
New Peptides Isolated from Marine Cyanobacteria, an Overview over the Past Decade


1
School of Marine Sciences, Laboratory of Marine Natural Products, Ningbo University, Ningbo 315211, China
2
Key Laboratory of Applied Marine Biotechnology of Ministry of Education, Ningbo University, Ningbo 315211, China
*
Author to whom correspondence should be addressed.
Mar. Drugs 2017, 15(5), 132; https://doi.org/10.3390/md15050132
Submission received: 15 March 2017
/
Revised: 28 April 2017
/
Accepted: 2 May 2017
/
Published: 5 May 2017
Abstract
:Marine cyanobacteria are significant sources of structurally diverse marine natural products with broad biological activities. In the past 10 years, excellent progress has been made in the discovery of marine cyanobacteria-derived peptides with diverse chemical structures. Most of these peptides exhibit strong pharmacological activities, such as neurotoxicity and cytotoxicity. In the present review, we summarized peptides isolated from marine cyanobacteria since 2007.
1. Introduction
Cyanobacteria, as some of the oldest aquatic and photosynthetic oxygenic prokaryotes, are widely distributed in the world [1]. In recent years, cyanobacteria from different habitats, particularly marine cyanobacteria, are found to contain a great deal of bioactive secondary metabolites. As chemical defenses, secondary metabolites from marine cyanobacteria can improve adaptability of marine cyanobacteria to various marine environments, which are characterized by hyperhaline, high-pressure, barren and complexity. These secondary metabolites from marine cyanobacteria not only greatly affect the growth and reproduction of cyanobacteria, but also show many biological activities, such as anti-tumor, antibacterial, enzyme inhibition, parasitic resistance, anti-inflammatory and other biological activities [2]. Therefore, they have attracted extensive attention of scholars in different subject fields, such as medicinal chemistry, pharmacology and marine chemical ecology. It is considered that more drug lead compounds can be found from marine cyanobacteria through the cooperative in-depth and systematic studies. Marine cyanobacteria-derived bioactive components have gained great popularity in research on marine natural products [3]. For example, dolastatin 10 analogue soblidotin (or TZT-1027, auristatin PE) has shown a good prospect in human colonic carcinoma, which has been developed to the phase II clinical trials [4]. Brentuximab vedotin (trade name Adcetris), a marine peptide-derived drug, was approved by the U.S. Food and Drug Administration (FDA) in 2011 for cancer treatment [5]. In the past 10 years (from 2007 to 2016), the programs for drug discovery from marine cyanobacteria, such as Panama International Cooperative Biodiversity Group (ICGB) program, have discovered more than 400 new natural compounds from marine cyanobacteria. Among these compounds, peptides and peptide-containing compounds are the major secondary metabolites.
In 2006, secondary metabolites from marine cyanobacteria were reviewed [6]. Biological targets and the mechanisms of action of bioactive natural products from marine cyanobacteria were also reviewed in 2015 [7]. In the present review, we systematically reviewed the current progress on the discovery of peptides and peptide-containing compounds from marine cyanobacteria since 2007. By the end of 2016, a total of 126 new peptide-compounds have been isolated from marine cyanobacteria, mainly from the genera Lyngbya, Oscillatoria and Symploca. However, two new genera Moorea and Okeania, which were previously identified as the polyphyletic cyanobacterial genus Lyngbya, have been proposed using genome sequence analysis in the past few years [8,9]. Another new genus Caldora, which was previously identified as Symploca, has also been proposed [10]. New peptide compounds, which were actually isolated from these new genera of cyanobacteria, were also mentioned in this review. Most of these peptides with chemical diversity exhibit strong biological activities, such as neurotoxicity and cytotoxicity. In addition, cyclic depsipeptides, including 76 compounds, are the main cyclic peptides discovered from marine cyanobacteria.
2. Linear Peptides
Linear peptides are common compounds isolated from marine cyanobacteria, most of which exhibit prodigious biological activities, such as anti-tumor, antimicrobial, antimalarial, enzyme inhibition and other biological activities [11]. Table 1 and Table 2 summarize 39 linear peptides derived from marine cyanobacteria, covering literature from January 2007 to December 2016.
2.1. Linear Depsipeptides
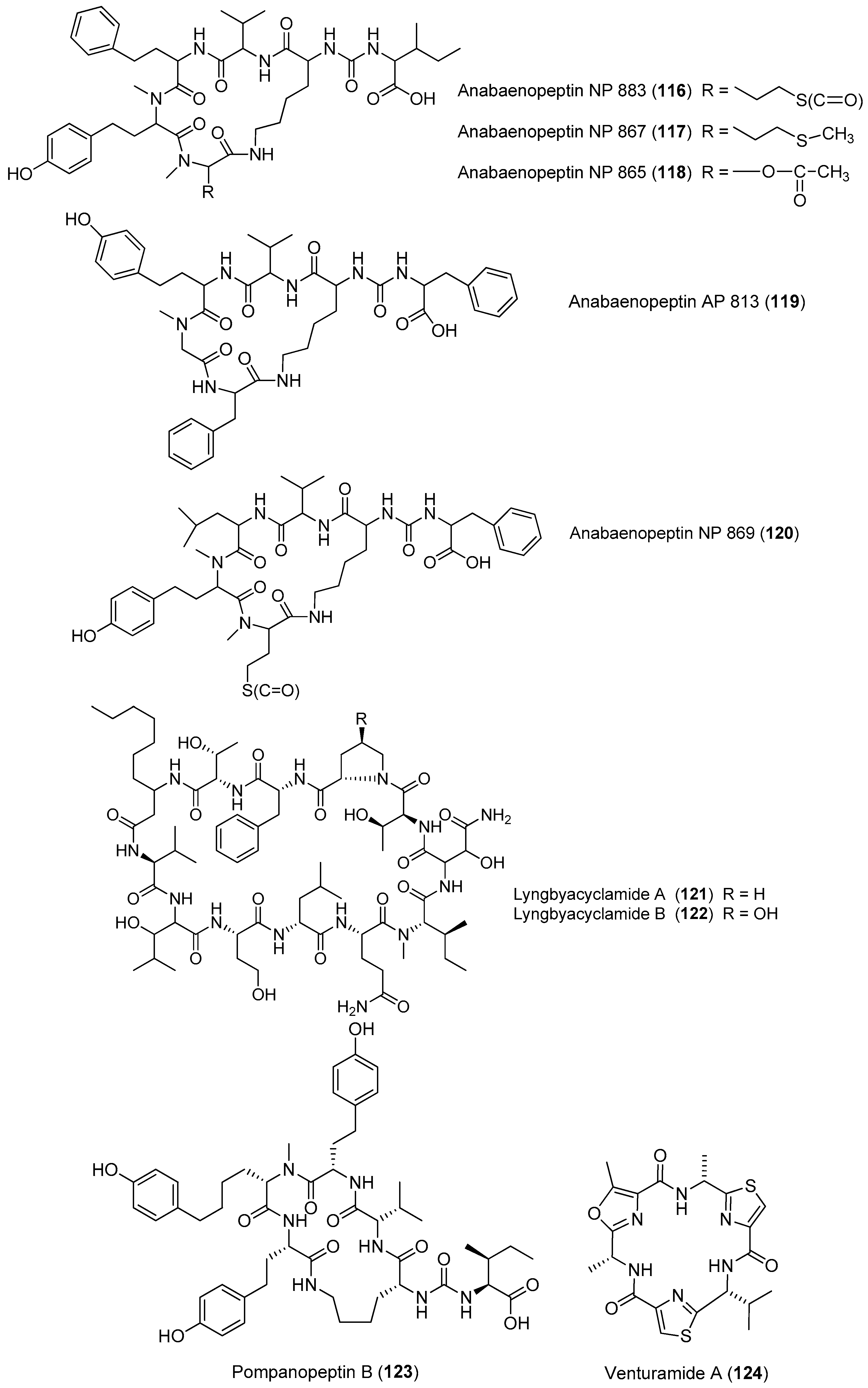
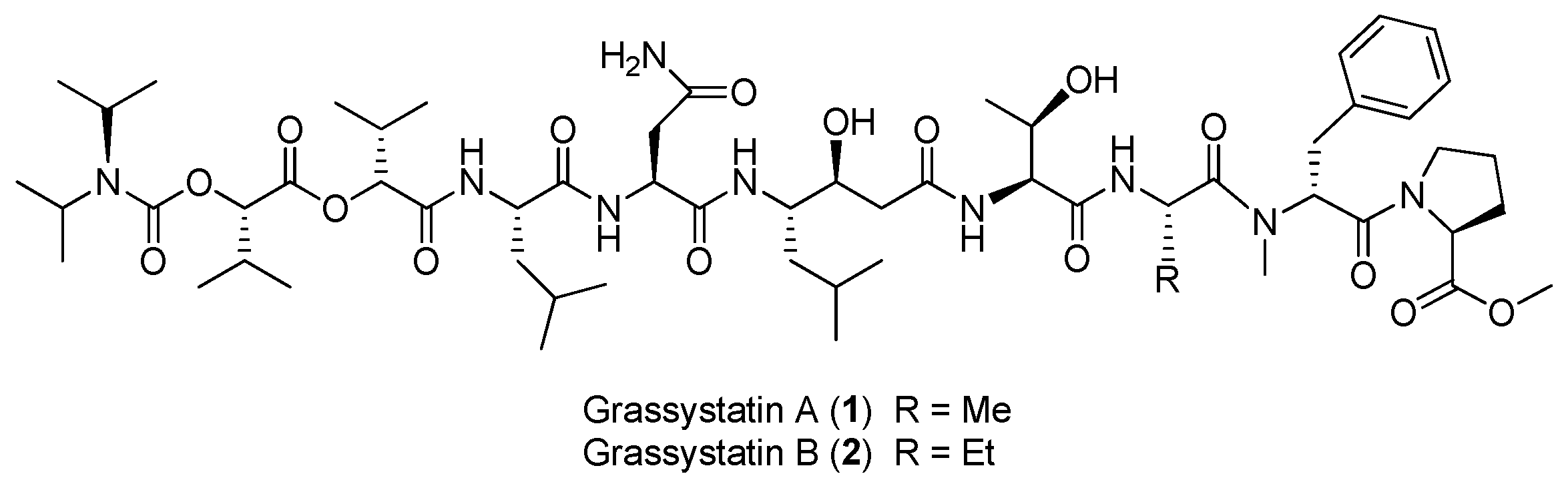
Two linear depsipeptides, grassystatins A and B (1, 2), have been purified from the marine cyanobacterium Okeania lorea (formerly Lyngbya cf. confervoides) collected from Key Largo [9], Florida (Figure 1). Compound 1 displays selectivity against cathepsins D and E with IC50 values of 26.5 nM and 886 pM, respectively. Compound 2 can also selectively inhibit cathepsins D and E with IC50 values of 7.27 nM and 354 pM, respectively. Selective inhibition of 1–2 against cathepsin E over cathepsin D (20- to 38-fold) suggests that these bioactive compounds 1 and 2 are useful tools to probe cathepsin E function [12]. Moreover, the total synthesis of 1 has been completed [13].
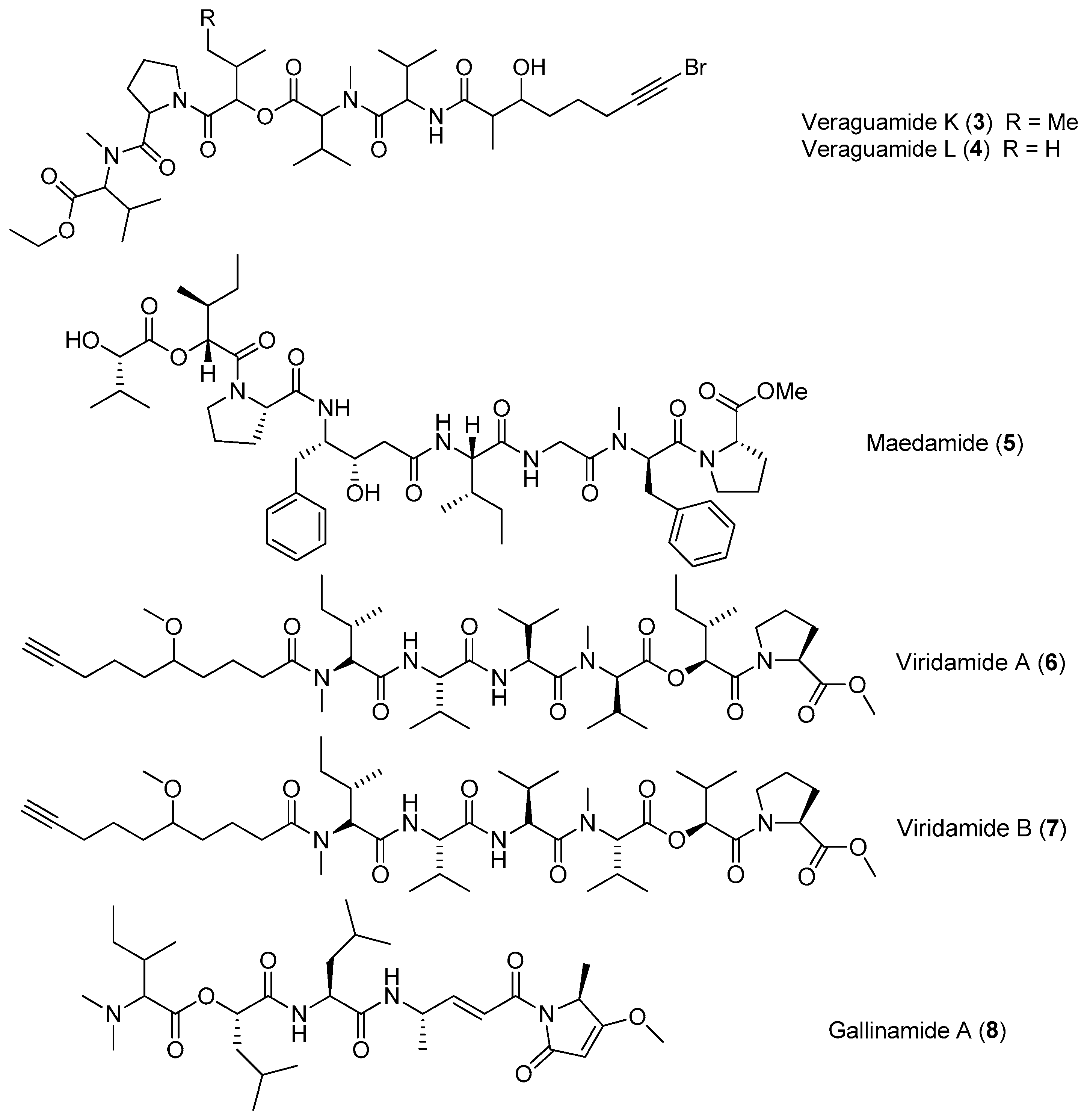
Two bromide-containing linear depsipeptides, veraguamides K–L (3–4), have been discovered from marine cyanobacterium cf. Oscillatoria margaritifera collected from Coiba Island National Park, Panama, which are supposed to exhibit the structural characteristics of marine natural products [14]. Maedamide (5) has been extracted from marine cyanobacterial assemblage of Lyngbya sp., which shows strong and selective inhibition against chymotrypsin (IC50 value of 45 μM), but not against elastase or trypsin. Moreover, compound 5 inhibits the growth of Hela cells and HL60 cells (IC50 values of 4.2 and 2.2 μM, respectively) and induces apoptosis in Hela cells [15]. The total synthesis of 5 has been achieved, leading to reassignment of the structure of 5 [16].
Two PKS-NRPS-derived metabolites, viridamides A, B (6, 7), have been discovered from the marine cyanobacterium Okeania comitata (formerly Oscillator nigroviridis) collected from Panama [9]. Compound 6 shows anti-trypanosomal and antileishmanial activities with IC50 values of 1.1 and 1.5 μM, respectively [17]. An antimalarial peptide, termed gallinamide A (8), has been purified from Panamanian marine cyanobacteria, showing moderate antimalarial activity against chloroquine-resistant strain (W2) of Plasmodium falciparum (IC50 = 8.4 μM) [18]. Total synthesis of compound 8 has been completed [19].
2.2. Other Linear Peptides
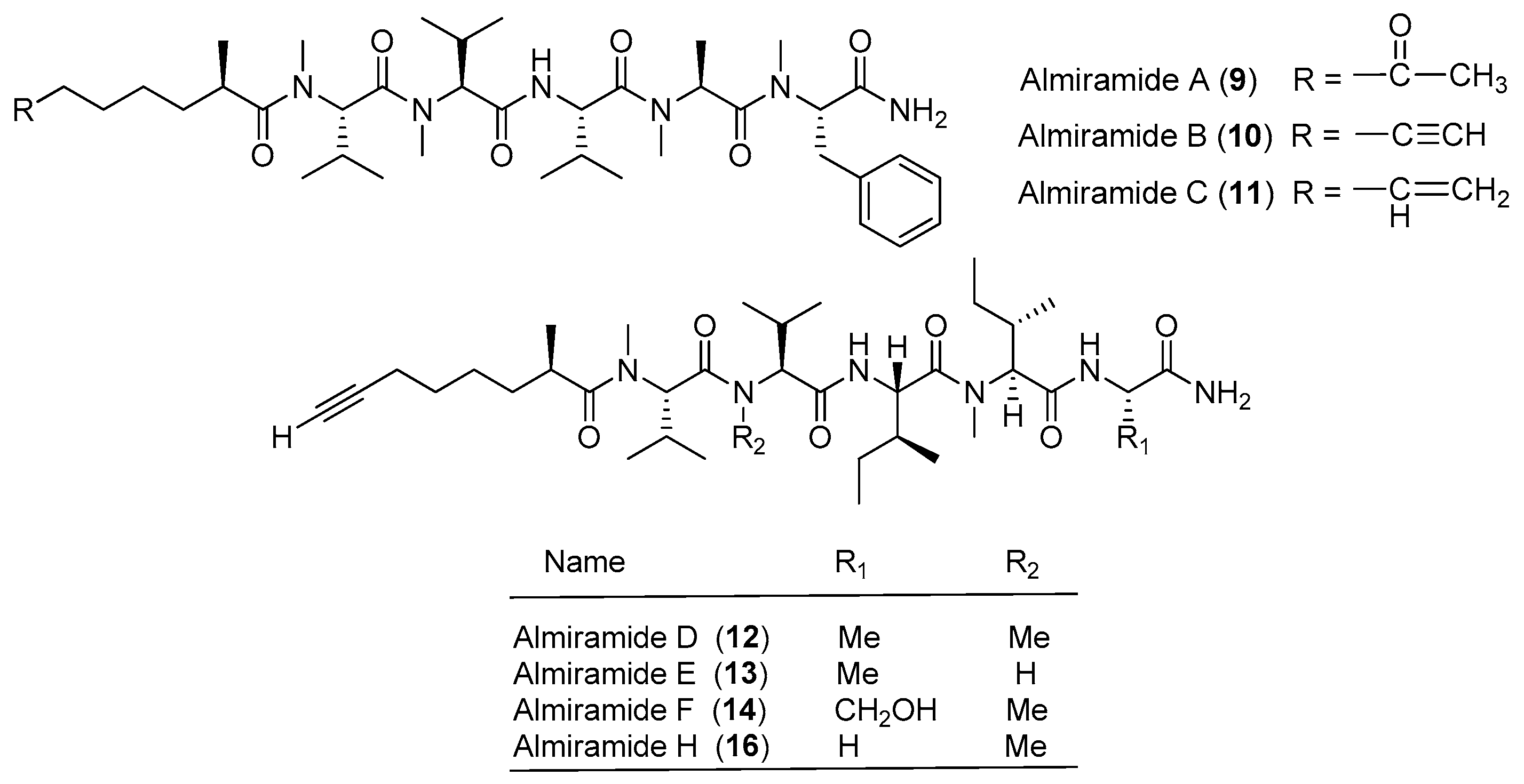
Three highly N-methylated linear lipopeptides, almiramides A–C (9–11), have been identified from screening of the marine cyanobacterium Lyngbya majuscula collected from Panama for antiparasitic activities against Leishmaniasis donovani (Figure 2). Compounds 9, 10 and 11 display strong antileishmanial activity with IC50 values of 13.5, 2.4 and 1.9 μM, respectively [20].
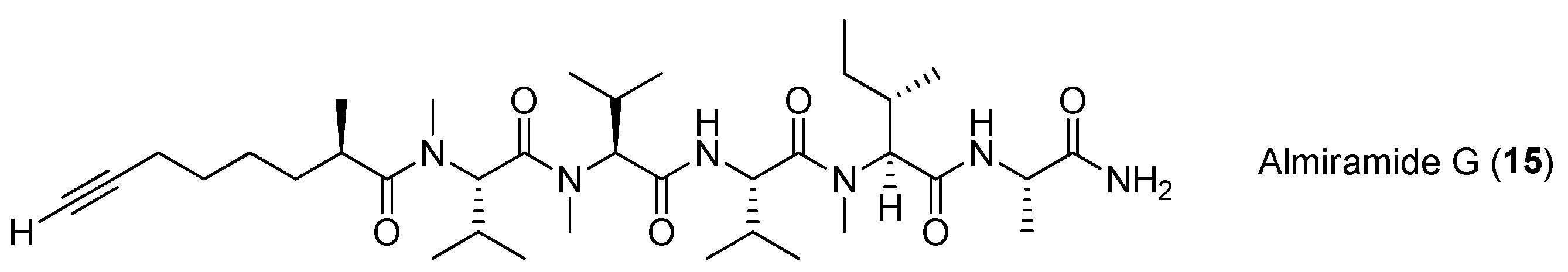
Five analogues of compound 9, almiramides D–H (12–16), have been derived from the marine cyanobacterium Oscillatoria nigroviridis collected from the Providence Island, Colombian Caribbean Sea. Compounds 10 and 12 exhibit mild toxicity against five human tumor cell lines (A549, MCF-7, HeLa, PC3 and MDA-MB231) and high toxicity against the gingival fibroblast cell line [21].
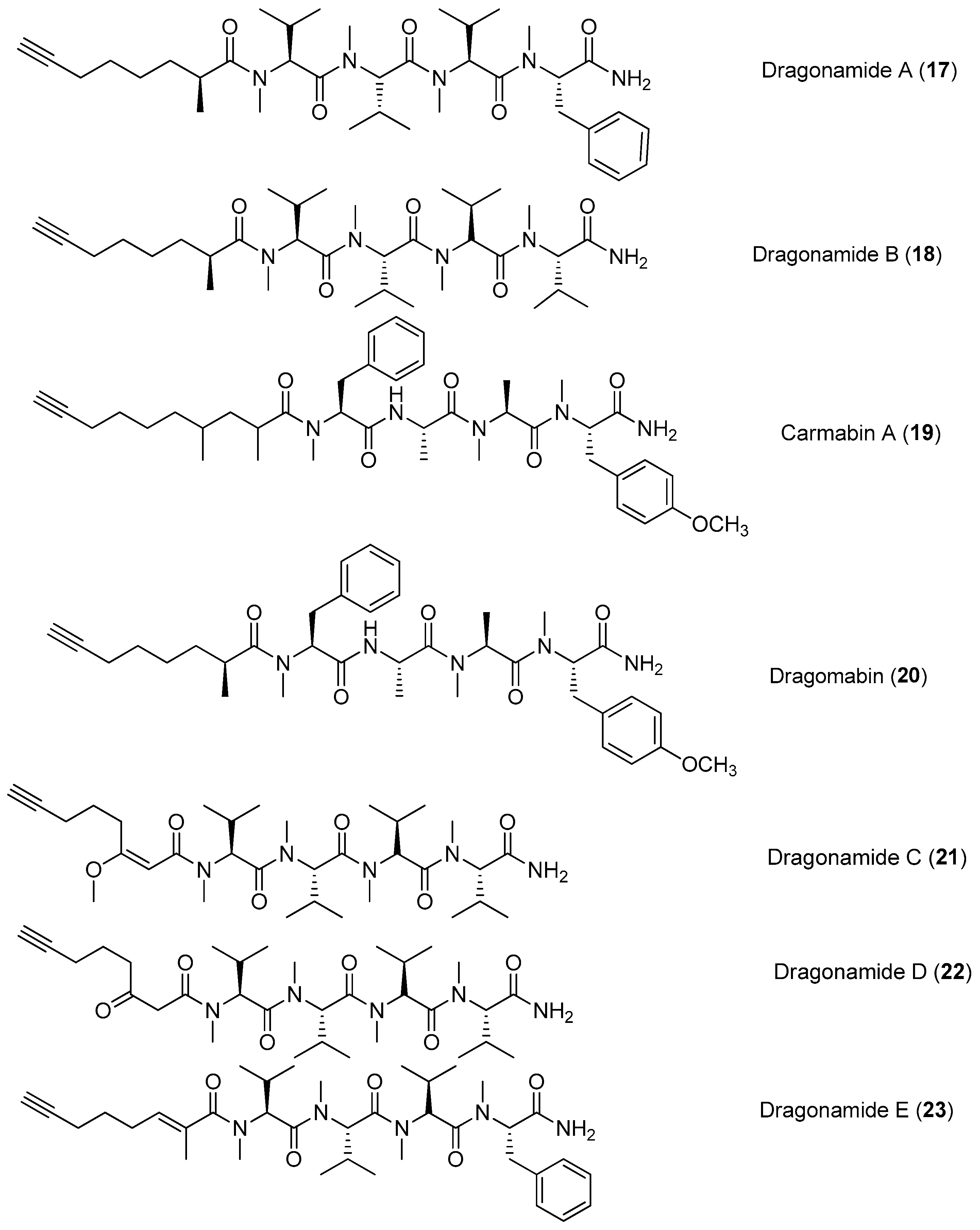
Four lipopeptides, named dragonamides A and B (17, 18), carmabin A (19) and dragomabin (20), have been identified from the antimalarial bioassay-guided isolation of the marine cyanobacterium Moorea producens (formerly Lyngbya majuscula) (Figure 3). Compounds 17, 19 and 20 exhibit good antimalarial activity (IC50 = 7.7, 4.3 and 6.0 μM, respectively) [8,22]. Two analogs of 17, named dragonamides C and D (21, 22), have been isolated from the marine cyanobacterium Moorea producens (formerly Lyngbya polychroa) collected from Hollywood Beach, Fort Lauderdale, Florida. Compounds 21 and 22 display weak cytotoxicity in cancer cell viability assays [23,24]. A new antimalarial peptide, termed dragonamide E (23), has been purified from the marine cyanobacterium Lyngbya majuscula and shows antileishmanial activity with an IC50 value of 5.1 μM [25].
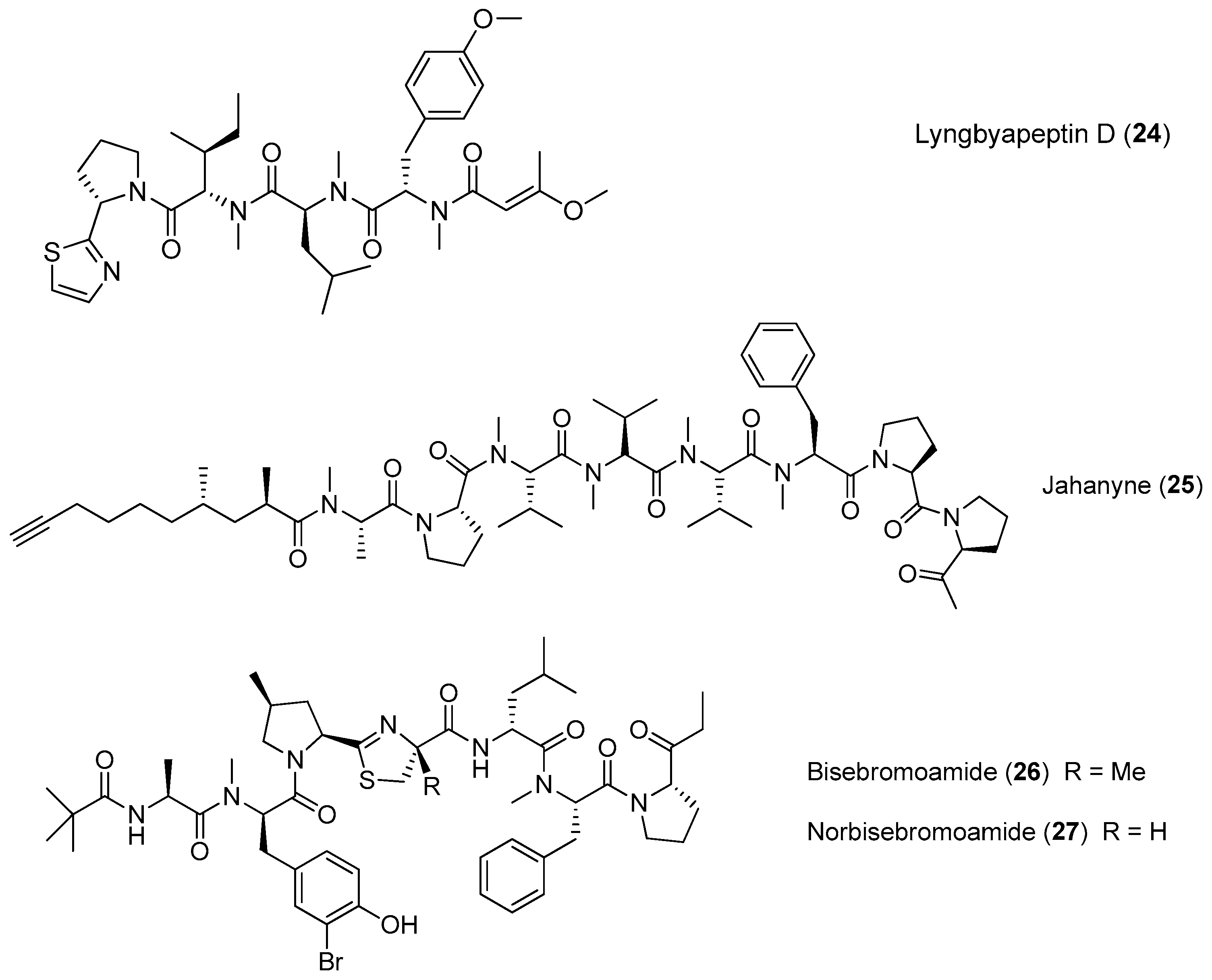
A linear lipopeptide, lyngbyapeptin D (24), has been purified from the marine cyanobacterium Moorea bouillonii (formerly Lyngbya bouillonii) collected from Apra Harbor, Guam [8,26]. Jahanyne (25), isolated from Lyngbya sp. collected in Okinawa, shows significant inhibitory effects on the growth of human cancer cells in vitro, and it can induce apoptosis of HeLa cells [27]. Two novel cytotoxic peptides, named bisebromoamide (26) and norbisebromoamide (27), have been identified from the marine cyanobacterium Lyngbya sp. (Figure 4). The rare peptide 26 possesses the combination of unusual structural features, including an N-pivalamide moiety, high degree of D-amino acids, N-methylated amino acids and several other modified amino acid residues of nonribosomal origin. Compound 26 exhibits cytotoxicity against HeLa S3 cells (IC50 = 0.04 μg/mL). Compound 26 can also inhibit phosphorylation of ERK (extracellular signal regulated protein kinase) in NRK cells, showing potent and selective inhibitory effects on protein kinase [28,29]. A revised configurational assignment for the marine peptide 26 has been proposed and validated by total synthesis [30,31].
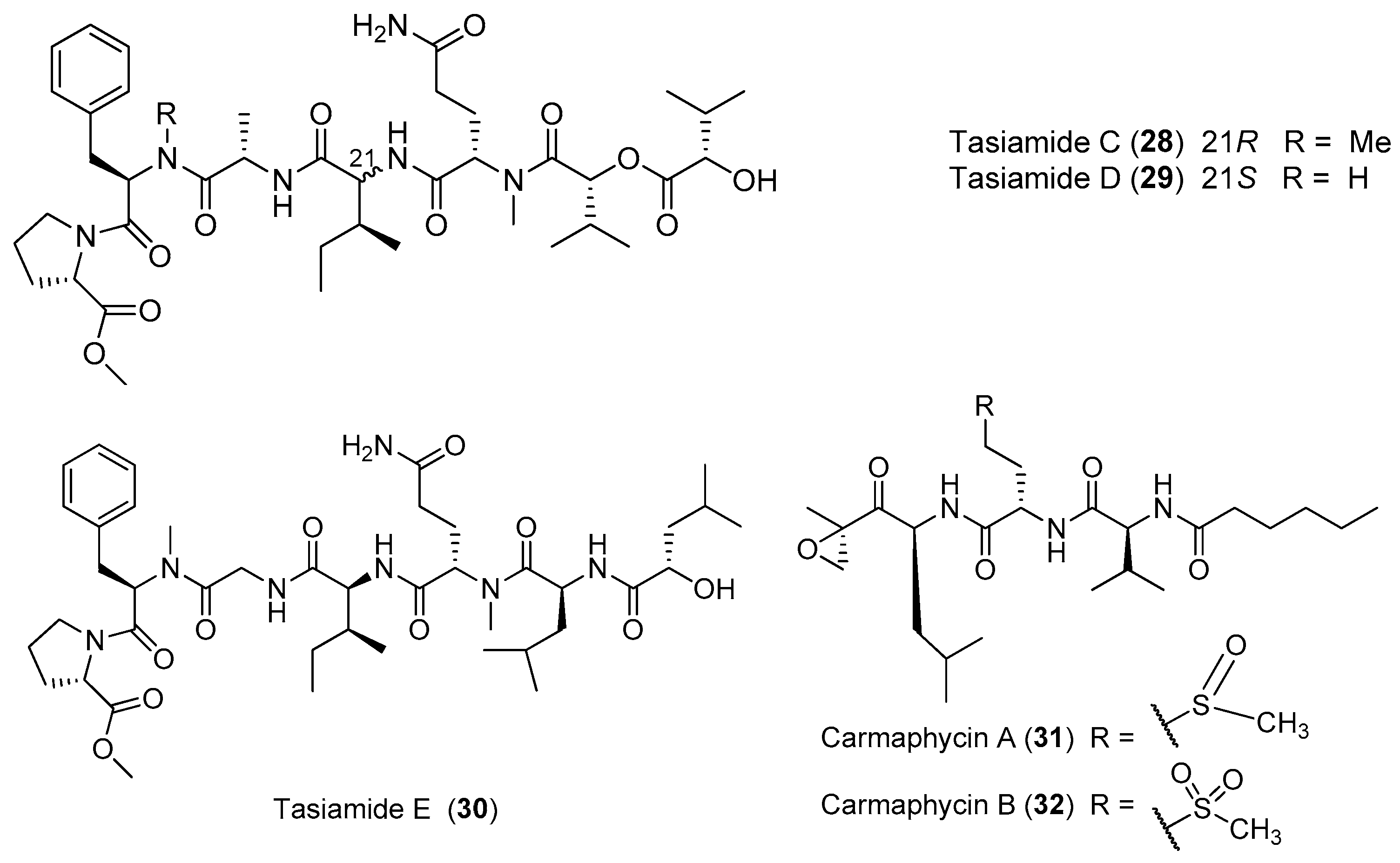
Three new lipopeptides, tasiamides C–E (28–30), have been derived from the tropical marine cyanobacterium Symploca sp. collected near Kimbe Bay, Papua New Guinea (Figure 5). The structural features of 28–30 are similar to some previously isolated peptides from the same marine cyanobacterium Symploca sp., such as tasiamides, grassystatins and symplocin [32]. Two novel proteasome inhibitors, carmaphycins A and B (31, 32), have been extracted from the marine cyanobacterium Symploca sp. collected from Curacao, and both of them possess a leucine-derived α, β-epoxyketone directly connected to either a sulfoxide or sulfone moiety. Compounds 31 and 32 strongly inhibit the β5 subunit of the S. cerevisiae 20S proteasome and show strong cytotoxicity against the lung and colon cancer cells. The total synthesis of 31 and 32 has been accomplished [33].
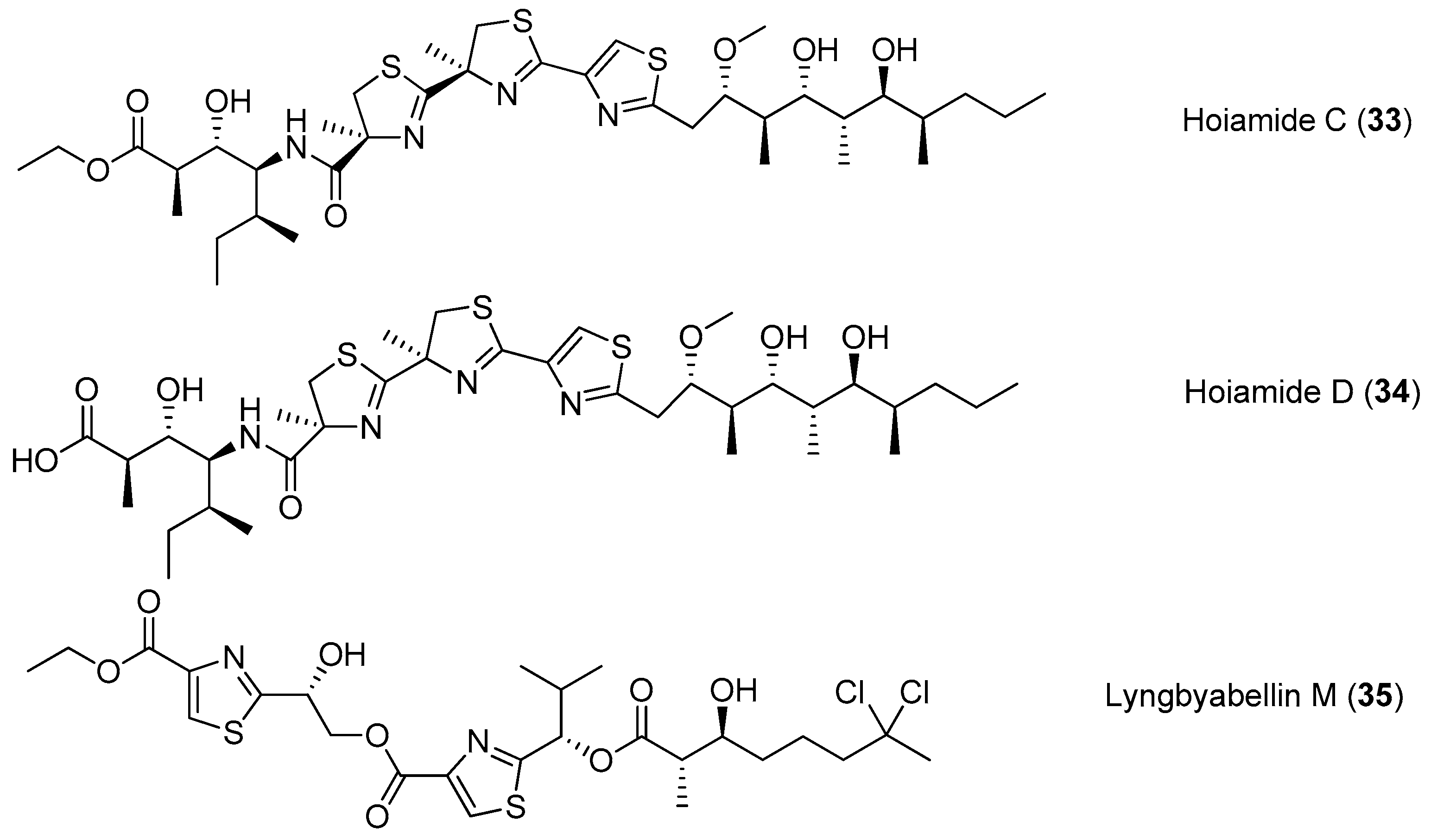
A structurally intriguing neurotoxic lipopeptide, hoiamide C (33), has been extracted from marine cyanobacteria collected in Papua New Guinea, and it possesses unique structural features of S-adenosyl methionine modified isoleucine unit, a central triheterocyclic system consisting of two R-methylated thiazolines and one thiazole moiety [34]. The total synthesis of 33 has been accomplished [35]. Hoiamide D (34), a new analogue of compound 33, has been purified from two cyanobacteria by bioassay-guided isolation (Figure 6). Compound 34 shows strong inhibitory activity against an attractive anticancer target p53/MDM2 interaction (EC50 = 4.5 μM) [36]. Another thiazole-containing lipopeptide, lyngbyabellin M (35), has been extracted from the cyanobacterium Moorea bouillonii from Palmyra Atoll, Central Pacific Ocean [37].
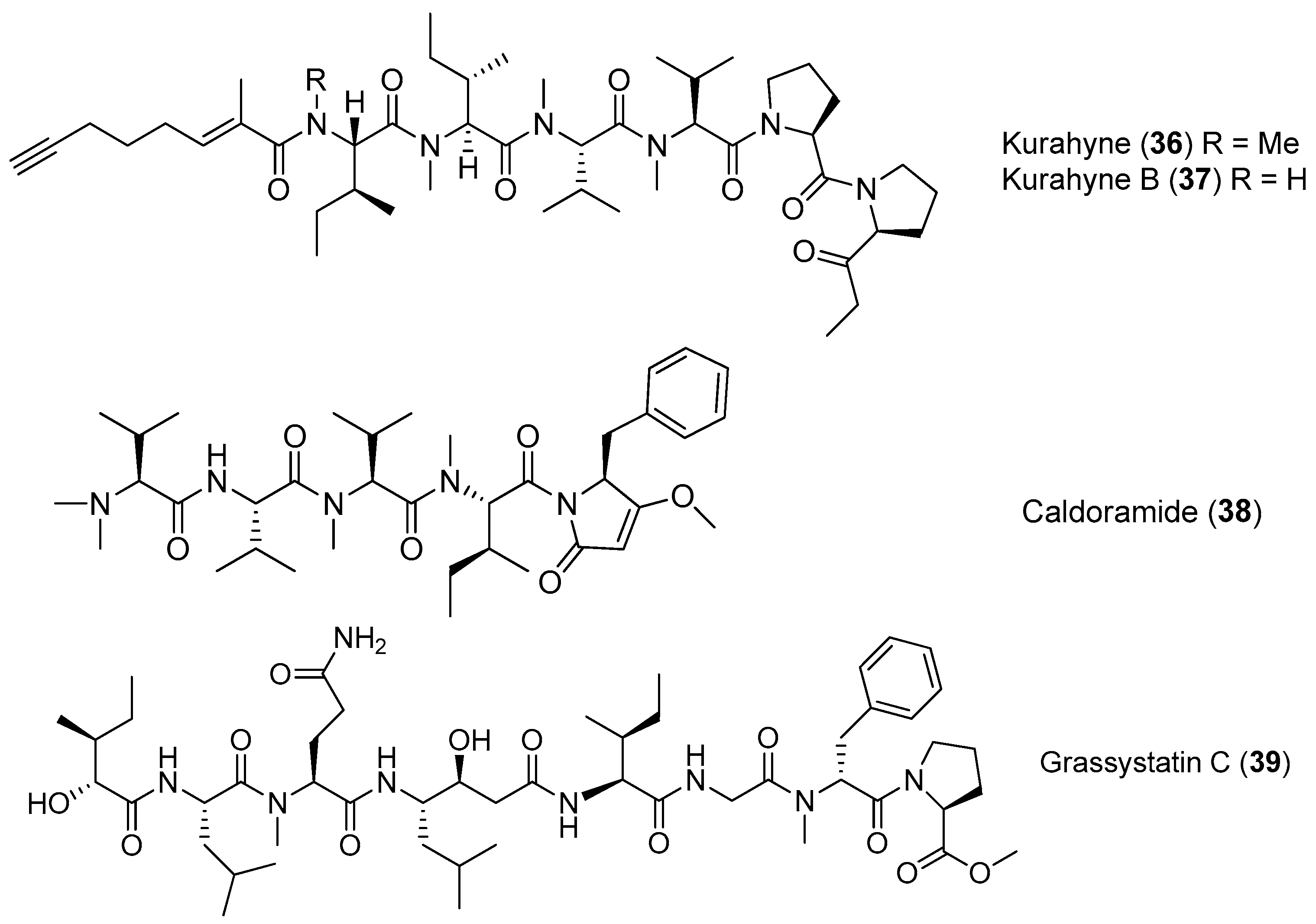
A new acetylene-containing lipopeptide, named Kurahyne (36), has been isolated from the cyanobacterial mixture consisting of Lyngbya sp. mostly. Compound 36 shows the inhibition against the growth of human cancer cells and induces the apoptosis of HeLa cells [38]. A new analogue of 36, kurahyne B (37), has been identified from the marine cyanobacterium Okeania sp. from Okinawa. Compound 37 inhibits the growth of HeLa cells and HL60 cells with IC50 values of 8.1 and 9.0 μM, respectively [39]. A cytotoxic pentapeptide caldoramide (38) has been extracted from the marine cyanobacterium Caldora penicillata from Big Pine Key, Florida (Figure 7). Compound 38 shows differential cytotoxicity against parental HCT116 colorectal cancer cells and isogenic cells lacking oncogenic KRAS or hypoxia-inducible factors 1α (HIF-1α) and 2α (HIF-2α) [40]. A linear peptide, grassystatin C (39), has been purified from the marine cyanobacterium Okeania lorea (formerly Lyngbya cf. confervoides) collected from Key Largo, Florida [9]. Compound 39, which consists of two fewer residues compared with 1 and 2, is less effective against both cathepsins D and E [12].
3. Cyclic Peptides
Cyclic peptides are a class of natural products with structural diversity and pharmacological perspective. In the past 10 years, the discovery of bioactive marine natural products from marine cyanobacteria has become a new research hotpoint in the field of marine natural products since a large number of bioactive cyclic peptides have been derived from marine cyanobacteria. The structural characteristics of cyclic peptides from marine cyanobacteria mainly manifest in tremendous diversity in new carbon skeletons, oxidation of the carbon skeletons of amino acids, complexes of holagen-containing molecules and complex spatial configuration [6]. Cyclic peptides from marine cyanobacteria can be further divided into cyclic depsipeptides, cyclic liopeptides and other cyclic peptides. A total of 87 cyclic peptides isolated from marine cyanobacteria, covering literature from January 2007 to December 2016, are summarized in Table 3, Table 4 and Table 5.
3.1. Cyclic Depsipeptides
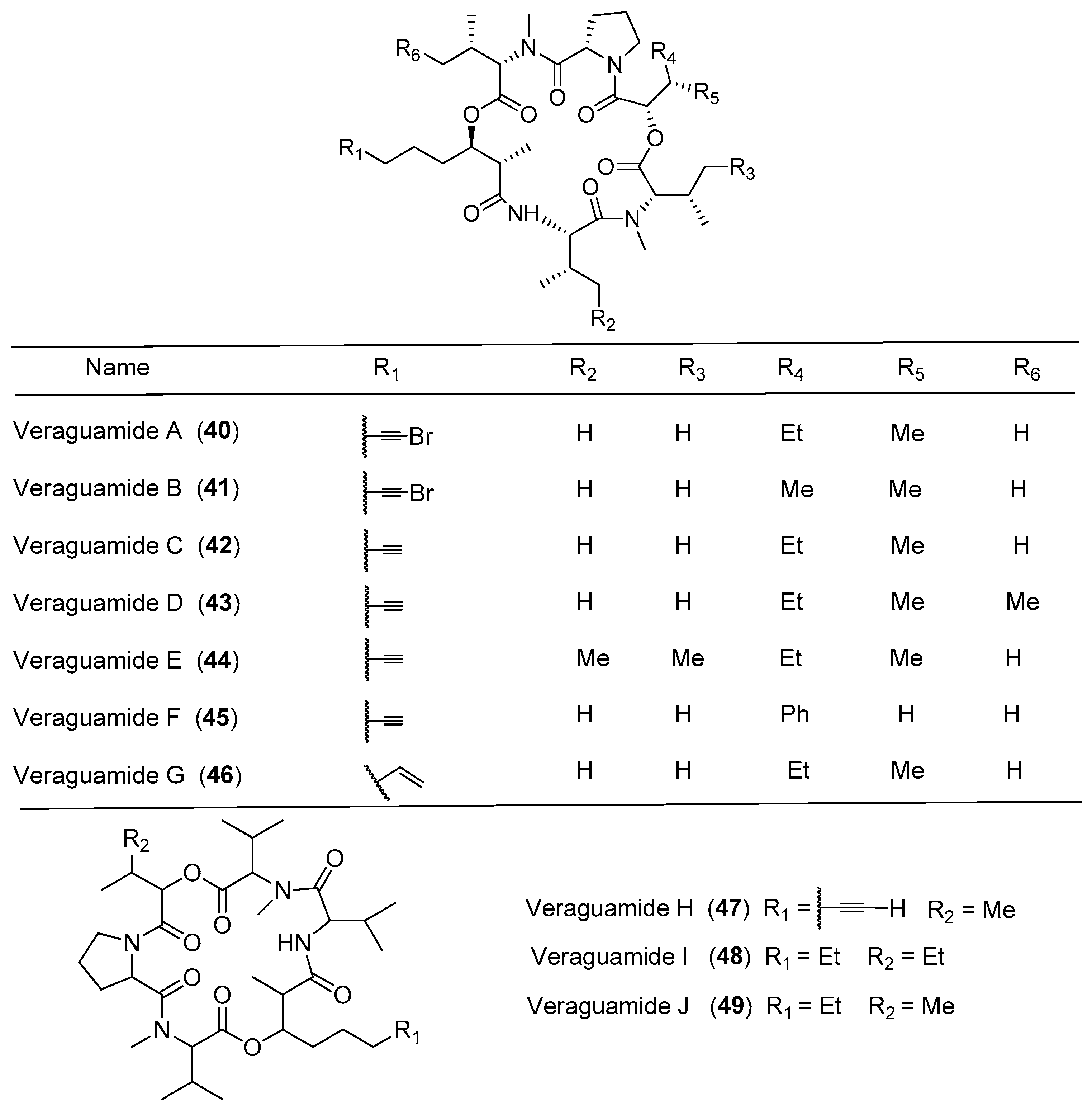
Seven new cyclic hexadepsipeptides, termed veraguamides A–G (40–46), have been identified through cytotoxicity-directed isolation of a marine cyanobacterium Symploca cf. hydnoides sample from Cetti Bay, Guam (Figure 8). Compounds 40–46 show moderate to weak cytotoxicity against HT29 colorectal adenocarcinoma and HeLa cell lines, and their cytotoxicities are determined at several sensitive positions in the veraguamide scaffold [41]. Six analogues of opunalide, 40–42 and veraguamides H–J (47–49), have been isolated from the marine cyanobacterium cf. Oscillatoria margaritifera collected from Coiba Island National Park, Pacific Panama. Compounds 40 and 41 contain bromine, conforming to the structural characteristics of the marine natural products. Compound 40 displays strong cytotoxicity to the H-460 human lung cancer cell lines with LD50 value of 141 nM [14]. The total synthesis of the proposed structure for compound 40 has been achieved, but the NMR data of the synthetic compound were significantly different from the natural product 40 [42].
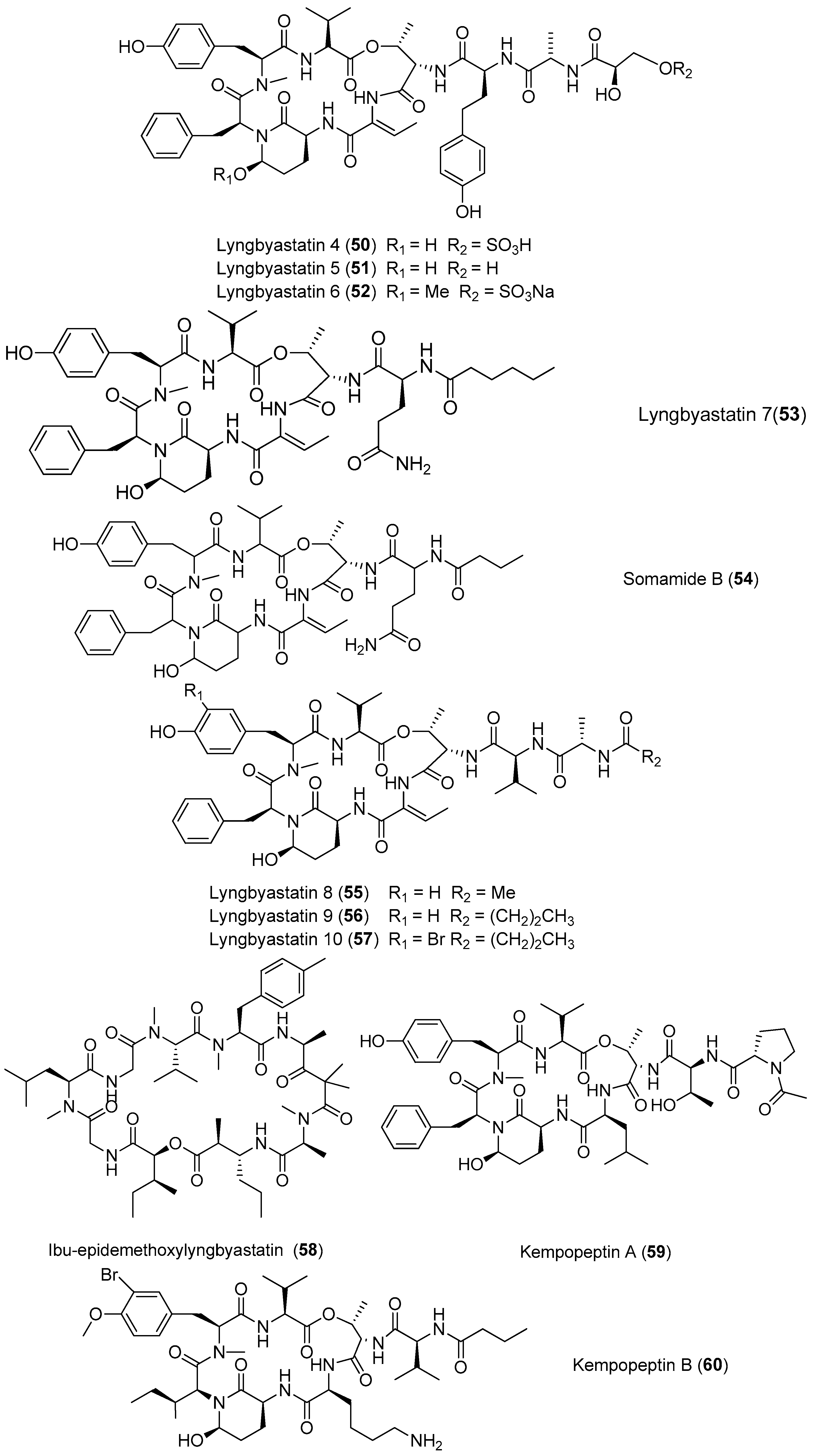
A large group of cyclic depsipeptides (lyngbyastatins) with various selectivity for elastase, chymotrypsin and trypsin has been purified from marine cyanobacteria, mainly from Lyngbya species (Figure 9). Three novel analogues of dolastatin 13, lyngbyastatin 4–6 (50–52) have been identified from the marine cyanobacterium Lyngbya confervoides from the Florida Atlantic coast and South Florida, and the presence of compounds 50–52 further supports the conclusion that cyanobacteria are the real origin of many dolastatins. Compound 50 shows potent and selective inhibitory effects on elastase as well as chymotrypsin in vitro over other serine proteases with IC50 values of 0.03 and 0.30 μM, respectively [32,43,44]. Another two novel analogues of dolastatin 13, lyngbyastatin 7 (53) and somamide B (54) have been purified from Lyngbya sp. from Florida. Compounds 51–54 show potent and selective inhibitory effects on porcine pancreatic elastase over some other serine proteases with IC50 values ranging from 3 to 10 nM [44]. The total synthesis of 53 has been completed [45]. Three cyclic depsipeptides with potent elastase inhibitory activity, termed lyngbyastatins 8–10 (55–57), have been isolated from the marine cyanobacterium Lyngbya semiplena collected in Tumon Bay, Guam. Like 50–53, compounds 55–57 show strong inhibitory activity against porcine pancreatic elastase with IC50 values of 123, 210 and 120 nM, respectively [46]. Ibu-epidemethoxylyngbyastatin 3 (58) has been purified from the marine cyanobacterium Leptolyngbya sp. from the SS Thistlegorm shipwreck in the Red Sea. Compound 58 shows weak cytotoxicity to neuro-2a cells (IC50 > 10 μM) [47]. Two analogues of lyngbyastatin, named kempopeptins A and B (59, 60), have been extracted from the marine cyanobacterium Lyngbya sp. from Florida. Compound 59 exhibits inhibitory activities against elastase and chymotrypsin (IC50 values of 0.32 and 2.6 μM, respectively), while 60 inhibits trypsin (IC50 value of 8.4 μM) [48].
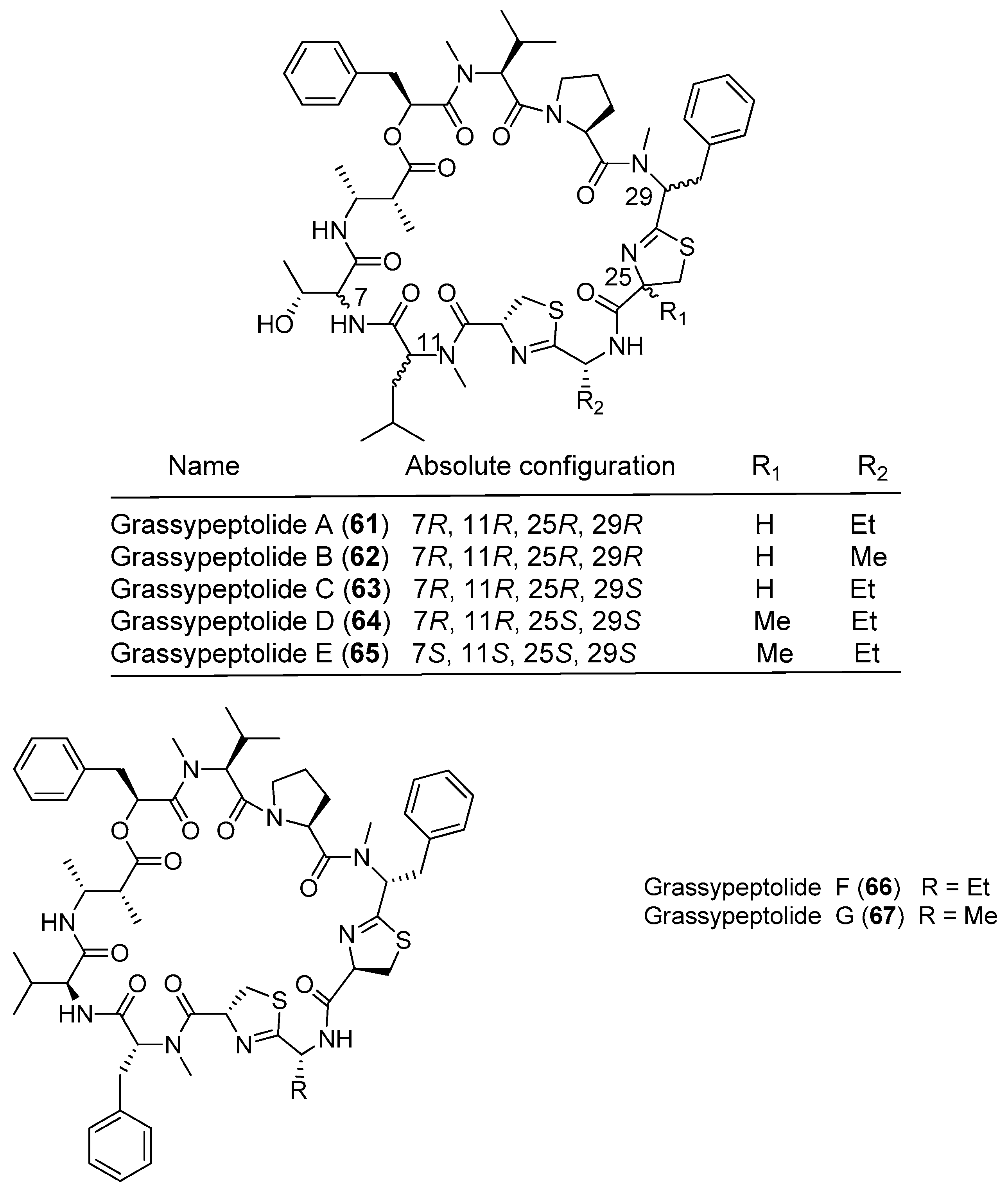
As a novel family of bis-thiazoline-containing macrocyclic depsipeptides, grassypeptolides containing d-amino acid residues and β-amino acid residues have been isolated from marine cyanobacteria (Figure 10). Grassypeptolide A (61) has been purified from the marine cyanobacterium Okeania lorea (formerly Lyngbya confervoides) off Grassy Key in Florida [9], and it inhibits the growth of four cancer cell lines with IC50 values ranging from 1.0 to 4.2 μM [49]. Two analogues of compound 61, grassypeptolides B and C (62, 63), have been extracted from the marine cyanobacterium Okeania lorea (formerly Lyngbya confervoides) from the Florida Keys [9]. The structure–activity relationship between the analogues shows that when the ethyl substituent of compound 61 is changed to a methyl substituent in 62, cytotoxic activity is only slightly reduced (3–4-fold), whereas inversion of the Phe unit flanking the bis-thiazoline moiety results in 16–23-fold greater potency. Both compounds 61 and 63 cause cell cycle arrest in G1 phase at lower concentrations, followed by G2/M phase arrest at higher concentrations, and these compounds bind Cu2+ and Zn2+ [50]. Total synthesis of compound 61 has been accomplished [51]. Grassypeptolides D and E (64, 65) have been derived from the marine cyanobacterium Leptolyngbya sp. collected from the SS Thistlegorm shipwreck in the Red Sea, and they show significant cytotoxicity against HeLa cell lines (IC50 = 335 and 192 nM, respectively) and mouse neuro-2a blastoma cells (IC50 = 599 and 407 nM, respectively) [47]. Grassypeptolides F and G (66, 67) have been purified from Palauan cyanobacterium Lyngbya majuscula, and they (66, 67) moderately inhibit the transcription factor AP-1 with IC50 values of 5.2 and 6.0 μM, respectively [52].
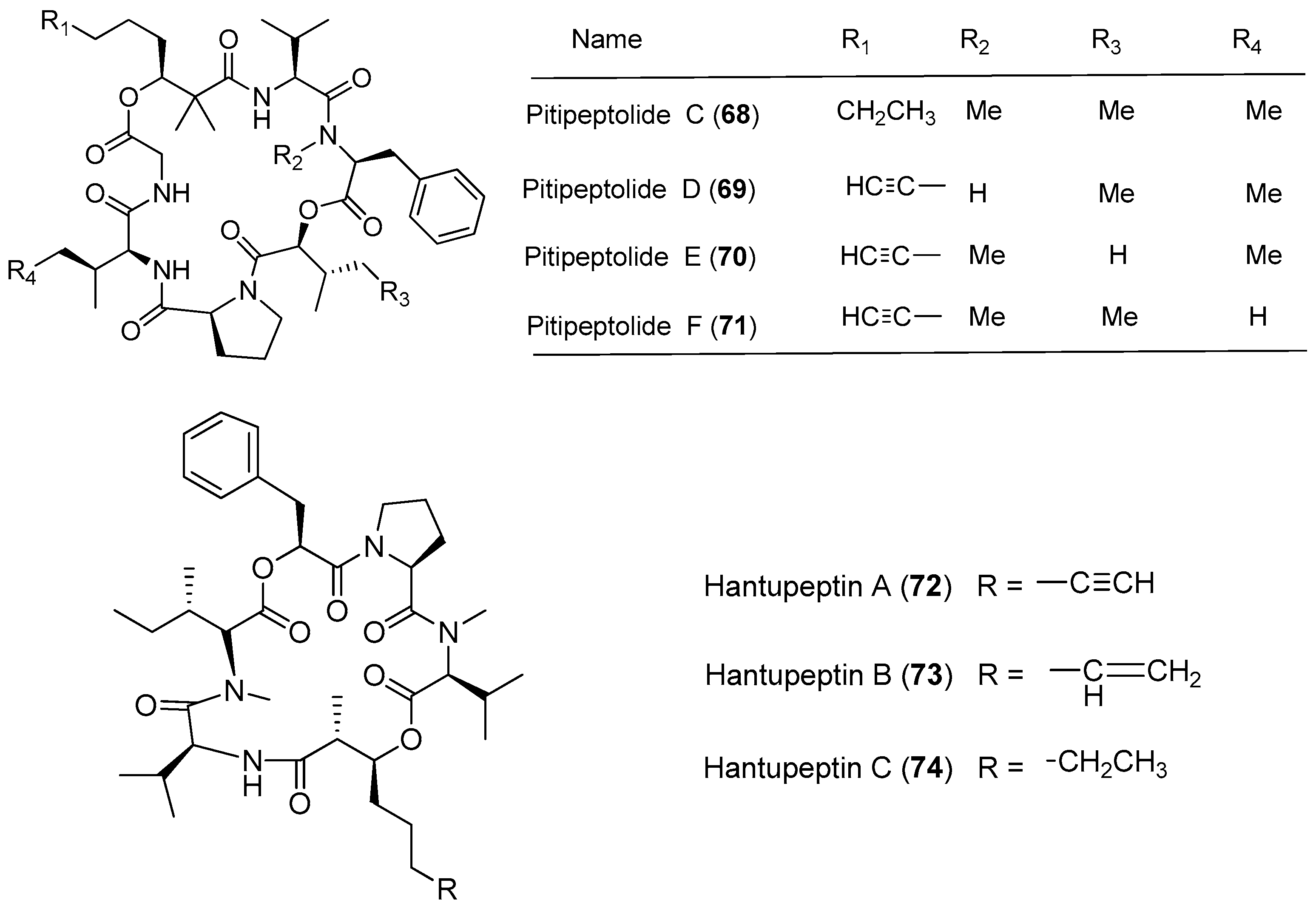
Lyngbya majuscula has been proved to be a chemically prolific species of cyanobacterium since a large number of natural products with structural diversity have been purified from Lyngbya majuscula (Figure 11 and Figure 12). Pitipeptolides C–F (68–71) are antimycobacterial cyclodepsipeptides isolated from the marine cyanobacterium Lyngbya majuscula from Piti Bomb Holes, Guam (Figure 11). They (68–71) show weak cytotoxicity against HT-29 colon adenocarcinoma and MCF7 breast cancer cells [53]. Three cytotoxic cyclic depsipeptides, hantupeptins A–C (72–74), have been derived from the marine cyanobacterium Lyngbya majuscula from Pulau Hantu Besar, Singapore [54,55]. Compound 72 shows strong cytotoxicity against leukemia cells and breast cancer MCF-7 cell (IC50 values of 32 and 4.0 μM, respectively) [54], while compounds 73 and 74 display moderate cytotoxicity against MOLT-4 (leukemia) and MCF-7 cell lines [55].
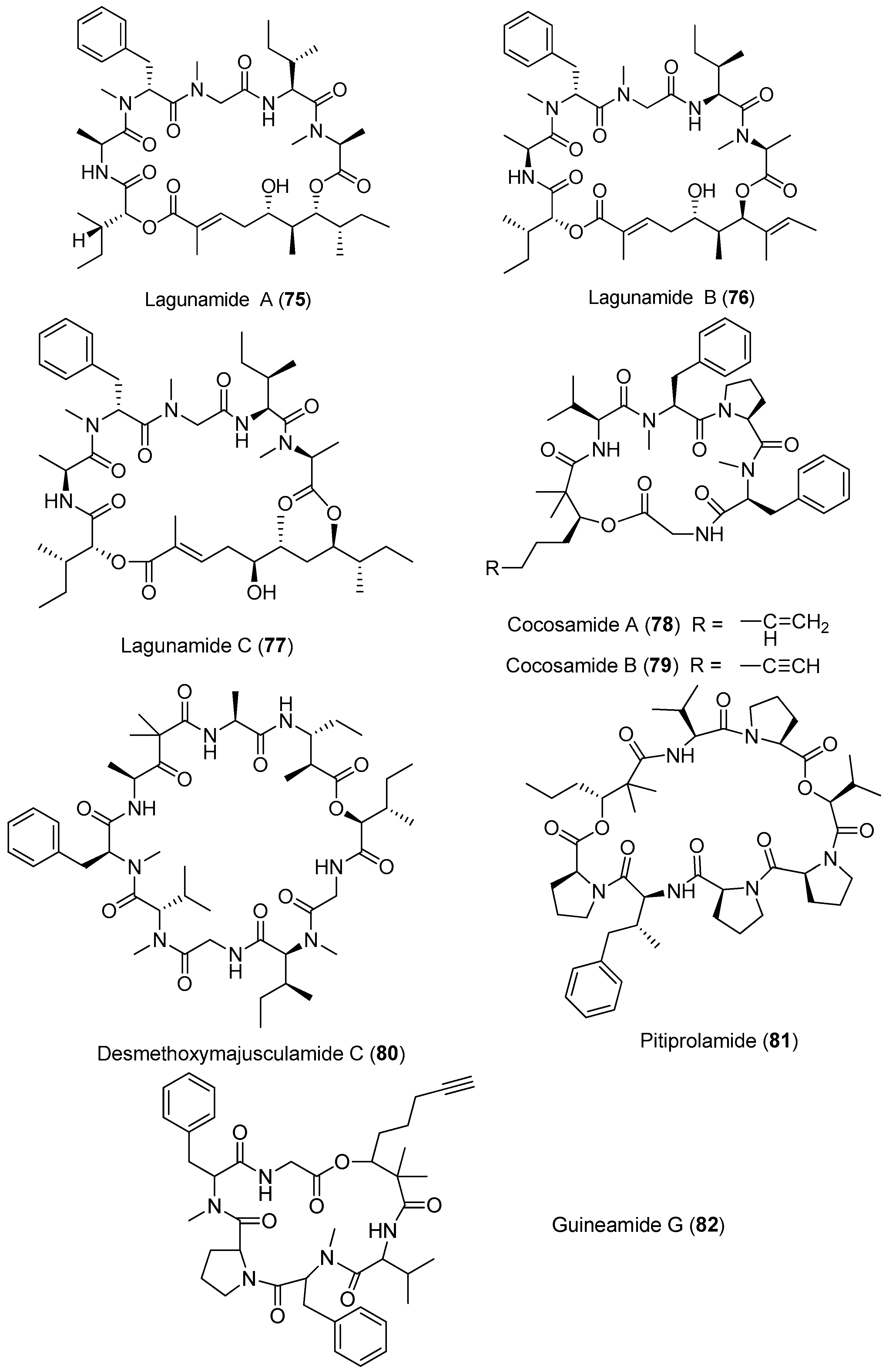
Three new cytotoxic cyclic depsipeptides, lagunamides A–C (75–77), have been extracted from Lyngbya majuscula collected from Pulau Hantu Besar, Singapore (Figure 12). Compounds 75–77 show antimalarial activity against Plasmodium falciparum with IC50 values of 0.19, 0.91 and 0.29 μM, respectively. Compounds 75 and 76 display significant cytotoxicity against P388 leukemia cells with IC50 values of 6.4 and 20.5 nM, respectively. Compound 77 exhibits potent cytotoxicity against several cancer cell lines, such as P388, A549, PC3, HCT8 and SK-OV3 cell lines, with IC50 values ranging from 2.1 to 24.4 nM. Furthermore, these compounds 75–77 display anti-swarming activities against Pseudomonas aeruginosa PA01 [56,57]. Total synthesis of 75 has been achieved, leading to revision of the structure of compound 75 [58].
Two cyclic depsipeptides, cocosamides A and B (78, 79), have been purified from marine cyanobacterium Lyngbya majuscula collected from the Cocos Lagoon, Guam, and they display weak cytotoxicity against MCF-7 breast cancer cells and HT-29 colon cancer cells [59]. A potent cytotoxic cyclic depsipeptide, desmethoxymajusculamide C (80) extracted from Lyngbya majuscula from the Fijian, displays potent and selective cytotoxicity against the HCT-116 human colon carcinoma cell line with an IC50 value of 20 nM [60]. A proline-rich analogue of dolastatin 16, pitiprolamide (81) isolated from Lyngbya majuscula collected from Guam, exhibits weak cytotoxicity against HCT116 colorectal cancer and MCF-7 breast cancer cell lines, and weak antibacterial activity against Mycobacterium tuberculosis and Bacillus cereus [61]. Guineamide G (82) has been extracted from Lyngbya majuscula. Compound 82 exhibits brine shrimp toxicity and shows potent cytotoxicity against mouse neuroblastoma cell line with an LC50 value of 2.7 μM [62].
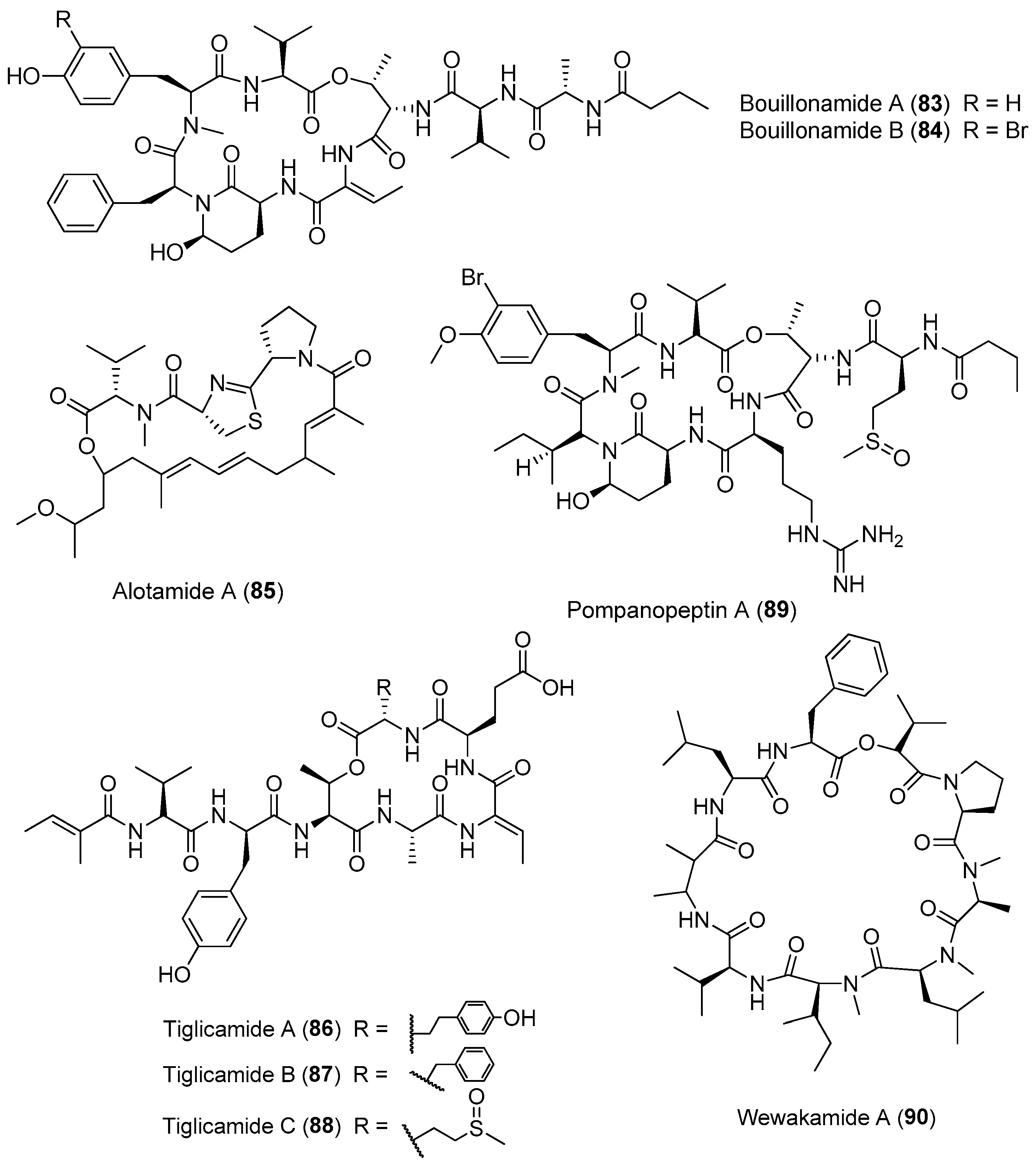
Except for Lyngbya majuscula, other cyanobacteria of the genus Lyngbya are important producers of bioactive cyclic depsipeptides (Figure 13). Two proteases inhibitors, bouillomides A and B (83, 84), have been isolated from the marine cyanobacterium Lyngbya bouillonii from Guam. Compounds 83 and 84 selectively inhibit serine protease elastase (IC50 = 1.9 μM for both) and chymotrypsin (IC50 = 0.17 and 9.3 μM, respectively), whereas no trypsin inhibition has been found (IC50 > 100 μM) [63]. A structurally intriguing cyclic depsipeptide, alotamide A (85) containing rare unsaturated heptaketide residues, has been extracted from Lyngbya bouillonii collected from Papua New Guinea. Compound 85 shows an unusual calcium influx in murine cerebrocortical neurons with an EC50 value of 4.18 μM [64]. Three protease inhibitors, named tiglicamides A–C (86–88) containing an unusual tiglic acid moiety, have been purified from the marine cyanobacterium Lyngbya confervoides from Florida. Compounds 86–88 display moderate inhibitory effects on porcine pancreatic elastase with IC50 values ranging from 2.14 to 7.28 μM [65]. Pompanopeptin A (89) has been purified from Lyngbya confervoides from the coast of Florida. Compound 89 containing an arginine residue in the cyclic core shows selective protease inhibition of trypsin (IC50 value of 2.4 μM) over elastase and chymotrypsin [66]. Wewakamide A (90) has been extracted from Lyngbya semiplena and exhibits brine shrimp toxicity [62].
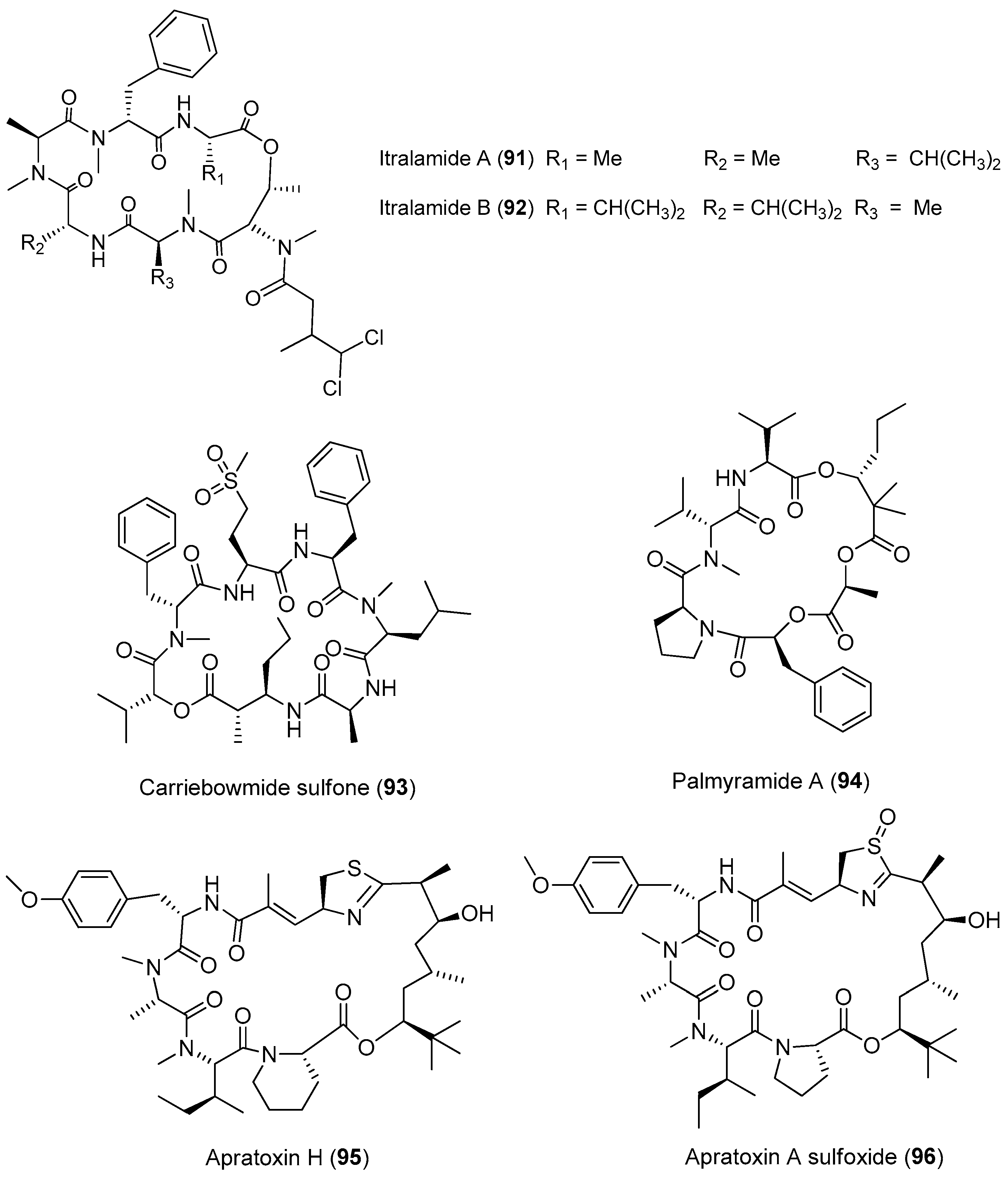
Three new cytotoxic cyclic depsipeptides, named itralamides A and B (91, 92) and carriebowmide sulfone (93), have been extracted from Moorea producens (formerly Lyngbya majuscula) from the Eastern Caribbean (Figure 14). Only compound 92 exhibits significant cytotoxicity with an IC50 value of 6 μM [24,67]. The total synthesis of the proposed structure for 92 has been achieved, but the true structure of the natural compound 92 is still unknown [68].
A cyclic depsipeptide, palmyramide A (94), has been identified through bioassay-guided isolation of the marine cyanobacterium Moorea producens (formerly Lyngbya majuscula) from Palmyra Atoll [24]. Compound 94 displays sodium channel-blocking activity in neuro-2a cells (IC50 value of 17.2 μM) and shows moderate cytotoxicity in H-460 human lung carcinoma cells (IC50 value of 39.7 μM) [69]. Cultivation of the Moorea producens (formerly Lyngbya majuscula) affords two new apratoxin analogues, termed apratoxin H (95) and apratoxin A sulfoxide (96). Compounds 95 and 96 exhibit strong cytotoxicity against human NCI-H460 lung cancer cells with IC50 values of 3.4 and 89.9 nM, respectively. The biosynthetic pathway of apratoxins supports the opinion that structural diversity of apratoxin can be the result of regional diversity [70].
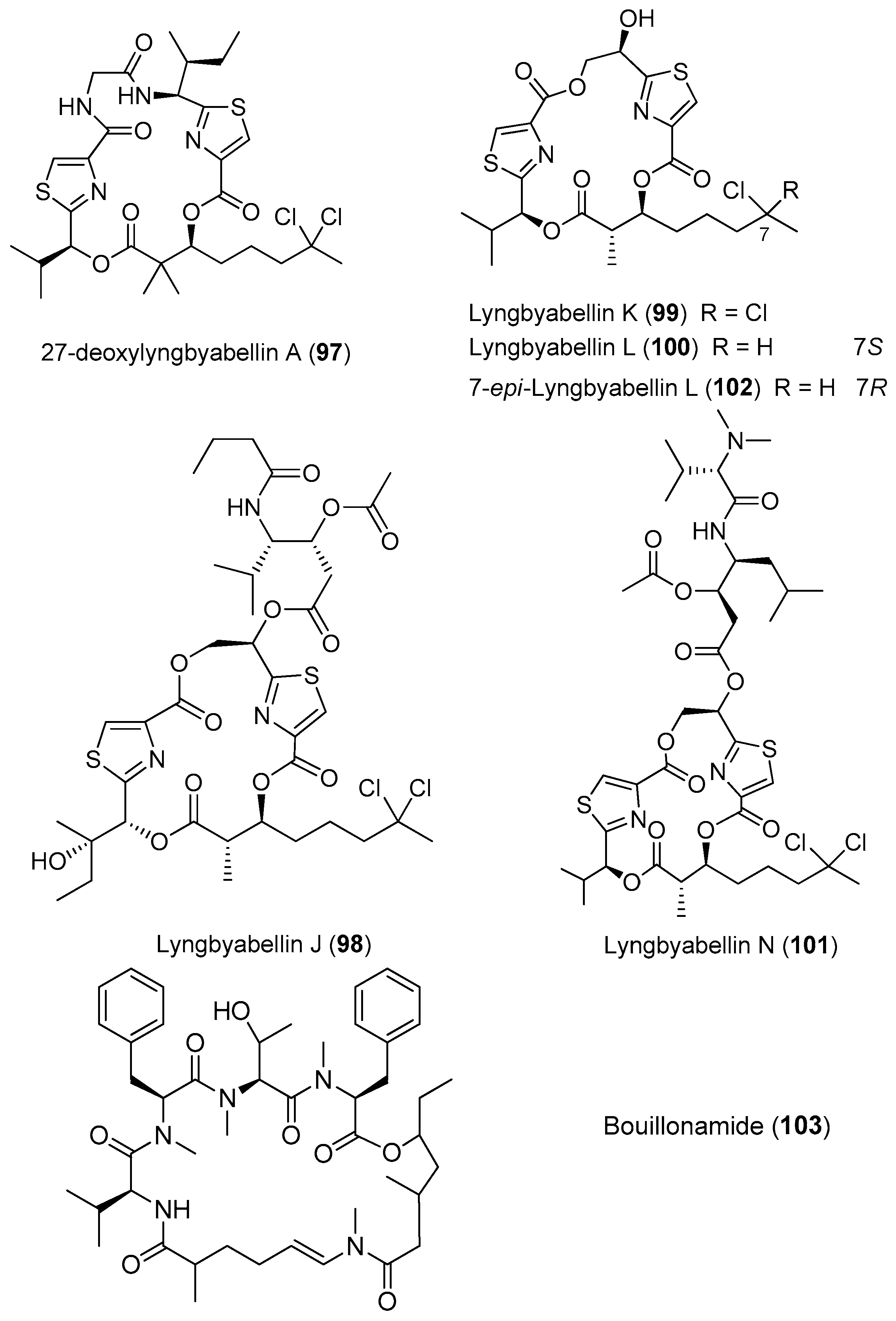
Two analogues of lyngbyabellin A, 27-deoxylyngbyabellin A (97) and lyngbyabellin J (98), have been isolated from Moorea bouillonii (formerly Lyngbya bouillonii) collected from shallow patch reefs in Apra Harbor, Guam [8]. Compounds 97 and 98 show weak cytotoxicity against HT29 colon cancer cells and HeLa cervical cancer cells [26]. Four novel cyclic petides, lyngbyabellins K and L (99, 100), lyngbyabellin N (101) and 7-epi-lyngbyabellin L (102), have been extracted from two collections of marine cyanobacteria Moorea bouillonii from Palmyra Atoll in the Central Pacific Ocean (Figure 15). Compound 101 containing an unusual N,N-dimethylvaline residue and a leucine statine residue shows potent cytotoxicity against HCT116 colon cancer cell line with an IC50 value of 40.9 nM [37]. A novel cytotoxic cyclic depsipeptide, bouillonamide (103), has been derived from the tropical marine cyanobacterium Moorea bouillonii collected from New Britain, Papua New Guinea. Compound 103 shows mild toxicity against the neuron 2a mouse neuroblastoma cells with IC50 value of 6.0 μM [71].
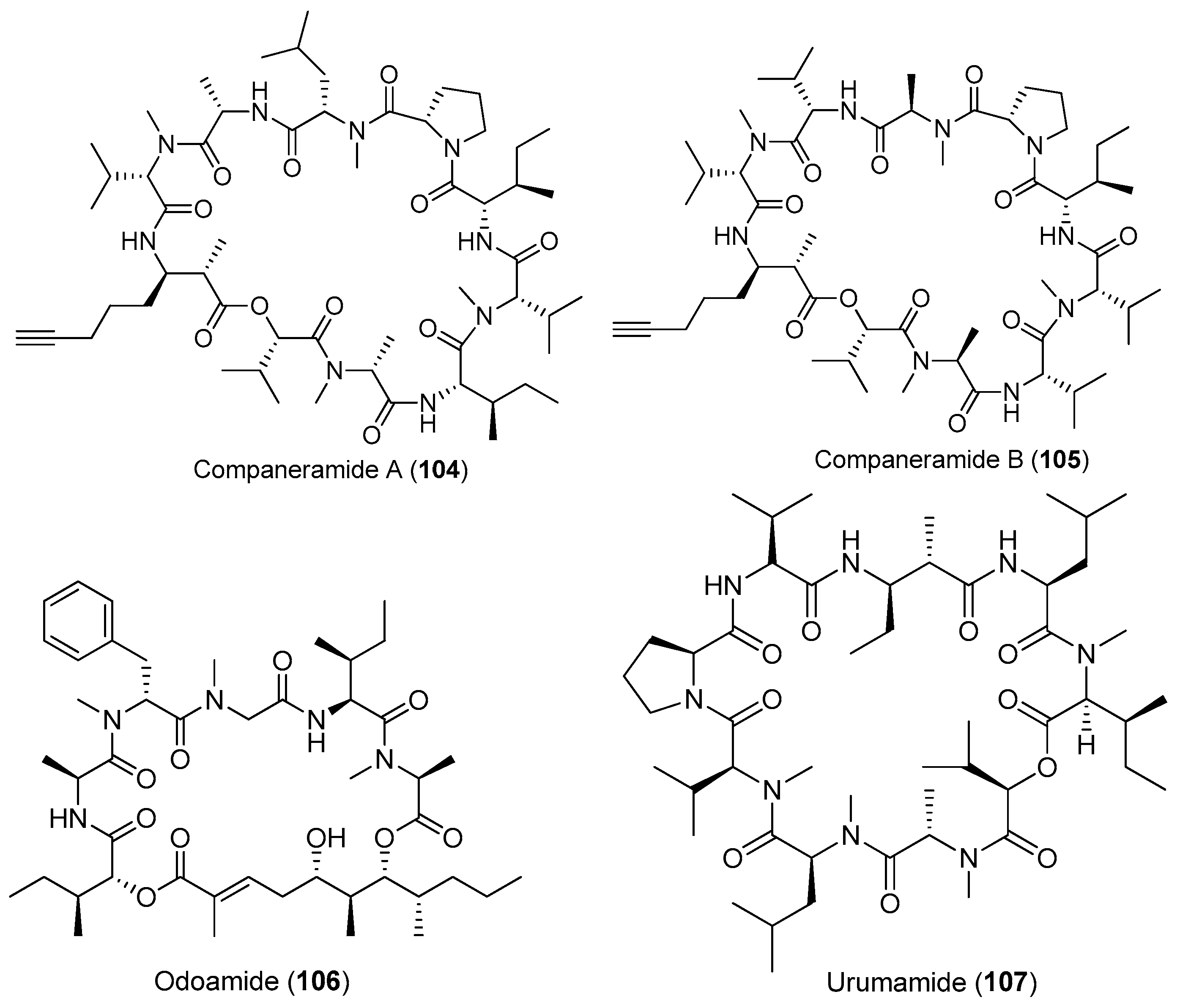
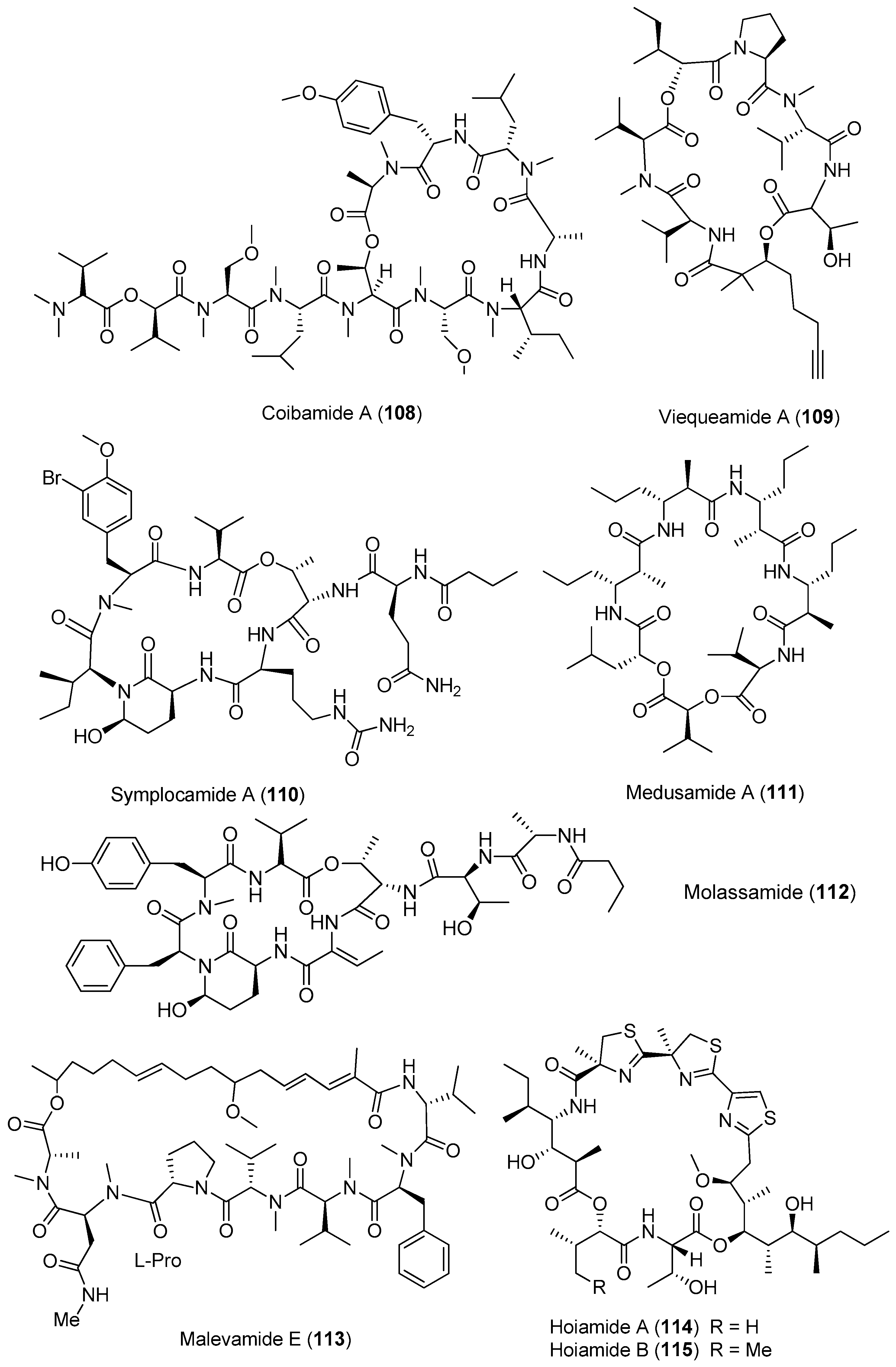
Except for cyanobacteria of the genera Lyngbya and Moorea, other species of marine cyanobacteria also have provided some bioactive cyclic depsipeptides (Figure 16). Two antimalarial cyclodepsipeptides, companeramides A and B (104, 105), have been isolated from the marine cyanobacterial assemblage collected from Coiba Island, Panama. Compounds 104 and 105 display high antiplasmodial activity [72]. Two new cyclic cyclodepsipeptides, odoamide (106) and urumamide (107), have been purified from Okeania sp. collected from Okinawa Prefecture and Ikei Island, Okinawa, respectively [73,74]. Compound 106 shows strong cytotoxicity against HeLa S3 human cervical cancer cells with an IC50 of 26.3 nM [73]. Compound 107 inhibits the growth of HeLa and HL60 cells with IC50 values of 18 and 13 μM, respectively. Compound 107 inhibits chymotrypsin with an IC50 value of 33 μM [74]. An unprecedented cytotoxic depsipeptide, coibamide A (108), has been identified from Caldora penicillata (formerly Leptolyngbya sp.) from Coiba Island National Park, Panama (Figure 16). Compound 108 contains eight N-methylated amino acid residues and shows significant and selective cytotoxicity against NCI-H460 lung cancer cells and mouse neuro-2a cells [10,75]. The structure of compound 108 has been revised by successful total synthesis [76]. A new marine cyclicdepsipeptide with potent cytotoxicity, viequeamide A (109), has been derived from the marine cyanobacterium Rivularia sp. from the Vieques Island, Puerto Rico. Compound 109 displays potent cytotoxicity against H460 human lung cancer cells with an IC50 value of 60 nM [77]. Total synthesis of 109 has been completed [78].
A potent cytotoxin, symplocamide A (110), has been identified from Symploca sp. collected from Papua New Guinea. Compound 110 shows potent cytotoxicity against H-460 lung cancer cells and neuro-2a neuroblastoma cells with IC50 values of 40 and 29 nM, respectively. Compound 110 selectively inhibits chymotrypsin with a greater potency (200-fold) than trypsin [79]. The first cyclic depsipeptide reported to contain multiple Amha residues and four contiguous β-amino acid residues, named medusamide A (111), has been isolated from a collection of marine cyanobacteria from Coiba Island on the Pacific coast of Panama [80]. The first peptide from the genus Dichothrix, termed molassamide (112), has been purified from the marine cyanobacterium Dichothrix utahensis collected from the Molasses Reef, Key Largo, Florida and from Brewer’s Bay, St. Thomas, U.S. Virgin Islands. Compound 112 exhibits serine protease inhibition against elastase and chymotrypsin with IC50 values of 0.032 and 0.234 μM, respectively. No inhibitory activity against trypsin has been found at the highest concentration tested (10 μM) [81]. Malevamide E (113) isolated from the marine cyanobacterium Symploca laeteviridis shows store-operated Ca2+ entry in thapsigargin-treated human embryonic kidney (HEK) cells with a dose-dependent inhibition manner (2–45 μM) [82].
Hoiamide A (114), isolated from Papua New Guinea marine cyanobacteria Lyngbya majuscula and Phormidium gracile, is an unusual cyclic depsipeptide that consists of an acetate extended and S-adenosyl methionine modified isoleucine moiety, a triheterocyclic fragment bearing two α-methylated thiazolines and one thiazole, and a highly oxygenated and methylated C15-polyketide substructure (Figure 16). Compound 114 is potent inhibitor of voltage-gated sodium channels (IC50 = 92.8 nM), and it can activate sodium influx (EC50 = 2.31 μM) in mouse neocortical neurons [83]. An analogue of compound 114, hoiamide B (115), has been derived from two different collections of marine cyanobacteria from Papua New Guinea. Compound 115 stimulates sodium influx and suppresses spontaneous Ca2+ oscillations in neocortical neurons with EC50 values of 3.9 μM and 79.8 nM, respectively [34].
3.2. Other Cyclic Peptides
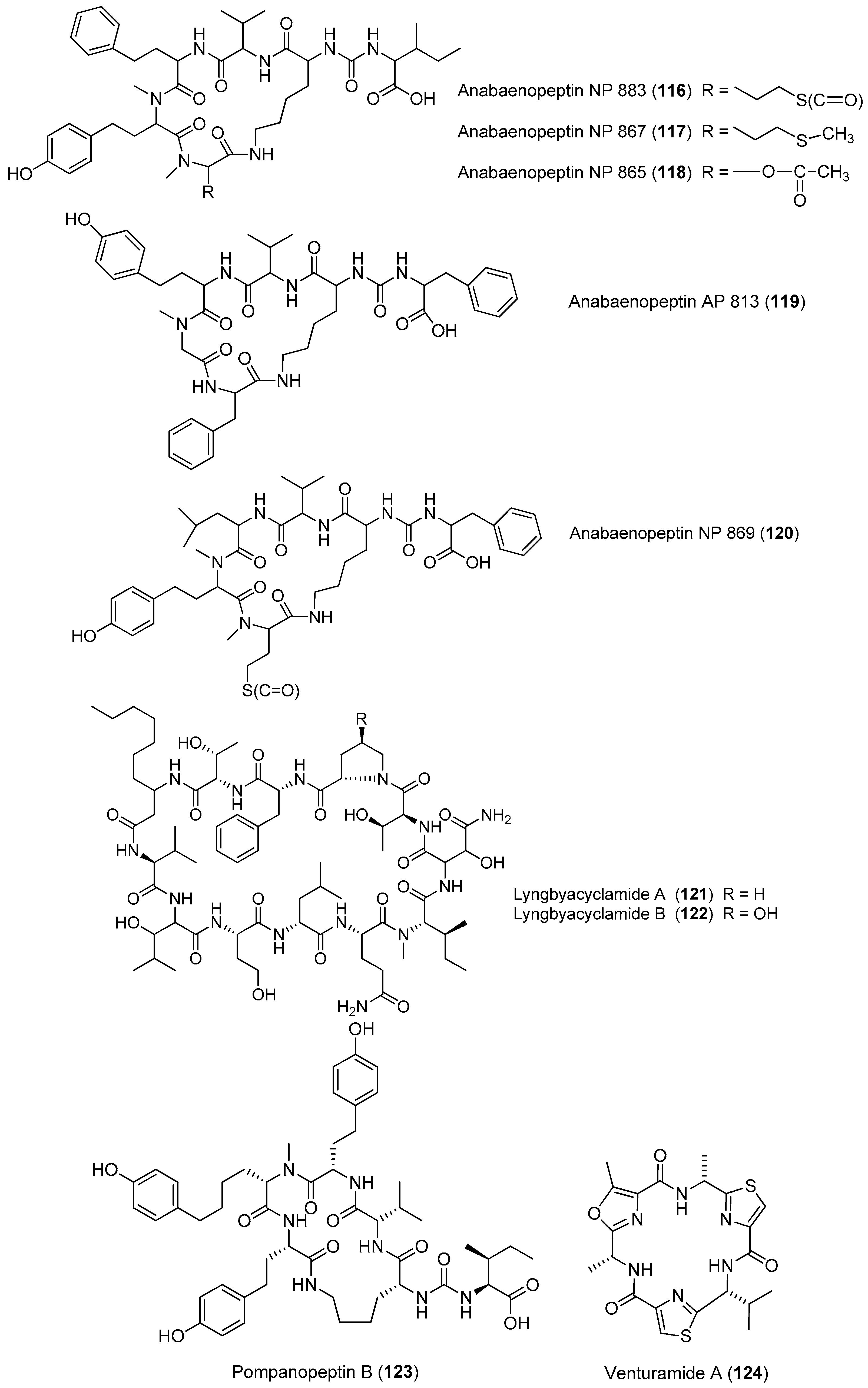
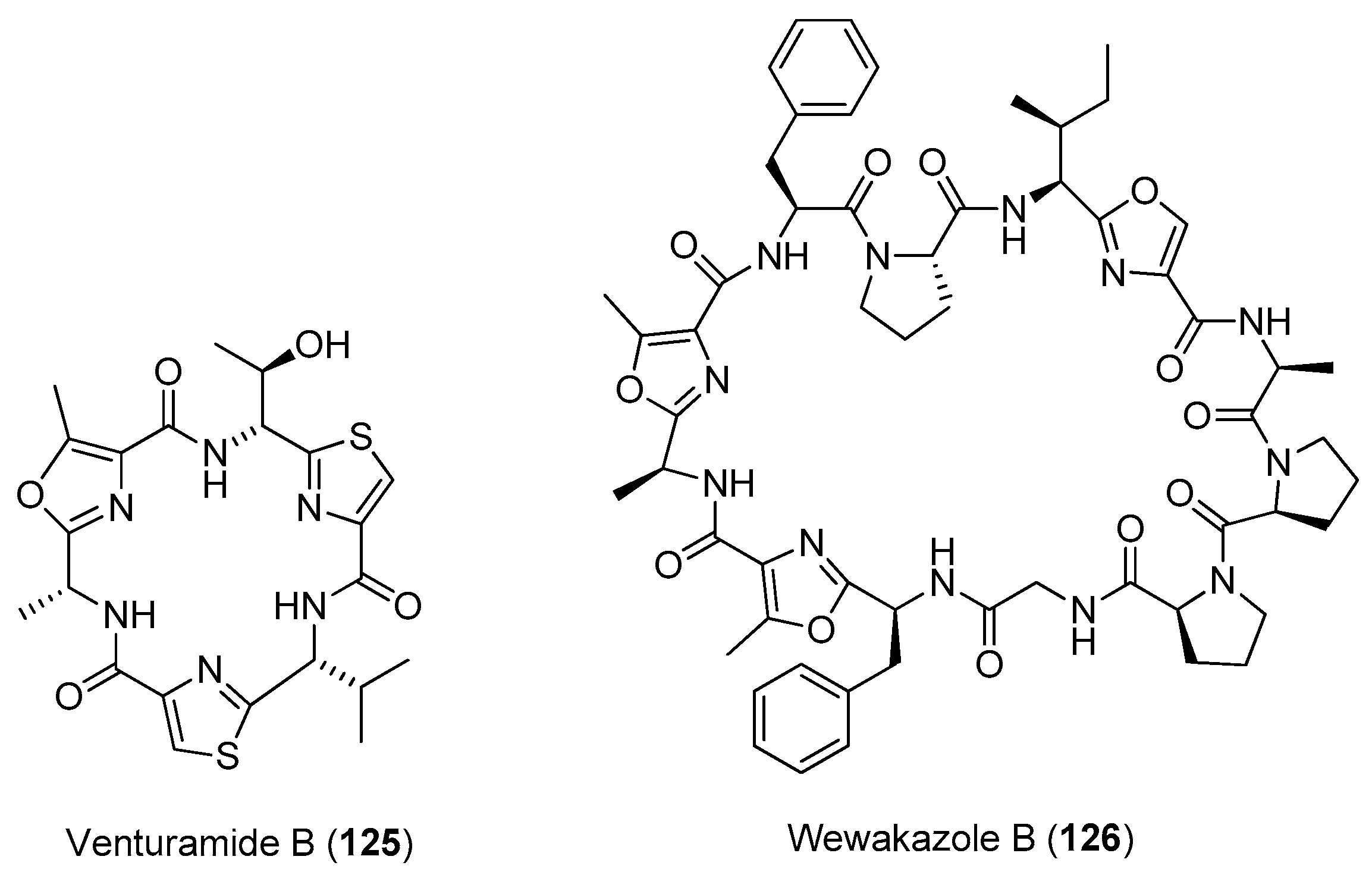
Five anabaenopeptin-related compounds (called by their molecular weight), termed anabaenopeptin NP883 (116), anabaenopeptin NP867 (117), anabaenopeptin NP865 (118), anabaenopeptin AP813 (119) and anabaenopeptin NP869 (120), have been isolated from a bloom sample of marine cyanobacteria of Baltic Sea (Figure 17). Compounds 116–120 inhibit carboxypeptidase A and protein phosphatase 1 with varying potency [84]. Two new cyclic peptides, lyngbyacyclamides A and B (121, 122), have been purified from the marine cyanobacterium Lyngbya sp. collected from Okinawa Prefecture, Japan. Compounds 121 and 122 show cytotoxicity against the growth of B16 melanoma cells with an IC50 of 0.7 μM [85]. Total synthesis of 121 has been completed [86]. Pompanopeptin B (123) has been purified from Lyngbya confervoides from the coast of Florida [66]. Two antimalarial cyclic hexapeptides, venturamides A and B (124, 125), have been identified from the Panamanian marine cyanobacterium Oscillatoria sp. through antimalarial bioassay-guided isolation [87]. A new cyanobacterial toxin, wewakazole B (126), has been isolated from Moorea producens collected in the Red Sea by mass spectrometry-guided isolation. Compound 126 shows cytotoxicity against human MCF7 breast cancer cells and human H460 lung cancer cells with IC50 values of 0.58 and 1.0 μM, respectively [88]. Total synthesis of 126 has been achieved [89].
4. Conclusions
Marine cyanobacteria are the significant sources of structurally diverse marine natural products with broad biological activities. Significant progress has been made in discovery of bioactive secondary metabolites from marine cyanobacteria over the past decade. The overwhelming majority of cyanobacterial secondary metabolites are peptides, especially cyclic depsipeptides (76 compounds), accounting for more than half of the total cyanobacterial peptides (126 compounds).
However, there are lots of problems in drug development from marine cyanobacteria, including evaluation of the taxonomy of cyanobacteria, new techniques developed to culture marine cyanobacteria in mass, total synthesis and multi-target screening assay. Firstly, the cyanobacterial genus Lyngbya, especially Lyngbya majuscula, has been proved to be important producers of marine peptides. Three new cyanobacterial genera Moorea, Okeania and Caldora, have been proposed in the past few years. The three new genera were previously identified as the chemically rich genera Lyngbya and Symploca, respectively [8,9,10]. Moreover, several important peptides with promising pharmaceutical potential, such as symplostatin 1 and dolastatin 10, were actually isolated from these new genera of cyanobacteria. At present, the chemically rich genus Lyngbya has been shown to be polyphyletic, and biodiversity in tropical marine cyanobacteria remains currently unclear [10]. It is necessary to evaluate the taxonomy of NP-rich marine cyanobacteria using a combined molecular, morphological and chemical approach in further research.
Secondly, cyanobacteria have great potential as sustainable sources for production of bioactive peptides because of their rapid growth, genetic tractability and cultivable property [3]. Although cyanobacteria possess the cultivable properties similar to those of microorganisms, cyanobacteria have attracted far less attention than microorganisms. More efforts should be invested in developing new techniques to culture marine cyanobacteria in mass. Thirdly, total synthesis of some bioactive cyanobacterial peptides has been successfully achieved, which might be beneficial for the structure revision of natural peptides, further evaluation and pharmacological applications. In addition, at present, the majority of cyanobacterial peptides exhibit a broad range of bioactivities, including cytotoxic, antibacterial, antimalarial, enzyme inhibition, parasitic resistance and channel-blocking activities. The overwhelming majority of cyanobacterial peptides display in vitro antitumor activity. Multi-target screening assays should be developed to accelerate the discovery of promising drug leader compounds.
At present, two scholars, including Luesch H. and Gerwick W., have greatly contributed to the discovery of new peptides from marine cyanobacteria. Programs for drug discovery from marine cyanobacteria, such as the Panama ICBG program, have led to the discovery of bioactive cyanobacterial peptides. Marine cyanobacteria have great potential as sustainable marine sources for production of bioactive peptides (such as dolastatins) because of their genetic tractability, cultivable property, rapid growth and peptide biosynthetic pathway. This review summarized new peptides derived from marine cyanobacteria over the past decade, providing useful information in the further discovery of novel cyanobacterial peptides.
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (41406163), the 863 Program of China (2013AA092902), the Ningbo Marine Algae Biotechnology Team (2011B81007), the Li Dak Sum Yip Yio Chin Kenneth Li Marine Biopharmaceutical Development Fund, the National 111 Project of China, the Scientific Research Foundation for Returned Scholars of ZJHRSS and the K.C.Wong Magna Fund in Ningbo University.
Author Contributions
All authors (Y.M., J.Z., S.H., X.Y.) contributed equally to the work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Silipo, A.; Molinaro, A.; Molteni, M.; Rossetti, C.; Parrilli, M.; Lanzetta, R. Full Structural Characterization of an Extracellular Polysaccharide Produced by the Freshwater Cyanobacterium Oscillatoria planktothrix FP1. Eur. J. Org. Chem. 2010, 29, 5594–5600. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Mahmood, N.A.; Hyde, E.G. Natural toxins from cyanobacteria (Blue-Green Algae). In Marine Toxins: Origin, Structure and Molecular Pharmacology, 1st ed.; Hall, S., Strichartz, G., Eds.; ACS: Washington, DC, USA, 1990; Volume 52, pp. 87–106. [Google Scholar]
- Martins, J.; Vasconcelos, V. Cyanobactins from cyanobacteria: Current genetic and chemical state of knowledge. Mar. Drugs 2015, 13, 6910–6946. [Google Scholar] [CrossRef] [PubMed]
- Natsume, T.; Watanabe, J.; Horiuchi, T.; Kobayashi, M. Combination effect of TZT-1027 (soblidotin) with other anticancer drugs. Anticancer Res. 2006, 26, 1145–1152. [Google Scholar] [PubMed]
- Deng, C.C.; Pan, B.Q.; O’Connor, O.A. Brentuximab Vedotin. Clin. Cancer Res. 2013, 19, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Reyesac, L.A.; Luesch, H. Biological targets and mechanisms of action of natural products from marine cyanobacteria. Nat. Prod. Rep. 2015, 32, 478–503. [Google Scholar] [CrossRef] [PubMed]
- Engene, N.; Rottacker, E.C.; Kaštovský, J.; Byrum, T.; Choi, H.; Ellisman, M.H.; Komárek, J.; Gerwick, W.H. Moorea producens gen. nov., sp. nov. and Moorea bouillonii comb. nov., tropical marine cyanobacteria rich in bioactive secondary metabolites. Int. J. Syst. Evol. Microbiol. 2012, 62, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Engene, N.; Paul, V.J.; Byrum, T.; Gerwick, W.H.; Thor, A.; Ellisman, M.H. Five chemically rich species of tropical marine cyanobacteria of the genus Okeania gen. nov. (Oscillatoriales, Cyanoprokaryota). J. Phycol. 2013, 49, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Engene, N.; Tronholm, A.; Salvador-Reyes, L.A.; Luesch, H.; Paul, V.J. Caldora penicillata gen. nov., comb. nov. (cyanobacteria), a pantropical marine species with biomedical relevance. J. Phycol. 2015, 51, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Rein, K.S. New Peptides Isolated from Lyngbya Species: A Review. Mar. Drugs 2010, 8, 1817–1837. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.C.; Eksioglu, E.A.; Liu, C.; Paul, V.J.; Luesch, H. Grassystatins A–C from marine cyanobacteria, potent cathepsin E inhibitors that reduce antigen presentation. J. Med. Chem. 2009, 52, 5732–5747. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.M.; Zhang, W.; Ding, N.; Lo, J.; Liu, Y.X.; Clare-Salzler, M.J.; Luesch, H.; Li, Y.X. Total synthesis of grassystatin A, a probe for cathepsin E function. Bioorg. Med. Chem. 2012, 20, 4774–4780. [Google Scholar] [CrossRef] [PubMed]
- Mevers, E.; Liu, W.T.; Engene, N.; Mohimani, H.; Byrum, T.; Pevzner, P.A.; Dorrestein, P.C.; Spadafora, C.; Gerwick, W.H. Cytotoxic veraguamides, alkynyl bromide-containing cyclic depsipeptides from the marine cyanobacterium cf. Oscillatoria margaritifera. J. Nat. Prod. 2011, 74, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Ohno, O.; Sumimoto, S.; Suda, S.; Suenaga, K. Maedamide, a novel chymotrypsin inhibitor from a marine cyanobacterial assemblage of Lyngbya sp. Tetrahedron Lett. 2014, 55, 4126–4128. [Google Scholar] [CrossRef]
- Takayanag, A.; Iwasaki, A.; Suenaga, K. Total synthesis and stereochemical reassignment of maedamide. Tetrahedron Lett. 2015, 56, 4947–4949. [Google Scholar] [CrossRef]
- Simmons, T.L.; Engene, N.; Ureña, L.D.; Romero, L.I.; Ortega-Barría, E.; Gerwick, L.; Gerwick, W.H. Viridamides A and B, lipodepsipeptides with anti-protozoa activity from the marine cyanobacterium Oscillatoria nigro-viridis. J. Nat. Prod. 2008, 71, 1544–1550. [Google Scholar] [CrossRef] [PubMed]
- Linington, R.G.; Clark, B.R.; Trimble, E.E.; Almanza, A.; Ureña, L.; Kyle, D.E.; Gerwick, W.H. Antimalarial peptides from marine cyanobacteria: Isolation and structural elucidation of gallinamide A. J. Nat. Prod. 2009, 72, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Conroy, T.; Guo, J.T.; Linington, G.R.; Hunt, N.H.; Payne, R.J. Total synthesis, stereochemical assignment, and antimalarial activity of gallinamide A. Chem. Eur. J. 2011, 17, 13544–13552. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, L.M.; Lopez, D.; Vesely, B.A.; Togna, G.D.; Gerwick, W.H.; Kyle, D.E.; Linington, R.G. Almiramides A–C: Discovery and development of a new class of Leishmaniasis lead compounds. J. Med. Chem. 2010, 53, 4187–4197. [Google Scholar] [CrossRef] [PubMed]
- Quintana, J.; Bayona, L.M.; Castellanos, L.; Puyana, M.; Camargo, P.; Aristizábal, F.; Edwards, C.; Tabudravu, J.N.; Jaspars, M.; Ramos, F.A. Almiramide D, cytotoxic peptide from the marine cyanobacterium Oscillatoria nigroviridis. Bioorg. Med. Chem. 2014, 22, 6789–6795. [Google Scholar] [CrossRef] [PubMed]
- McPhail, K.L.; Correa, J.; Linington, R.G.; González, J.; Ortega-Barría, E.; Capson, T.L.; Gerwick, W.H. Antimalarial linear lipopeptides from a panamanian strain of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2007, 70, 984–988. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Ross, C.; Paul, V.J.; Matthew, S.; Luesch, H. Dragonamides C and D, linear lipopeptides from the marine cyanobacterium brown Lyngbya polychroa. J. Nat. Prod. 2008, 71, 887–890. [Google Scholar] [CrossRef] [PubMed]
- Engene, N.; Gunasekera, S.P.; Gerwick, W.H.; Paula, V.J. Phylogenetic inferences reveal a large extent of novel biodiversity in chemically rich tropical marine cyanobacteria. Appl. Environ. Microbiol. 2013, 79, 1882–1888. [Google Scholar] [CrossRef] [PubMed]
- Balunas, M.J.; Linington, R.G.; Tidgewell, K.; Fenner, A.M.; Ureña, L.; Togna, G.D.; Kyle, D.E.; Gerwick, W.H. Dragonamide E, a modified linear lipopeptide from Lyngbya majuscula with antileishmanial activity. J. Nat. Prod. 2010, 73, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Matthew, S.; Salvador, L.A.; Schupp, P.J.; Paul, V.J.; Luesch, H. Cytotoxic halogenated macrolides and modified peptides from the apratoxin-producing marine cyanobacterium Lyngbya bouillonii from Guam. J. Nat. Prod. 2010, 73, 1544–1552. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Ohno, O.; Sumimoto, S.; Ogawa, H.; Nguyen, K.A.; Suenaga, K. Jahanyne, an apoptosis-inducing lipopeptide from the marine cyanobacterium Lyngbya sp. Org. Lett. 2015, 17, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Teruya, T.; Sasaki, H.; Fukazawa, H.; Suenaga, K. Bisebromoamide, a potent cytotoxic peptide from the marine cyanobacterium Lyngbya sp.: Isolation, stereostructure, and biological activity. Org. Lett. 2009, 11, 5062–5065. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Teruya, T.; Fukazawa, H.; Suenaga, K. Revised structure and structureeactivity relationship of bisebromoamide and structure of norbisebromoamide from the marine cyanobacterium Lyngbya sp. Tetrahedron 2011, 67, 990–994. [Google Scholar] [CrossRef]
- Gao, X.G.; Liu, Y.Q.; Kwong, S.Q.; Xu, Z.S.; Ye, T. Total synthesis and stereochemical reassignment of bisebromoamide. Org. Lett. 2010, 12, 3018–3021. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Yu, S.Y.; Jin, M.Z.; Xia, H.G.; Ma, D. Total synthesis and cytotoxicity of bisebromoamide and its analogues. Tetrahedron Lett. 2011, 52, 2124–2127. [Google Scholar] [CrossRef]
- Mevers, E.; Haeckl, J.; Boudreau, P.D.; Byrum, T.; Dorrestein, P.C.; Valeriote, F.A.; Gerwick, W.H. Lipopeptides from the tropical marine cyanobacterium Symploca sp. J. Nat. Prod. 2014, 77, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Pereiraa, A.R.; Kalea, A.J.; Fenleyb, A.T.; Byruma, T.; Debonsia, H.M.; Gilsonb, M.K.; Valerioted, F.A.; Moorea, B.S.; Gerwicka, W.H. The Carmaphycins, new proteasome inhibitors exhibiting an α,β-epoxyketone warhead from a marine cyanobacterium. Chembiochem 2012, 4, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Pereira, A.R.; Cao, Z.Y.; Shuman, C.F.; Engene, N.; Byrum, T.; Matainaho, T.; Murray, T.F.; Mangoni, A.; Gerwick, W.H. The hoiamides, structurally intriguing neurotoxic lipopeptides from Papua New Guinea marine cyanobacteria. J. Nat. Prod. 2010, 73, 1411–1421. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, Z.S.; Ye, T. Total synthesis of hoiamide C. Org. Lett. 2011, 13, 2506–2509. [Google Scholar] [CrossRef] [PubMed]
- Malloy, K.L.; Choi, H.; Fiorilla, C.; Valeriote, F.A.; Matainahoe, T.; Gerwick, W.H. Hoiamide D, a marine cyanobacteria-derived inhibitor of p53/MDM2 interaction. Bioorg. Med. Chem. Lett. 2012, 22, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Mevers, E.; Byrumb, T.; Valerioted, F.A.; Gerwick, W.H. Lyngbyabellins K-N from two palmyra atoll collections of the marine cyanobacterium Moorea bouillonii. Eur. J. Org. Chem. 2012, 27, 5141–5150. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Ohno, O.; Sumimoto, S.; Suda, S.; Suenaga, K. Kurahyne, an acetylene-containing lipopeptide from a marine cyanobacterial assemblage of Lyngbya sp. RSC Adv. 2014, 4, 12840–12843. [Google Scholar] [CrossRef]
- Okamoto, S.; Iwasaki, A.; Ohno, O.; Suenaga, K. Isolation and structure of kurahyne B and total synthesis of the kurahynes. J. Nat. Prod. 2015, 78, 2719–2725. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Imperial, L.; Garst, C.; Ratnayake, R.; Dang, L.H.; Paul, V.J.; Luesch, H. Caldoramide, a modified pentapeptide from the marine cyanobacterium Caldora penicillata. J. Nat. Prod. 2016, 7, 1867–1871. [Google Scholar] [CrossRef] [PubMed]
- Salvador, L.A.; Biggs, J.S.; Paul, V.J.; Luesch, H. Veraguamides A-G, cyclic hexadepsipeptides from a dolastatin16-producing cyanobacterium Symploca cf. hydnoides from Guam. J. Nat. Prod. 2011, 74, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Y.; Jia, X.; Zhang, A. Total synthesis of the proposed structure of cyclic hexadepsipeptide veraguamide A. Org. Biomol. Chem. 2012, 10, 7027–7030. [Google Scholar] [CrossRef] [PubMed]
- Matthew, S.; Ross, C.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatin 4, a dolastatin 13 analog with elastase and chymotrypsin inhibitory activity from the marine cyanobacterium Lyngbya confervoides. J. Nat. Prod. 2007, 70, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Taori, K.; Matthew, S.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatins 5–7, potent elastase inhibitors from Floridian marine cyanobacteria, Lyngbya spp. J. Nat. Prod. 2007, 70, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.M.; Chen, Q.Y.; Luesch, H. Total synthesis of the potent marine-derived elastase inhibitor lyngbyastatin 7 and in vitro biological evaluation in model systems for pulmonary diseases. J. Org. Chem. 2016, 2, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.C.; Taori, K.; Paul, V.J.; Luesch, H. Lyngbyastatins 8–10, elastase inhibitors with cyclic depsipeptide scaffolds isolated from the marine cyanobacterium Lyngbya semiplena. Mar. Drugs 2009, 7, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Thornburg, C.C.; Thimmaiah, M.; Shaala, L.A.; Hau, A.M.; Malmo, J.M.; Ishmael, J.E.; Youssef, D.T.A.; McPhail, K.L. Cyclic depsipeptides, grassypeptolides D and E and ibu-epidemethoxylyngbyastatin 3, from a Red Sea Leptolyngbya cyanobacterium. J. Nat. Prod. 2011, 74, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Taori, K.; Paul, V.J.; Luesch, H. Kempopeptins A and B, serine protease inhibitors with different selectivity profiles from a marine cyanobacterium, Lyngbya sp. J. Nat. Prod. 2008, 71, 1625–1629. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.C.; Rocca, J.R.; Abboud, K.A.; Paul, V.J.; Luesch, H. Total Structure Determi nation of Grassypeptolide, a New Marine Cyanobacterial Cytotoxin. Org. Lett. 2008, 10, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.C.; Ratnayake, R.; Abboud, K.A.; Paul, V.J.; Luesch, H. Grassypeptolides A–C, cytotoxic bis-thiazoline containing marine cyclodepsipeptides. J. Org. Chem. 2010, 75, 8012–8023. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, Y.Q.; Wang, Z.; Xing, X.Y.; Maguire, A.R.; Luesch, H.; Zhang, H.; Xu, Z.S.; Ye, T. Total synthesis and biological evaluation of grassypeptolide A. Chem. Eur. J. 2013, 19, 6774–6784. [Google Scholar] [CrossRef] [PubMed]
- Popplewell, W.L.; Ratnayake, R.; Wilson, J.A.; Beutler, J.A.; Colburn, N.H.; Henrich, C.J.; McMahon, J.B.; McKee, T.C. Grassypeptolides F and G, cyanobacterial peptides from Lyngbya majuscula. J. Nat. Prod. 2011, 74, 1686–1691. [Google Scholar] [CrossRef] [PubMed]
- Montaser, R.; Paul, V.J.; Luesch, H. Pitipeptolides C–F, antimycobacterial cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula from Guam. Phytochemistry 2011, 72, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Lee, P.P.F.; Tan, L.T. Hantupeptin A, a cytotoxic cyclic depsipeptide from a Singapore collection of Lyngbya majuscula. J. Nat. Prod. 2009, 72, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Lee, P.P.F.; Tan, L.T. Hantupeptins B and C, cytotoxic cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2010, 71, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Tan, L.T. Lagunamides A and B: Cytotoxic and antimalarial cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 1810–1814. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Chan, K.P.; Chen, D.Y.; Tan, L.T. Lagunamide C, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2011, 72, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Chen, B.; Lei, H.H.; Wang, Z.; Liu, Y.Q.; Xu, Z.S.; Ye, T. Total synthesis and stereochemical revision of lagunamide A. Chem. Commun. 2012, 48, 8697–8699. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Owle, C.S.; Montaser, R.; Luesch, H.; Paul, V.J. Malyngamide 3 and Cocosamides A and B from the marine cyanobacterium Lyngbya majuscula from Cocos Lagoon, Guam. J. Nat. Prod. 2011, 74, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.L.; Nogle, L.M.; Media, J.; Valeriote, F.A.; Mooberry, S.L.; Gerwick, W.H. Desmethoxymajusculamide C, a cyanobacterial depsipeptide with potent cytotoxicity in both cyclic and ring-opened forms. J. Nat. Prod. 2009, 72, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Montaser, R.; Abboud, K.A.; Paul, V.J.; Luesch, H. Pitiprolamide, a proline-rich dolastatin 16 analogue from the marine cyanobacterium Lyngbya majuscula from Guam. J. Nat. Prod. 2011, 74, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Bingnan, H.; Gross, H.; McPhail, K.L.; Goeger, D.; Maier, C.S.; Gerwick, W.H. Wewakamide A and guineamide G, cyclic depsipeptides from the marine cyanobacteria Lyngbya semiplena and Lyngbya majuscula. J. Microbiol. Biotechnol. 2011, 21, 930–936. [Google Scholar]
- Rubio, B.K.; Parrish, S.M.; Yoshida, W.; Schupp, P.J.; Schils, T.; Williams, P.G. Depsipeptides from a Guamanian marine cyanobacterium, Lyngbya bouillonii, with selective inhibition of serine proteases. Tetrahedron 2010, 51, 6718–6721. [Google Scholar] [CrossRef] [PubMed]
- Soria-Mercado, I.E.; Pereira, A.; Cao, Z.Y.; Murray, T.F.; Gerwick, W.H. Alotamide A, a novel neuropharmacological agent from the marine cyanobacterium Lyngbya bouillonii. Org. Lett. 2009, 11, 4704–4707. [Google Scholar] [CrossRef] [PubMed]
- Matthew, S.; Paul, V.J.; Luesch, H. Tiglicamides A–C, cyclodepsipeptides from the marine cyanobacterium Lyngbya confervoides. Phytochemistry 2009, 70, 2058–2063. [Google Scholar] [CrossRef] [PubMed]
- Matthew, S.; Ross, C.; Paul, V.J.; Luesch, H. Pompanopeptins A and B, new cyclic peptides from the marine cyanobacterium Lyngbya confervoides. Tetrahedron 2008, 64, 4081–4089. [Google Scholar] [CrossRef]
- Jiménez, J.I.; Vansach, T.; Yoshida, W.Y.; Sakamoto, B.; Pörzgen, P.; Horgen, F.D. Halogenated fatty acid amides and cyclic depsipeptides from an eastern caribbean collection of the cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2009, 72, 1573–1578. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Lv, C.S.; Liu, J.Y.; Tang, L.J.; Feng, J.M.; Tang, S.B.; Wang, Z.; Liu, Y.Q.; Meng, Y.; Ye, T.; et al. Total synthesis of the proposed structure for itralamide B. Synlett 2014, 25, 1014–1018. [Google Scholar] [CrossRef]
- Taniguchi, M.; Nunnery, J.K.; Engene, N.; Esquenazi, E.; Byrum, T.; Dorrestein, P.C.; Gerwick, W.H. Palmyramide A, a cyclic depsipeptide from a Palmyra Atoll collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Thornburg, C.C.; Cowley, E.S.; Sikorska, J.; Shaala, L.A.; Ishmael, J.E.; Youssef, D.T.A.; McPhail, K.L. Apratoxin H and apratoxin A sulfoxide from the Red Sea cyanobacterium Moorea producens. J. Nat. Prod. 2013, 76, 1781–1788. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T.; Okino, T.; Gerwick, W.H. Bouillonamide: A mixed polyketide–peptide cytotoxin from the marine cyanobacterium Moorea bouillonii. Mar. Drugs 2013, 11, 3015–3024. [Google Scholar] [CrossRef] [PubMed]
- Vining, O.B.; Medina, R.A.; Mitchell, E.A.; Videau, P.; Li, D.; Serrill, J.D.; Kelly, J.X.; Gerwick, W.H.; Proteau, P.J.; Ishmael, J.E.; et al. Depsipeptide companeramides from a Panamanian marine cyanobacterium associated with the coibamide producer. J. Nat. Prod. 2015, 3, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Sueyoshi, K.; Kaneda, M.; Sumimoto, S.; Oishi, S.; Fujii, N.; Suenaga, K.; Teruya, T. Odoamide, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Okeania sp. Tetrahedron 2016, 72, 5472–5478. [Google Scholar] [CrossRef]
- Kanamori, Y.; Iwasaki, A.; Sumimoto, S.; Suenaga, K. Urumamide, a novel chymotrypsin inhibitor with a b-amino acid from a marine cyanobacterium Okeania sp. Tetrahedron 2016, 57, 4213–4216. [Google Scholar] [CrossRef]
- Medina, R.A.; Goeger, D.E.; Hills, P.; Mooberry, S.L.; Huang, N.; Romero, L.I.; Ortega-Barría, E.; Gerwick, W.H.; McPhail, K.L. Coibamide A, a Potent antiproliferative cyclic depsipeptide from the Panamanian marine cyanobacterium Leptolyngbya sp. J. Am Chem. Soc. 2008, 130, 6324–6325. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.Y.; Pan, Z.Y.; Wu, C.L.; Wang, W.; Fang, L.J.; Su, W. Efficient synthesis and stereochemical revision of coibamide A. J. Am. Chem. Soc. 2015, 42, 13488–13491. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, P.D.; Byrum, T.; Liu, W.; Dorrestein, P.C.; Gerwick, W.H. Viequeamide A, a cytotoxic member of the kulolide superfamily of cyclic depsipeptides from a marine button cyanobacterium. J. Nat. Prod. 2012, 75, 1560–1570. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Y.; Song, S.S.; Tian, Y.; Xu, Y.J.; Miao, Z.H.; Zhang, A. Total synthesis of the marine cyclic depsipeptide viequeamide A. J. Nat. Prod. 2013, 76, 974–978. [Google Scholar] [CrossRef] [PubMed]
- Linington, R.G.; Edwards, D.J.; Shuman, C.F.; McPhail, K.L.; Matainaho, T.; Gerwick, W.H. Symplocamide A, a potent cytotoxin and chymotrypsin inhibitor from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2008, 71, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Fenner, A.M.; Engene, N.; Spadafora, C.; Gerwick, W.H.; Balunas, M.J. Medusamide A, a Panamanian cyanobacterial depsipeptide with multiple β-Amino acids. Org. Lett. 2016, 3, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Miller, M.W.; Kwan, J.C.; Luesch, H.; Paul, V.J. Molassamide, a depsipeptide serine protease inhibitor from the marine cyanobacterium Dichothrix utahensis. J. Nat. Prod. 2010, 73, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.; Pörzgen, P.; Pittman, E.; Yoshida, W.Y.; Westenburg, H.E.; Horgen, F.D. Isolation and structure determination of malevamide E, a dolastatin 14 analogue, from the marine cyanobacterium Symploca laete-Wiridis. J. Nat. Prod. 2008, 71, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Cao, Z.Y.; Murray, T.F.; Gerwick, W.H. Hoiamide A, a sodium channel activator of unusual architecture from a consortium of two Papua New Guinea cyanobacteria. Chem. Biol. 2009, 16, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Spoof, L.; Błaszczyk, J.M.; Cegłowska, M.; Marzec, H.M. Structures and activity of new anabaenopeptins produced by Baltic Sea cyanobacteria. Mar. Drugs 2016, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Maru, N.; Ohno, O.; Uemura, D. Lyngbyacyclamides A and B, novel cytotoxic peptides from marine cyanobacteria Lyngbya sp. Tetrahedron 2010, 51, 6384–6387. [Google Scholar] [CrossRef]
- Boyaud, F.; Mahiout, Z.; Lenoir, C.; Tang, S.; Wdzieczak-Bakala, J.; Witczak, A.; Bonnard, I.; Banaigs, B.; Ye, T. First total synthesis and stereochemical revision of laxaphycin B and Its Extension to lyngbyacyclamide A and Nicolas Inguimbert. Org. Lett. 2013, 15, 3898–3901. [Google Scholar] [CrossRef] [PubMed]
- Linington, R.G.; González, J.; Ureña, L.; Romero, L.I.; Ortega-Barría, E.; Gerwick, W.H. Venturamides A and B: Antimalarial constituents of the Panamanian marine cyanobacterium Oscillatoria sp. J. Nat. Prod. 2007, 70, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.A.V.; Al-Lihaibi, S.S.; Alarif, W.M.; Abdel-Lateff, A.; Nogata, Y.; Washio, K.; Morikawa, M.; Okino, T. Wewakazole B, a Cytotoxic Cyanobactin from the Cyanobacterium Moorea producens Collected in the Red Sea. J. Nat. Prod. 2016, 79, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Long, B.H.; Zhang, J.Z.; Tang, X.D.; Wu, Z.Z. Total synthesis of wewakazole B. Org. Biomol. Chem. 2016, 14, 9712–9715. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structures of compounds 1–8.
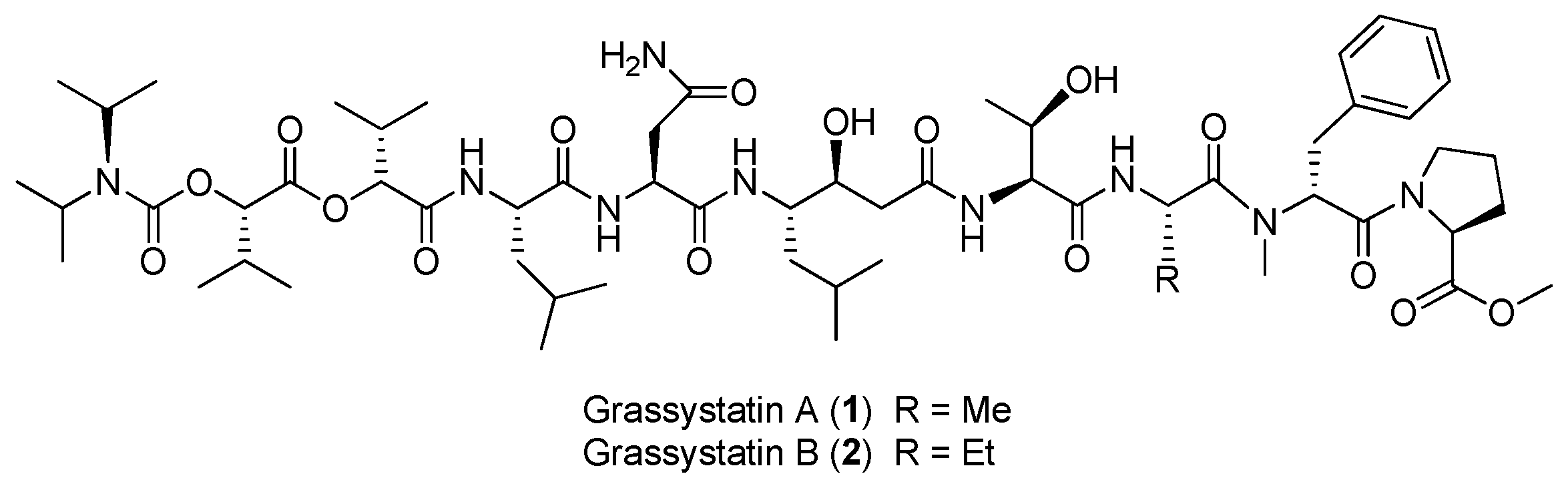
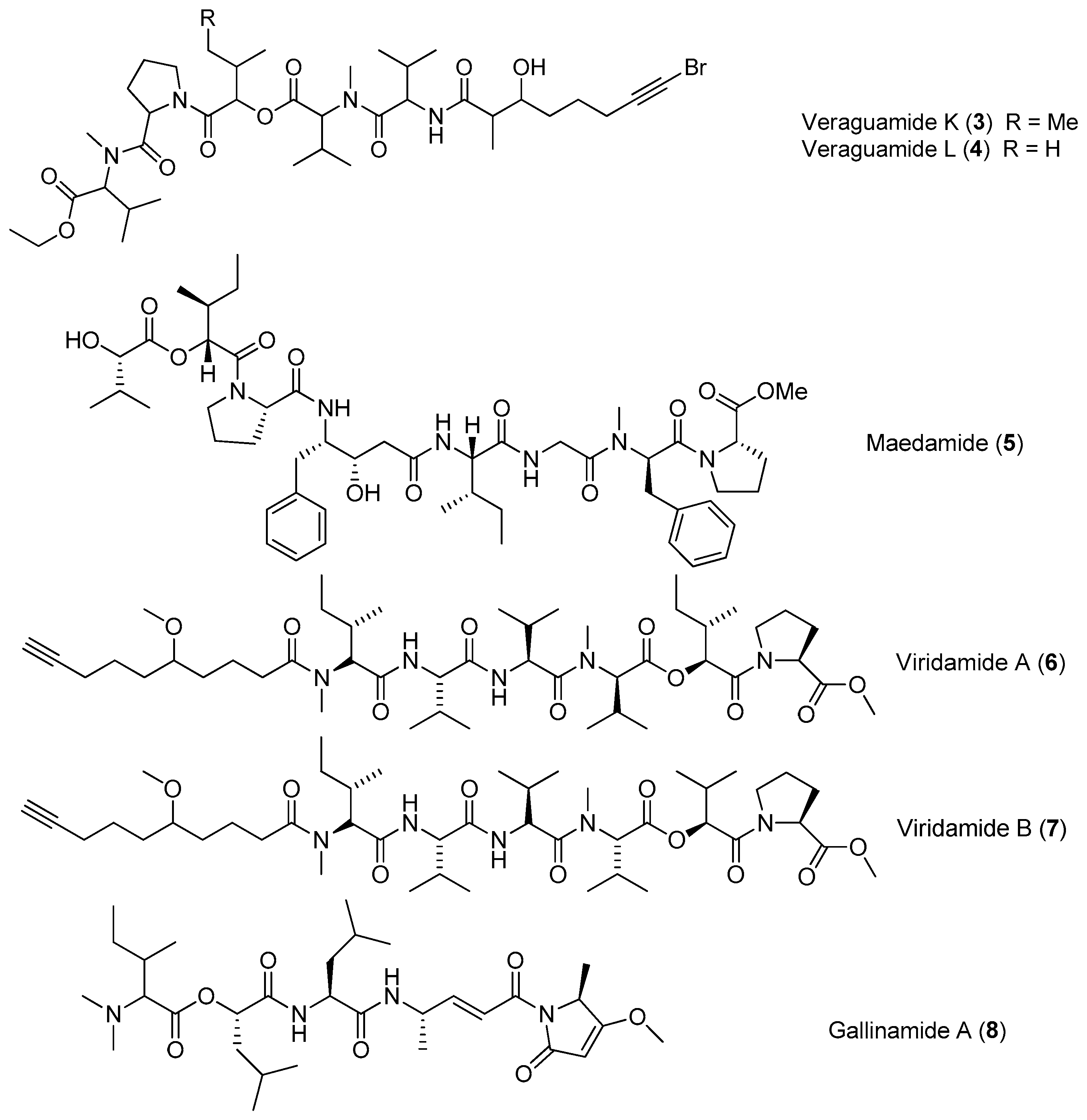
Figure 2.
Chemical structures of compounds 9–16.

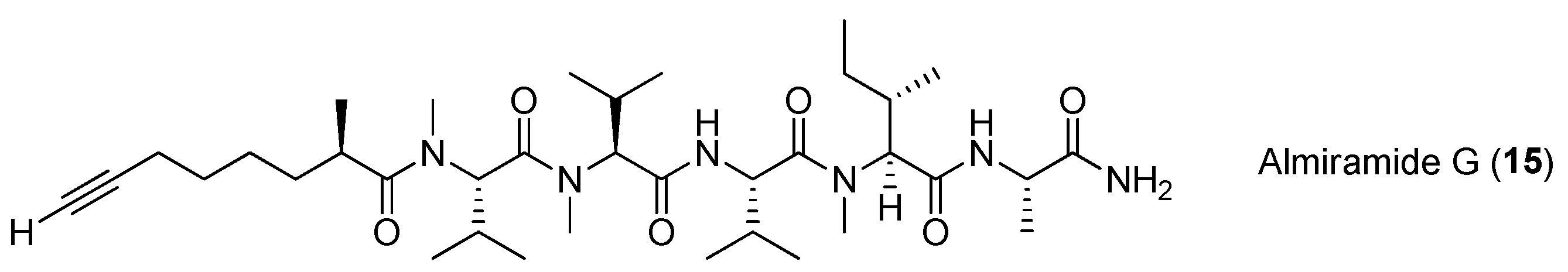
Figure 3.
Chemical structures of compounds 17–23.
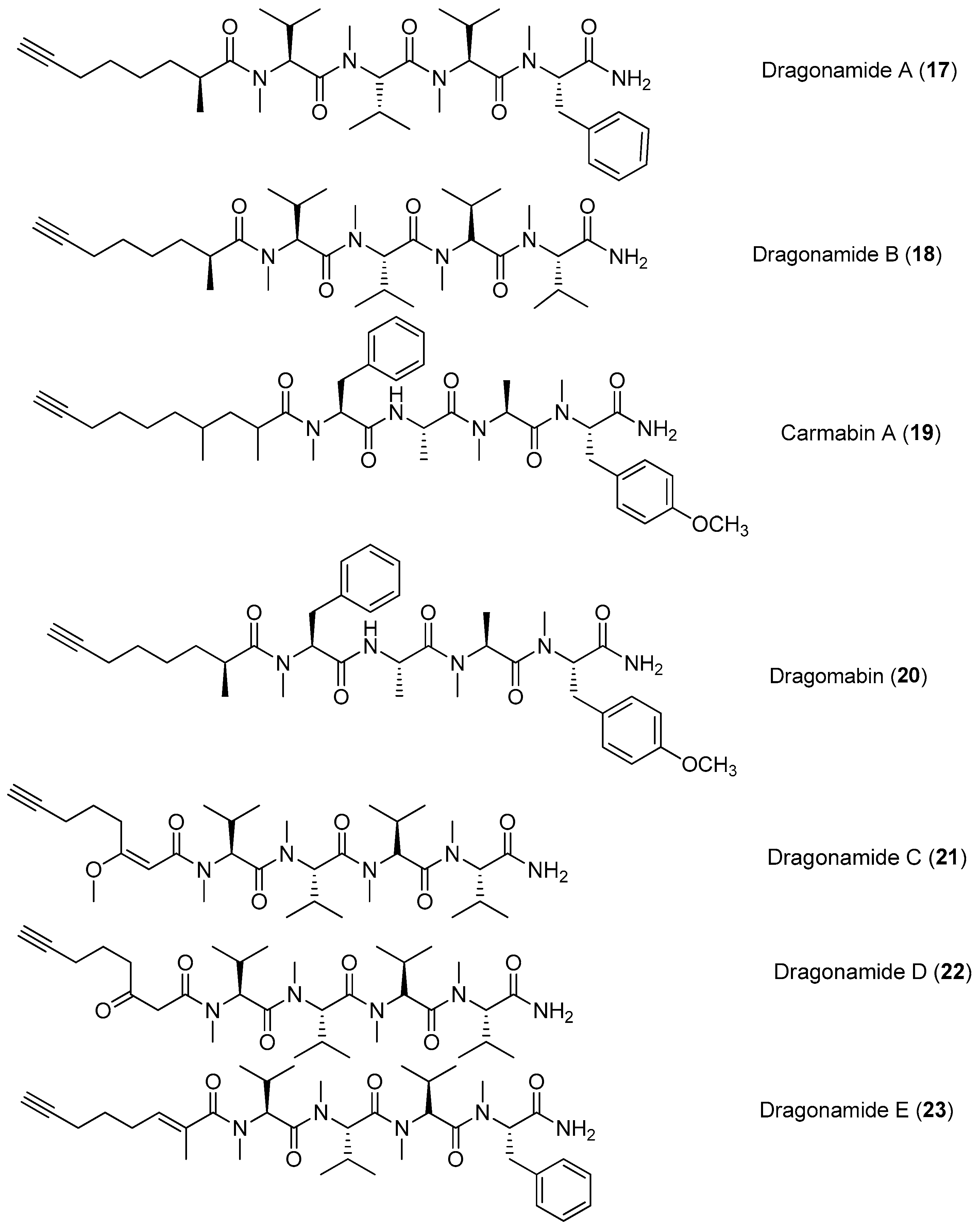
Figure 4.
Chemical structures of compounds 24–27.

Figure 5.
Chemical structures of compounds 28–32.
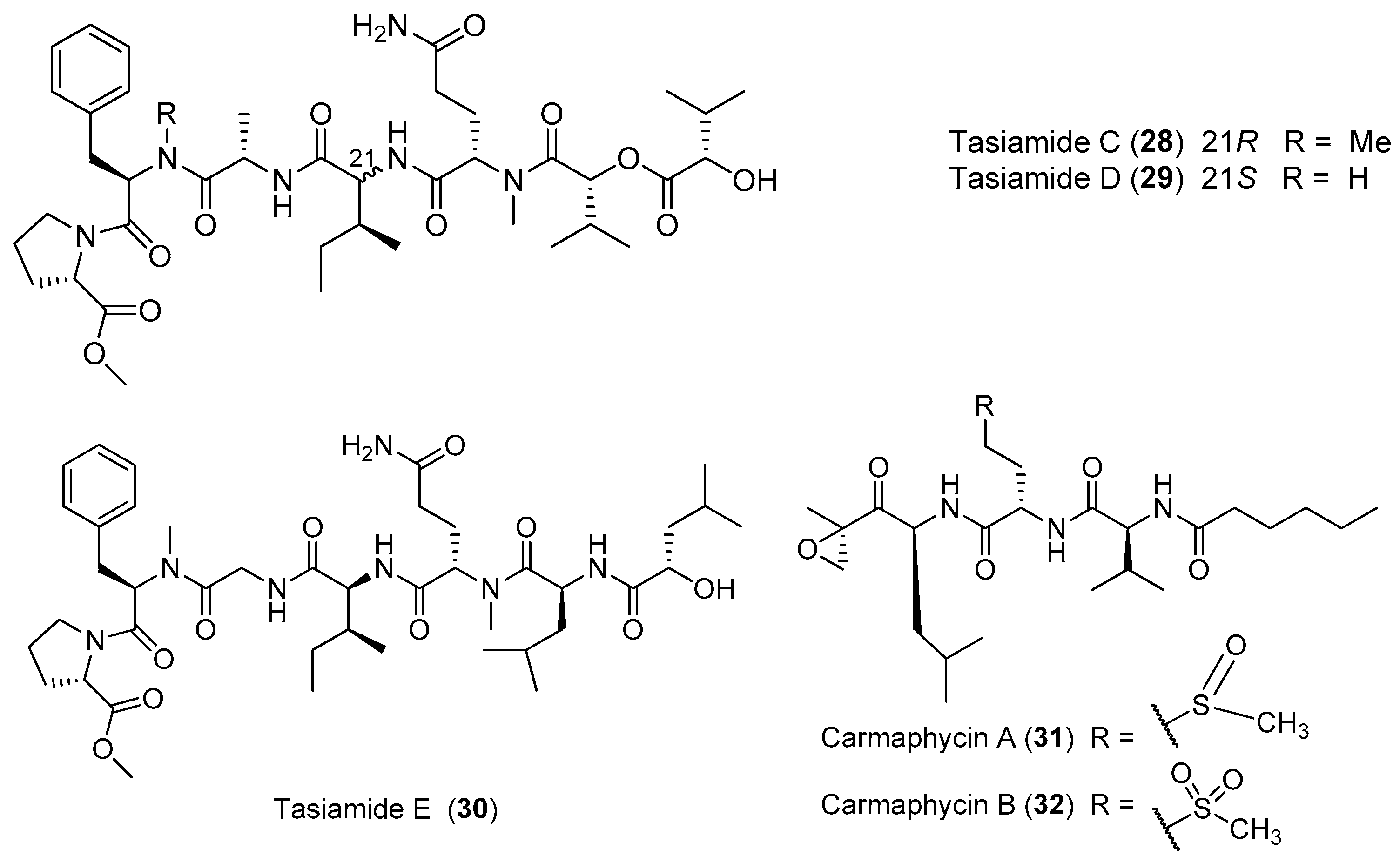
Figure 6.
Chemical structures of compounds 33–35.

Figure 7.
Chemical structures of compounds 36–39.

Figure 8.
Chemical structures of compounds 40–49.

Figure 9.
Chemical structures of compounds 50–60.
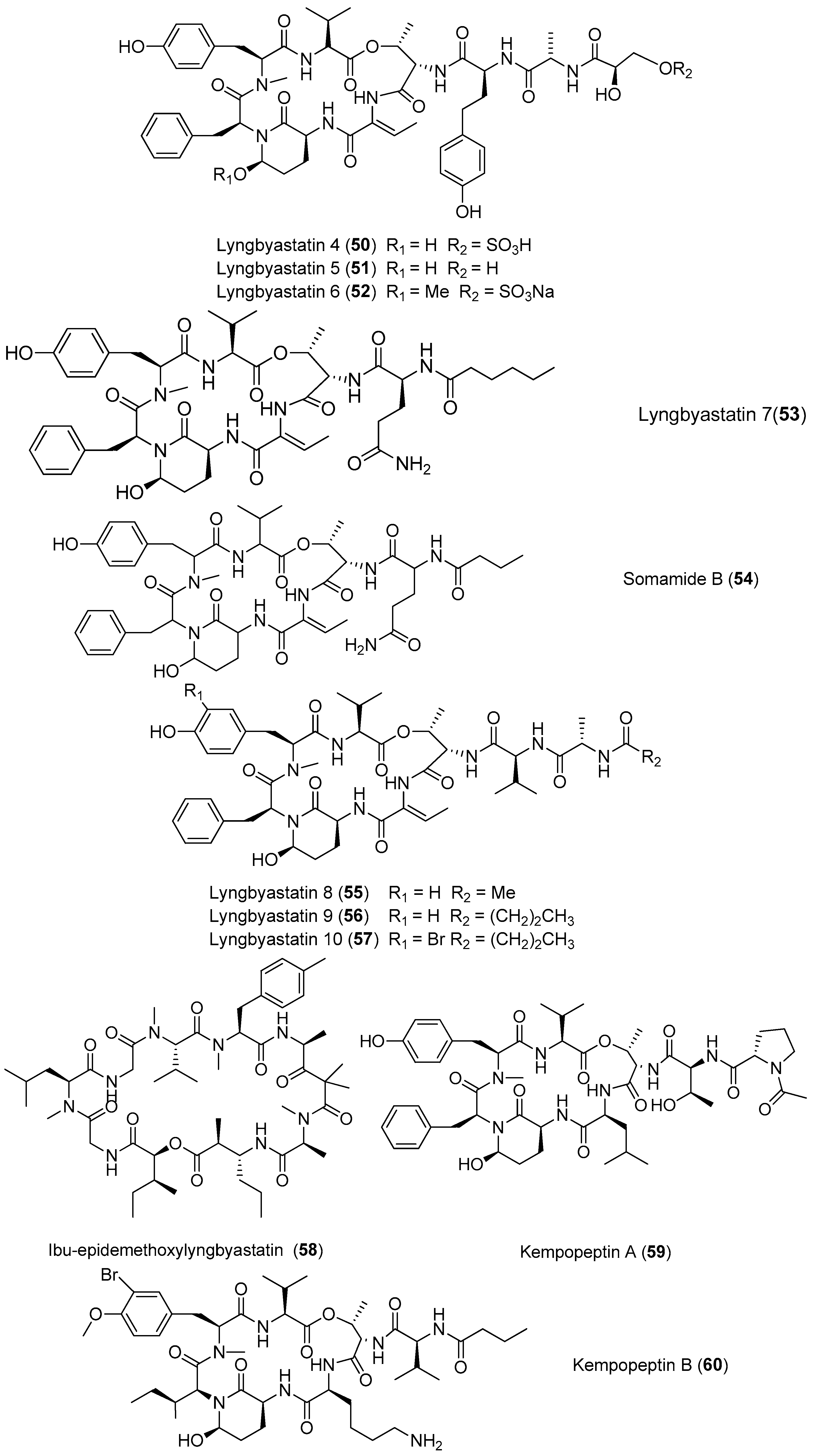
Figure 10.
Chemical structures of compounds 61–67.
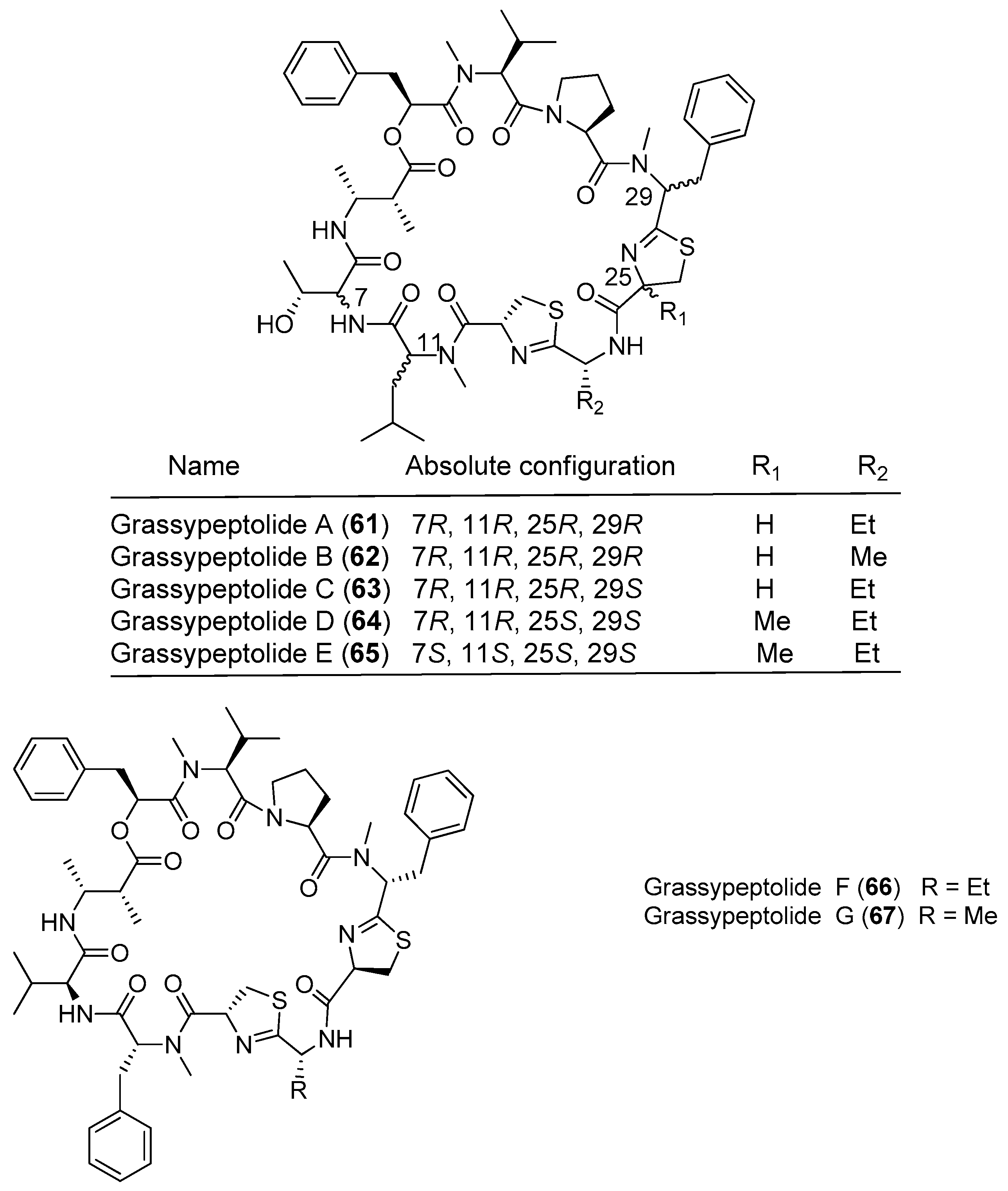
Figure 11.
Chemical structures of compounds 68–74.
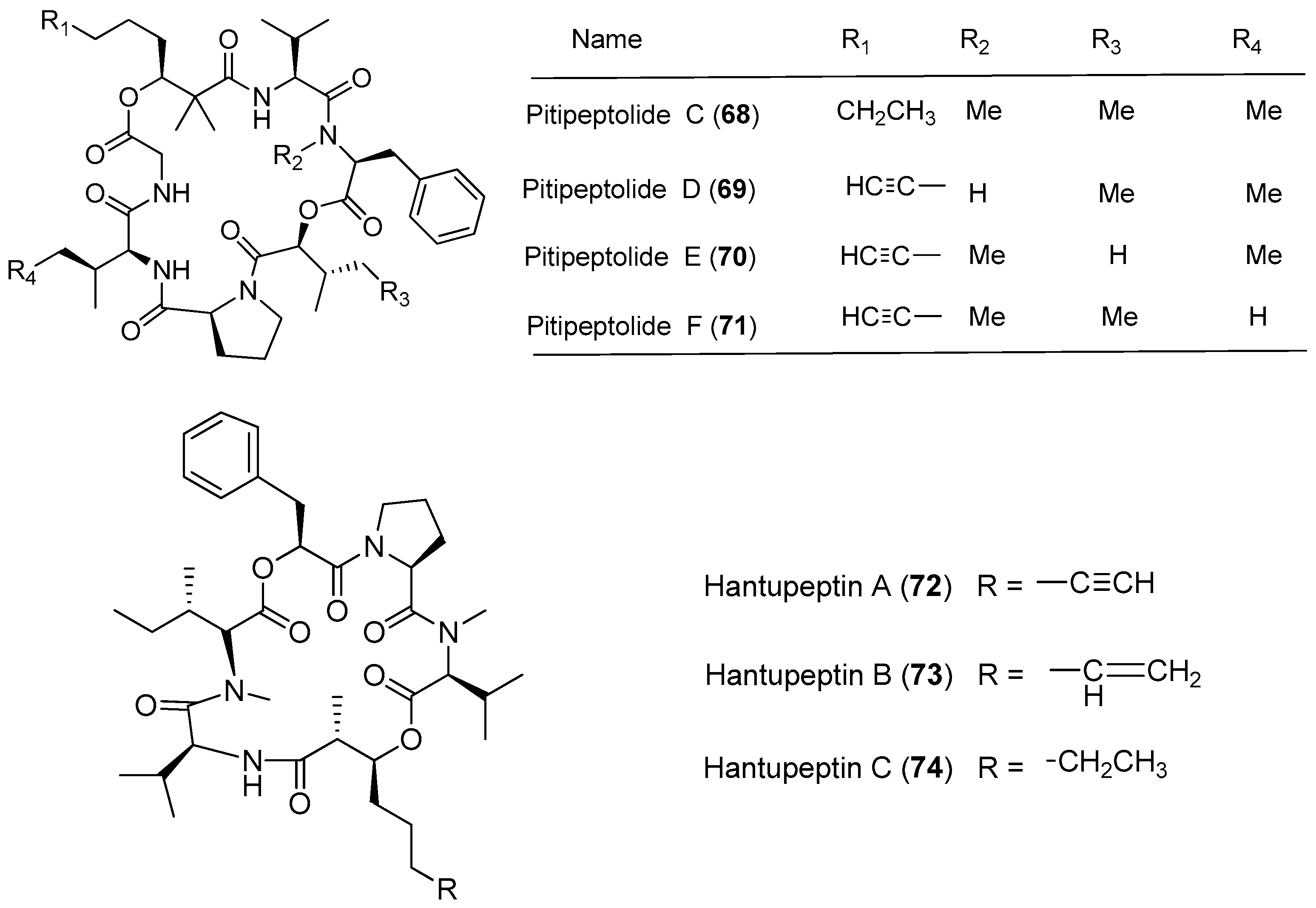
Figure 12.
Chemical structures of compounds 75–82.

Figure 13.
Chemical structures of compounds 83–90.
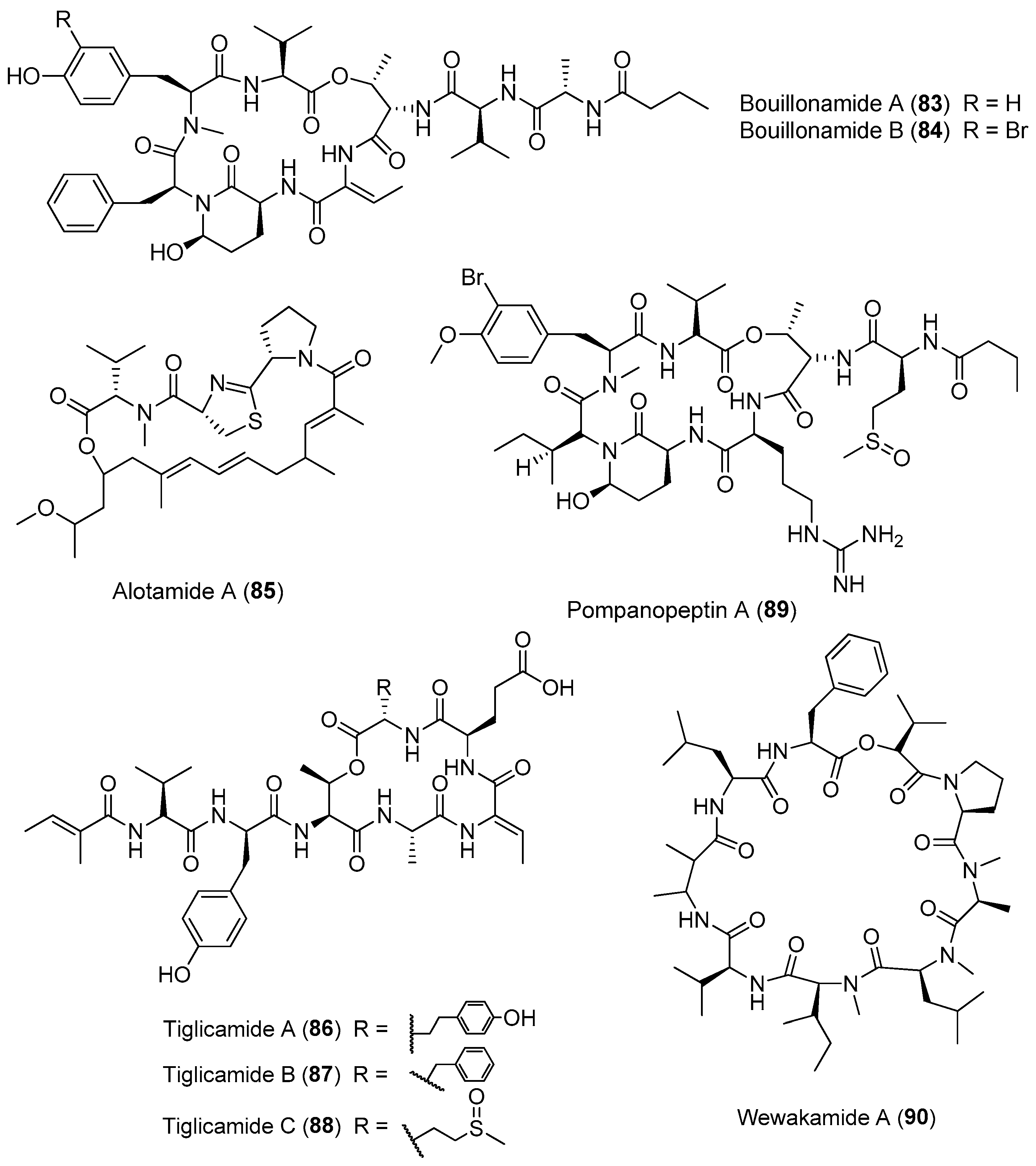
Figure 14.
Chemical structures of compounds 91–96.
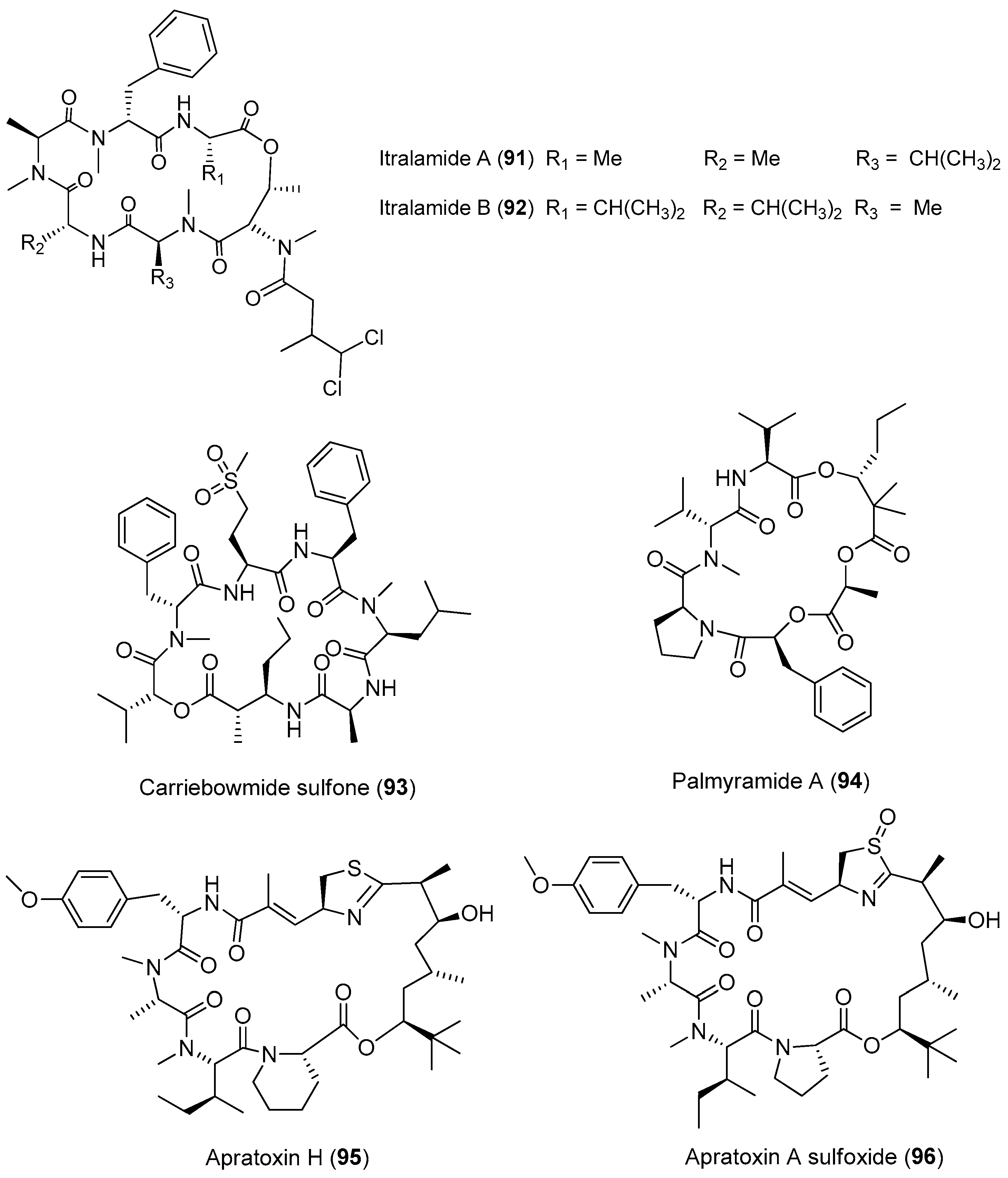
Figure 15.
Chemical structures of compounds 97–103.
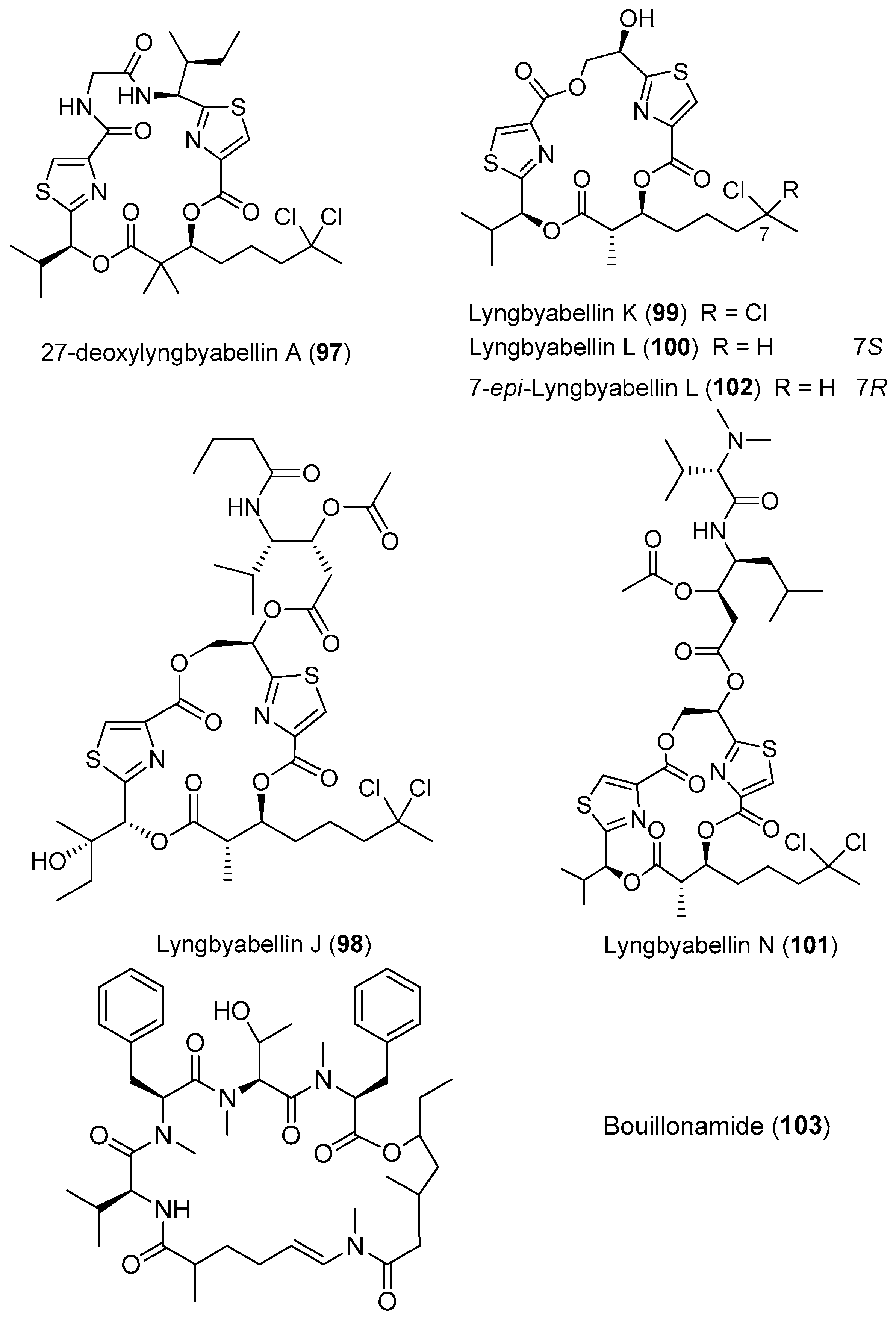
Figure 16.
Chemical structures of compounds 104–115.
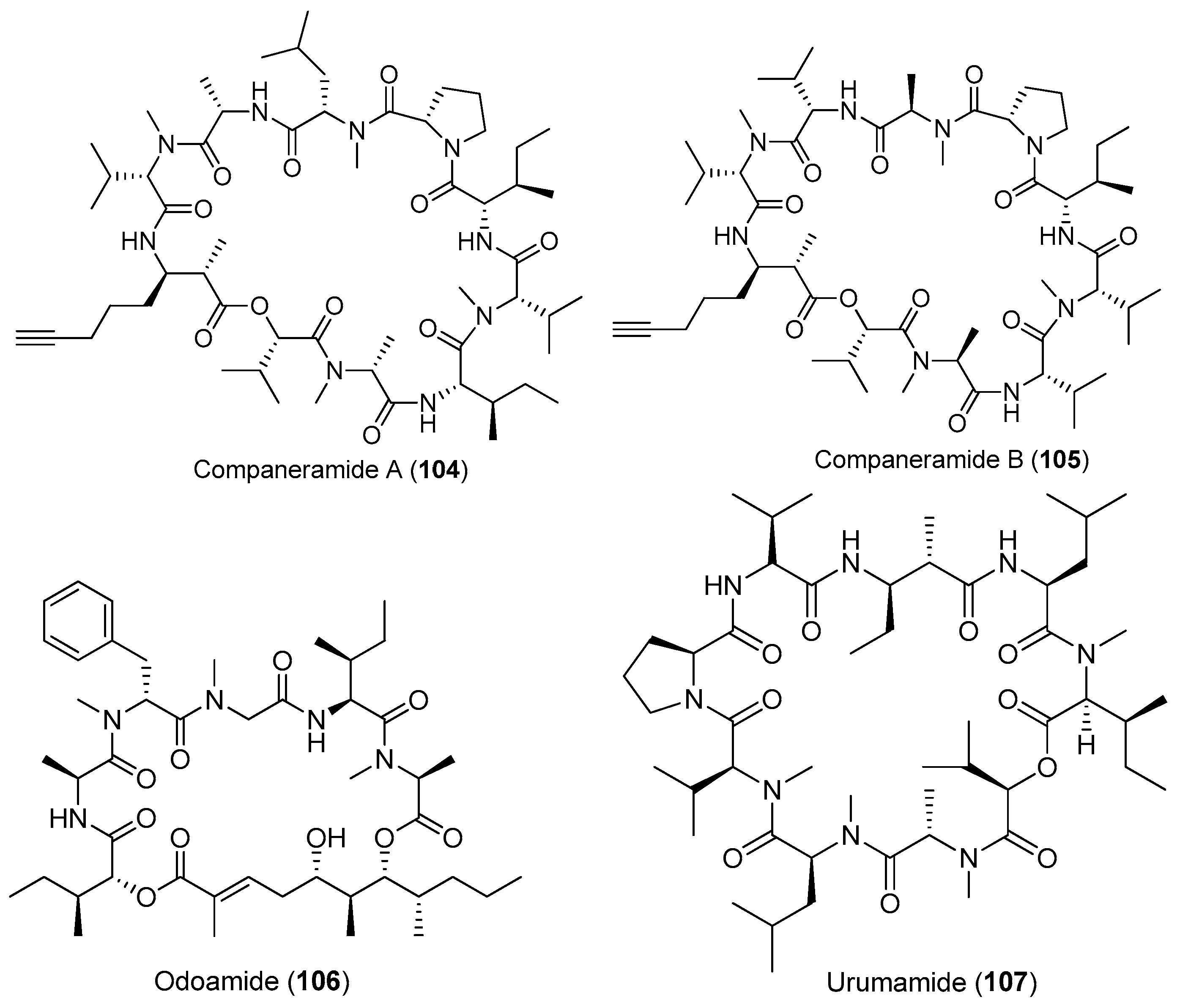
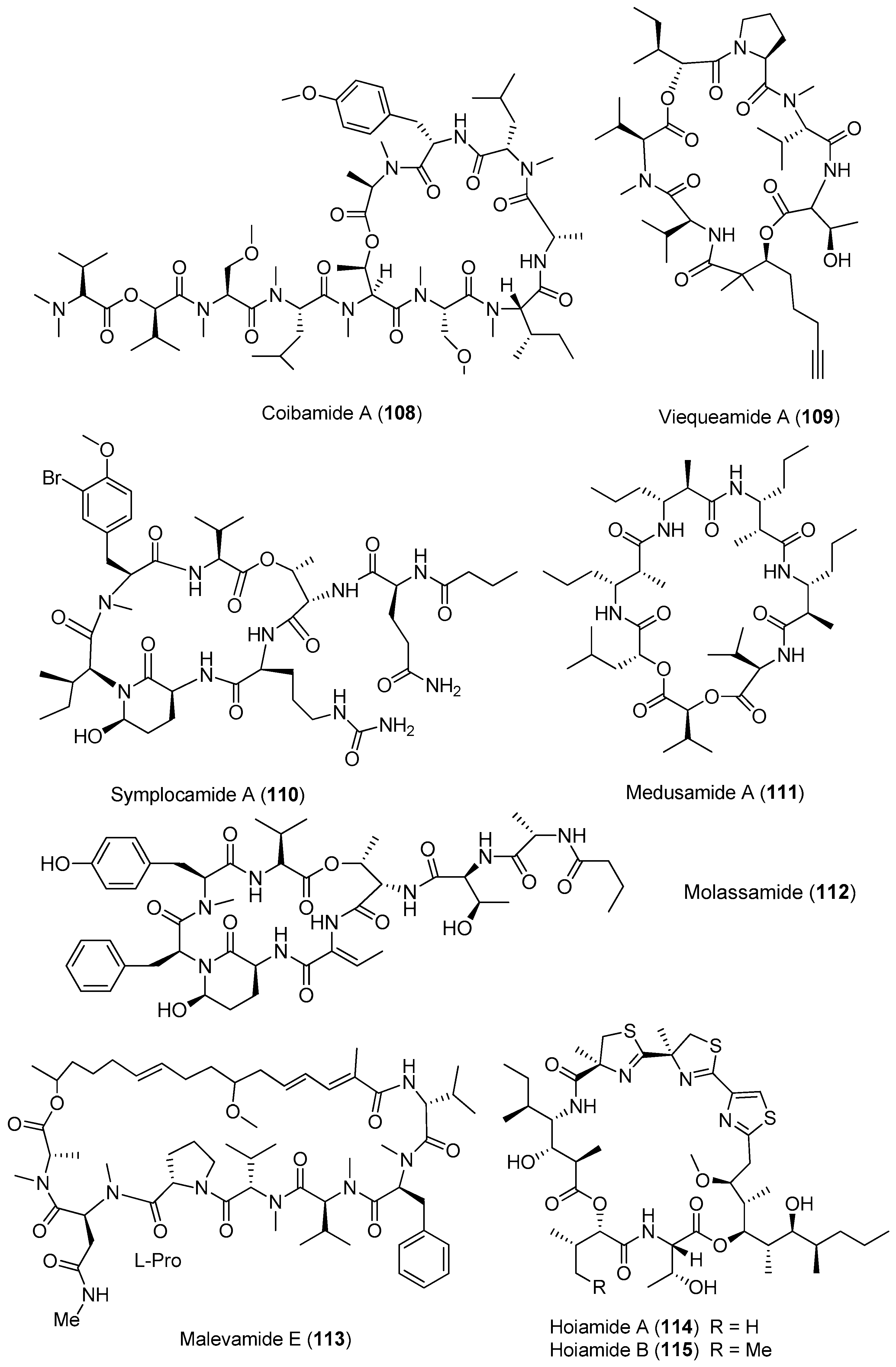
Figure 17.
Chemical structures of compounds 116–126.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bioactivities of linear depsipeptides from marine cyanobacteria.
| Metabolites | Sources | Activities | References |
|---|---|---|---|
| Grassystatins A–B (1–2) | Okeania lorea (formerly Lyngbya cf. confervoides) | Cathepsin inhibition | [12,13] |
| Veraguamides K–L (3–4) | cf. Oscillatoria margaritifera Coiba, Panama | nd a | [14] |
| Maedamide (5) | Lyngbya sp. | Potent antitumor cytotoxicity Protease inhibition | [15,16] |
| Viridamides A–B (6–7) | Okeania comitata (formerly Oscillator nigroviridis) Panama | Antitrypanosomal activity Antileishmanial activity | [17] |
| Gallinamide A (8) | Schizothrix iedras Gallinas | Antimalarial activity | [18,19] |
a Not determined.
Table 2.
Bioactivities of linear peptides from marine cyanobacteria.
| Metabolites | Sources | Bioactivities | References |
|---|---|---|---|
| Almiramides A–C (9–11) | Lyngbya majuscula Panama | General antileishmanial activity Antitumor cytotoxicity | [20,21] |
| Almiramide D (12) | Oscillatoria nigroviridis Island of Providence (Colombia, S.W. Caribbean Sea) | Antitumor cytotoxicity | [21] |
| Almiramide E–H (13–16) | Oscillatoria nigroviridis Island of Providence (Colombia, S.W. Caribbean Sea) | nd a | [21] |
| Dragonamides A–B (17–18) Carmabin A (19) Dragomabin (20) | Moorea producens (formerly Lyngbya polychroa) Panama | Antimalarial activity | [22] |
| Dragonamides C–D (21–22) | Moorea producens (formerly Lyngbya polychroa) Florida, Fort Lauderdale, Hollywood | Weak antitumor cytotoxicity | [23,24] |
| Dragonamide E (23) | Lyngbya majuscula | Antileishmanial activity | [25] |
| Lyngbyapeptin D (24) | Moorea bouillonii (formerly Lyngbya bouillonii) Apra Harbor, Guam | nd a | [26] |
| Jahanyne (25) | Lyngbya sp. | Potent antitumor cytotoxicity | [27] |
| Bisebromoamide (26) | Lyngbya sp. | Protein kinase inhibition | [28,29,30,31] |
| Norbisebromoamide (27) | Lyngbya sp. | nd a | [29] |
| Tasiamides C–D (28–29) | Symploca sp. Papua New Guinea | Weak antitumor cytotoxicity | [32] |
| Tasiamide E (30) | Symploca sp. Papua New Guinea | nd a | [32] |
| Carmaphycins A–B (31–32) | Symploca sp. Curacao | Protease inhibition Potent antitumor cytotoxicity | [33] |
| Hoiamides C–D (33–34) | Cyanobacterium Papua New Guinea | nd a | [34,35,36] |
| Lyngbyabellin M (35) | cyanobacterium from Palmyra Atoll Central Pacific Ocean | nd a | [37] |
| Kurahyne (36) | cyanobacterial mixture | Antitumor cytotoxicity | [38] |
| Kurahyne B (37) | Okeania sp. | Mild antitumor cytotoxicity | [39] |
| Caldoramide (38) | Caldora penicillata Florida | Antitumor cytotoxicity | [40] |
| Grassystatin C (39) | Okeania lorea (formerly Lyngbya cf. confervoides) | Cathepsin inhibition | [12] |
a Not determined.
Table 3.
Bioactivities of cyclic depsipeptides (40–67) from marine cyanobacteria.
| Structure Class | Metabolites | Sources | Activities | References |
|---|---|---|---|---|
| Veraguamides | Veraguamides A–G (40–46) | cf. Oscillatoria margaritifera, Panama Symploca cf. hydnoides Cetti Bay, Guam | Weak antitumor cytotoxicity | [14,41,42] |
| Veraguamides H–J (47–49) | cf. Oscillatoria margaritifera Panama | nd a | [14] | |
| Lyngbyastatins | Lyngbyastatins 4–6 (50–52) | Lyngbya confervoides off the coast of Florida | Potent protease inhibition | [43,44] |
| Lyngbyastatin 7 (53) Somamide B (54) | Lyngbya sp. from Florida | Potent protease inhibition | [44,45] | |
| Lyngbyastatins 8–10 (55–57) | Lyngbya semiplena Tumon Bay, Guam | Potent protease inhibition | [46] | |
| Ibu-epidemethoxylyngbyastatin 3 (58) | Leptolyngbya sp. SS Thistlegorm shipwreck, Red Sea | Weak cytotoxicity to neuro-2a cells | [47] | |
| Kempopeptins A and B (59, 60) | Lyngbya sp. Florida | Potent protease inhibition | [48] | |
| Grassypeptolide A (61) | Okeania lorea (formerly Lyngbya confervoides) off Grassy Key in Florida | Antitumor cytotoxicity | [49] | |
| Grassypeptolides A–C (61–63) | Okeania lorea (formerly Lyngbya confervoides) | Cause G1 and G2/M phase cell cycle arrest | [50,51] | |
| Grassypeptolides | Grassypeptolides D and E (64, 65) | Leptolyngbya sp. SS Thistlegorm shipwreck, Red Sea | Potent antitumor cytotoxicity | [47] |
| Grassypeptolides F and G (66, 67) | Lyngbya majuscula Panama | Moderate inhibitory activity against the transcription factor AP-1 | [52] |
Table 4.
Bioactivities of cyclic depsipeptides (68–115) from marine cyanobacteria.
| Sources | Metabolites | Sources/Location | Activities | References |
|---|---|---|---|---|
| Lyngbya majuscula | Pitipeptolides C–E (68–70) | Guam, Piti Bomb Holes | nd a | [53] |
| Pitipeptolide F (71) | Guam, Piti Bomb Holes | Antibacterial activity | [53] | |
| Hantupeptins A–C (72–74) | Pulau Hantu Besar Singapore | Moderate antitumor cytotoxicity | [54,55] | |
| Lagunamides A–C (75–77) | Pulau Hantu Besar Singapore | Antimalarial activity Potent antitumor cytotoxicity | [56,57,58] | |
| Cocosamides A and B (78, 79) | Cocos Lagoon, Guam | Slight antitumor cytotoxicity | [59] | |
| Desmethoxymajusculamide C (80) | Fijian | Potent antitumor cytotoxicity | [60] | |
| Pitiprolamide (81) | Piti Bomb Holes, Guam | Weak antitumor cytotoxicity Weak antibacterial activity | [61] | |
| Guineamide G (82) | Papua New Guinea | Brine shrimp toxicity Cytotoxicity against neuroblastoma cell | [62] | |
| Genus Lyngbya | Bouillomides A and B (83, 84) | Lyngbya bouillonii, Guam | Protease inhibition | [63] |
| Alotamide A (85) | Lyngbya bouillonii Papua New Guinea | Influx of Ca2+ in murine cerebrocortical neurons | [64] | |
| Tiglicamides A–C (86–88) | Lyngbya confervoides Florida | Protease inhibition | [65] | |
| Pompanopeptin A (89) | Lyngbya confervoides Florida | Protease inhibition | [66] | |
| wewakamide A (90) | Lyngbya semiplena Papua New Guinea | Potent brine shrimp toxicity | [62] | |
| Moorea producens | Itralamides A and B (91, 92) | eastern Caribbean | Antitumor cytotoxicity | [67,68] |
| Carriebowmide sulfone (93) | eastern Caribbean | nd a | [67] | |
| Palmyramide A (94) | Palmyra Atoll | Blocks sodium channel in neuro-2a cells Antitumor cytotoxicity | [69] | |
| Apratoxin H (95) Apratoxin A sulfoxide (96) | Gulf of Aqaba, Nabq Mangrovs | Potent antitumor cytotoxicity | [70] | |
| Moorea bouillonii | 27-deoxylyngbyabellin A (97) Lyngbyabellin J (98) | Apra Bay, Guam | Moderate antitumor cytotoxicity | [26] |
| Lyngbyabellins K–L, N (99–101) 7-epi-Lyngbyabellin L (102) | Palmyra Atoll Central Pacific Ocean | Antitumor cytotoxicity | [37] | |
| Bouillonamide (103) | New Britain, Papua New Guinea | Mild toxicity to neuron 2a cells | [71] | |
| Other marine cyanobacteria | Companeramides A andB (104, 105) | cyanobacterium from Panama | Moderate antimalaria parasites | [72] |
| Odoamide (106) | Okeania sp. | Potent antitumor cytotoxicity | [73] | |
| Urumamide (107) | Okeania sp. | Mild antitumor cytotoxicity | [74] | |
| Coibamide A (108) | Caldora penicillata (formerly Leptolyngbya sp.) Panama | Antitumor cytotoxicity | [75,76] | |
| Viequeamide A (109) | Rivularia sp. viequeamides uerto Rico, Vieque | Potent antitumor cytotoxicity | [77,78] | |
| Symplocamide A (110) | Symploca sp. Papua New Guinea | Potent antitumor cytotoxicity | [79] | |
| Medusamide A (111) | cyanobacterium from Panama | nd a | [80] | |
| Molassamide (112) | Dichothrix utahensis Molasses Reef, Key Largo, Florida | Protease inhibition | [81] | |
| Malevamide E (113) | Symploca laeteviridis | Inhibits Ca2+ release activated Ca2+ (CRAC) channels | [82] | |
| hoiamide A (114) | Lyngbya majuscula and Phormidium gracile Papua New Guinea | nd a | [83] | |
| Hoiamide B (115) | two different cyanobacterium from Papua New Guinea | Activates sodium chanal in mouse neocortical neurons Suppresses spontaneous Ca2+ oscillations in neocortical neurons | [34] |
a Not determined.
Table 5.
Bioactivities of cyclic peptides from marine cyanobacteria.
| Metabolites | Sources | Activities | References |
|---|---|---|---|
| Anabaenopeptins NP 883, NP 867, NP 865, AP813, NP 869 (116–120) | bloom sample of marine cyanobacteria Baltic Sea | nd a | [84] |
| Lyngbyacyclamides A–B (121–122) | Lyngbya sp. Okinawa, Japan | Moderate antitumor cytotoxicity | [85,86] |
| Pompanopeptin B (123) | Lyngbya confervoides Florida | Protease inhibition | [66] |
| Venturamides A and B (124, 125) | Oscillatoria sp. | Antimalaria parasites | [87] |
| Wewakazole B (126) | Moorea producens Red Sea | Moderate antitumor cytotoxicity | [88] |
a Not determined.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mi, Y.; Zhang, J.; He, S.; Yan, X. New Peptides Isolated from Marine Cyanobacteria, an Overview over the Past Decade. Mar. Drugs 2017, 15, 132. https://doi.org/10.3390/md15050132
AMA Style
Mi Y, Zhang J, He S, Yan X. New Peptides Isolated from Marine Cyanobacteria, an Overview over the Past Decade. Marine Drugs. 2017; 15(5):132. https://doi.org/10.3390/md15050132
Chicago/Turabian StyleMi, Yue, Jinrong Zhang, Shan He, and Xiaojun Yan. 2017. "New Peptides Isolated from Marine Cyanobacteria, an Overview over the Past Decade" Marine Drugs 15, no. 5: 132. https://doi.org/10.3390/md15050132
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.