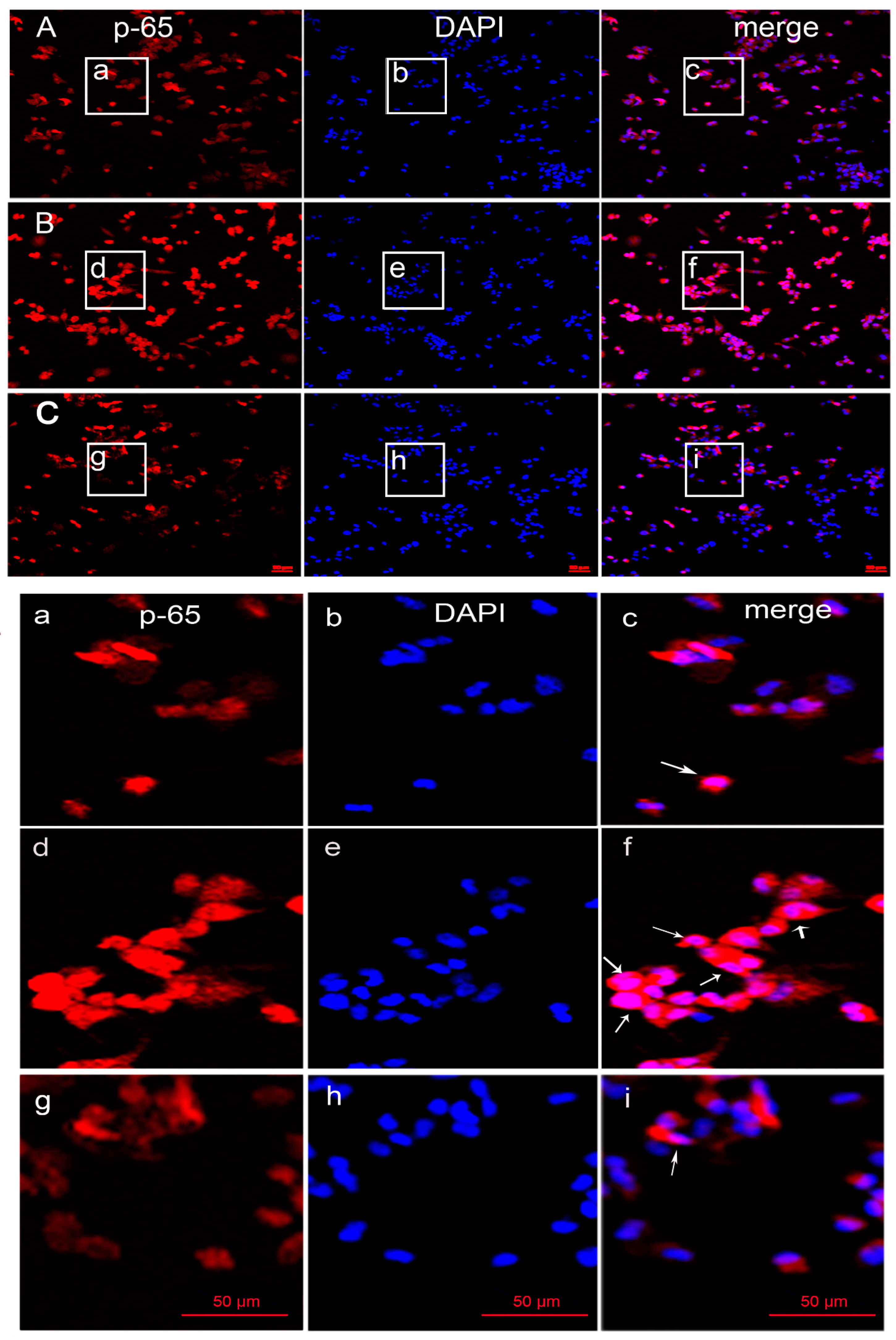
3.7. MP-A Prevented LPS-Induced Activation and the Nuclear Translocation of NF-κB in THP-1 Cells
3.8. Affinity of MP-A for Recombinant Human TLR4, TLR2, and Dectin-1
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| LPS | lipopolysaccharide |
| TLR4 | Toll-like receptor 4 |
| TLR2 | Toll-like receptor 2 |
| TLRs | toll like receptors |
| TNF-α | tumor necrosis factor α |
| iNOS | inducible nitric oxide synthase |
| COX-2 | cyclooxygenase-2 |
| PGE2 | prostaglandin E2 |
| SPR | surface plasmon resonance |
| CCK-8 | Cell Counting Kit-8 |
| PMA | phorbol-12-myristate-13-acetate |
| DAPI | 2-(4-amidinophenyl)-6-indolecarbamidine dihydrochloride |
| MAPK | mitogen-activated protein kinase |
| ERK | extracellular signal regulated kinase |
| JNK | c-Jun N terminal kinase |
| NF-κB | nuclear factor-kappa B |
References
- Xu, H.; Guo, T.; Guo, Y.F.; Zhang, J.; Li, Y.; Feng, W.; Jiao, B. Characterization and protection on acute liver injury of a polysaccharide mp-i from mytilus coruscus. Glycobiology 2008, 18, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-K.; Oh, H.-J.; Kim, Y.-S.; Hwang, J.-W.; Ahn, C.-B.; Lee, J.S.; Jeon, Y.-J.; Moon, S.-H.; Sung, S.H.; Jeon, B.-T.; et al. Purification of a novel peptide derived from mytilus coruscus and in vitro/in vivo evaluation of its bioactive properties. Fish Shellfish Immunol. 2013, 34, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Ahn, C.B.; Je, J.Y. Anti-inflammatory action of high molecular weight mytilus edulis hydrolysates fraction in lps-induced raw264.7 macrophage via nf-kappab and mapk pathways. Food Chem. 2016, 202, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fu, Y.; Zheng, J.; Li, D. Anti-inflammatory activity and mechanism of a lipid extract from hard-shelled mussel (mytilus coruscus) on chronic arthritis in rats. Mar. Drugs 2014, 12, 568–588. [Google Scholar] [CrossRef] [PubMed]
- Tzianabos, A.O. Polysaccharide immunomodulators as therapeutic agents: Structural aspects and biologic function. Clin. Microbiol. Rev. 2000, 13, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Cavaillon, J.M. Cytokines and macrophages. Biomed. Pharmacother. 1994, 48, 445–453. [Google Scholar] [CrossRef]
- Miller, T.E.; Ormrod, D.J.; Geddes, R. Anti-inflammatory activity of glycogen extracted from perna canaliculus (nz green-lipped mussel). Agents Actions 1993, 38, 139–142. [Google Scholar] [CrossRef]
- Cesaretti, M.; Luppi, E.; Maccari, F.; Volpi, N. Isolation and characterization of a heparin with high anticoagulant activity from the clam tapes phylippinarum: Evidence for the presence of a high content of antithrombin iii binding site. Glycobiology 2004, 14, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.; Xin, T.; Chu, Z.Y.; Wang, J.C.; Zhang, S.; Shen, Y. Immunomodulation of mytihus edulis linnaeus polysaccharides in mice. Chin. J. Mar. Drugs 2010, 29, 39–41. [Google Scholar]
- Mao, J.; Hou, Z.; Chen, D.; Zhang, W. Anti-hyperlipi-demic effects of polysaccharide ma from mytilus coruscus on experimental atherosclerosis in rabitts. J. Pharm. Pract. 2014, 32, 27–30. [Google Scholar]
- Czop, J.K. The role of beta-glucan receptors on blood and tissue leukocytes in phagocytosis and metabolic activation. Pathol. Immunopathol. Res. 1986, 5, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.L. Overview of (1→3)-β-d-glucan immunobiology. Mediat. Inflamm. 1997, 6, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Ross, G.D.; Vetvicka, V.; Yan, J.; Xia, Y.; Vetvickova, J. Therapeutic intervention with complement and beta-glucan in cancer. Immunopharmacology 1999, 42, 61–74. [Google Scholar] [CrossRef]
- Muller, A.; Rice, P.J.; Ensley, H.E.; Coogan, P.S.; Kalbfleish, J.H.; Kelley, J.L.; Love, E.J.; Portera, C.A.; Ha, T.; Browder, L.W.; et al. Receptor binding and internalization of a water-soluble (1→3)-β-d-glucan biologic response modifier in two monocyte/macrophage cell lines. J. Immunol. 1996, 156, 3418–3425. [Google Scholar] [PubMed]
- Xu, X.; Yasuda, M.; Nakamura-Tsuruta, S.; Mizuno, M.; Ashida, H. Beta-glucan from lentinus edodes inhibits nitric oxide and tumor necrosis factor-alpha production and phosphorylation of mitogen-activated protein kinases in lipopolysaccharide-stimulated murine raw 264.7 macrophages. J. Biol. Chem. 2012, 287, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.B.; Gao, X.D.; Han, F.; Xu, B.S.; Song, Y.C.; Tan, R.X. Purification, characterization and enzymatic degradation of ycp, a polysaccharide from marine filamentous fungus phoma herbarum ys4108. Biochimie 2005, 87, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Zhang, X.; Liu, W.; Zhou, Y.; Ding, R.; Yao, W.; Gao, X. Preparation and immunomodulating activities of a library of low-molecular-weight alpha-glucans. Carbohydr. Polym. 2014, 111, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.J.; Wichers, H.J. Thp-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B.; Rietschel, E.T. Innate immune sensing and its roots: The story of endotoxin. Nat. Rev. Immunol. 2003, 3, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Soyoola, E.; Chanmugam, P.; Hart, S.; Sun, W.; Zhong, H.; Liou, S.; Simmons, D.; Hwang, D. Selective expression of mitogen-inducible cyclooxygenase in macrophages stimulated with lipopolysaccharide. J. Biol. Chem. 1992, 267, 25934–25938. [Google Scholar] [PubMed]
- Buchanan, M.M.; Hutchinson, M.; Watkins, L.; Yin, H. Toll-like receptor 4 in cns pathologies. J. Neurochem. 2010, 114, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Fujii, M.; Hou, D.-X. 6-(methylsulfinyl)hexyl isothiocyanate suppresses inducible nitric oxide synthase expression through the inhibition of janus kinase 2-mediated jnk pathway in lipopolysaccharide-activated murine macrophages. Biochem. Pharmacol. 2005, 70, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Zar, P.P.K.; Morishita, A.; Hashimoto, F.; Sakao, K.; Fujii, M.; Wada, K.; Hou, D.-X. Anti-inflammatory effects and molecular mechanisms of loquat (eriobotrya japonica) tea. J. Funct. Foods 2014, 6, 523–533. [Google Scholar] [CrossRef]
- Zhou, J.; Zhu, P.; Jiang, J.L.; Zhang, Q.; Wu, Z.B.; Yao, X.Y.; Tang, H.; Lu, N.; Yang, Y.; Chen, Z.N. Involvement of cd147 in overexpression of mmp-2 and mmp-9 and enhancement of invasive potential of pma-differentiated thp-1. BMC Cell Biol. 2005, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, X.; Pang, X.; Zhang, W.; Wu, W.; Zhao, J.; Yang, H.; Qu, W. Effects of zinc and manganese on advanced glycation end products (ages) formation and ages-mediated endothelial cell dysfunction. Life Sci. 2012, 90, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Lin, S.; Wu, W.; Tan, H.; Wang, Z.; Cheng, C.; Lu, L.; Zhang, X. Ghrelin prevents doxorubicin-induced cardiotoxicity through tnf-alpha/nf-kappab pathways and mitochondrial protective mechanisms. Toxicology 2008, 247, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Wonderling, R.S.; Ghaffar, A.; Mayer, E.P. Lipopolysaccharide-induced suppression of phagocytosis: Effects on the phagocytic machinery. Immunopharmacol. Immunotoxicol. 1996, 18, 267–289. [Google Scholar] [CrossRef] [PubMed]
- De Lima, T.M.; Sampaio, S.C.; Petroni, R.; Brigatte, P.; Velasco, I.T.; Soriano, F.G. Phagocytic activity of lps tolerant macrophages. Mol. Immunol. 2014, 60, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, X.; Cheng, Y.; Li, J.; Xiao, Y.; Zhang, Q.; Zong, A.; Zhong, C.; Wang, F. Preparation and in vitro immunomodulatory effect of curdlan sulfate. Carbohydr. Polym. 2014, 102, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, X.; Ling, P.; Liao, J.; Zhao, M.; Mei, L.; Shao, H.; Jiang, P.; Song, Z.; Chen, Q.; et al. Immunomodulatory effects of xanthan gum in lps-stimulated raw 264.7 macrophages. Carbohydr. Polym. 2017, 169, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.L.; Martin, E.; Turko, I.V.; Murad, F. Novel effects of nitric oxide. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 203–236. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C. Nitric oxide as a secretory product of mammalian cells. FASEB J. 1992, 6, 3051–3064. [Google Scholar] [PubMed]
- Ohno, N.; Hashimoto, T.; Adachi, Y.; Yadomae, T. Corrigendum to: “Conformation dependency of nitric oxide synthesis of murine peritoneal macrophages by β-glucans in vitro” [immunol. Lett. 52 (1996) 1–7]. Immunol. Lett. 1996, 53, 157–163. [Google Scholar] [CrossRef]
- Gupta, S.C.; Sundaram, C.; Reuter, S.; Aggarwal, B.B. Inhibiting nf-kappab activation by small molecules as a therapeutic strategy. Biochim. Biophys. Acta 2010, 1799, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Iezzi, A.; Ferri, C.; Mezzetti, A.; Cipollone, F. Cox-2: Friend or foe? Curr. Pharm. Des. 2007, 13, 1715–1721. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Wang, G.; Zhu, X.; Shao, H.; Liu, F.; Yang, P.; Ying, Y.; Wang, F.; Ling, P. Preparation of xanthan gum injection and its protective effect on articular cartilage in the development of osteoarthritis. Carbohydr. Polym. 2012, 87, 1837–1842. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ko, H.M.; Kang, N.I.; Song, C.H.; Zhang, X.; Chung, W.C.; Kim, J.H.; Choi, I.H.; Park, Y.M.; Kim, G.Y. Mast cells play a key role in the developmentof late airway hyperresponsiveness through tnf-αin a murine model of asthma. Eur. J. Immunol. 2007, 37, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Roux, C.H.; Brocq, O.; Breuil, V.; Albert, C.; Euller-Ziegler, L. Safety of anti-tnf-alpha therapy in rheumatoid arthritis and spondylarthropathies with concurrent b or c chronic hepatitis. Rheumatology 2006, 45, 1294–1297. [Google Scholar] [CrossRef] [PubMed]
- Stokkers, P.; Camoglio, L.; Van Deventer, S. Tumor necrosis factor (tnf) in inflammatory bowel disease: Gene polymorphisms, animal models, and potential for anti-tnf therapy. J. Inflamm. 1994, 47, 97–103. [Google Scholar]
- Brown, G.D. Dectin-1: A signalling non-tlr pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Gantner, B.N.; Simmons, R.M.; Canavera, S.J.; Akira, S.; Underhill, D.M. Collaborative induction of inflammatory responses by dectin-1 and toll-like receptor 2. J. Exp. Med. 2003, 197, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, A.; Kamiryou, Y.; Yamada, H.; Eto, M.; Shibata, K.; Haruna, K.; Naito, S.; Yoshikai, Y. Oral administration of xanthan gum enhances antitumor activity through toll-like receptor 4. Int. Immunopharmacol. 2009, 9, 1562–1567. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, R.; Akashi, S.; Ogata, H.; Nagai, Y.; Fukudome, K.; Miyake, K.; Kimoto, M. Md-2, a molecule that confers lipopolysaccharide responsiveness on toll-like receptor 4. J. Exp. Med. 1999, 189, 1777–1782. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; Yamazaki, S.; Eto-Kimura, A.; Takeshige, K.; Muta, T. The amino-terminal region of toll-like receptor 4 is essential for binding to md-2 and receptor translocation to the cell surface. J. Biol. Chem. 2004, 279, 47431–47437. [Google Scholar] [CrossRef] [PubMed]
- Moynagh, P.N. The nf-κb pathway. J. Cell Sci. 2005, 118, 4589–4592. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. Nf-κb: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Zhang, X.; Li, Y.; Chen, Q.; Liu, F.; Zhu, X.; Mei, L.; Song, X.; Liu, X.; Song, Z.; et al. Anti-Inflammatory Effects of a Mytilus coruscus α-d-Glucan (MP-A) in Activated Macrophage Cells via TLR4/NF-κB/MAPK Pathway Inhibition. Mar. Drugs 2017, 15, 294. https://doi.org/10.3390/md15090294
Liu F, Zhang X, Li Y, Chen Q, Liu F, Zhu X, Mei L, Song X, Liu X, Song Z, et al. Anti-Inflammatory Effects of a Mytilus coruscus α-d-Glucan (MP-A) in Activated Macrophage Cells via TLR4/NF-κB/MAPK Pathway Inhibition. Marine Drugs. 2017; 15(9):294. https://doi.org/10.3390/md15090294
Chicago/Turabian StyleLiu, Fuyan, Xiaofeng Zhang, Yuqiu Li, Qixin Chen, Fei Liu, Xiqiang Zhu, Li Mei, Xinlei Song, Xia Liu, Zhigang Song, and et al. 2017. "Anti-Inflammatory Effects of a Mytilus coruscus α-d-Glucan (MP-A) in Activated Macrophage Cells via TLR4/NF-κB/MAPK Pathway Inhibition" Marine Drugs 15, no. 9: 294. https://doi.org/10.3390/md15090294