Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications



1
Department of Biology and Evolution of Marine Organisms, Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy
2
Department of Biology, University of Naples Federico II, Complesso Universitario di Monte Sant’Angelo, Via Cinthia, 80126 Napoli, Italy
3
Bio-Organic Chemistry Unit, Institute of Biomolecular Chemistry-CNR, Via Campi Flegrei 34, Pozzuoli, 80078 Naples, Italy
4
Unità di Farmacologia Sperimentale, Istituto Nazionale Tumori “Fondazione G. Pascale”, IRCCS, 80131 Napoli, Italy
5
Department of Integrative Marine Ecology, Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy
6
Institute of Biosciences and BioResources, CNR, 80131 Napoli, Italy
*
Author to whom correspondence should be addressed.
Mar. Drugs 2017, 15(6), 173; https://doi.org/10.3390/md15060173
Submission received: 5 May 2017
/
Revised: 6 June 2017
/
Accepted: 8 June 2017
/
Published: 12 June 2017
Abstract
:Enzymes play key roles in different cellular processes, for example, in signal transduction, cell differentiation and proliferation, metabolic processes, DNA damage repair, apoptosis, and response to stress. A deregulation of enzymes has been considered one of the first causes of several diseases, including cancers. In the last several years, enzyme inhibitors, being good candidates as drugs in the pathogenic processes, have received an increasing amount of attention for their potential application in pharmacology. The marine environment is considered a challenging source of enzyme inhibitors for pharmacological applications. In this review, we report on secondary metabolites with enzyme inhibitory activity, focusing our attention on marine sponges and bacteria as promising sources. In the case of sponges, we only reported the kinase inhibitors, because this class was the most representative isolated so far from these marine organisms.
1. An Introduction to Enzyme Inhibitors in Marine Environments
An important challenge of the last several decades has been the search for active compounds from natural sources, about 50% of which have pharmacological applications [1,2].
Among natural compounds, enzyme inhibitors have been considered useful tools mainly for their biotechnological potential in pharmacology [3] and agriculture [4]. In particular, protease inhibitors represent important examples of enzyme inhibitors, able to inactivate target proteases in the presence of human diseases (as for example in high blood pressure, arthritis, muscular dystrophy, pancreatitis, thrombosis, different cancers, as well as AIDS [5,6]). In the case of carbohydrate-dependent diseases, such as diabetes, obesity and hyperlipemia, amylase inhibitors represent useful tools for controlling them [7,8]. Enzyme inhibitors have been also isolated from different terrestrial organisms, including microorganisms (mainly actinomycetes; [9]), even if they are able to produce structurally identical inhibitors [10]. Differently, in the marine organisms occurs considerably different characteristics of enzyme inhibitors in comparison with the terrestrial ones [11].
In fact, the oceans (covering 70% of the earth) represent a widely unexplored and promising source of new biologically active natural compounds, also linked to the highest biodiversity of the marine environment in comparison with the terrestrial one [12,13,14,15,16]. Concerning the marine environment, it is important to consider that sessile marine invertebrates, such as sponges, cnidaria, and ascidians, constitute the largest amount of biomass of the marine macrofauna. They are able to store several secondary metabolites for their ecological success in different marine habitats so as to counterattack predation and pathogenic organisms [17,18,19,20]. These chemical defenses of sessile marine invertebrates not only have ecological roles but also exhibit several different biological activities with pharmacological, nutraceutical, and cosmaceutical applications [13,21,22,23].
Marine microorganisms isolated for the first time in the marine environment in the last century are another key source of active natural compounds. In more detail, marine microorganisms are very different from terrestrial ones, for their metabolic and physiological processes, because about 90% of them are Gram-negative psychrophiles that also require high Na+ concentration for growth [24,25]. Maeda and Taga [25] isolated a deoxyribonuclease from a marine Vibrio sp. and studied its activity, demonstrating that this enzyme was activated by Mg2+ and stabilized by Ca2+. Moreover, another marine bacterium has been isolated, able to produce a phosphatase with its activity at a high level of hydrostatic pressure (1000 atm; [26]), suggesting that marine enzymes acted in a different way in comparison to terrestrial ones. Moreover, many enzymes have significant roles in the maintenance of homeostasis and diseases are the results of the breakdown of homeostasis. In fact, there is a very strong correlation between the biological functions of several enzymes and diseases. Enzyme inhibitors are molecules that reduce the catalytic activity or the complete blocking of enzymes, thus causing either the complete cell death or modification in the pathways. On this line, some enzyme inhibitors correlated with specific diseases are very important as drugs [27]. For example, the angiotensin-converting enzyme (ACE) inhibitors act as antihypertensive drugs, reducing hypertension. Adenosine deaminase inhibitors alter adenosine and deoxyadenosine levels together with lympocytic growth and function, thus enhancing the chemotherapeutic effects of adenoside analogs in cancer chemotherapy [28].
Taking this into account, this review reports examples of many enzyme inhibitors from marine microorganisms with pharmaceutical significance. In particular, we report on two important sources of enzyme inhibitors from the marine environment: the sponges and the bacteria. In the case of sponges, we will only focus our attention on the kinase inhibitors since this class is the most representative that has so far been isolated from these marine organisms.
2. Marine Sponges
Although marine sponges are considered very simple marine organisms, they represent “chemical factories” because they are able to produce a great number of biologically active compounds [29]. There is still an ongoing debate about whether natural products isolated from sponges originated from sponges or from associated bacteria. Several experiments have evidenced that some compounds isolated from sponges are synthesized by their associated microorganisms [15,30]. In the case of enzyme inhibitors, there are also several evidences on their bacterial origin (see below).
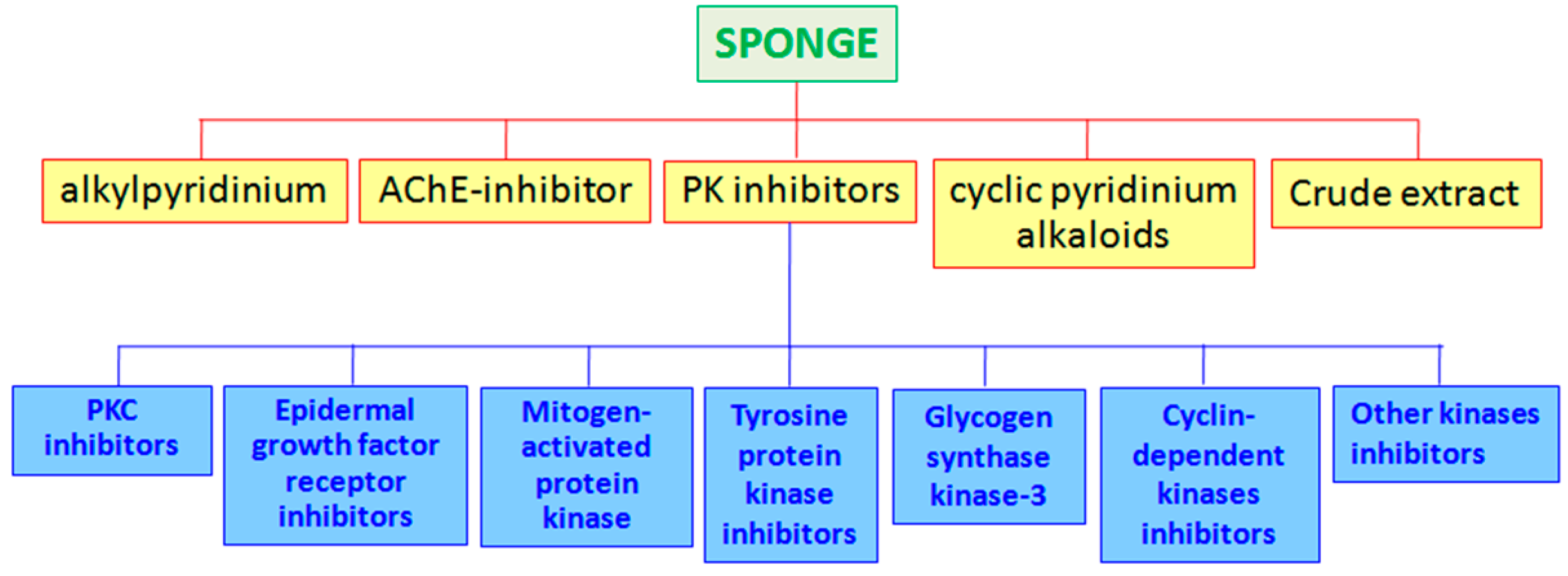
Polyketides, terpenoids, and peptides are the most abundant products isolated from sponges, showing inhibitory activities against many enzymes (Figure 1; see also Figure 2, Figure 3 and Figure 4 for the chemical structures of enzyme inhibitors reported in this paragraph).
Recently, the alkylpyridinium salts have been isolated from sponges, showing potent biological activities mainly as enzyme inhibitors. The marine sponge Reniera sarai produce some polymeric 3-alkylpyridinium salts [29]. Cyclostelletamines, cyclic pyridinium alkaloids, from the sponge Stelletta maxima [31] are able to inhibit the reaction of methyl quinuclidinyl benzylate with muscarinic acetylcholine receptors. The sponge Callyspongia fibrosa is the source of some polymers, which inhibit the epidermal growth factors [32]. Among acetylcholinesterase (AChE) inhibitors, an irreversible inhibitor (the onchidal) has been isolated from the mollusk Onchidella binneyi [33], and a pseudozoanthoxanthin-like compound from the coral Parazoanthus axinellae [34].
The enzyme adenine phosphoribosyl transferase of Leishmania tarentolae (L-APRT) has been inhibited by crude extracts from several marine invertebrates, as for example from the ascidian Polysyncraton sp. and from the bryozoan Bugula sp. [35]. Concerning the sponges, an inhibitor of L-APRT activity has been found in Dragmacidon sp. and Polymastia sp. The haplosclerid Callyspongia sp. SS97-23 was the source of three meroterpenoids, ilhabelanol, ilhabrene, and isoakaterpin, very potent inhibitors of L-APRT [36]. The marine sponge Petromica sp. BA99-103 produced halistanol sulfate, another inhibitor of L-APRT [35].
Marine sponges have been also proven as sources of protein kinase (PK) inhibitors. The enzymes, belonging to the protein kinase family, chemically catalyzed the transfer of a phosphate group to a defined substrate from a high-energy molecule. About 2% of all eukaryotic genes are protein kinases, organized in eight main groups [37]: (1) TK (tyrosine kinase); (2) TKL (tyrosine kinase-like); (3) STE (STE20, STE11, and STE7); (4) CK1 (casein kinase 1); (5) AGC (protein kinase A, protein kinase G, and protein kinase C); (6) CAMK (Ca2+/calmodulin-dependent kinases); (7) CMGC (Cdk, MAPK, GSK, Cdk-like); (8) RGC (receptor guanylyl cyclase). Kinases play key roles in different regulatory cellular processes, signal transduction, cell proliferation and differentiation, metabolic processes, apoptosis, and so on [2,38]. Because of the very different roles of the kinases, several diseases have as causes the deregulation of these enzymes. Furthermore, misregulation of various kinases has very often been reported in cancerous cells, so anticancer treatments involve kinases to specifically target cancer cells [39]. Kinase inhibitors represented a good challenge for cancer treatments, considering that to date about 130 kinase inhibitors are in different phases of clinical trials [40]. This is the case, for example, of Imitinib (Gleevec, Novartis), a tyrosine kinase inhibitor used in the prognosis for sufferers of chronic myeloid leukemia, now in the pharmaceutical market [41]. Inhibitors for different PK isolated from marine sponges are reported below (Table 1).
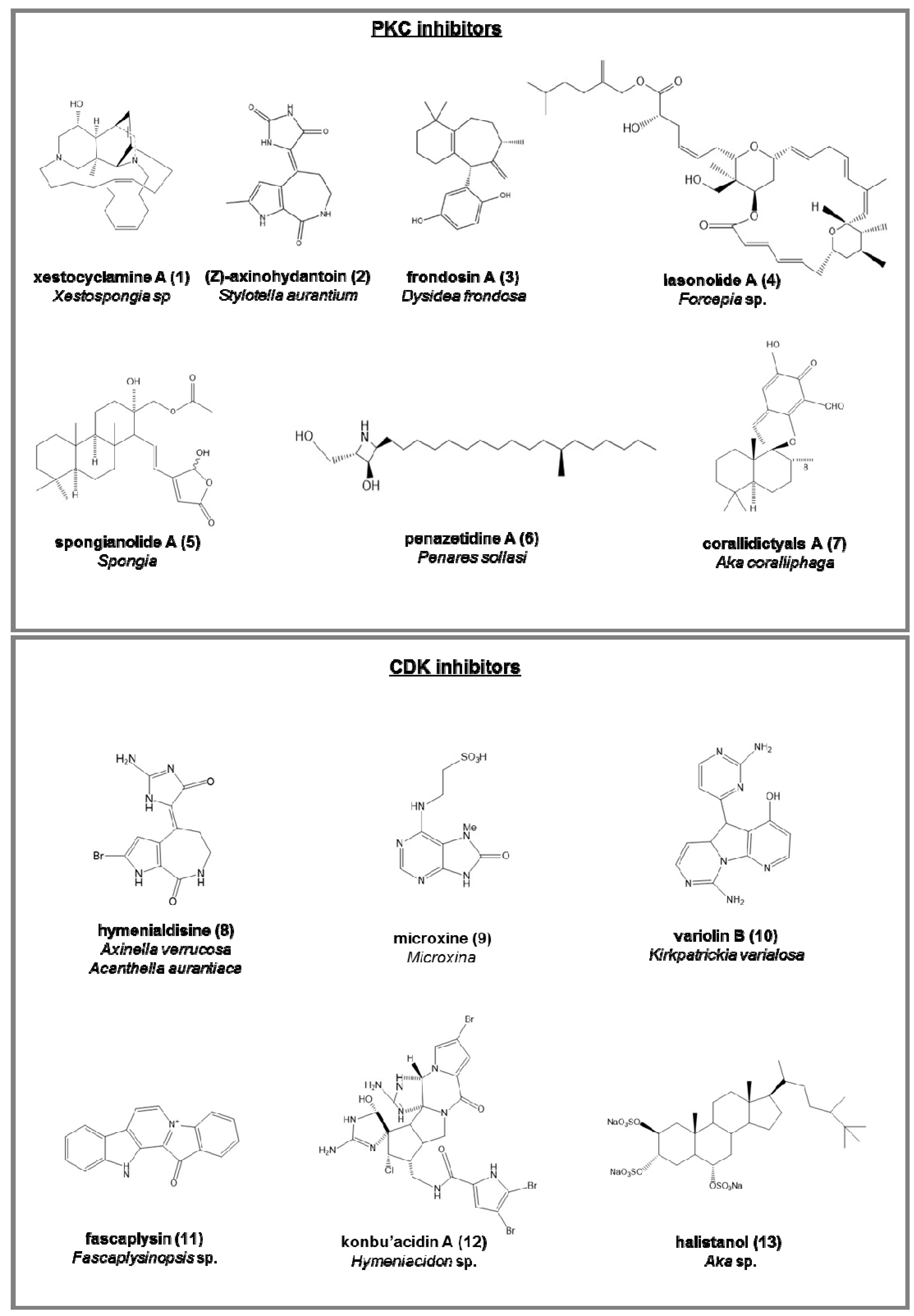
PKC inhibitors (serine/threonine kinases involved in cell differentiation, apoptosis, and inhibition cancer; Figure 2):
- -
- -
- (Z)-Axinohydantoin (2) and debromo-Z-axinohydantoin, from Stylotella aurantium [45];
- -
- -
- BRS1, a lipid from a calcarea sponge [47];
- -
- -
- -
- -
- penazetidine A (6), new azetidine compound from Penares sollasi, cytotoxic against human and murine cancer cell lines (A549, HT-29, B16/F10 and P388) [55];
- -
- corallidictyals A (7) and B, two diastereomeric spirosesquiterpene aldehydes, from Aka coralliphaga [56], selectively inhibiting the α-PKC isoform.
Cyclin-dependent kinases (CDK) inhibitors (serine/threonine kinases, involved in the regulation of the cell cycle; Figure 2):
CDK-1 (mitosis phase)
- -
- hymenialdisine (8), from Axinella verrucosa and Acanthella aurantiaca, able to act on NF-kappa B signaling process and indicated as possible pharmaceuticals in treating rheumatoid arthritis, multiple sclerosis, and Alzheimer’s diseases by some recent patents [57];
- -
- microxine (9), a purine derivative from genus Microxina [58];
- -
CDK-4 (G1 phase)
- -
- fascaplysin (11), a red pigment isolated from Fascaplysinopsis sp. It is a potent inhibitor of angiogenesis, inhibiting the proliferation of endothelial cells through VEGF [61].
- -
- konbu’acidin A (12), a bromopyrrole alkaloid from the Okinawan Hymeniacidon sp. [62];
- -
- two quinols and halistanol sulfate (13), novel sesquiterpene, from Aka sp. [63];
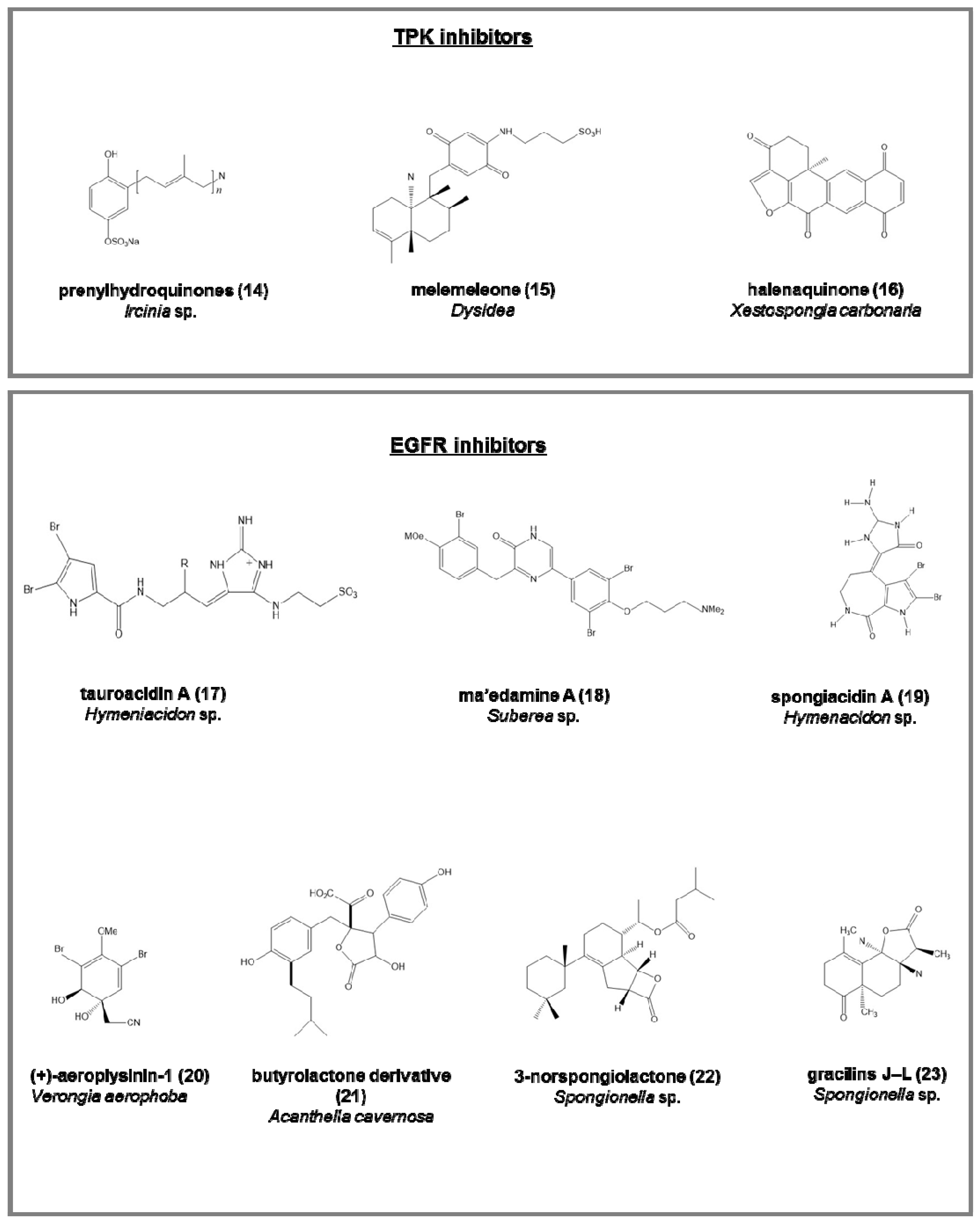
Tyrosine protein kinase (TPK) inhibitors (involved in the phosphorylation of tyrosine residues in proliferative diseases; Figure 3):
- -
- the penta-, hexa-, and hepta-prenylhydroquinone 4-sulfates (14), from the deep-sea Ircinia sp. [64], with cytotoxic activity against HIV-1 integrase enzyme and epidermal carcinoma cell line;
- -
- melemeleone (15), a novel sesquiterpene, from two species of Dysidea, with activity against tyrosine kinase pp60V-SRC [65];
- -
- halenaquinone (16), halenaquinol, halenaquinol sulfate, and xestoquinone from Xestospongia carbonaria [66].
Epidermal growth factor receptor (EGFR) inhibitors (tyrosine kinases involved mainly in breast cancer; Figure 3):
- -
- tauroacidins A (17) and B, bromopyrrole alkaloids from Hymeniacidon sp. [67];
- -
- ma’edamine A (18), bromotyrosine alkaloid, from Suberea sp. [68];
- -
- spongiacidins A (19) and B, bromopyrrole alkaloids from Hymenacidon sp. [69];
- -
- (+)-aeroplysinin-1 (20), from Verongia aerophoba, antitumoral on EGFR-dependent human breast cancer cell lines MCF-7 and ZR-75-1 [70];
- -
- butyrolactone derivative (21), from Acanthella cavernosa [71];
- -
- 3-norspongiolactone (22) and gracilins J–L (23), bioactive diterpenes from Spongionella sp. [72].
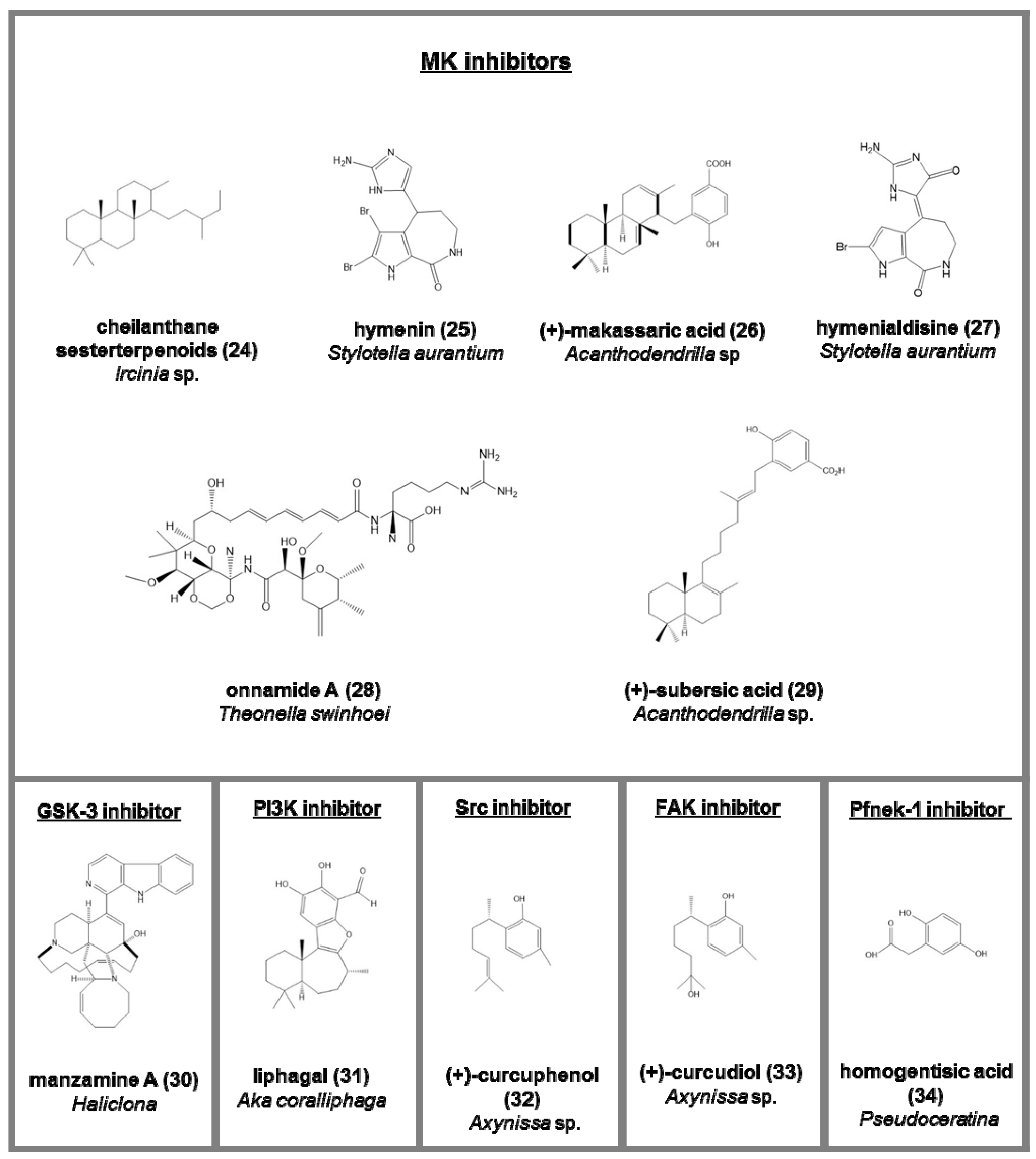
Mitogen-activated protein kinase (MK) inhibitors (serine/threonine protein kinases associated to stress; Figure 4):
- -
- -
- hymenin (25) and hymenialdisine (27), from Stylotella aurantium [75];
- -
- a methanol fraction from Batzella sp. [76];
- -
- onnamide A (28), heterocyclic compounds belonging to pederin family from Theonella swinhoei. Its activity is through the stimulation of plasminogen activator inhibitor-1 (PAI-1) considered a drug target against metastasis in human cancer cells [77];
- -
- (+)-makassaric acid (26) and (+)-subersic acid (29), meroterpenoids from Acanthodendrilla sp., acting inflammatory responses and cellular stress processes [78].
Glycogen synthase kinase-3 (GSK-3) inhibitors (a serine/threonine protein kinase, involved in neurodegenerative diseases; Figure 4):
- -
Other kinases inhibitors (Figure 4)
- -
- liphagal (31), a meroterpenoid from Aka coralliphaga, with inhibitory activity against PI3K and cytotoxic against human colon and human breast cancer [81];
- -
- (+)-curcuphenol (32) (Src protein kinase inhibitor) and (+)-curcudiol (33) (focal adhesion kinase, FAK, inhibitor), two bisabolenes type sesquiterpenoids Axynissa sp. [82];
- -
- -
- hymenialdisine, debromohymenialdisine, and four novel dihydrohymenialdisine derivatives from Cymbastela cantharella, able to inhibit the Polo-Like kinase-1 [84].
3. Marine Bacteria
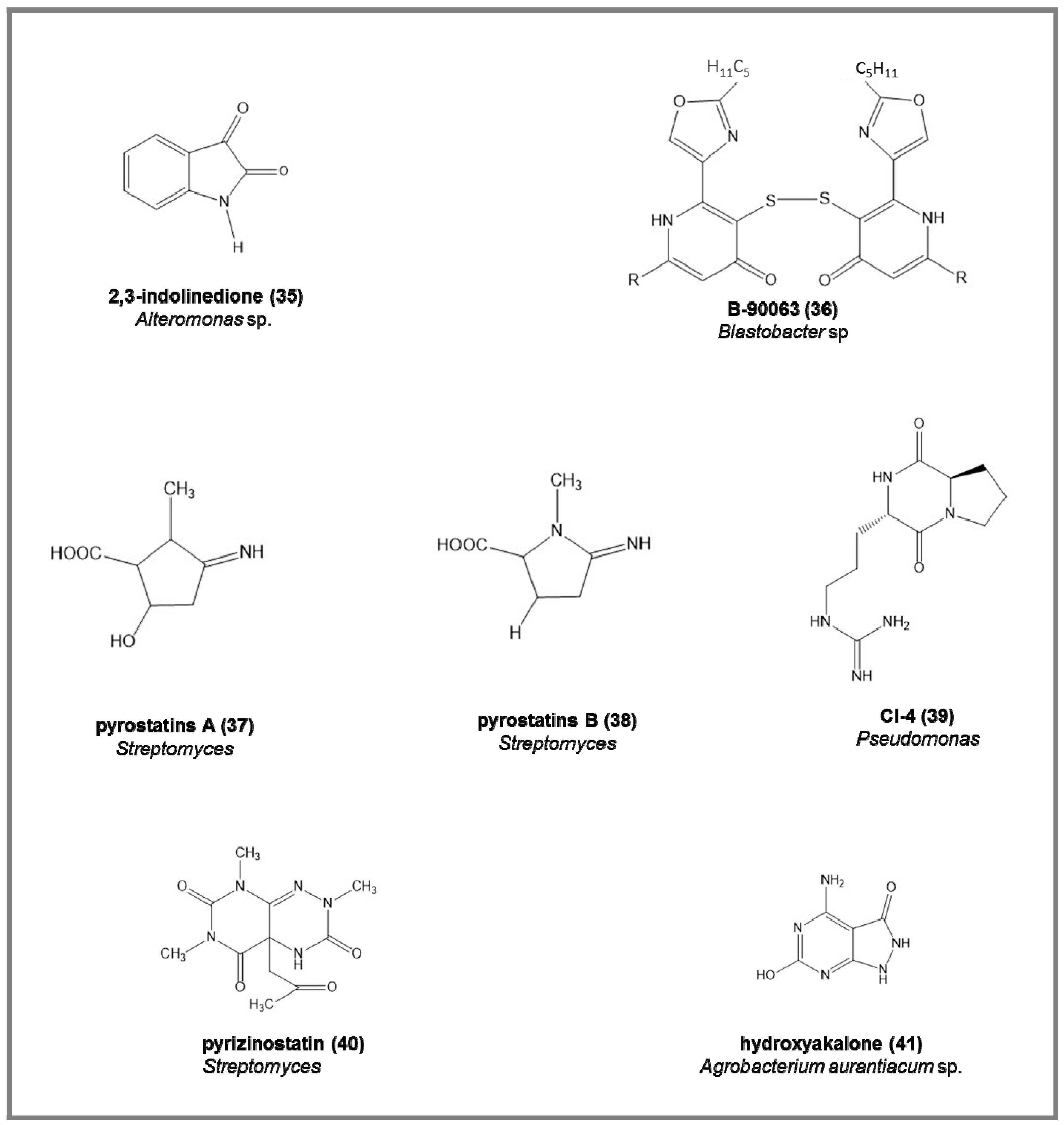
In the marine environment, several enzyme inhibitors have been isolated from bacteria and actinomycetes for their industrial applications and their use in medicine and agriculture [11] (see Figure 5).
One of the first isolated marine microbial source of enzyme inhibitors were represented by the Alteromonas sp., from which serine and cysteine protease inhibitors have been isolated [85]. An example of serine protease inhibitors are represented by the marinostatins C-1 and C-2, used as drugs in pancreatitis pathogenesis thanks to its properties to inhibit α-chymotrypsin (IC50 = 1.0–3.2 µM) [86,87].
Inhibitors for different PK isolated from marine bacteria are summarized in Table 2.
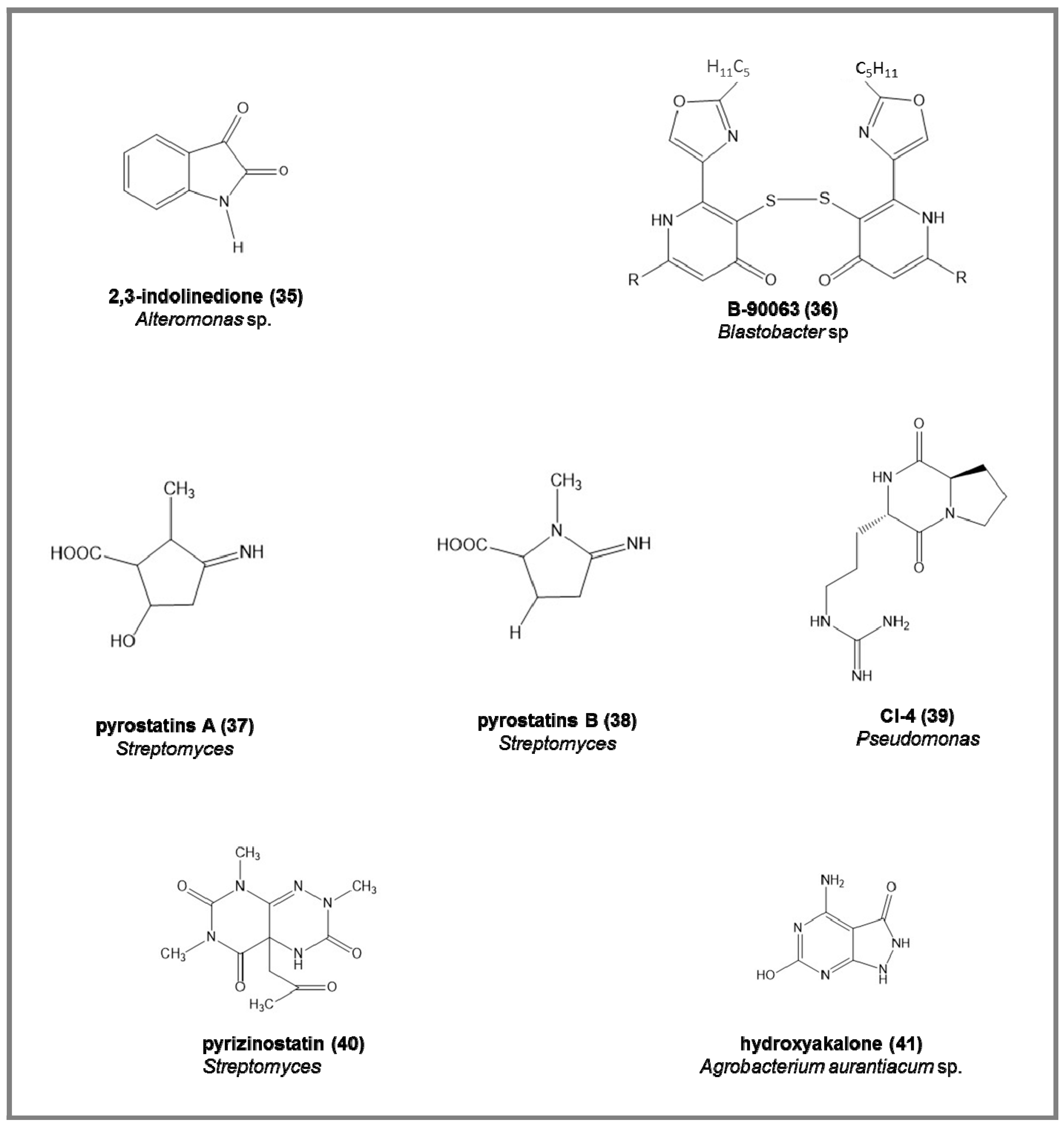
Some examples of the chemical structures of enzyme inhibitors from marine bacteria are reported in Figure 6:
- -
- endogenous monoamine oxidase (MAO) inhibitors, the 2,3-indolinedione (35), from marine Alteromonas sp., increasing the acetylcholine and dopamine in neurotransmission processes (IC50 = 9.2 µM) [88].
- -
- -
- -
- -
- -
- -
- angiotensin-converting enzyme (ACE) inhibitors and adenosine deaminase inhibitors (ADA) [27], from different Streptomyces strains. ACE inhibitors were able to reduce hypertension either by the suppression of angiotensin II biosynthesis or by the stimulation of bradykinin breakdown; ADA inhibitors were responsible for the alteration in adenosine and deoxyadenosine levels and in lymphocytic growth and functions, and enhance the effects of chemotherapeutic effects of adenosine analogs. Among 94 Streptomyces strains screened, 8 and 4 strains were positive for ACE and ADE inhibitors, respectively [27].
- -
Recently, the actinobacteria have also been considered as producers of enzyme inhibitors. This is the case for 30 strains of marine actinobacteria reported by Ganesan et al. [104], used in treating diabetes.
Marine bacteria associated with the sponge Jaspis sp. have been characterized for their production of protease inhibitors [105]. In particular, the associated bacteria Providencia sp. showed inhibitory activity against the protease subtilisin, and Bacillus sp. had inhibitory effects against the metalloproteinase thermolysin.
4. General Conclusions
The increased incidence of severe diseases, including cancer, prompted the scientific research to find new drugs. This also pushed the scientific communities to explore the marine environment for new pharmaceuticals. In fact, the marine environment has a great richness and biodiversity of micro- and macroorganisms, also exhibiting biosynthetic pathways to produce secondary metabolites useful both against predators and active compounds for human health attracting the attention of pharmaceutical industries. A very significant example is represented by several classes of enzyme inhibitors isolated from marine sponges and bacteria, as reported in the review. These enzyme inhibitors show cytotoxic activity against many different cancer cell lines (see Table 1). Marine-derived enzyme inhibitors are considered viable alternatives in pharmaceuticals for replacing synthetic drugs to combat, for example, cancer as well as viral, amtiinflammatory, and neurodegenerative diseases. Nonetheless, further studies will be necessary to investigate the human consumer’s well being. The general significance of the topic of this review is well underlined by the number of EU-funded projects, as well as by Horizon 2020, which are aimed at improving the exploitation of marine organisms for drug discovery.
Acknowledgments
Nadia Ruocco (Ph.D. in Biology, University of Naples Federico II) was supported by a Ph.D. fellowship co-funded by the Stazione Zoologica Anton Dohrn and Bio-Organic Chemistry Unit of the Institute of Biomolecular Chemistry-CNR, Pozzuoli, Naples, Italy.
Author Contributions
M.C., N.R. and S.C. conceived and designed the scheme of the review and wrote the paper. F.P. created the illustrations in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef] [PubMed]
- Skropeta, D.; Pastro, N.; Zivanovic, A. Kinase inhibitors from marine sponges. Mar. Drugs 2011, 9, 2131–2154. [Google Scholar] [CrossRef] [PubMed]
- Bode, W.; Huber, R. Natural protein proteinase inhibitors and their interaction with proteinases. Eur. J. Biochem. 1992, 204, 433–451. [Google Scholar] [CrossRef] [PubMed]
- Terashita, T.; Kono, M.; Murao, S. Promoting effect of S-PI on fruiting of Lentinus edodes. Trans. Mycol. Soc. Jpn. 1980, 21, 137–140. [Google Scholar]
- Demuth, H.U. Recent developments in inhibiting cysteine and serine proteases. J. Enzym. Inhib. 1990, 3, 249–278. [Google Scholar] [CrossRef] [PubMed]
- Delston, R.B.; Kothary, M.H.; Shangraw, K.A.; Tall, B.D. Isolation and characterization of a zinc-containing metalloprotease expressed by Vibrio tubiashii. Can. J. Microbiol. 2003, 49, 525. [Google Scholar] [CrossRef] [PubMed]
- Bo-Linn, G.W.; Santa Ana, C.A.; Morawski, S.G.; Fordtran, J.S. Starch blockers-their effect on calorie absorption from a high-starch meal. N. Engl. J. Med. 1982, 307, 1413–1416. [Google Scholar] [CrossRef] [PubMed]
- Puls, W.; Keup, U. Influence of an a-amylase inhibitor (BAY d 7791) on blood glucose, serum insulin and NEFA in starch loading tests in rats, dogs and man. Diabetologia 1973, 9, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, H. Enzyme Inhibitor of Microbial Origin; University of Tokyo Press: Tokyo, Japan, 1972. [Google Scholar]
- Hamato, N.; Takano, R.; Kamei-Hayashi, T.; Imada, C.; Hara, S. Leupeptins produced by the marine Alteromonas sp. B-10-31. Biosci. Biotechnol. Biochem. 1992, 56, 1316–1318. [Google Scholar] [CrossRef]
- Imada, C. Enzyme inhibitors of marine microbial origin with pharmaceutical importance. Mar. Biotechnol. 2004, 6, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Aneiros, A.; Garateix, A. Bioactive peptides from marine sources: Pharmacological properties and isolation procedures. J. Chromatogr. B. Anal. Technol. Biomed. Life Sci. 2004, 15, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2012, 29, 144–222. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed]
- Mehbub, M.F.; Lei, J.; Franco, C.; Zhang, W. Marine sponge derived natural products between 2001 and 2010: Trends and opportunities for discovery of bioactives. Mar. Drugs 2014, 12, 4539–4577. [Google Scholar] [CrossRef] [PubMed]
- Reen, F.; Gutierrez-Barranquero, J.; Dobson, A.; Adams, C.; O’Gara, F. Emerging concepts promising new horizons for marine biodiscovery and synthetic biology. Mar. Drugs 2015, 13, 2924–2954. [Google Scholar] [CrossRef] [PubMed]
- Amsler, C.D.; Iken, K.B.; McClintock, J.B.; Baker, B.J. Secondary metabolites from Antarctic marine organisms and their ecological implications. In Marine ChemicalEcology; McClintock, J.B., Baker, J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 267–300. [Google Scholar]
- Lindquist, N. Chemical defense of early stages of benthic marine invertebrates. J. Chem. Ecol. 2002, 28, 1987–2000. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Hu, W.P.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2007, 24, 31–86. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, D.J. Marine pharmacology. Antonie Leeuwenhoek 2000, 77, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2013, 30, 237–323. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2016, 33, 382–431. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, R.A. On the role of inorganic ions in the physiology of marine bacteria, VI. In Advances in Microbiology of the Sea; Droop, M.R., Wood, E.J.F., Eds.; Academic Press: New York, NY, USA, 1968; Volume 1, pp. 95–126. [Google Scholar]
- Maeda, M.; Taga, N. Extracellular nuclease produced by a marine bacterium, II: Purification and properties of extracellular nuclease from a marine Vibrio sp. Can. J. Microbiol. 1976, 22, 1443–1452. [Google Scholar] [CrossRef]
- Kobori, H.; Taga, N. Extracellular alkaline phosphatase from marine bacteria: Purification and properties of extracellular phosphatase from a marine Pseudomonas sp. Can. J. Microbiol. 1980, 26, 833–838. [Google Scholar] [CrossRef]
- Chellaram, C.; Prem Anand, T.; Felicia Shanthini, C.; Chandrika, M.; Gladis, C. Screening for enzyme inhibitors in marine bacteria. Int. J. PharmTech Res. 2014, 6, 351–355. [Google Scholar]
- Agarwal, R.P. Inhibitors of adenosine deaminase. Pharmacol. Ther. 1982, 17, 399–429. [Google Scholar] [CrossRef]
- Sepčić, K.; Marcel, V.; Klaebe, A.; Turk, T.; Šuput, D.; Fournier, D. Inhibition of acetylcholinesterase by an alkylpyridinium polymer from the marine sponge, Reniera sarai. Biochim. Biophys. Acta 1998, 1387, 217–225. [Google Scholar] [CrossRef]
- Lee, Y.K.; Lee, J.-H.; Lee, H.K. Microbial symbiosis in marine sponges. J. Microbiol. 2001, 39, 254–264. [Google Scholar]
- Fusetani, N.; Asai, N.; Matsunaga, S.; Honda, K.; Yasumuro, K. Cyclostellettamines A–F, pyridine alkaloids which inhibit binding of methyl quinuclidinyl benzilate (QNB) to muscarinic acetylcholine receptors, from the marine sponge, Stelletta maxima. Tetrahedron Lett. 1994, 35, 3967–3970. [Google Scholar] [CrossRef]
- Davies-Coleman, M.T.; Faulkner, J.; Dubowchik, G.; Roth, G.P.; Polson, C.; Fairchild, C. A new EGF-active polymeric pyridinium alkaloid from the sponge Callyspongia fibrosa. J. Org. Chem. 1993, 58, 5925–5930. [Google Scholar] [CrossRef]
- Abramson, S.N.; Radic, Z.; Manker, D.; Faulkner, J.; Taylor, P. Onchidal: A naturally occurring irreversible inhibitor of acetylcholinesterase with a novel mechanism of action. Mol. Pharmacol. 1989, 36, 349–354. [Google Scholar] [PubMed]
- Turk, T.; Macek, P.; Sŭput, D. Inhibition of acetylcholinesterase by a pseudozoanthoxanthin-like compound isolated from the zoanthid Parazoanthus axinellae (O. Schmidt). Toxicon 1995, 33, 133–142. [Google Scholar] [CrossRef]
- Seleghim, M.H.R.; Lira, S.P.; Kossuga, M.H.; Batista, T.; Berlinck, R.G.S.; Hajdu, E.; Muricy, G.; Rocha, R.M.; Nascimento, G.F.; Silva, M.; et al. Antibiotic, cytotoxic and enzyme inhibitory activity of crude extracts from Brazilian marine invertebrates. Rev. Bras. Farmacogn. 2007, 17, 287–318. [Google Scholar] [CrossRef]
- Gray, C.A.; de Lira, S.P.; Silva, M.; Pimenta, E.F.; Thiemann, O.H.; Oliva, G.; Hajdu, E.; Andersen, R.J.; Berlinck, R.G.S. Sulfated meroterpenoids from the Brazilian sponge Callyspongia sp. are inhibitors of the antileishmaniasis target adenosine phosphoribosyl transferase. J. Org. Chem. 2006, 71, 8685–8690. [Google Scholar] [CrossRef] [PubMed]
- Duong-Ly, K.C.; Peterson, J.R. The human kinome and kinase inhibition. Curr. Protoc. Pharmacol. 2013. [Google Scholar] [CrossRef]
- Novak, K. Conference report-protein kinase inhibitors in cancer treatment: Mixing and matching? In Proceedings of the Keystone Symposium on Protein Kinases and Cancer, Medscape General Medicine, Lake Tahoe, CA, USA, 24–29 February 2004. [Google Scholar]
- Mariaule, G.; Belmont, P. Cyclin-dependent kinase inhibitors as marketed drugs: Where are we now? A short survey. Molecules 2014, 19, 14366–14382. [Google Scholar] [CrossRef] [PubMed]
- Norman, P. Overview: Kinase Therapeutics Pipelines: An Assessment of Targets and Agents in Development; Cambridge Healthtech Institute: Needham, MA, USA, 2007. [Google Scholar]
- Goldstein, D.; Gray, N.; Zarrinkar, P. High-throughput kinase profiling as a platform for drug discovery. Nat. Rev. Drug Discov. 2008, 7, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Peters, B.M.; Kurz, L.; Schatzman, R.C.; McCarley, D.; Lou, L.; Crews, P. An alkaloid protein kinase C inhibitor, xestocyclamine A, from the marine sponge Xestospongia sp. J. Am. Chem. Soc. 1993, 115, 10436–10437. [Google Scholar] [CrossRef]
- Rodriguez, J.; Crews, P. Revised structure of xestocyclaminee A and description of new analogue. Tetrahedon Lett. 1994, 35, 4719–4722. [Google Scholar] [CrossRef]
- Yun, H.; Gagnon, A.; Danishefsky, S.J. Toward the synthesis of xestocyclamine A: Investigation of double Michael reaction and direct aza Diels-Alder reaction. Tetrahedron Lett. 2006, 47, 5311–5315. [Google Scholar] [CrossRef]
- Patil, A.D.; Freyer, A.J.; Killmer, L.; Hofmann, G.; Johnson, R.K. Z-Axinohydantoin and debromo-Z-axinohydantoin from the sponge Stylotella aurantium: Inhibitors of protein kinase C. Nat. Prod. Res. 1997, 9, 201–207. [Google Scholar]
- Freyer, A.J.; Patil, A.D.; Killmer, L.; Offen, P.; Carte, B.; Jurewicz, A.J.; Johnson, R.K. Frondosins, five new sesquiterpene hydroquinone derivatives with novel skeletons from the sponge Dysidea frondosa: Inhibitors of interleukin-8 receptors. Tetrahedron 1997, 53, 5047–5060. [Google Scholar]
- Willis, R.H.; de Vries, D.J. BRS1, a C30 bis-amino, bis-hydroxy polyunsaturated lipid from an Australian calcareous sponge that inhibits protein kinase C. Toxicon 1997, 35, 1125–1129. [Google Scholar] [CrossRef]
- Shigemori, H.; Madono, T.; Sasaki, T.; Mikami, Y.; Kobayashi, J. Nakijiquinones A and B, new antifungal sesquiterpenoid quinones with an amino acid residue from an Okinawan marine sponge. Tetrahedron 1994, 50, 8347–8354. [Google Scholar] [CrossRef]
- Kobayashi, J.; Madono, T.; Shigemori, H. Nakijiquinones C and D, new sesquiterpenoid quinones with a hydroxy amino acid residue from a marine sponge inhibiting c-erbB-2 kinase. Tetrahedron 1995, 51, 10867–10874. [Google Scholar] [CrossRef]
- Kissau, L.; Stahl, P.; Mazitschek, R.; Giannis, A.; Waldmann, H. Development of natural product-derived receptor tyrosine kinase inhibitors based on conservation of protein domain fold. J. Med. Chem. 2003, 46, 2917–2931. [Google Scholar] [CrossRef] [PubMed]
- Longley, R.E.; Harmody, D. A rapid colorimetric microassay to detect agonist/antagonists of protein-kinase-C based on adherence of EL-4.IL-2 cells. J. Antibiot. 1991, 44, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.A.; Koehn, F.E.; Longley, R.E.; McConnell, O.J. Lasonolide A, a new cytotoxic macrolide from the marine sponge Forcepia sp. J. Am. Chem. Soc. 1994, 116, 6015–6016. [Google Scholar] [CrossRef]
- Isbrucker, R.A.; Guzman, E.A.; Pitts, T.P.; Wright, A.E. Early effects of lasonolide A on pancreatic cancer cells. J. Pharmacol. Exp. Ther. 2009, 331, 733–739. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Kulanthaivel, P.; Baker, B.J. New cytotoxic sesterterpenes from the marine sponge Spongia sp. Tetrahedron Lett. 1994, 35, 7189–7192. [Google Scholar] [CrossRef]
- Alvi, K.A.; Jaspars, M.; Crews, P.; Strulovici, B.; Oto, E. Penazetidine-A, an alkaloid inhibitor of protein kinase C. Bioorg. Med. Chem. Lett. 1994, 4, 2447–2450. [Google Scholar] [CrossRef]
- Chan, J.A.; Freyer, A.J.; Carte, B.K.; Hemling, M.E.; Hofmann, G.A.; Mattern, M.R.; Mentzer, M.A.; Westley, J.W. Protein kinase C inhibitors: Novel spirosesquiterpene aldehydes from a marine sponge Aka (=Siphonodictyon) coralliphagum. J. Nat. Prod. 1994, 57, 1543–1548. [Google Scholar] [CrossRef] [PubMed]
- Meijer, L.; Thunnissen, A.M.; White, A.W.; Garnier, M.; Nikolic, M.; Tsai, L.H.; Walter, J.; Cleverley, K.E.; Salinas, P.C.; Wu, Y.Z.; et al. Inhibition of cyclin-dependent kinases, GSK-3β and CK1 by hymenialdisine, a marine sponge constituent. Chem. Biol. 2000, 7, 51–63. [Google Scholar] [CrossRef]
- Killday, B.; Yarwood, D.; Sills, M.; Murphy, P.; Hooper, J.; Wright, A. Microxine, a new cdc2 kinase inhibitor from the Australian marine sponge Microxima species. J. Nat. Prod. 2001, 64, 525–526. [Google Scholar] [CrossRef] [PubMed]
- Trimurtulu, G.; Faulkner, D.J.; Perry, N.B.; Ettouati, L.; Litaudon, M.; Blunt, J.W.; Munro, M.H.G.; Jameson, G.B. Alkaloids from the antarctic sponge Kirkpatrickia varialosa. Part 2: Variolin A and N(3′)-methyl tetrahydrovariolin B. Tetrahedron 1994, 50, 3993–4000. [Google Scholar] [CrossRef]
- Walker, S.R.; Carter, E.J.; Huff, B.C.; Morris, J.C. Variolins and related alkaloids. Chem. Rev. 2009, 109, 3080–3098. [Google Scholar] [CrossRef] [PubMed]
- Soni, R.; Muller, L.; Furet, P.; Schoepfer, J.; Stephan, C.; Zumstein-Mecker, S.; Fretz, H.; Chaudhuri, B. Inhibition of cyclin-dependent kinase 4 (Cdk4) by fascaplysin, a marine natural product. Biochem. Biophys. Res. Commun. 2000, 275, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Suzuki, M.; Tsuda, M. Konbu’acidin A, a new bromopyrrole alkaloid with cdk4 inhibitory activity from hymeniacidon sponge. Tetrahedron 1997, 53, 15681–15684. [Google Scholar] [CrossRef]
- Mukku, V.; Edrada, R.A.; Schmitz, F.J.; Shanks, M.K.; Chaudhuri, B.; Fabbro, D. New sesquiterpene quinols from a micronesian sponge, Aka sp. J. Nat. Prod. 2003, 66, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Bifulco, G.; Bruno, I.; Minale, L.; Riccio, R.; Debitus, C.; Bourdy, G.; Vassas, A.; Lavayre, J. Bioactive prenylhydroquinone sulfates and a novel C31 furanoterpene alcohol sulfate from the marine sponge, Ircinia sp. J. Nat. Prod. 1995, 58, 1444–1449. [Google Scholar] [CrossRef]
- Alvi, K.A.; Diaz, M.C.; Crews, P.; Slate, D.L.; Lee, R.H.; Moretti, R. Evaluation of new sesquiterpene quinones from two Dysidea sponge species as inhibitors of protein tyrosine kinase. J. Org. Chem. 1992, 57, 6604–6607. [Google Scholar] [CrossRef]
- Lee, R.H.; Slate, D.L.; Moretti, R.; Alvi, K.A.; Crews, P. Marine sponge polyketide inhibitors of protein tyrosine kinase. Biochem. Biophys. Res. Commun. 1992, 184, 765–772. [Google Scholar] [CrossRef]
- Kobayashi, J.; Inaba, K.; Tsuda, M. Tauroacidins A and B, new bromopyrrole alkaloids possessing a taurine residue from hymeniacidon sponge. Tetrahedron 1997, 53, 16679–16682. [Google Scholar] [CrossRef]
- Kobayashi, J.; Hirano, K.; Kubota, T.; Tsuda, M.; Watanabe, K.; Fromont, J. Ma’edamines A and B, cytotoxic bromotyrosine alkaloids with a unique 2(1H)pyrazinone ring from sponge Suberea sp. Tetrahedron 2000, 56, 8107–8110. [Google Scholar]
- Inaba, K.; Sato, H.; Tsuda, M.; Kobayashi, J. Spongiacidins A–D, new bromopyrrole alkaloids from hymeniacidon sponge. J. Nat. Prod. 1998, 61, 693–695. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, M.-H.; Leake, R.E.; Rinaldi, F.; Müller-Klieser, W.; Maidhof, A.; Müller, W.E.G.; Schröder, H.C. Inhibition of intrinsic protein tyrosine kinase activity of EGF-receptor kinase complex from human breast cancer cells by the marine sponge metabolite (+)-aeroplysinin-1. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1990, 97, 151–158. [Google Scholar] [CrossRef]
- Rateb, M.E.; Houssen, W.E.; Legrave, N.M.; Clements, C.; Jaspars, M.; Ebel, R. Dibenzofurans from the marine sponge-derived ascomycete Super1F1-09. Bot. Mar. 2010, 53, 499–506. [Google Scholar] [CrossRef]
- Rateb, M.E.; Houssen, W.E.; Schumacher, M.; Harrison, W.T.A.; Diederich, M.; Ebel, R.; Jaspars, M. Bioactive diterpene derivatives from the marine sponge Spongionella sp. J. Nat. Prod. 2009, 72, 1471–1476. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, M.S.; Edser, A.; King, G.; Whitmore, J.; Quinn, R.J. Cheilanthane sesterterpenes, protein kinase inhibitors, from a marine sponge of the genus Ircinia. J. Nat. Prod. 2001, 64, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Kesler, C.; Neary, J.; Fishman, L. Effects of marine sponge extracts on mitogen-activated protein kinase (MAPK/ERK1,2) activity in SW-13 human adrenal carcinoma cells. Toxicon 2001, 39, 1835–1839. [Google Scholar] [CrossRef]
- Tasdemir, D.; Mallon, R.; Greenstein, M.; Feldberg, L.; Kim, S.; Collins, K.; Wojciechowicz, D.; Mangalindan, G.; Concepcion, G.; Harper, M.K.; et al. Aldisine alkaloids from the Philippine sponge Stylissa massa are potent Inhibitors of mitogen-activated protein kinase kinase-1 (MEK-1). J. Med. Chem. 2002, 45, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.; Malpezzi, E.; Costa, L.; Berlinck, R.; Almeida, A.; Ogawa, C.; Sanchez, M.; Hajdu, E. Cytotoxic and neurotoxic effects induced by halitoxin isolated from Amphimedon viridis (Porifera). Toxicon 1996, 34, 335. [Google Scholar] [CrossRef]
- Lee, K.H.; Nishimura, S.; Matsunaga, S.; Fusetani, N.; Horinouchi, S.; Yoshida, M. Inhibition of protein synthesis and activation of stress-activated protein kinases by onnamide A and theopederin B, antitumor marine natural products. Cancer Sci. 2005, 96, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.E.; Telliez, J.B.; Liu, J.; Tahir, A.; van Soest, R.; Andersen, R.J. Meroterpenoid MAPKAP (MK2) inhibitors isolated from the Indonesian marine sponge Acanthodendrilla sp. J. Nat. Prod. 2004, 67, 2127–2129. [Google Scholar] [CrossRef] [PubMed]
- Hamann, M.; Alonso, D.; Martin-Aparicio, E.; Fuertes, A.; Perez-Puerto, M.; Castro, A.; Morales, S.; Navarro, M.; Monte-Millan, M.; Medina, M.; et al. Glycogen synthase kinase-3 (GSK-3) inhibitory activity and structure activity relationship (SAR). Studies of the manzamine alkaloids. Potential for Alzheimer’s disease. J. Nat. Prod. 2007, 70, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Ang, K.; Holmes, M.; Higa, T.; Hamann, M.; Kara, U. In vivo antimalarial activity of the beta-carboline alkaloid manzamine A. Antimicrob. Agents Chemother. 2000, 44, 1645–1649. [Google Scholar] [CrossRef] [PubMed]
- Marion, F.; Williams, D.E.; Patrick, B.O.; Hollander, I.; Mallon, R.; Kim, S.C.; Roll, D.M.; Feldberg, L.; Van Soest, R.; Andersen, R.J. Liphagal, a selective inhibitor of PI3 kinase alpha isolated from the sponge Aka coralliphaga: Structure elucidation and biomimetic synthesis. Org. Lett. 2006, 8, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Hertiani, T.; Edrada-Ebel, R.A.; Kubbutat, M.H.G.; van Soest, R.W.M.; Proksch, P. Protein kinase inhibitors from Indonesian sponge Axynissa sp. Maj. Farm. Indones. 2008, 19, 78–85. [Google Scholar]
- Lebouvier, N.; Jullian, V.; Desvignes, I.; Maurel, S.; Parenty, A.; Dorin-Semblat, D.; Doerig, C.; Sauvain, M.; Laurent, D. Antiplasmodial activities of homogentisic acid derivative protein kinase inhibitors isolated from a vanuatu marine sponge Pseudoceratina sp. Mar. Drugs 2009, 7, 640–653. [Google Scholar] [CrossRef] [PubMed]
- Sauleau, P.; Retailleau, P.; Nogues, S.; Carletti, I.; Marcourt, L.; Raux, R.; Al Mourabit, A.; Debitus, C. Dihydrohymenialdisines, new pyrrole-2-aminoimidazole alkaloids from the marine sponge Cymbastela cantharella. Tetrahedron Lett. 2011, 52, 2676–2678. [Google Scholar] [CrossRef]
- Imada, C.; Taga, N.; Maeda, M. Cultivation conditions for subtilisin inhibitor-producing bacterium and general properties of the inhibitor “marinostatin”. Bull. Jpn. Soc. Sci. Fish 1985, 51, 805–810. [Google Scholar] [CrossRef]
- Imada, C.; Hara, S.; Maeda, M.; Simidu, U. Aminoacid sequences of marinostatins C-1 and C-2 from marine Alteromonas sp. Bull. Jpn. Soc. Sci. Fish 1986, 52, 1455–1459. [Google Scholar] [CrossRef]
- Imada, C.; Maeda, M.; Hara, S.; Taga, N.; Simidu, U. Purification and characterization of subtilisin inhibitors ‘‘marinostatin’’ produced by marine Alteromonas sp. J. Appl. Bacteriol. 1986, 60, 469–476. [Google Scholar] [CrossRef]
- Gil-Turnes, M.S.; Hay, M.E.; Fenical, W. Symbiotic marine bacteria defend crustacean embryos from a pathogenic fungus. Science 1989, 240, 116–118. [Google Scholar] [CrossRef]
- Imada, C.; Maeda, M.; Taga, N. Purification and characterization of the protease inhibitor “monastatin” from a marine Alteromonas sp. with reference to inhibition of the protease produced by a bacterium pathogenic to fish. Can. J. Microbiol. 1985, 31, 1089–1094. [Google Scholar] [CrossRef]
- Imada, C.; Nishimoto, S.; Hara, S. The effect of addition of protease inhibitor from marine bacterium on the strength of gel formation of sardine meat gel (Kamaboko). Nippon Suisan Gakkaishi 2001, 67, 85–89. (In Japanese) [Google Scholar] [CrossRef]
- Takaishi, S.; Tuchiya, N.; Sato, A.; Negishi, T.; Takamatsu, Y.; Matsushita, Y.; Watanabe, T.; Iijima, Y.; Haruyama, H.; Kinoshita, T.; et al. B-90063, a novel endothelin converting enzyme inhibitor isolated from a new marine bacterium, Blastobacter sp. SANK 71894. J. Antibiot. 1998, 51, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Hasegawa, H.; Umekawa, K.; Ueki, Y.; Ohashi, N.; Kanaoka, M. Design, synthesis and biological activity of novel non-peptidyl endothelin converting enzyme inhibitors, 1-phenyl-tetrazole-formazan analogues. Bioorg. Med. Chem. Lett. 2002, 12, 1275–1278. [Google Scholar] [CrossRef]
- Aoyama, T.; Kojima, F.; Imada, C.; Muraoka, Y.; Naganawa, H.; Okami, Y.; Takeuchi, T.; Aoyagi, T. Pyrostatins A and B, new inhibitors of N-acetyl-b-glucosaminidase, produced by Streptomyces sp. SA-3501. J. Enzym. Inhib. 1995, 8, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Alhadeff, J.A.; Holzinger, R.T. Characterization of bhexosaminidase from liver and sera of diabetic patients and control. Biochem. Med. 1983, 27, 214–225. [Google Scholar] [CrossRef]
- Drexler, H.G.; Gaedicke, G. Isoenzyme studies in human leukemia, III: B-hexosaminidase (E.C. 3.3.1.30). Leuk. Res. 1983, 7, 611–619. [Google Scholar] [CrossRef]
- Pluncinsky, M.C.; Prorok, J.J.; Alhadeff, J.A. Variant serum beta-hexosaminidase as a biochemical marker of malignancy. Cancer 1986, 58, 1484–1487. [Google Scholar] [CrossRef]
- Aoyagi, T.; Hatsu, M.; Imada, C.; Naganawa, H.; Okami, Y.; Takeuchi, T. Pyrizinostatin: A new inhibitor of pyroglutamyl peptidase. J. Antibiot. 1992, 45, 1795–1796. [Google Scholar] [CrossRef] [PubMed]
- Prasad, C.A.; Peterkofsky, A. Determination of pyroglutamylpeptidase and amidase activities toward thyrotropinreleasing hormone in hamster hypothalamus extracts. J. Biol. Chem. 1976, 251, 3229–3234. [Google Scholar] [PubMed]
- Izumida, H.; Miki, W.; Sano, H.; Endo, M. Agar plate method, a new assay for chitinase inhibitors using a chitindegrading bacterium. J. Mar. Biotechnol. 1995, 2, 163–166. [Google Scholar]
- Izumida, H.; Imamura, N.; Sano, H. A novel chitinase inhibitor from a marine bacterium, Pseudomonas sp. J. Antibiot. 1996, 49, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Van Aalten, D.M.F.; Komander, D.; Systad, B.; Gaseidnes, S.; Peter, M.G.; Eijsink, V.G.H. Structural insights into the catalytic mechanism of a family 18 exo-chitinase. Proc. Natl. Acad. Sci. USA 2001, 98, 8979–8984. [Google Scholar] [CrossRef] [PubMed]
- Izumida, H.; Adachi, K.; Nishijima, M.; Endo, M.; Miki, W. Akalone: A novel xanthine oxidase inhibitor produced by the marine bacterium, Agrobacterium aurantiacum sp. nov. J. Mar. Biotechnol. 1985, 2, 115–118. [Google Scholar]
- Borges, F.; Fernandes, E.; Roleira, F. Progress towards the discovery of xanthine oxidase inhibitors. Curr. Med. Chem. 2002, 9, 195–217. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Raja, S.; Sampathkumar, P.; Sivakumar, K.; Thangaradjou, T. Isolation and screening of α-glucosidase enzyme inhibitor producing marine actinobacteria. Afr. J. Microbiol. Res. 2011, 5, 3437–3445. [Google Scholar] [CrossRef]
- Wahyudi, A.T.; Mubarik, N.R. Screening and characterization of protease inhibitors from marine bacteria associated with sponge Jaspis sp. HAYATI J. Biosci. 2010, 4, 173–178. [Google Scholar] [CrossRef]
Figure 1.
Enzyme inhibitors isolated from marine sponges.
Figure 2.
Chemical structures of some natural PKC and CDK inhibitors isolated from sponges, reported as examples.
Figure 2.
Chemical structures of some natural PKC and CDK inhibitors isolated from sponges, reported as examples.
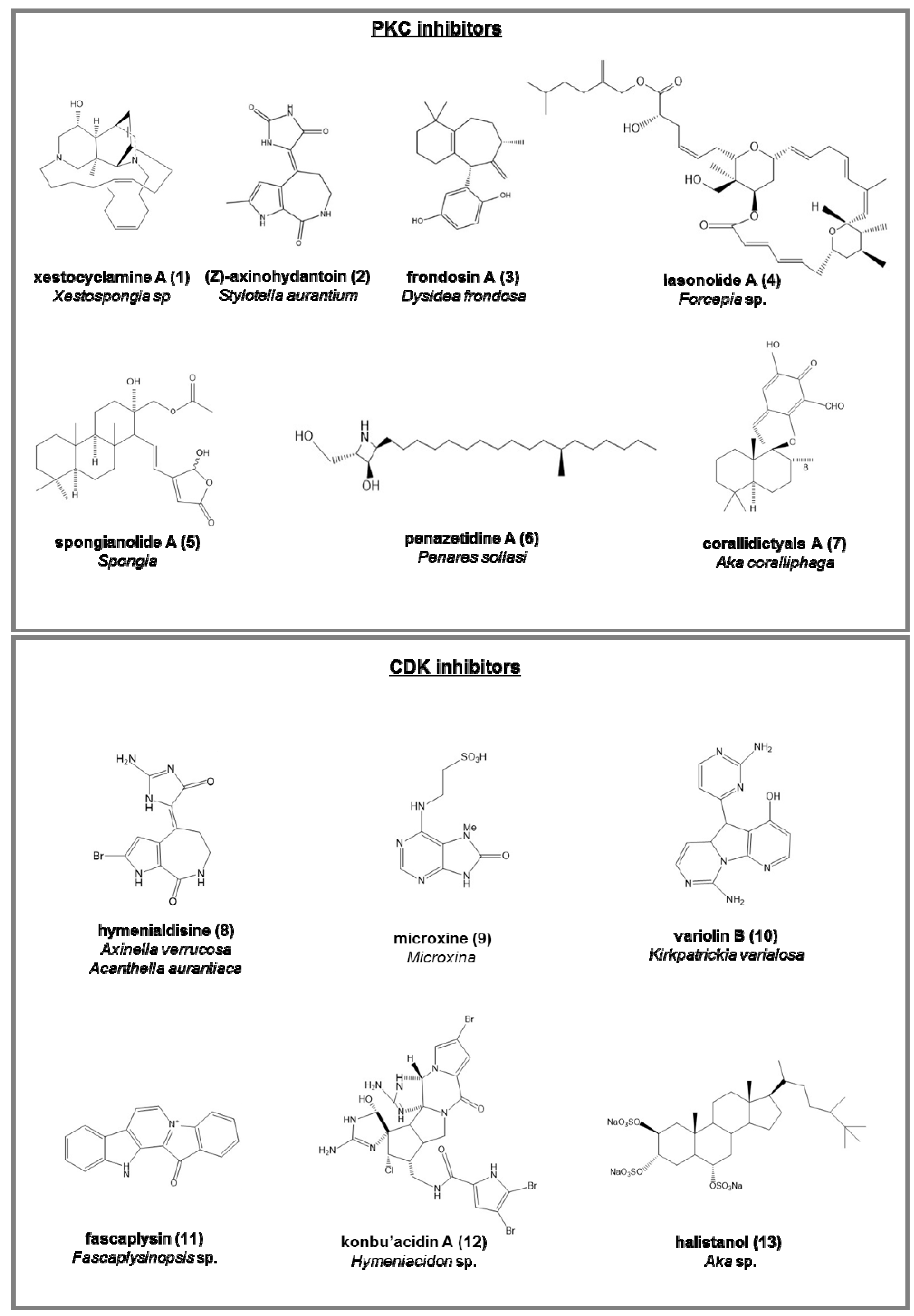
Figure 3.
Chemical structure of some natural TPK and EGFR inhibitors isolated from sponges, reported as examples.
Figure 3.
Chemical structure of some natural TPK and EGFR inhibitors isolated from sponges, reported as examples.
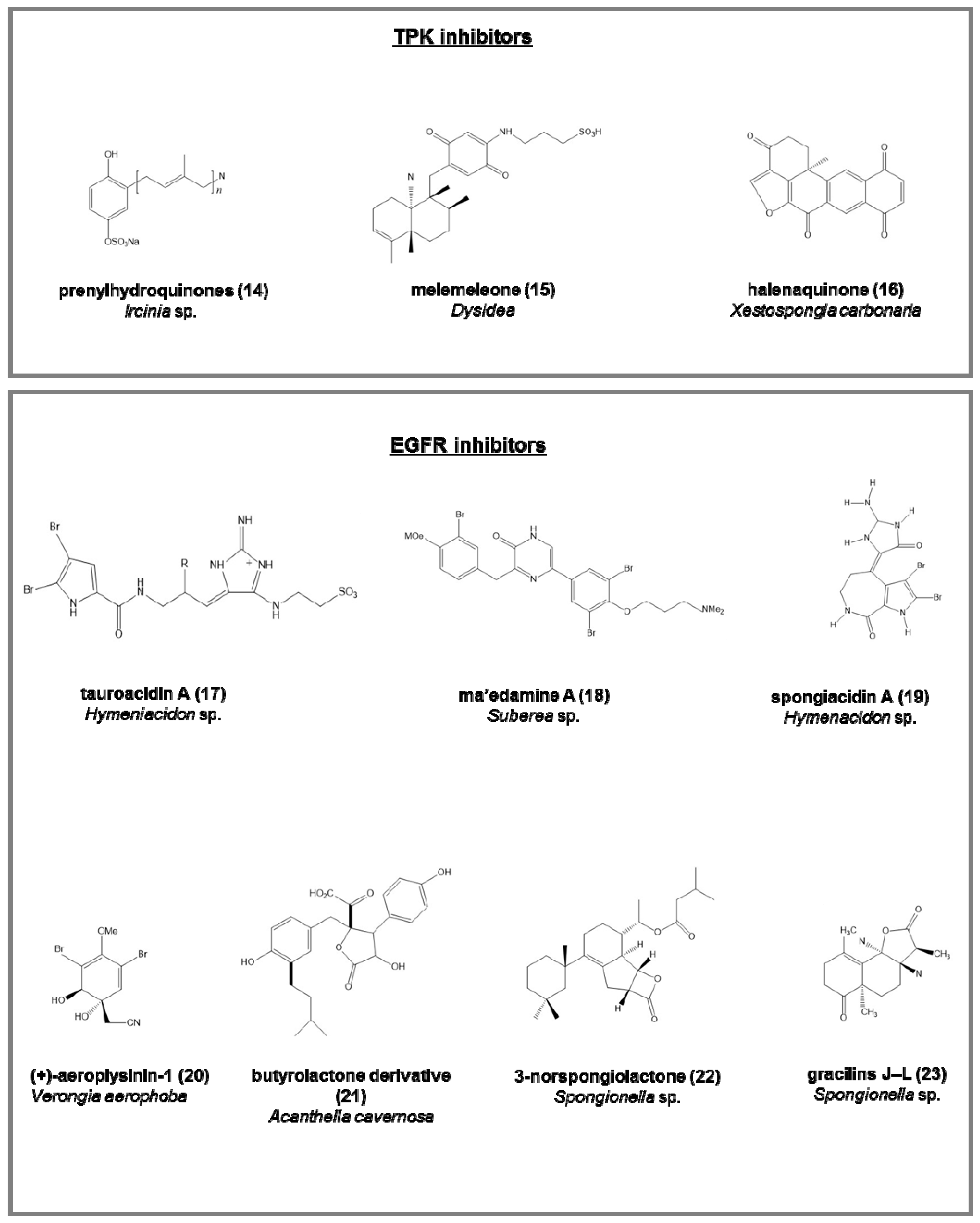
Figure 4.
Chemical structure of some natural mitogen-activated protein kinase (MK) and GSK-3 inhibitors and other kinases inhibitors isolated from sponges, reported as examples.
Figure 4.
Chemical structure of some natural mitogen-activated protein kinase (MK) and GSK-3 inhibitors and other kinases inhibitors isolated from sponges, reported as examples.

Figure 5.
Enzyme inhibitors isolated from marine bacteria.
Figure 6.
Examples of chemical structure of some enzyme inhibitors isolated from marine bacteria.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Kinase enzyme, enzyme inhibitors names (together with number compounds in the case the chemical structure have been reported in Figure 2, Figure 3, Figure 4 and Figure 5), references, pharmacological applications, and IC50 values (in micromolar, μM) from marine sponges: the four less potent enzyme inhibitors were indicated with (*), the five more potent with (**).
Table 1.
Kinase enzyme, enzyme inhibitors names (together with number compounds in the case the chemical structure have been reported in Figure 2, Figure 3, Figure 4 and Figure 5), references, pharmacological applications, and IC50 values (in micromolar, μM) from marine sponges: the four less potent enzyme inhibitors were indicated with (*), the five more potent with (**).
| Kinase Enzyme | Compound | Reference | Pharmacological Application | IC50 (μM) |
|---|---|---|---|---|
| PKC | ||||
| xestocyclamine A (1) | [42,43,44] | anticancer | 10 | |
| (Z)-Axinohydantoin (2) | [45] | 9 | ||
| frondosin A (3) | [46] | HIV virus | 1.8 | |
| frondosin B | [46] | HIV virus | 4.8 | |
| frondosin C | [46] | HIV virus | 20.9 | |
| frondosin D | [46] | HIV virus | 26 | |
| frondosin E | [46] | HIV virus | 30.6 | |
| nakijiquinones A | [48,49] | anticancer | 270 * | |
| nakijiquinones B | [48,49] | anticancer | 200 * | |
| nakijiquinones C | [48,49] | anticancer | 23 | |
| nakijiquinones D | [48,49,50] | anticancer | 220 * | |
| lasonolide A (4) | [51,52,53] | thymoma cells | 0.03 ** | |
| spongianolides A (5) –E | [54] | 20–30 | ||
| penazetidine A (6) | [55] | anticancer | 1 | |
| corallidictyals A (7) –B | [56] | 28 | ||
| CDK | ||||
| hymenialdisine (8) | [57] | rheumatoid arthritis | 0.02 ** | |
| microxine (9) | [58] | 13 | ||
| variolin B (10) | [59,60] | antiviral, anticancer | 0.03 | |
| fascaplysin (11) | [61] | anticancer, angiogenesis | 0.4 | |
| konbu’acidin A (12) | [62] | anticancer | 20 | |
| halistanol (13) | [63] | anticancer | 0.013 ** | |
| penta-prenylhydroquinone 4-sulfates (14) | [64] | antiviral, anticancer | 8 | |
| hexa-prenylhydroquinone 4-sulfates | [64] | antiviral, anticancer | 4 | |
| hepta-prenylhydroquinone 4-sulfates | [64] | antiviral, anticancer | 8 | |
| melemeleone (15) | [65] | anticancer | 28 | |
| halenaquinone (16) | [66] | anticancer | 1.5 | |
| EGFR | ||||
| tauroacidin A (17) | [67] | anticancer | 0.001 ** | |
| ma’edamine A (18) | [68] | anticancer | 11 | |
| spongiacidin A (19) | [69] | 8.5 | ||
| spongiacidin B | [69] | 6 | ||
| (+)-aeroplysinin-1 (20) | [70] | anticancer | 0.25–0.5 | |
| butyrolactone derivative (21) | [71] | anticancer | 22.9 | |
| 3-norspongiolactone (22) | [72] | anticancer | 0.6–15 | |
| gracilins J–L (23) | [73] | anticancer | 0.6–15 | |
| MK | ||||
| cheilanthane (24) | [74] | anticancer | 4 | |
| hymenin (25) | [75] | 128.8–250 * | ||
| hymenialdisine (27) | [76] | antitumor | 0.003–0.006 ** | |
| onnamide A (28) | [77] | anti-inflammatory | 30 | |
| (+)-makassaric acid (26) | [78] | anti-inflammatory | 20 | |
| (+)-subersic acid (29) | [78] | anti-inflammatory | 9.6 | |
| GSK-3 | ||||
| manzamine A (30) | [79,80] | Alzheimer’s disease | 10.2 | |
| Other kinases inhibitors | ||||
| liphagal (31) | [81] | anticancer | 0.1 | |
| (+)-curcuphenol (32) | [82] | 36 | ||
| homogentisic acid (34) | [83] | antimalarial | 1.8 | |
Table 2.
Enzyme, enzyme inhibitors names (together with number compounds in the case the chemical structure have been reported in Figure 6), references, biotechnological applications, and IC50 values (in micromolar, μM) from marine bacteria.
Table 2.
Enzyme, enzyme inhibitors names (together with number compounds in the case the chemical structure have been reported in Figure 6), references, biotechnological applications, and IC50 values (in micromolar, μM) from marine bacteria.
| Kinase Enzyme | Compound | Reference | Application | IC50 (μM) |
|---|---|---|---|---|
| Serine protease | marinostatins C1–C2 | [86,87] | pancreatitis pathogenesis | 1.0–3.2 |
| Monoamine oxidase | 2,3-indolinedone (35) | [88] | neurodegenrative diseases | 9.2 |
| Protease | monostatin | [89,90] | cooked fish meat gel | |
| ECE | B-90063 (36) | [92] | hypertension, renal disease | 1.0–3.2 |
| GluNAc-ase | pyrostatins A–B (37,38) | [93,94,95,96] | diabetes, leukemia, cancer | 1 |
| PGP | pyrizinostatin (40) | [97,98] | hormone diseases | 21 |
| Chitinase | CI-4 (39) | [99,100,101] | antifungal and insecticidal | |
| Xanhine oxidase | hydroxyakalone (41) | [102,103] | uric acid accumulation | 4.6 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ruocco, N.; Costantini, S.; Palumbo, F.; Costantini, M. Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications. Mar. Drugs 2017, 15, 173. https://doi.org/10.3390/md15060173
AMA Style
Ruocco N, Costantini S, Palumbo F, Costantini M. Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications. Marine Drugs. 2017; 15(6):173. https://doi.org/10.3390/md15060173
Chicago/Turabian StyleRuocco, Nadia, Susan Costantini, Flora Palumbo, and Maria Costantini. 2017. "Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications" Marine Drugs 15, no. 6: 173. https://doi.org/10.3390/md15060173
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.