
High-EPA Biomass from Nannochloropsis salina Cultivated in a Flat-Panel Photo-Bioreactor on a Process Water-Enriched Growth Medium


Abstract
:
1. Introduction
2. Results and Discussion
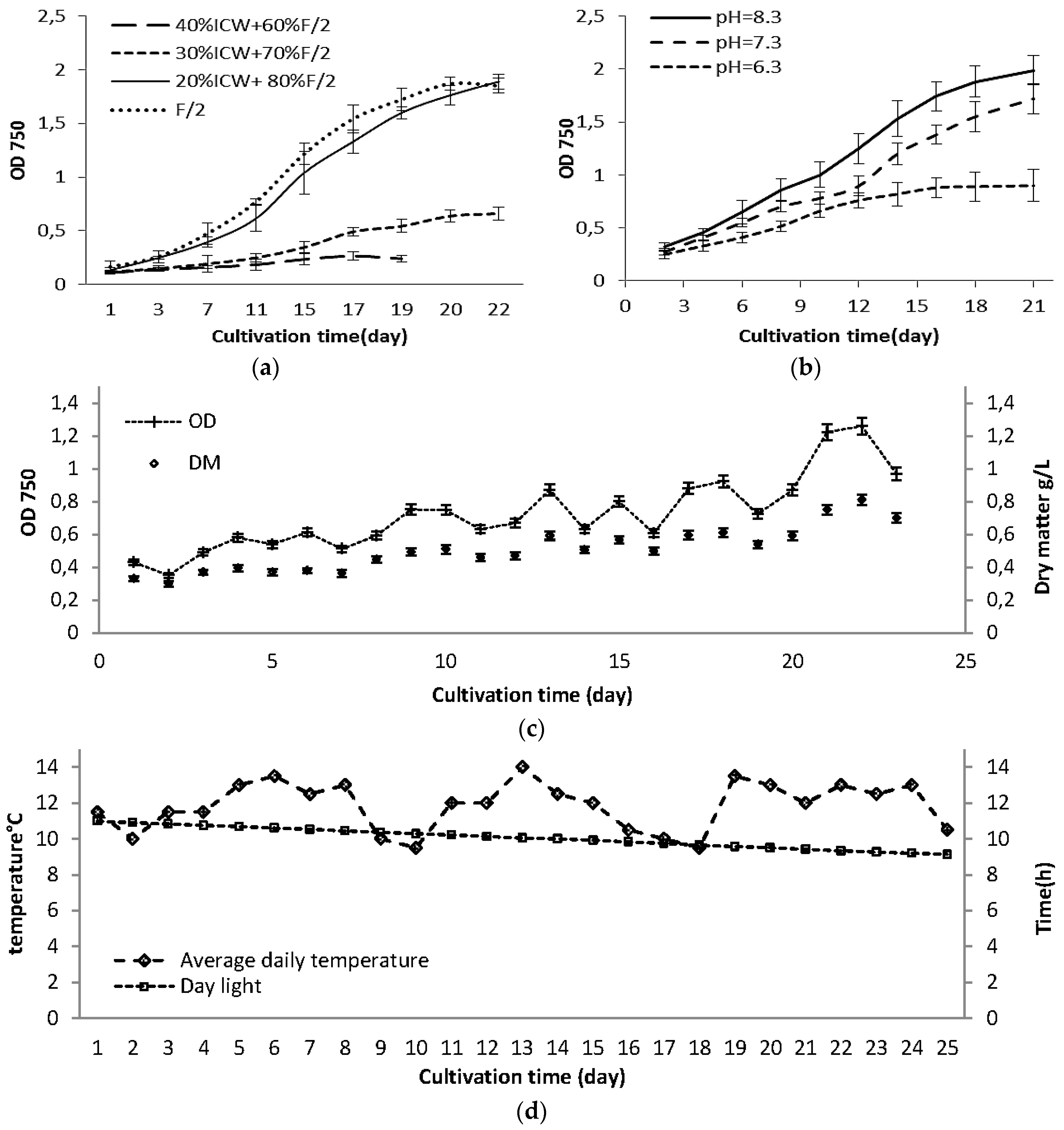
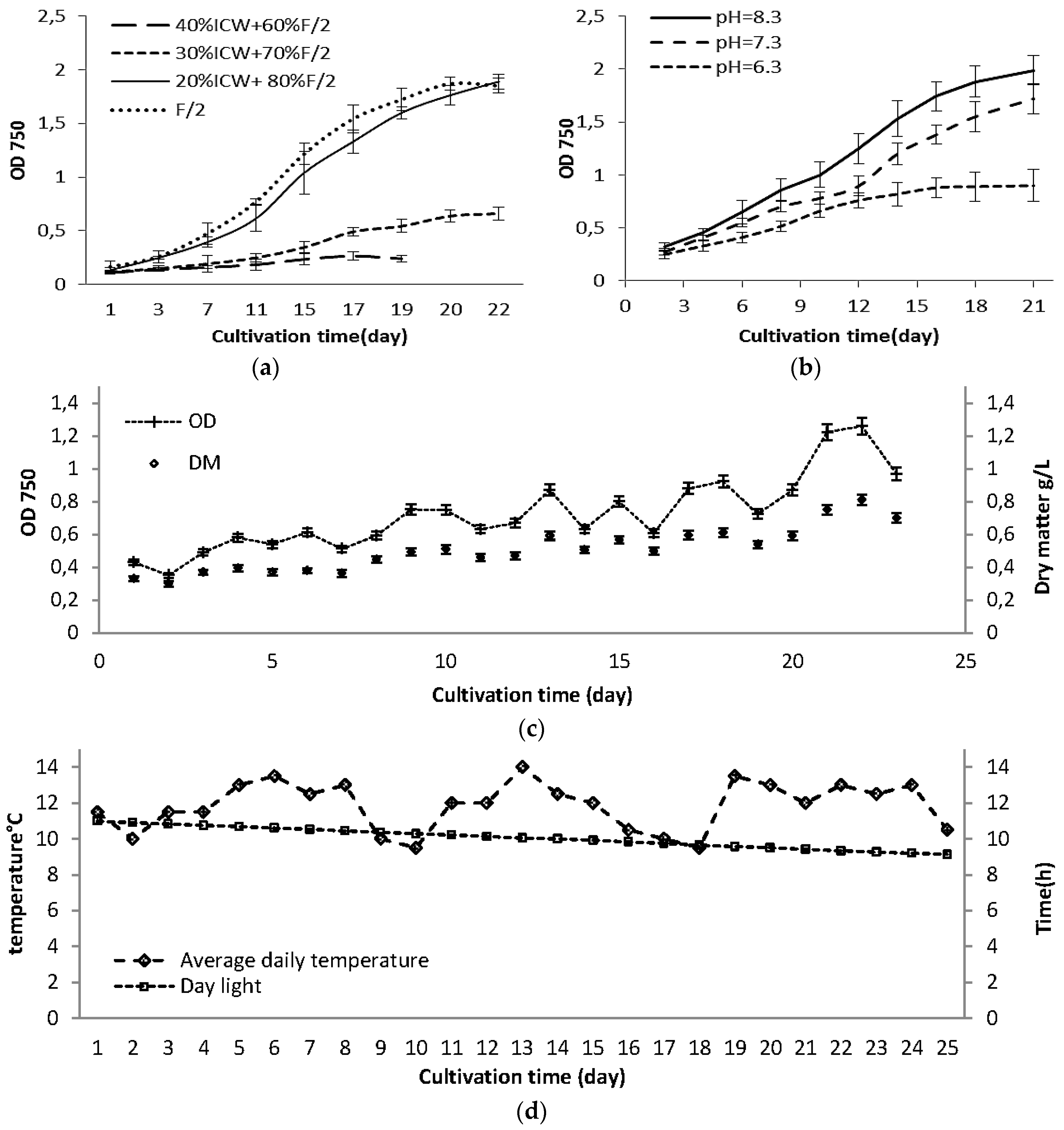
2.1. Growth Rate in Laboratory Scale Experiments
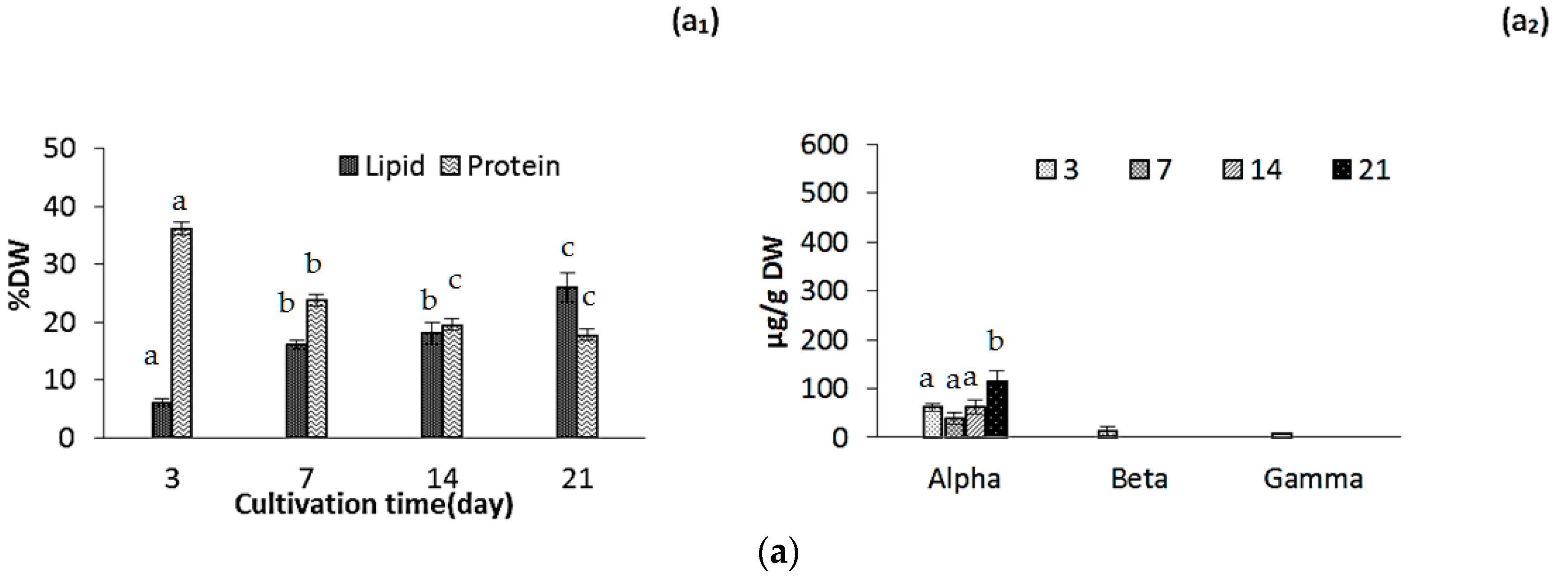
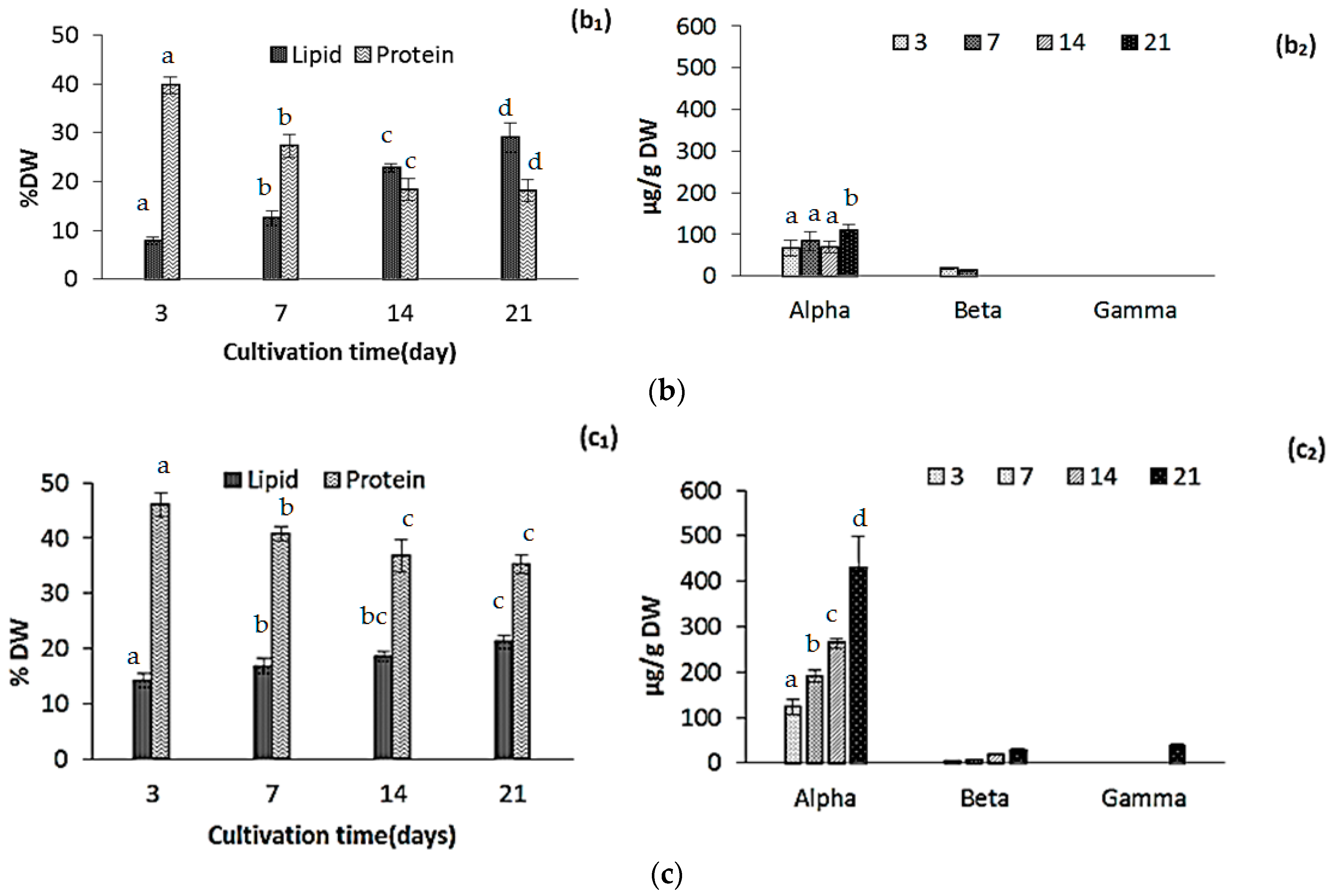
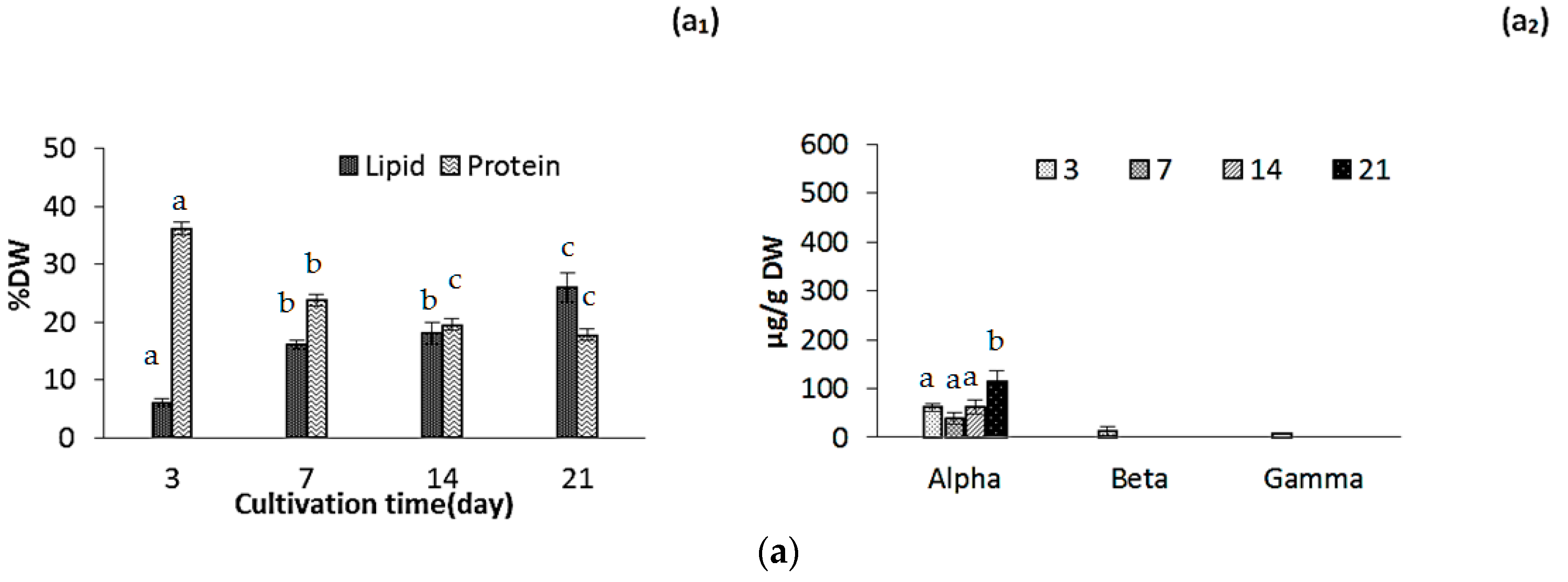
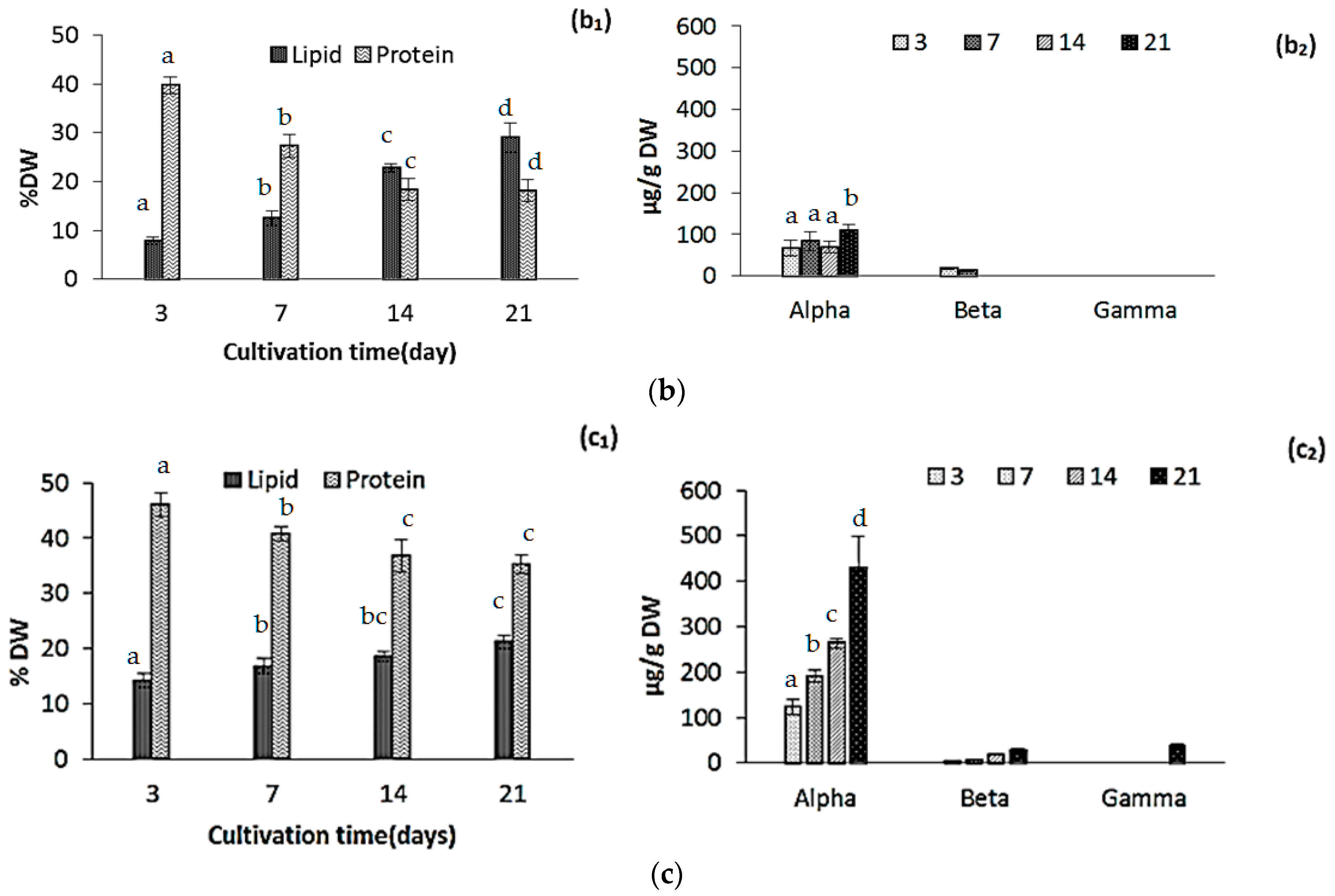
2.2. Changes in Biomass Composition in Response to Growth Media
2.2.1. Protein, Lipids and Tocopherols
2.2.2. Fatty Acid Composition
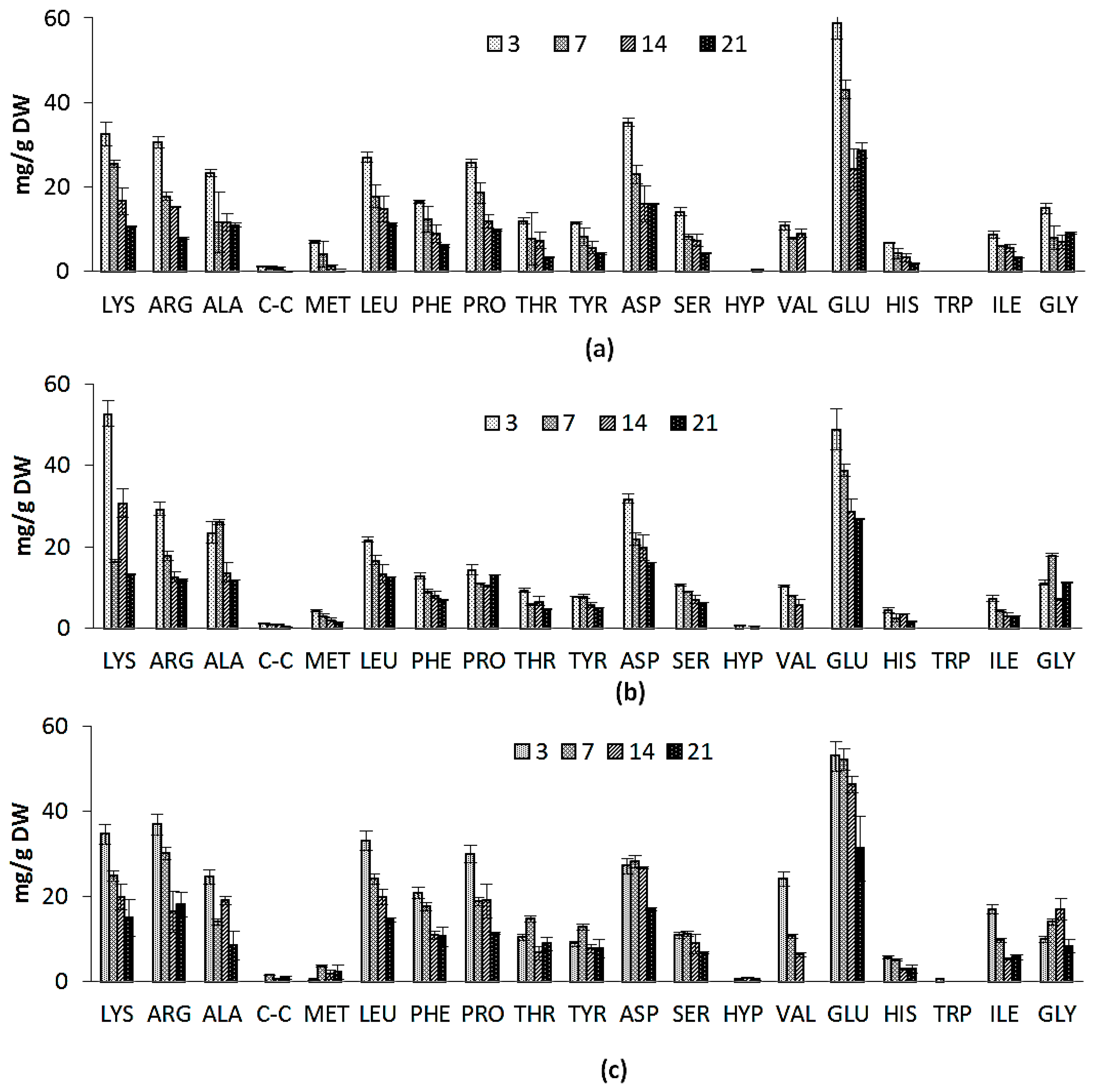
2.2.3. Amino Acid Composition
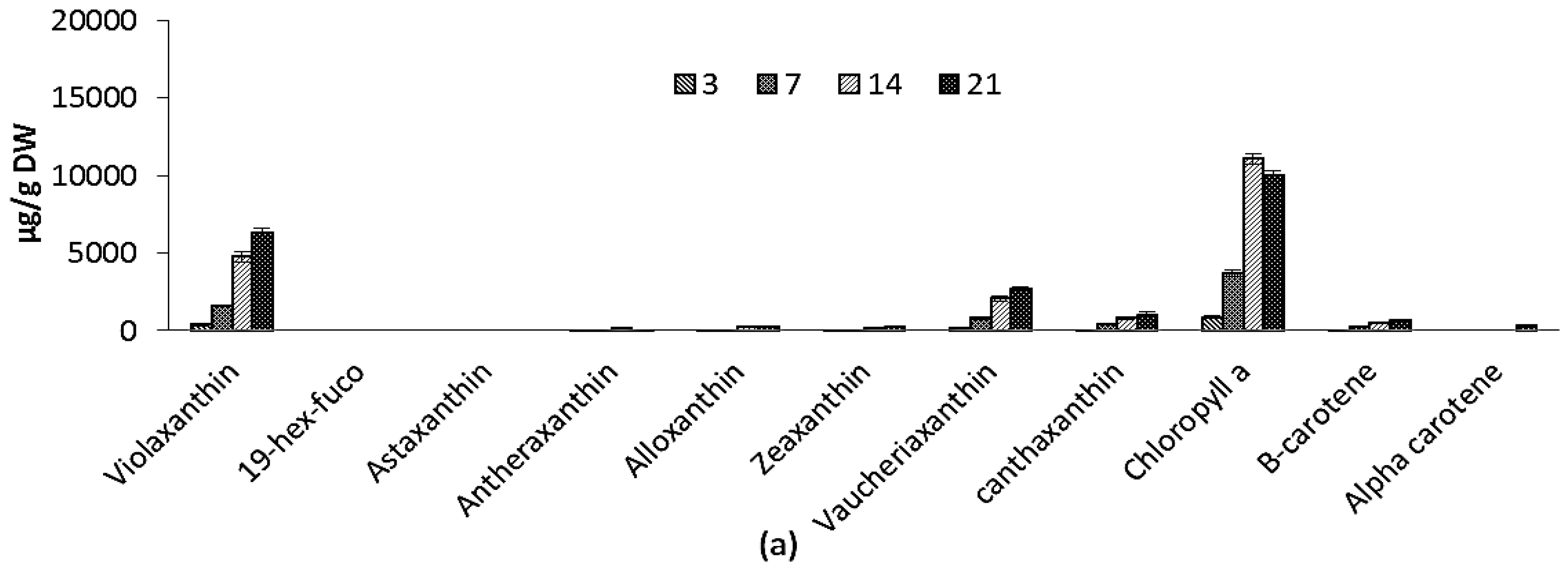
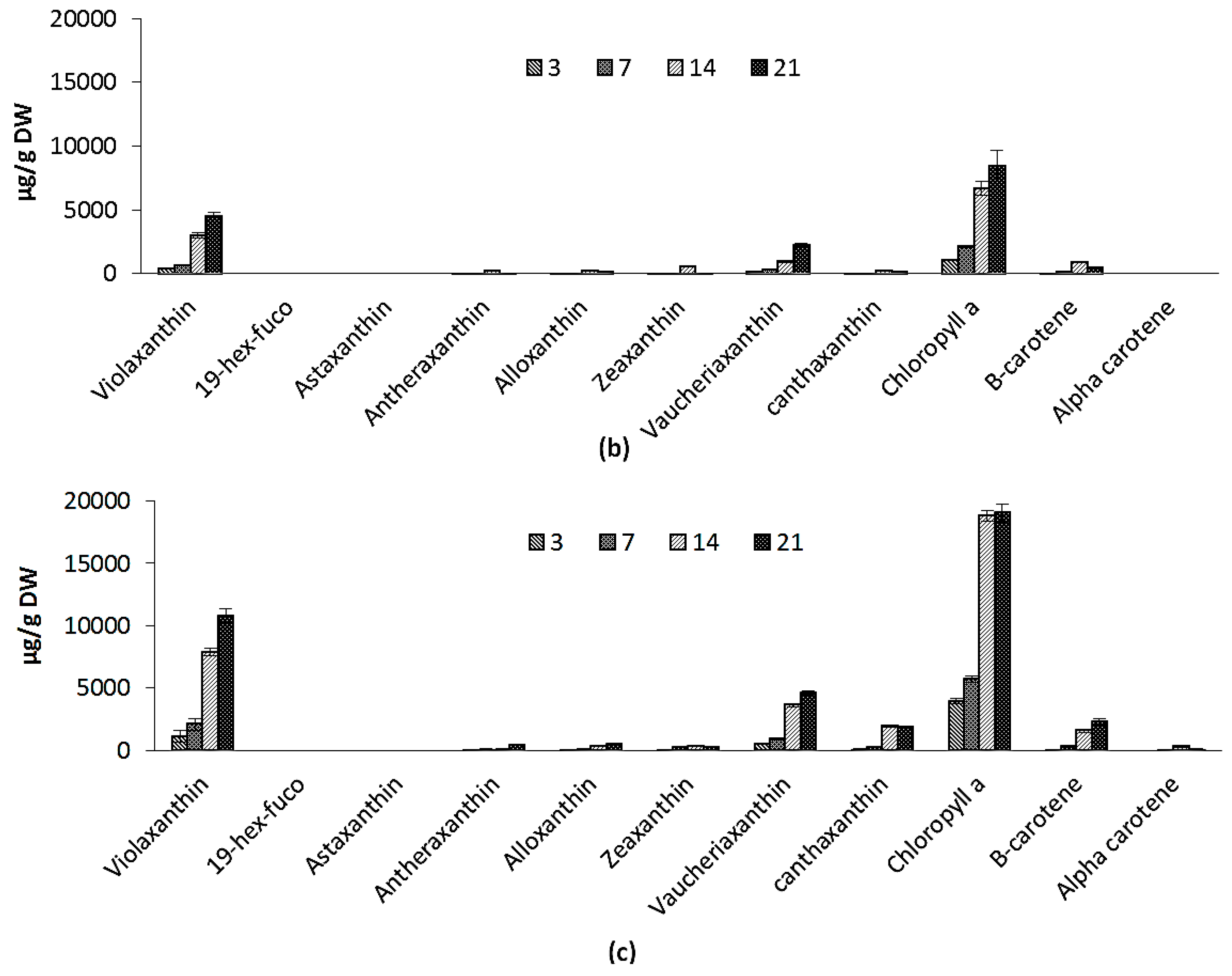
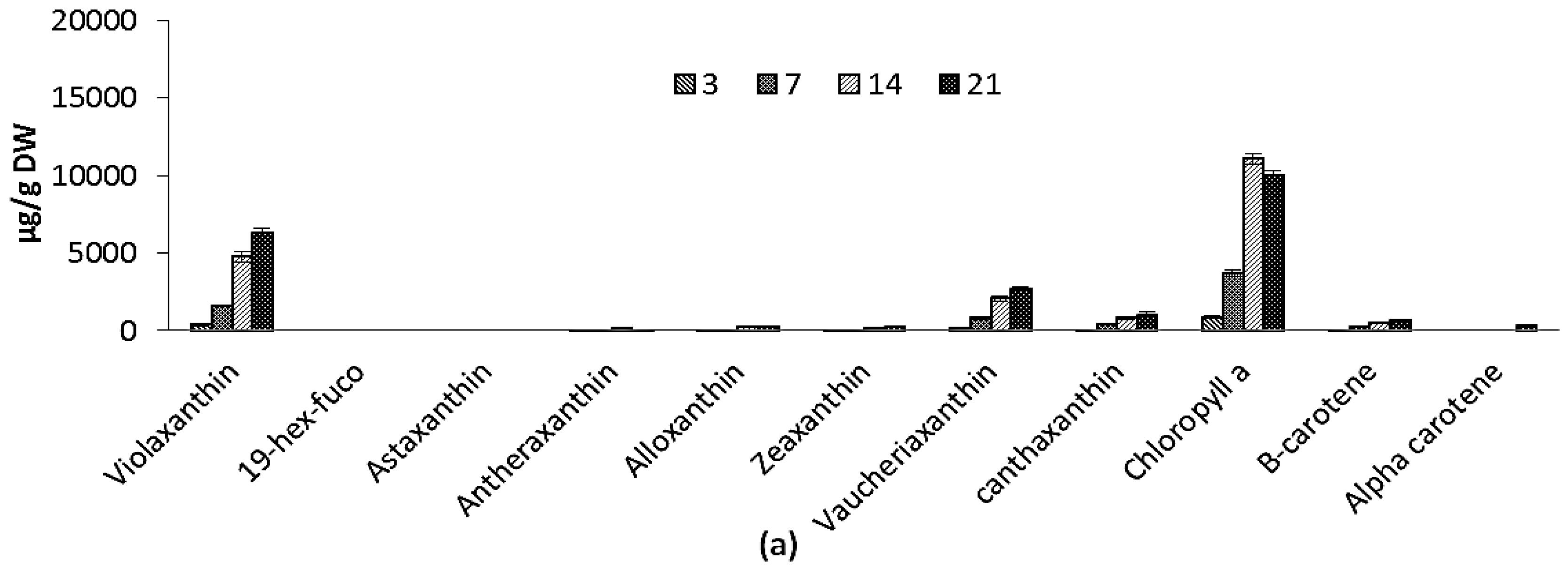
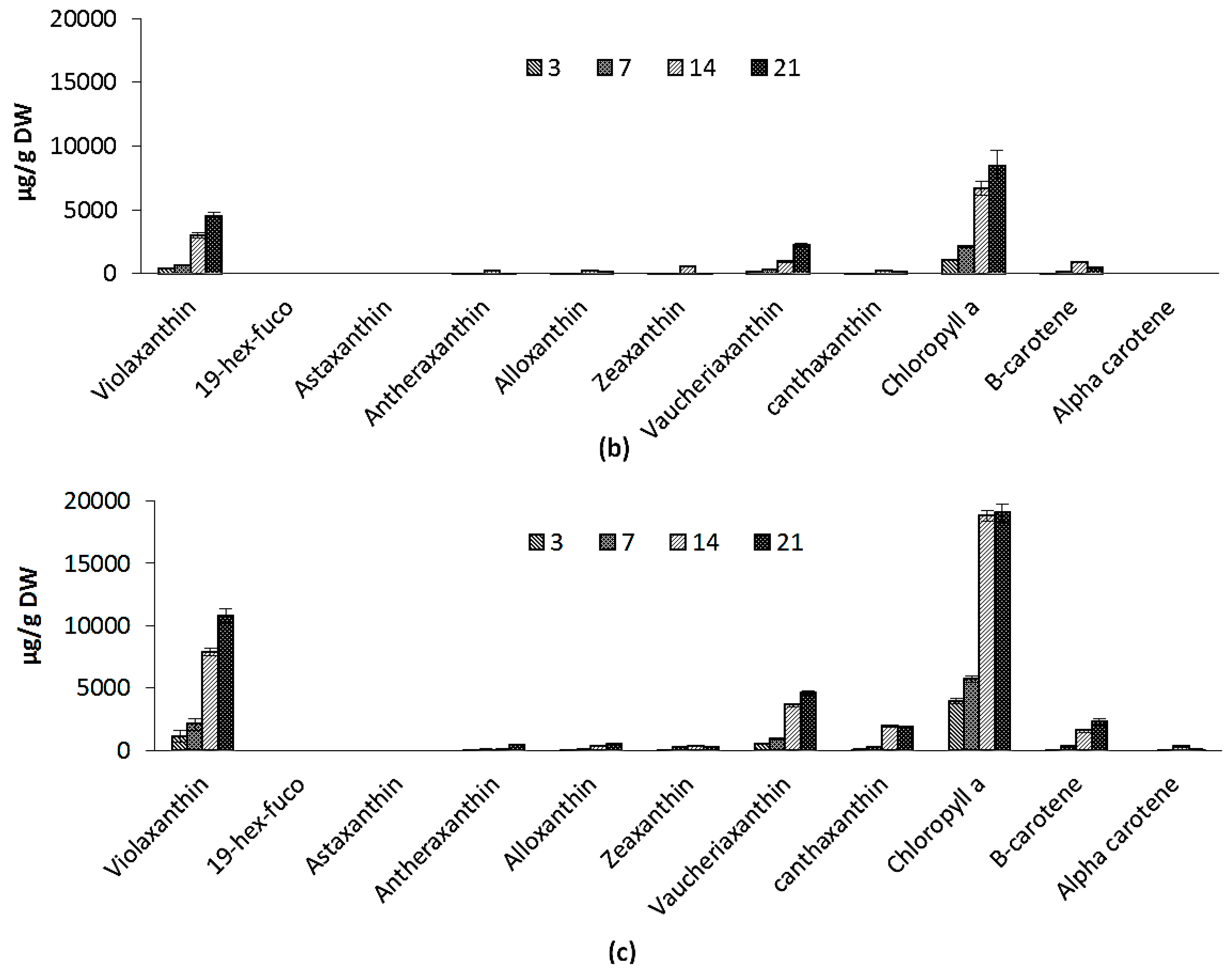
2.2.4. Pigments
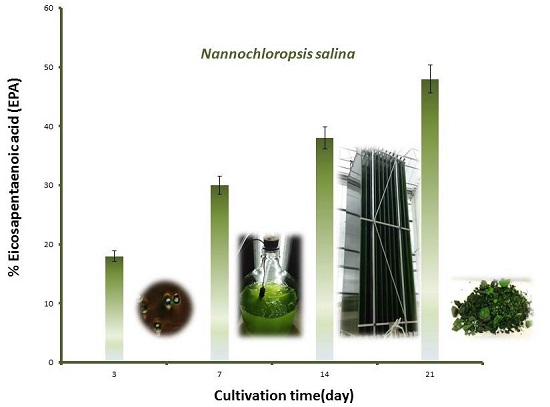
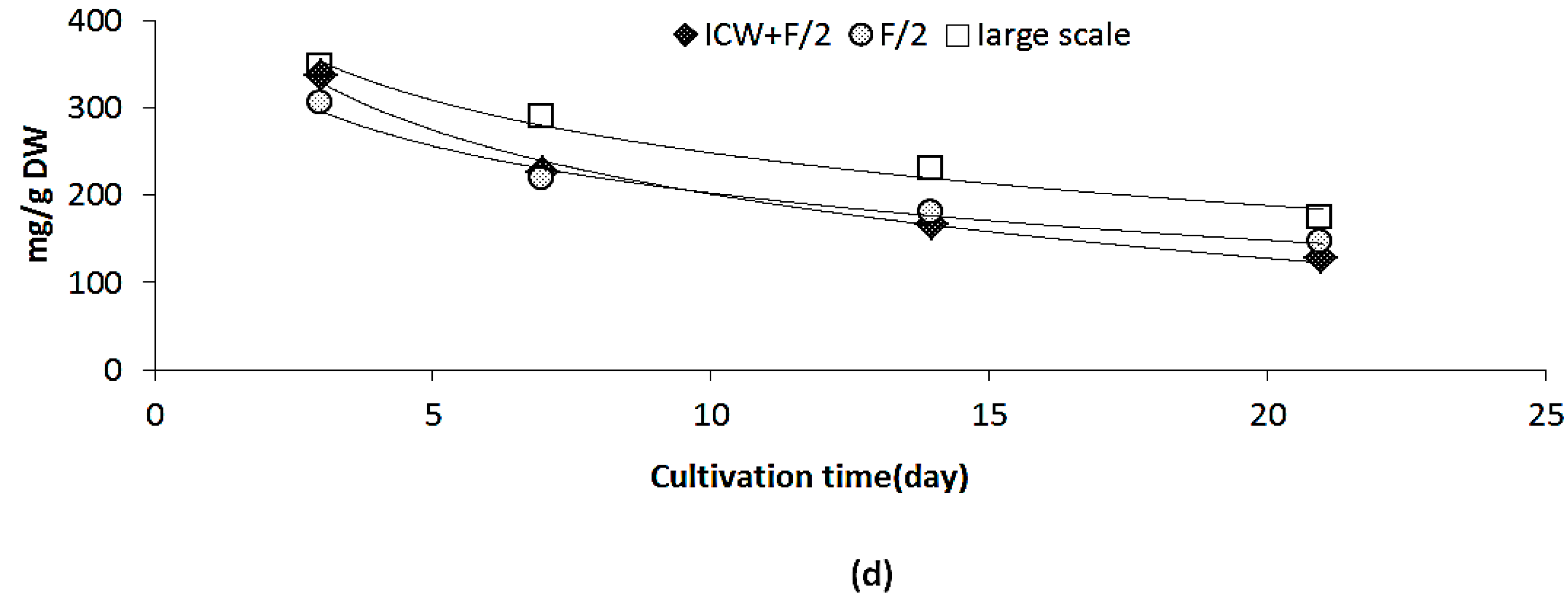
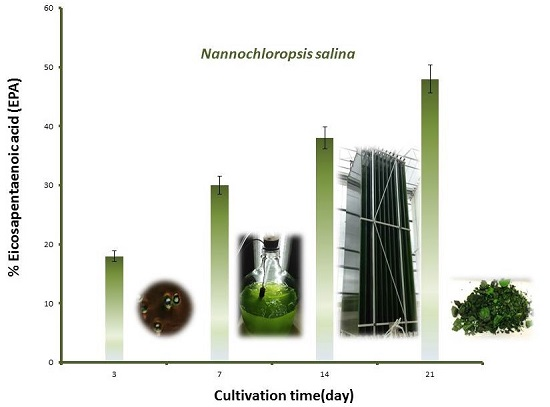
2.3. Changes in Growth and Biomass Composition of Large Scale Cultivated Nannochloropsis salina
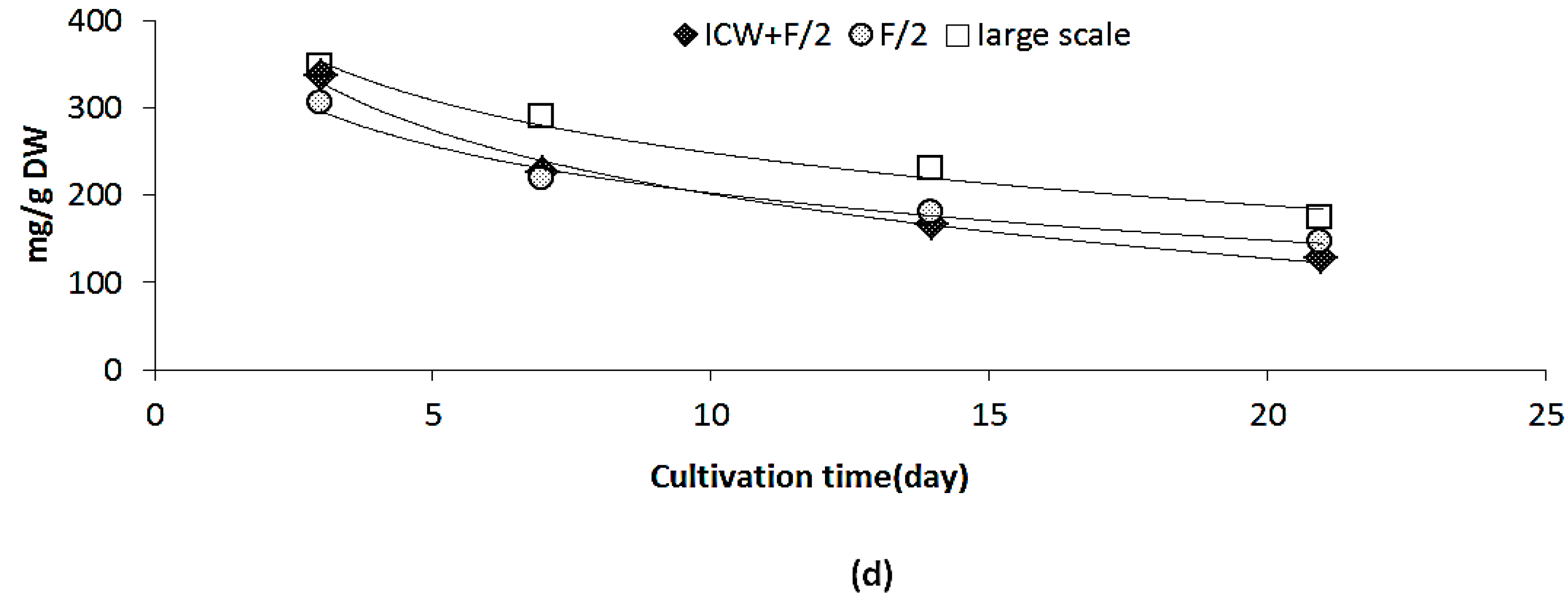
2.3.1. Growth Rate and Biomass Productivity
2.3.2. Biomass Chemical Composition
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Growth Media
3.3. Laboratory Scale Cultivation of Microalgae
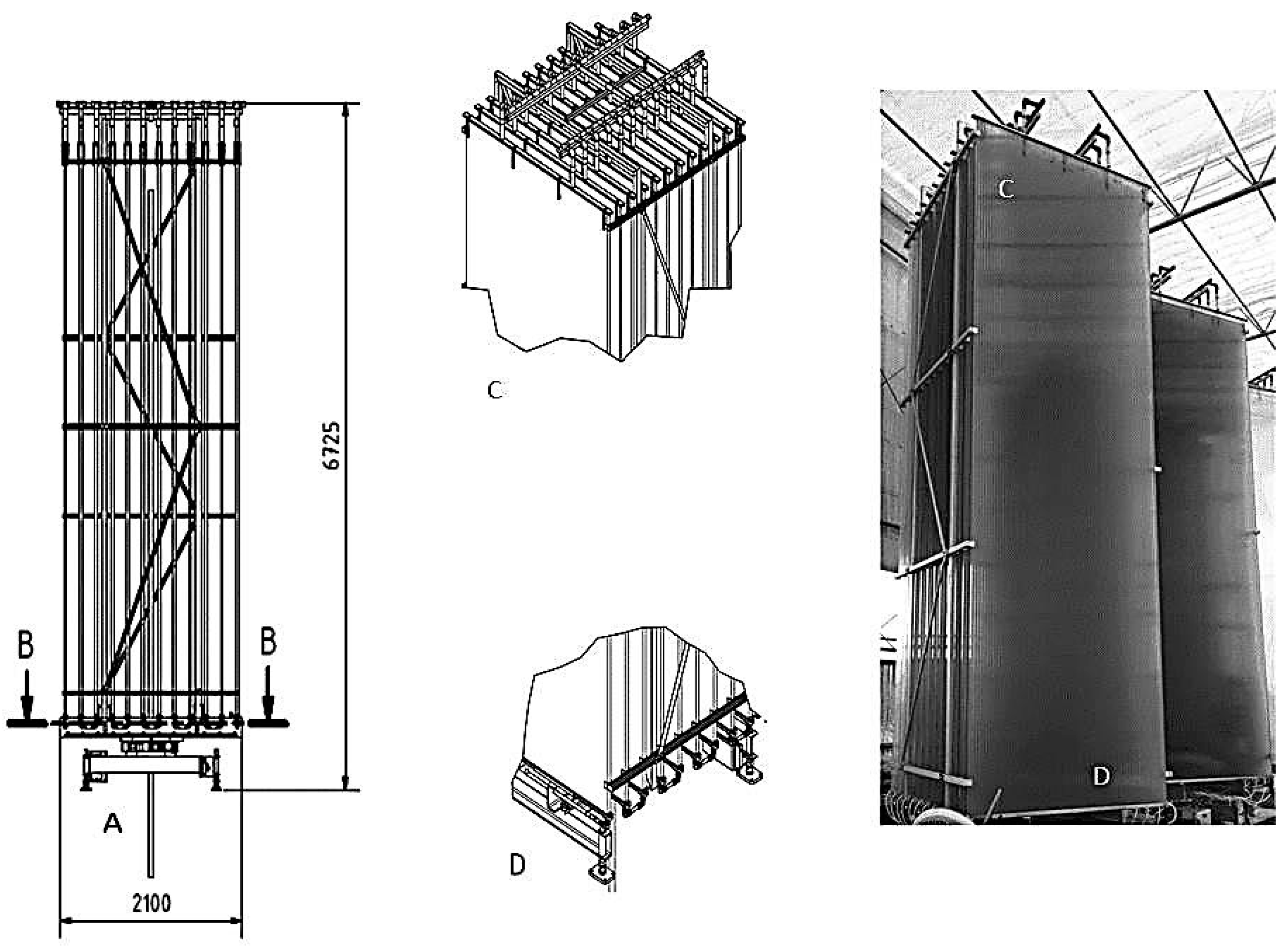
3.4. Large-Scale Cultivation of Microalgae
3.5. Analytical Methods
3.5.1. Growth Rate
3.5.2. Protein and Amino Acids
3.5.3. Lipids, Fatty Acid and Tocopherols
3.5.4. Pigments
3.5.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| EPA | Eicosa pentanoic acid |
| ICW | Industrial process water |
| BHT | Butylated hydroxyl toluene |
| FPPBR | Flat panel photo bioreactor |
References
- Mostafa, S. Microalgal biotechnology: Prospects and applications. In Agricultural and Biological Sciences; Dhal, N.K., Sahu, S.C., Eds.; InTech Open publication: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef]
- Benemann, J.R. CO2 mitigation with microalgae systems. Energy Convers. Manag. 1997, 38, 475–479. [Google Scholar]
- Se-Kwon, K. Chapter 1: Marine microalgae biotechnology present trends and future advances. In Handbook of Marine Microalgae; Elsevier Publications: Amsterdam, The Netherlands, 2015; p. 27. [Google Scholar]
- Becker, E.W. Microalgae for aquaculture: Nutritional Aspects. In Handbook of Microalgal Culture: Applied Phycology Biotechnology; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 671–691. [Google Scholar]
- Mourente, G.; Lubian, L.M.; Odriozola, J.M. Total fatty acid composition as a taxonomic index of some marine microalgae used as food in marine aquaculture. Hydrobiologia 1990, 203, 147–154. [Google Scholar]
- Roy, S.S.; Pal, R. Microalgae in Aquaculture: A Review with Special References to Nutritional Value and Fish Dietetics. Proc. Zool. Soc. 2015, 68, 1–8. [Google Scholar]
- Muller-Feuga, A. Microalgae for aquaculture: The current global situation and future trends. In Handbook of Microalgal Culture: Applied Phycology Biotechnology; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 613–627. [Google Scholar]
- Muller-Feuga, A. The role of microalgae in aquaculture: Situation and trends. J. Appl. Phycol. 2000, 12, 527–534. [Google Scholar]
- Fawley, K.P.; Fawley, M.W. Observations on the Diversity and Ecology of Freshwater Nannochloropsis (Eustigmatophyceae), with Descriptions of New Taxa. Protist 2007, 158, 325–336. [Google Scholar] [PubMed]
- Andersen, R.A.; Brett, R.W.; Potter, D.; Sexton, J.P. Phylogeny of the Eustigmatophyceae Based upon 18S rDNA, with Emphasis on Nannochloropsis. Protist 1998, 149, 61–74. [Google Scholar] [PubMed]
- Hoffmann, M.; Marxen, K.; Schulz, R.; Vanselow, K.H. TFA and EPA productivities of Nannochloropsis salina influenced by temperature and nitrate stimuli in turbidostatic controlled experiments. Mar. Drugs 2010, 8, 2526–2545. [Google Scholar] [PubMed]
- Ma, Y.; Wang, Z.; Yu, C.; Yin, Y.; Zhou, G. Evaluation of the potential of 9 Nannochloropsis strains for biodiesel production. Bioresour. Technol. 2014, 167, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Gao, K. Response of growth and fatty acid compositions of Nannochloropsis sp. to environmental factors under elevated CO2 concentration. Biotechnol. Lett. 2006, 28, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Sforza, E.; Bertucco, A.; Morosinotto, T.; Giacometti, G.M. Photo-bioreactors for microalgal growth and oil production with Nannochloropsis salina: From lab-scale experiments to large-scale design. Chem. Eng. Res. Des. 2012, 90, 1151–1158. [Google Scholar] [CrossRef]
- Tibaldi, E.; Zittelli, G.C.; Parisi, G.; Bruno, M.; Giorgi, G.; Tulli, F.; Venturini, S.; Tredici, M.R.; Poli, B.M. Growth performance and quality traits of European sea bass (D. labrax) fed diets including increasing levels of freeze-dried Isochrysis sp. (T-ISO) biomass as a source of protein and n-3 long chain PUFA in partial substitution of fish derivatives. Aquaculture 2015, 440, 60–68. [Google Scholar] [CrossRef]
- Zitteli, G.C.; Lavista, F.; Bastianini, A.; Rodolfi, L.; Vincenzini, M.; Tredici, M.R. Production of eicosapentaenoic acid by Nannochloropsis sp. cultures in outdoor tubular photo-bioreactors. J. Biotechnol. 1999, 70, 299–312. [Google Scholar] [CrossRef]
- Zitteli, G.C.; Pastorelli, R.; Tredici, M.R. A Modular Flat Panel Photo-bioreactor (MFPP) for indoor mass cultivation of Nannochloropsis sp. under artificial illumination. J. Appl. Phycol. 2000, 12, 521–526. [Google Scholar] [CrossRef]
- Olofsson, M.; Lamela, T.; Nilsson, E.; Bergé, J.P.; del Pino, V.; Uronen, P.; Legrand, C. Seasonal variation of lipids and fatty acids of the microalgae Nannochloropsis oculata grown in outdoor large-scale photo-bioreactors. Energies 2012, 5, 1577–1592. [Google Scholar] [CrossRef]
- Zou, N.; Richmond, A. Effect of light-path length in outdoor fiat plate reactors on output rate of cell mass and of EPA in Nannochloropsis sp. Prog. Ind. Microbiol. 1999, 35, 351–356. [Google Scholar]
- Olofsson, M.; Lamela, T.; Nilsson, E.; Bergé, J.P.; del Pino, V.; Uronen, P.; Legrand, C. Combined effects of nitrogen concentration and seasonal changes on the production of lipids in Nannochloropsis oculata. Mar. Drugs 2014, 12, 1891–1910. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Rodríguez, J.; González-Céspedes, A.M.; Cerón-García, M.C.; Fernández-Sevilla, J.M.; Acién-Fernández, F.G.; Molina-Grima, E. A quantitative study of eicosapentaenoic acid (EPA) production by Nannochloropsis gaditana for aquaculture as a function of dilution rate, temperature and average irradiance. Appl. Microbiol. Biotechnol. 2014, 98, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- San Pedro, A.; González-López, C.V.; Acién, F.G.; Molina-Grima, E. Outdoor pilot production of Nannochloropsis gaditana: Influence of culture parameters and lipid production rates in raceway ponds. Algal Res. 2015, 8, 205–213. [Google Scholar] [CrossRef]
- De Sousa, L.L.; da Hora, D.S.; Sales, E.A.; Perelo, L.W. Cultivation of Nannochloropsis sp. in brackish groundwater supplemented with municipal wastewater as a nutrient source. Braz. Arch. Biol. Technol. 2014, 57, 171–177. [Google Scholar] [CrossRef]
- Cai, T.; Park, S.Y.; Racharaks, R.; Li, Y. Cultivation of Nannochloropsis salina using anaerobic digestion effluent as a nutrient source for biofuel production. Appl. Energy 2013, 108, 486–492. [Google Scholar] [CrossRef]
- Dong, B.; Ho, N.; Ogden, K.L.; Arnold, R.G. Cultivation of Nannochloropsis salina in municipal wastewater or digester centrate. Ecotoxicol. Environ. Saf. 2014, 103, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Emdadi, D.; Berland, B. Variation in lipid class composition during batch growth of Nannochloropsis salina and Pavlova lutheri. Mar. Chem. 1989, 26, 215–225. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Hii, Y.S.; Soo, C.L.; Chuah, T.S.; Mohd-Azmi, A.; Abol-Munafi, A. Interactive effect of ammonia and nitrate on the nitrogen uptake by Nannochloropsis sp. J. Sustain. Sci. Manag. 2011, 6, 60–68. [Google Scholar]
- Hii, Y.S.; Soo, C.L.; Chuah, T.S.; Mohd-Azmi, A.; Abol-Munafi, A. The effects of temperature on the growth of and ammonia uptake by marine microalgae. Bot. Stud. 2012, 53, 125–133. [Google Scholar]
- Hecky, R.E.; Kilham, P. Nutrient limitation of phytoplankton in freshwater and marine environments: A review of recent evidence on the effects of enrichment. Limnol. Oceanogr. 1988, 33, 796–822. [Google Scholar] [CrossRef]
- Collos, Y.; Harrison, P.J. Acclimation and toxicity of high ammonium concentrations to unicellular algae. Mar. Pollut. Bull. 2014, 80, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q. Environmental effects on cell composition. In Handbook of Microalgal Culture: Applied Phycology Biotechnology; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 114–122. [Google Scholar]
- Chen, Y.; Tang, X.; Kapoore, R.V.; Xu, C.; Vaidyanathan, S. Influence of nutrient status on the accumulation of biomass and lipid in Nannochloropsis salina and Dunaliella salina. Energy Convers. Manag. 2015, 106, 62–72. [Google Scholar] [CrossRef]
- Safafar, H.; van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, phenolic compounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durmaz, Y. Vitamin E (α-tocopherol) production by the marine microalgae Nannochloropsis oculata (Eustigmatophyceae) in nitrogen limitation. Aquaculture 2007, 272, 717–722. [Google Scholar] [CrossRef]
- Solovchenko, A.; Khozin-Goldberg, I.; Recht, L.; Boussiba, S. Stress-Induced Changes in Optical Properties, Pigment and Fatty Acid Content of Nannochloropsis sp.: Implications for Non-destructive Assay of Total Fatty Acids. Mar. Biotechnol. 2011, 13, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Hoshida, H.; Ohira, T.; Minematsu, A.; Akada, R.; Nishizawa, Y. Accumulation of eicosapentaenoic acid in Nannochloropsis sp. in response to elevated CO2 concentrations. J. Appl. Phycol. 2005, 17, 29–34. [Google Scholar] [CrossRef]
- Brown, M.R. The amino-acid and sugar composition of 16 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- Owens, T.G.; Gallager, J.C.; Alberte, R.S. Photosynthetic light harvesting function of viloaxanthin in Nannochloropsis spp. (Eustigmatophyceae). J. Phycol. 1987, 23, 79–85. [Google Scholar] [CrossRef]
- Mulders, K.J.M.; Lamers, P.P.; Martens, D.E.; Wijffels, R.H. Phototrophic Pigment Production with Microalgae: Biological Constraints and Opportunities. J. Phycol. 2014, 50, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Rebolloso-Fuentes, M.M.; Navarro-Pérez, A.; García-Camacho, F.; Ramos-Miras, J.J.; Guil-Guerrero, J.L. Biomass nutrient profiles of the microalga Nannochloropsis. J. Agric. Food Chem. 2001, 49, 2966–2972. [Google Scholar] [CrossRef] [PubMed]
- Dragsted, J.; Furbo, S. Solar Radiation and Thermal Performance of Solar Collectors for Denmark. Available online: http://orbit.dtu.dk/files/51295325/Solar_radiation_and_thermal_performance_of_solar_collectors_for_Denmark.pdf (accessed on 28 July 2016).
- Chen, Y.; Vaidyanathan, S. Simultaneous assay of pigments, carbohydrates, proteins and lipids in microalgae. Anal. Chim. Acta 2013, 776, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Firestone, D. Official Methods and Recommended Practices of the AOCS, 5th ed.; American Oil Chemist’s Society: Urbana, IL, USA, 2009. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
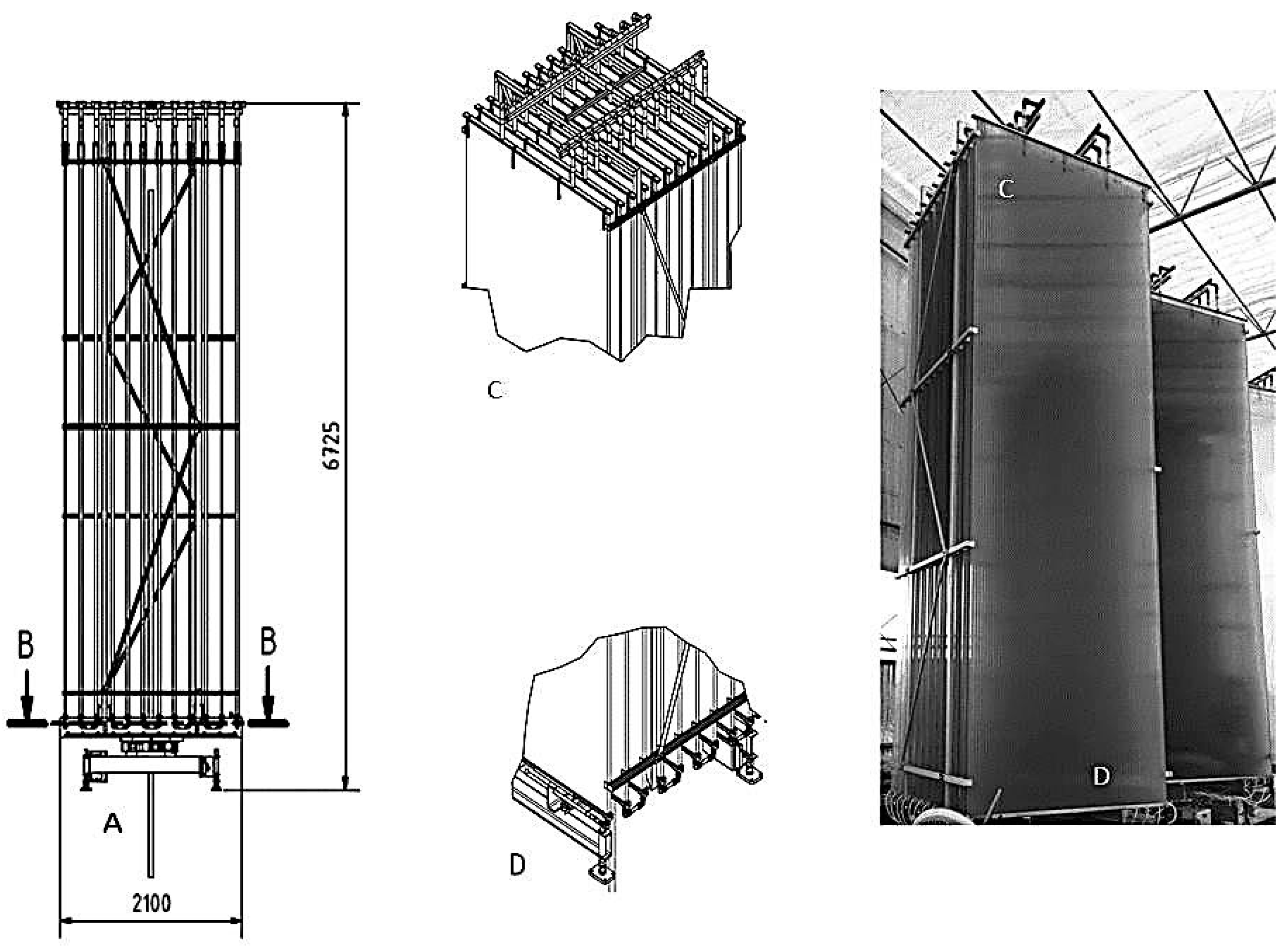{kind=link}
| Item | Unit | Amount |
|---|---|---|
| pH | - | 8.1 |
| Suspended solids | mg/L | 20 |
| Total N | mg/L | 190 |
| Ammonia + ammonium-N | mg/L | 150 |
| Nitrite + nitrate | mg/L | <0.1 |
| Total P | mg/L | 11 |
| Sulphate | mg/L | 3.6 |
| Total Alkalinity | mmol/L | 62.5 |
| EDTA | mg/L | <0.5 |
| Sodium(Na) | mg/L | 1500 |
| Copper (Cu) | µg/L | 3.4 |
| Iron (Fe) | mg/l | 0.23 |
| Growth Media * | Total N | Total P | ||
|---|---|---|---|---|
| F/2 | 75 | - | 75 | 5.0 |
| 20% ICW + 80% F/2 | 60 | 30 | 90 | 6.2 |
| 30% ICW + 70% F/2 | 52 | 45 | 97 | 6.8 |
| 40% ICW + 60% F/2 | 45 | 60 | 105 | 7.7 |
| Cultivation Time (Day) | 3 | 7 | 14 | 21 | ||||
|---|---|---|---|---|---|---|---|---|
| Fatty Acid | F/2 | ICW + F/2 | F/2 | ICW + F/2 | F/2 | ICW + F/2 | F/2 | ICW + F/2 |
| 14:0 | 1.45 ± 0.44 | 3.22 ± 0.50 | 3.52 ± 0.12 | 0.51 ± 0.05 | 1.40 ± 0.13 | 0.56 ± 0.30 | 2.05 ± 0.50 | 3.53 ± 0.13 |
| 15:0 | 0.04 ± 0.00 | nd | nd | nd | 0.54 ± 0.01 | 1.10 ± 0.40 | 0.30 ± 0.09 | 1.46 ± 0.03 |
| 16:0 | 48.8 ± 0.59 a | 43.3 ± 0.98 b | 32.9 ± 0.53 c | 31.9 ± 0.77 c | 23.7 ± 0.40 d | 20.6 ± 0.64 de | 21.3 ± 0.30 de | 19.1 ± 0.40 e |
| 16:1 (n-7) | 30.3 ± 0.40 a | 30.9 ± 0.10 a | 24.0 ± 0.09 b | 20.4 ± 0.02 c | 25.1 ± 0.51 b | 23.0 ± 0.02 b | 31.9 ± 0.66 a | 17.7 ± 0.51 c |
| 16:2 (n-4) | 0.18 ± 0.01 | 0.23 ± 0.02 | 1.24 ± 0.00 | 0.65 ± 0.00 | 0.33 ± 0.01 | 0.76 ± 0.07 | 0.64 ± 0.02 | 0.38 ± 0.01 |
| 16:3 (n-4) | 0.11 ± 0.03 | 3.57 ± 0.09 | 0.61 ± 0.02 | 0.56 ± 0.03 | 0.75 ± 0.04 | 0.63 ± 0.08 | 0.24 ± 0.05 | 1.01 ± 0.04 |
| 16:4 (n-1) | 0.36 ± 0.22 | 3.35 ± 0.08 | 0.45 ± 0.04 | 0.14 ± 0.19 | 0.44 ± 0.01 | 0.28 ± 0.10 | 0.43 ± 0.03 | 0.66 ± 0.06 |
| 18:0 | 2.35 ± 0.20 | 1.69 ± 0.19 | 5.32 ± 0.09 | 1.82 ± 0.06 | 2.20 ± 0.01 | 2.10 ± 0.06 | 0.51 ± 0.10 | 1.86 ± 0.07 |
| 18:1 (n-9) | 7.03 ± 0.23 | 3.31 ± 0.06 | 9.43 ± 0.08 | 5.61 ± 0.06 | 9.06 ± 0.20 | 6.53 ± 0.06 | 4.22 ± 0.06 | 4.38 ± 0.20 |
| 18:1 (n-7) | 0.75 ± 0.20 | 0.00 ± 0.06 | 0.45 ± 0.08 | 6.31 ± 0.60 | 7.52 ± 0.21 | 7.50 ± 0.45 | 0.57 ± 0.06 | 3.32 ± 0.21 |
| 18:2 (n-6) | 1.13 ± 0.01 | 0.74 ± 0.06 | 1.45 ± 0.03 | 1.35 ± 0.06 | 2.23 ± 0.08 | 1.67 ± 0.07 | 1.63 ± 0.03 | 1.46 ± 0.09 |
| 18:3(n-3) | 0.64 ± 0.10 | 0.50 ± 0.01 | 0.49 ± 0.04 | 0.46 ± 0.03 | 0.04 ± 0.02 | nd | 0.30 ± 0.01 | nd |
| 20:1(n-7) | 0.10 ± 0.00 | nd | 1.20 ± 0.12 | nd | 0.10 ± 0.01 | nd | 0.10 ± 0.01 | nd |
| 20:4 (n-6) | 1.21 ± 0.20 | 0.73 ± 0.06 | 1.92 ± 0.05 | 4.82 ± 0.08 | 3.82 ± 0.07 | 5.63 ± 0.08 | 2.33 ± 0.06 | 4.91 ± 0.07 |
| 20:3 (n-3) | 0.22 ± 0.08 | 0.33 ± 0.08 | 1.57 ± 0.04 | 0.26 ± 0.02 | nd | 0.31 ± 0.02 | 0.17 ± 0.08 | 0.93 ± 0.02 |
| 20:4 (n-3) | 0.10 ± 0.00 | 0.33 ± 0.02 | 0.43 ± 0.03 | nd | 0.31 ± 0.08 | nd | 0.10 ± 0.00 | nd |
| 20:5 (n-3) | 4.95 ± 0.78 a | 7.47 ± 0.23 ab | 10.9 ± 0.11 b | 22.2 ± 0.21 c | 19.9 ± 0.42 c | 26.2 ± 0.25 d | 32.0 ± 0.82 e | 37.1 ± 0.77 f |
| 22:5 (n-3) | 0.66 ± 0.05 | nd | 0.40 ± 0.01 | 0.26 ± 0.04 | 1.35 ± 0.05 | 0.14 ± 0.03 | 0.66 ± 0.03 | 0.27 ± 0.05 |
| 22:6 (n-3) | 0.46 ± 0.09 | 0.15 ± 0.04 | 0.24 ± 0.02 | 1.04 ± 0.40 | nd | 0.69 ± 0.40 | 0.19 ± 0.02 | 0.16 ± 0.05 |
| ∑ SAFA | 51.6 ± 3.25 a | 48.3 ± 2.33 a | 41.8 ± 0.63 b | 34.2 ± 1.25 b | 27.9 ± 0.51 c | 24.4 ± 1.44 c | 24.2 ± 1.63 c | 25.9 ± 0.62 c |
| ∑ n-3 | 6.92 ± 2.41 a | 8.77 ± 0.45 a | 13.6 ± 0.23 b | 24.3 ± 0.70 c | 21.66 ± 0.66 c | 27.4 ± 0.74 d | 33.37 ± 1.0 e | 38.6 ± 1.00 f |
| Cultivation Time (Day) | 3 | 7 | 14 | 21 |
|---|---|---|---|---|
| Fatty Acid | ||||
| 14:0 | 5.28 ± 0.57 | 4.20 ± 0.50 | 3.58 ± 0.37 | 3.37 ± 0.37 |
| 15:0 | 0.67 ± 0.44 | 0.38 ± 0.04 | 0.26 ± 0.04 | 0.24 ± 0.04 |
| 16:0 | 24.9 ± 2.28 a | 19.9 ± 0.59 b | 18.5 ± 0.19 b | 17.4 ± 0.24 b |
| 16:1 (n-7) | 26.5 ± 2.59 a | 28.7 ± 0.30 a | 28.1 ± 0.44 a | 26.4 ± 0.19 a |
| 16:2 (n-4) | 0.18 ± 0.03 | 0.23 ± 0.02 | 0.10 ± 0.02 | 0.10 ± 0.00 |
| 16:3 (n-4) | 0.20 ± 0.01 | 0.45 ± 0.02 | 0.22 ± 0.03 | 0.10 ± 0.02 |
| 16:4 (n-1) | 0.44 ± 0.03 | 1.01 ± 0.02 | 0.50 ± 0.02 | 0.21 ± 0.02 |
| 18:0 | 1.91 ± 0.22 | 0.37 ± 0.08 | 0.26 ± 0.03 | 0.47 ± 0.02 |
| 18:1 (n-9) | 6.06 ± 0.76 | 3.93 ± 0.19 | 1.87 ± 0.10 | 1.76 ± 0.21 |
| 18:1 (n-11) | 2.62 ± 0.20 | 1.31 ± 0.06 | 0.94 ± 0.03 | 0.24 ± 0.00 |
| 18:2 (n-6) | 2.39 ± 0.23 | 1.30 ± 0.06 | 0.16 ± 0.01 | 0.88 ± 0.03 |
| 18:3 (n-3) | 0.10 ± 0.01 | nd | nd | nd |
| 20:1 (n-9) | 0.10 ± 0.02 | 0.11 ± 0.01 | nd | nd |
| 20:1(n-7) | 0.24 ± 0.02 | nd | 0.14 ± 0.01 | nd |
| 20:4 (n-6) | 0.05 ± 0.02 | nd | nd | 0.13 ± 0.01 |
| 20:3 (n-3) | 1.10 ± 0.03 | 6.45 ± 0.06 | 3.38 ± 0.05 | 2.75 ± 0.03 |
| 20:4 (n-3) | 0.61 ± 0.01 | 0.10 ± 0.08 | 1.02 ± 0.34 | 0.91 ± 0.01 |
| 20:5 (n-3) | 23.9 ± 1.78 a | 30.1 ± 1.21 b | 39.3 ± 1.10 c | 44.2 ± 2.30 d |
| 22:1 (n-11) | nd | 0.16 ± 0.03 | 0.10 ± 0.03 | 0.12 ± 0.03 |
| 21:5 (n-3) | 1.82 ± 0.39 | 0.81 ± 0.35 | 1.03 ± 0.01 | 0.09 ± 0.01 |
| 22:5 (n-3) | 0.32 ± 0.05 | 0.23 ± 0.03 | 0.07 ± 0.01 | 0.06 ± 0.02 |
| 22:6 (n-3) | nd | 0.12 ± 0.20 | 0.10 ± 0.04 | 0.10 ± 0.01 |
| ∑ SAFA | 32.0 ± 3.10 a | 24.5 ± 1.21 b | 22.3 ± 0.72 b | 21.2 ± 0.74 b |
| ∑ n-3 | 27.8 ± 2.30 a | 37.7 ± 2.00 b | 44.9 ± 1.52 bc | 48.1 ± 2.43 c |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Safafar, H.; Hass, M.Z.; Møller, P.; Holdt, S.L.; Jacobsen, C. High-EPA Biomass from Nannochloropsis salina Cultivated in a Flat-Panel Photo-Bioreactor on a Process Water-Enriched Growth Medium. Mar. Drugs 2016, 14, 144. https://doi.org/10.3390/md14080144
Safafar H, Hass MZ, Møller P, Holdt SL, Jacobsen C. High-EPA Biomass from Nannochloropsis salina Cultivated in a Flat-Panel Photo-Bioreactor on a Process Water-Enriched Growth Medium. Marine Drugs. 2016; 14(8):144. https://doi.org/10.3390/md14080144
Chicago/Turabian StyleSafafar, Hamed, Michael Z. Hass, Per Møller, Susan L. Holdt, and Charlotte Jacobsen. 2016. "High-EPA Biomass from Nannochloropsis salina Cultivated in a Flat-Panel Photo-Bioreactor on a Process Water-Enriched Growth Medium" Marine Drugs 14, no. 8: 144. https://doi.org/10.3390/md14080144