
Micrometam C Protects against Oxidative Stress in Inflammation Models in Zebrafish and RAW264.7 Macrophages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
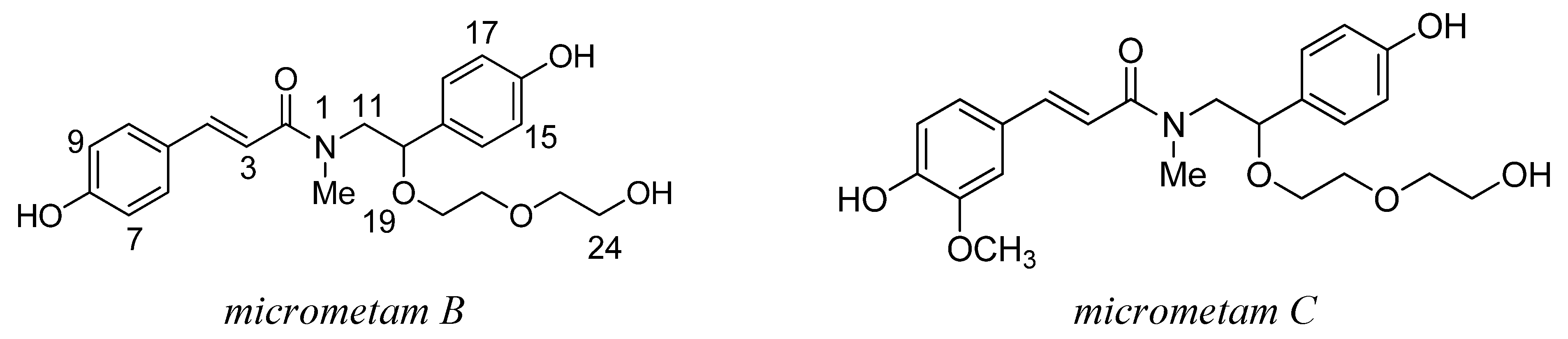
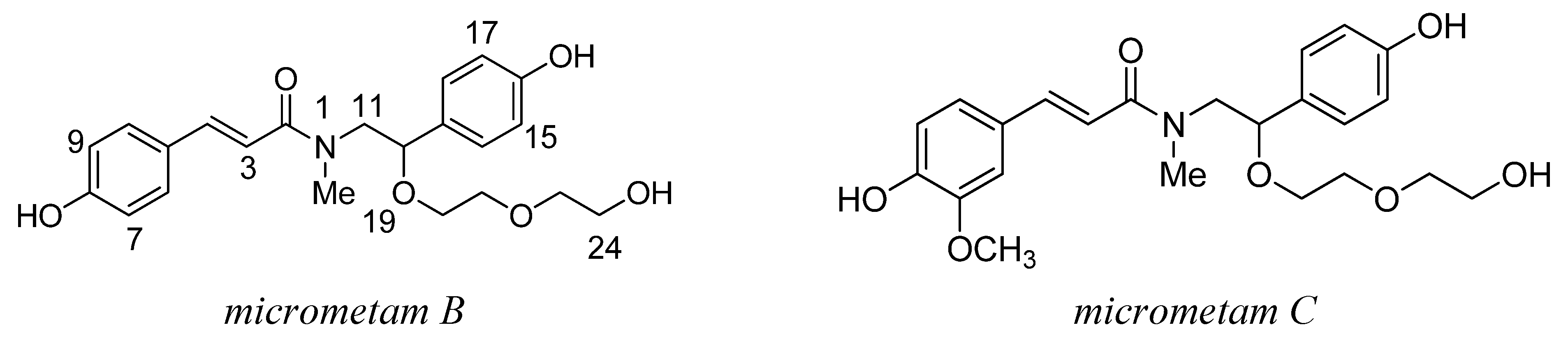
2.1. The Isolation and Preparation of Micrometam B and Micrometam C
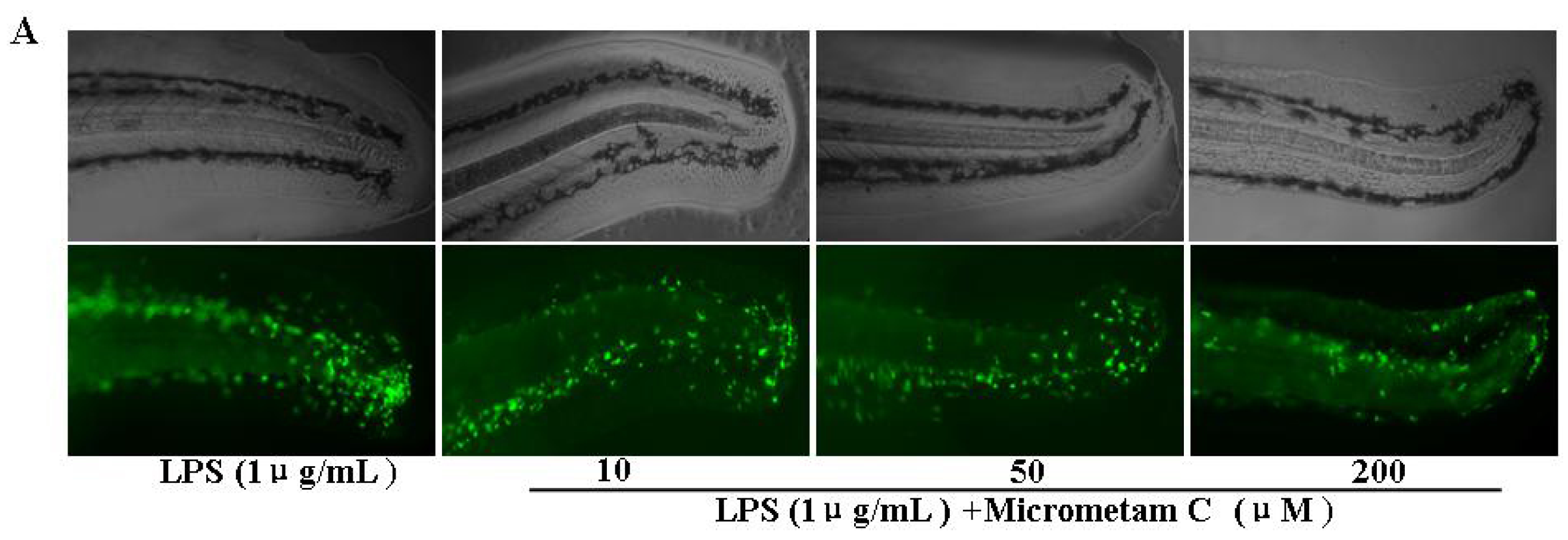
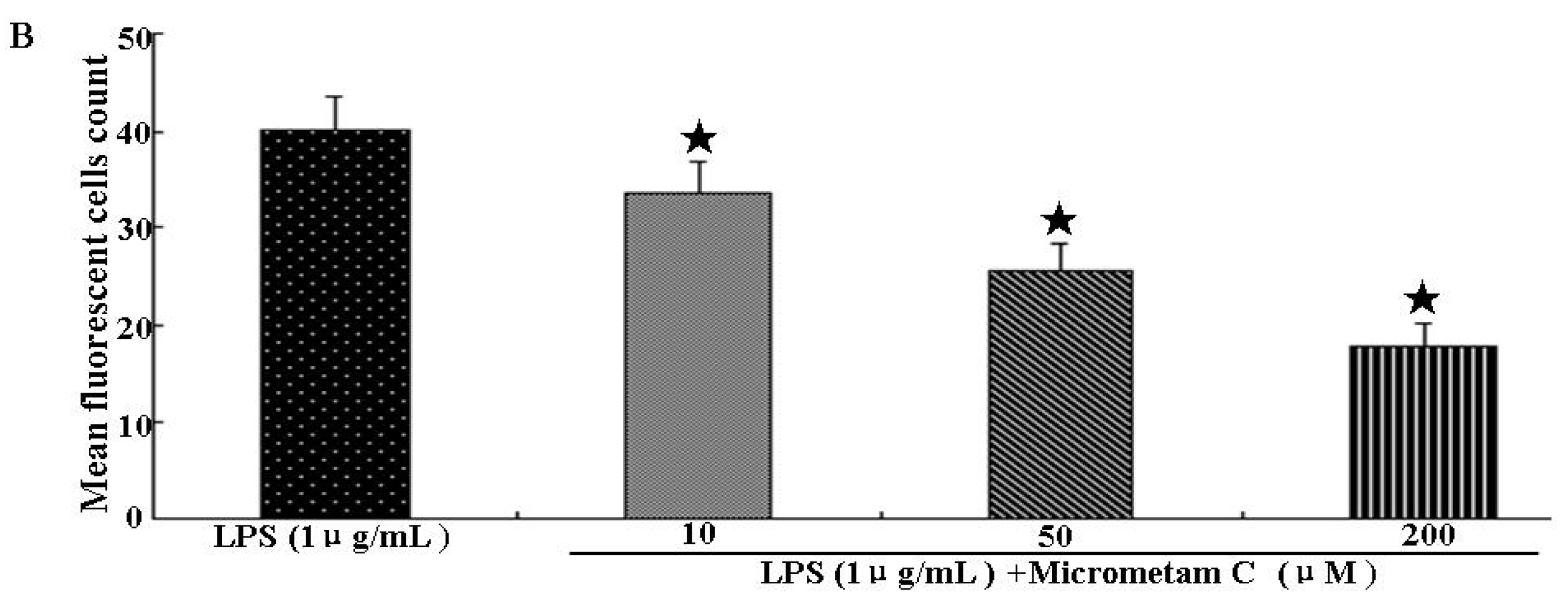
2.2. Effect of Micrometam C in Tail Cutting-Induced Inflammation in Zebrafish Model
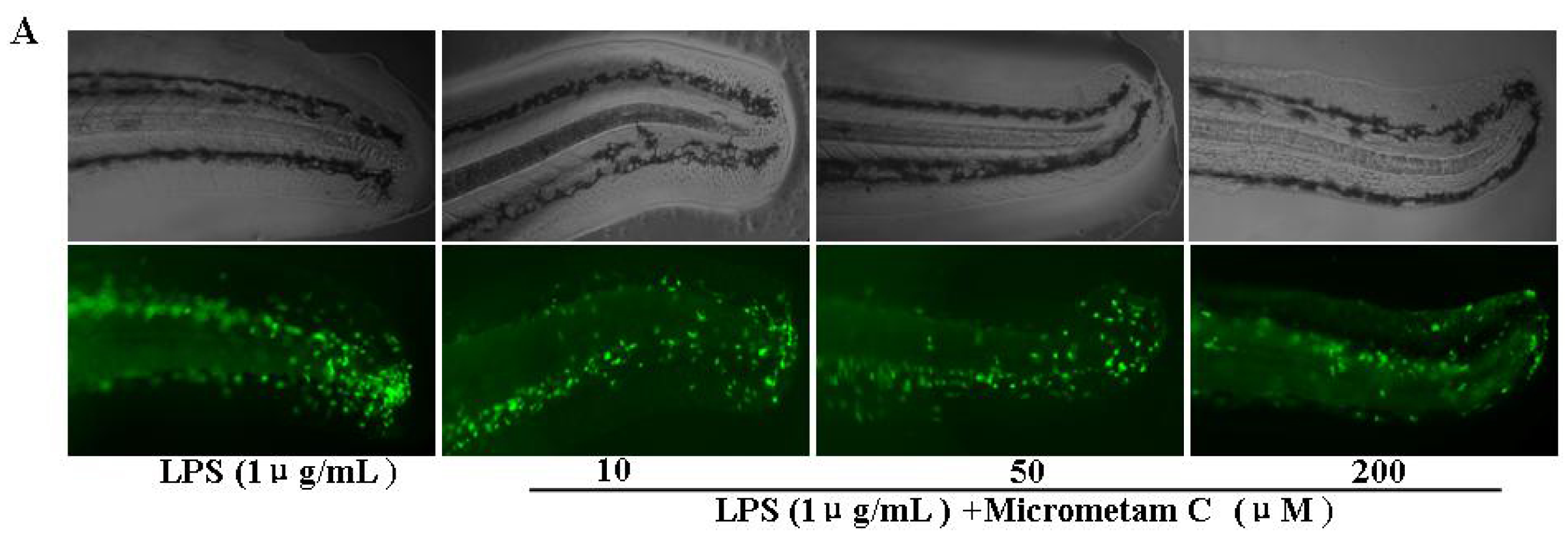
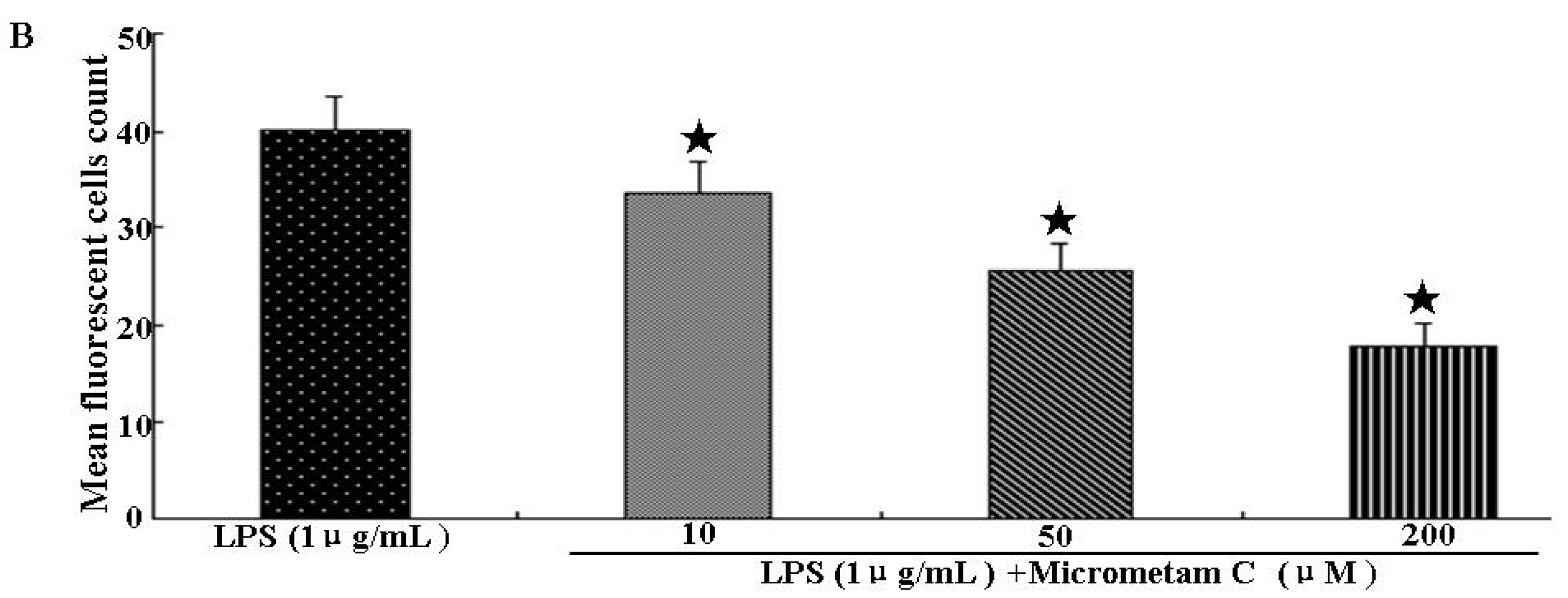
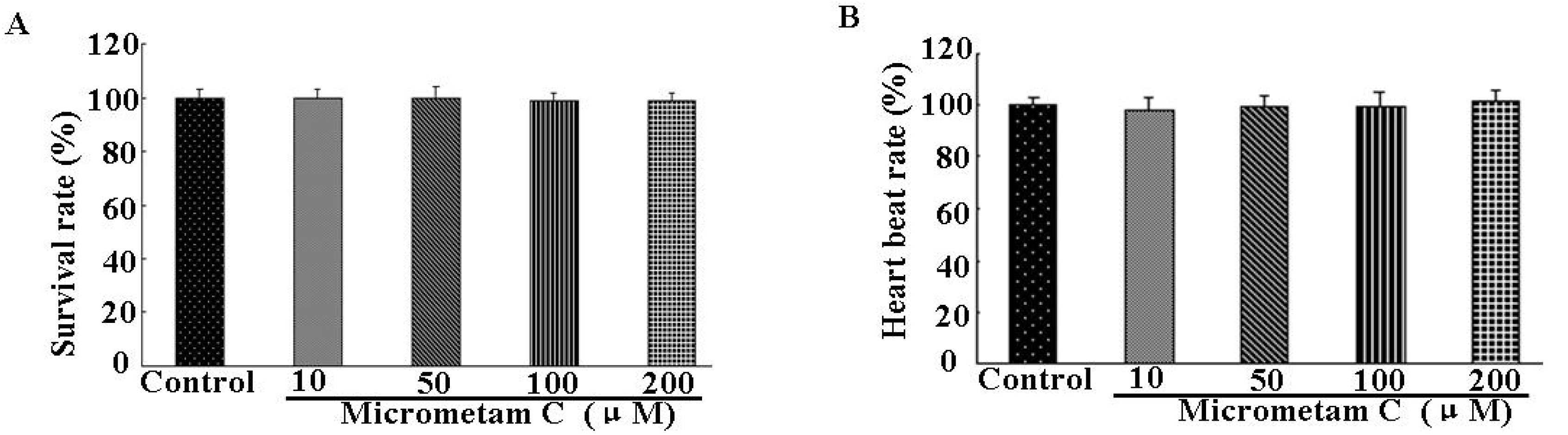
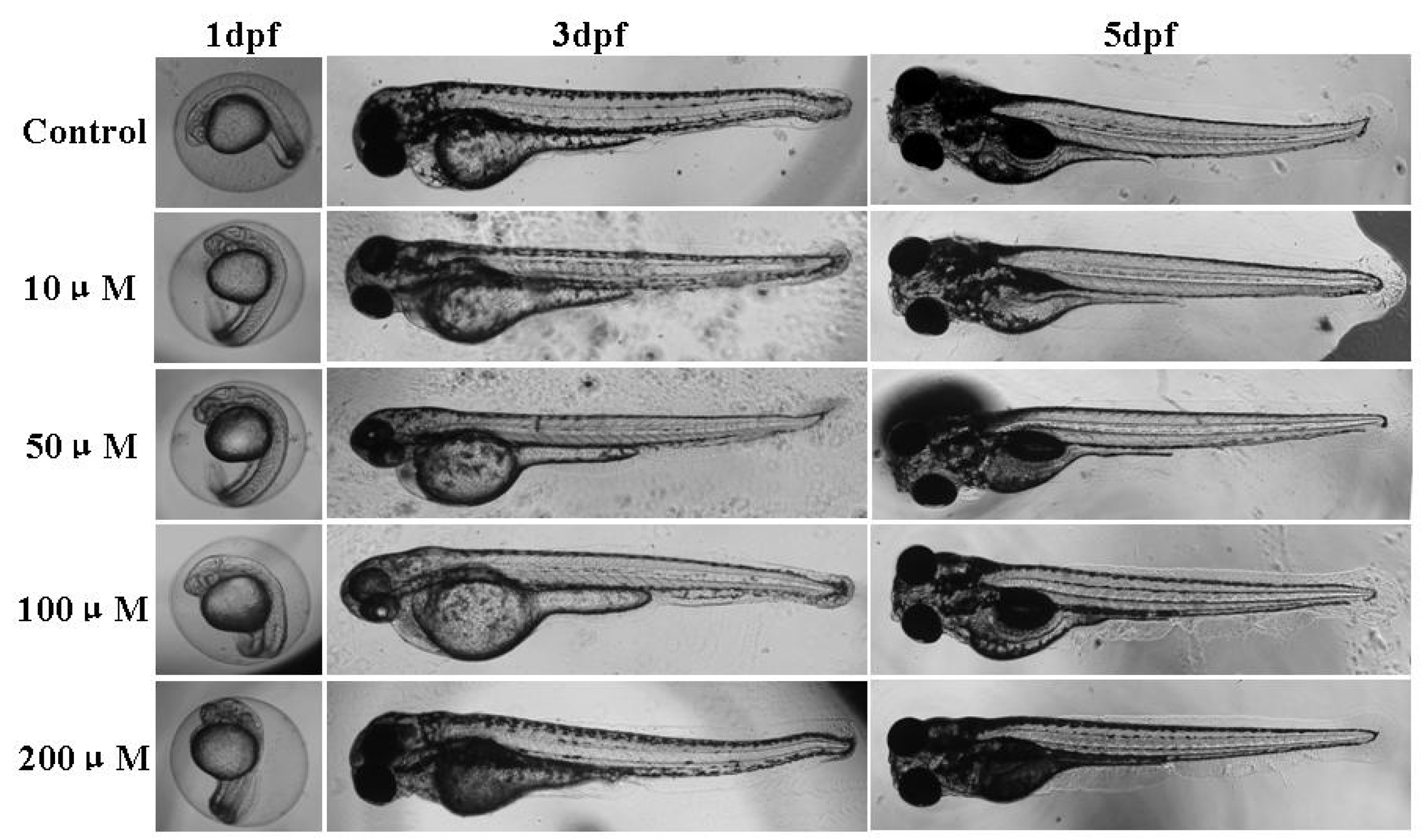
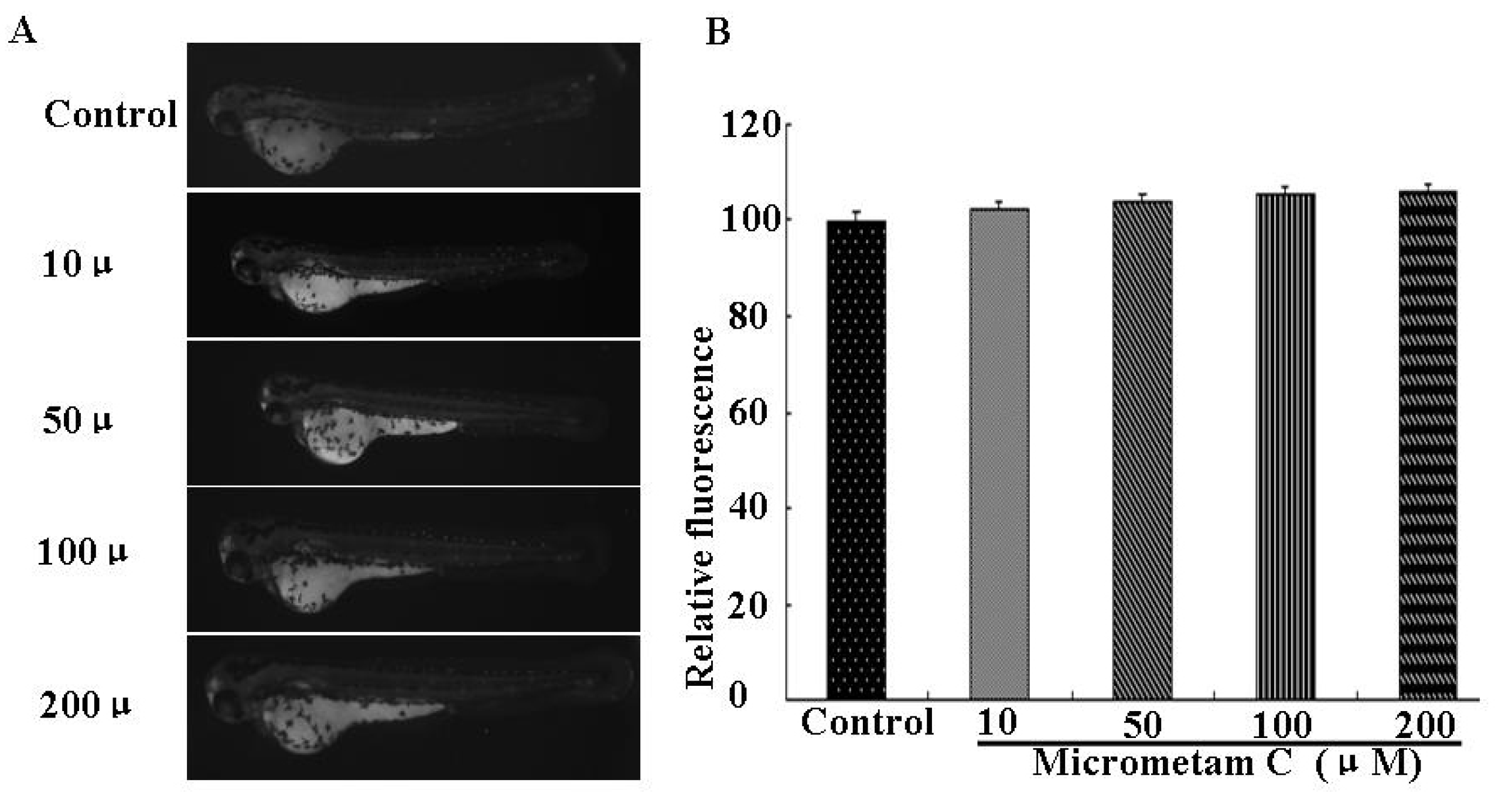
2.3. Effect of Micrometam C on Survival Rate, Heart Beat Rate and Morphological Changes in Zebrafish
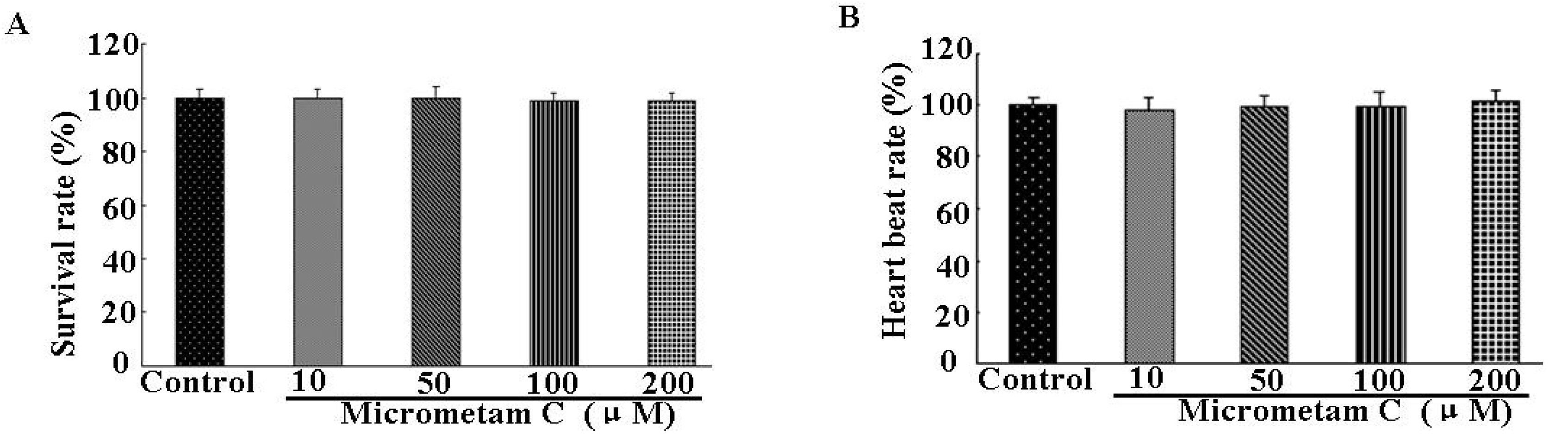
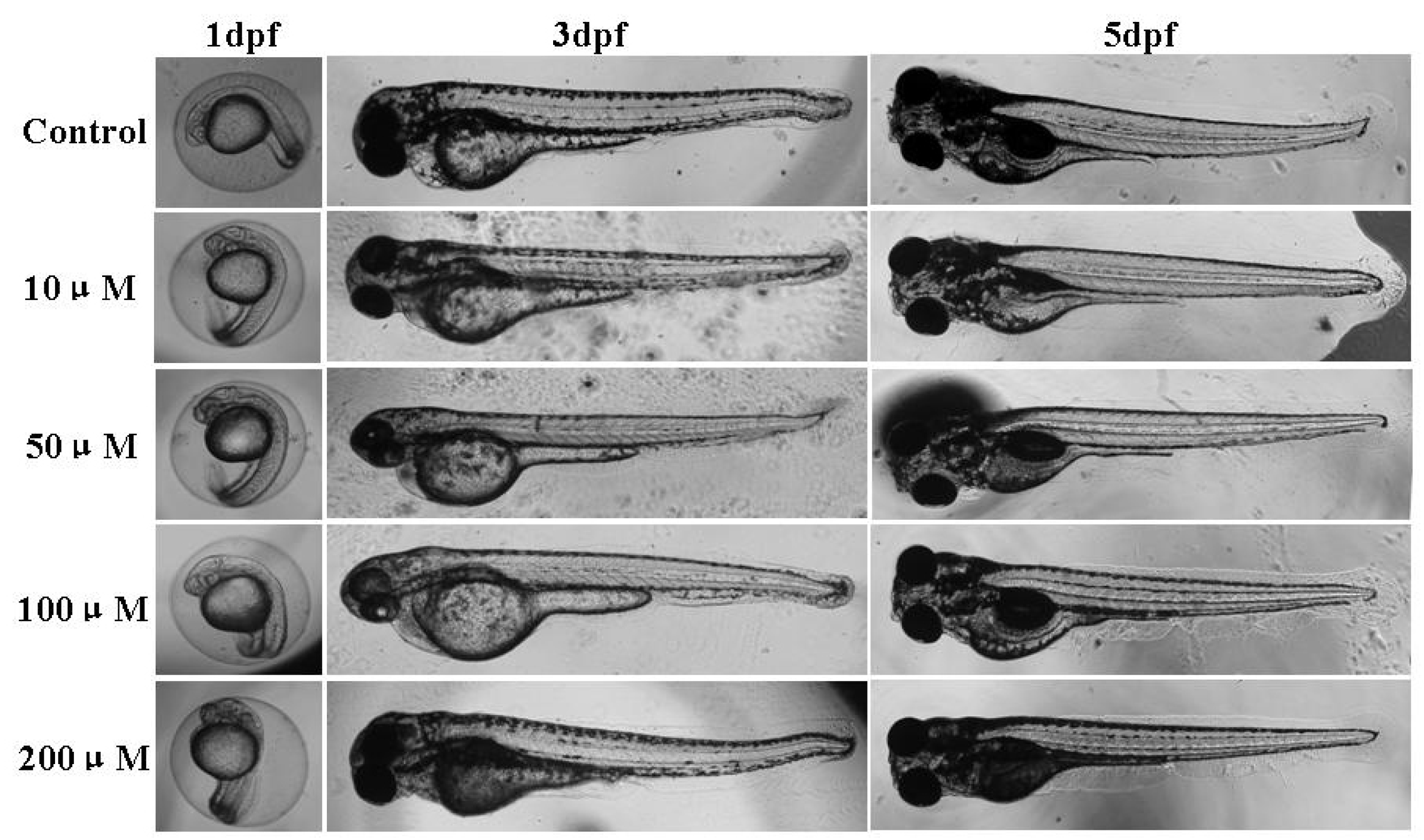
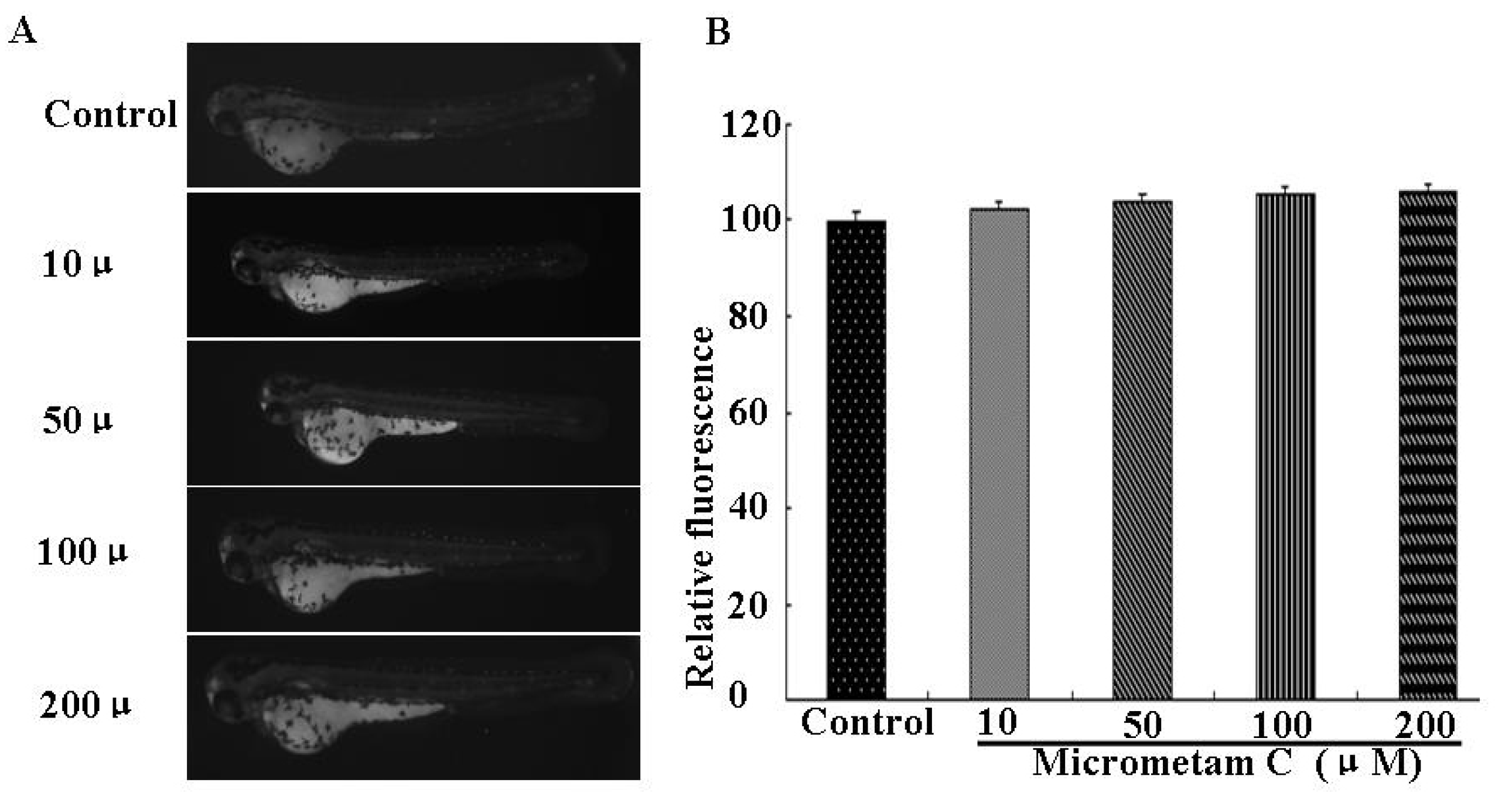
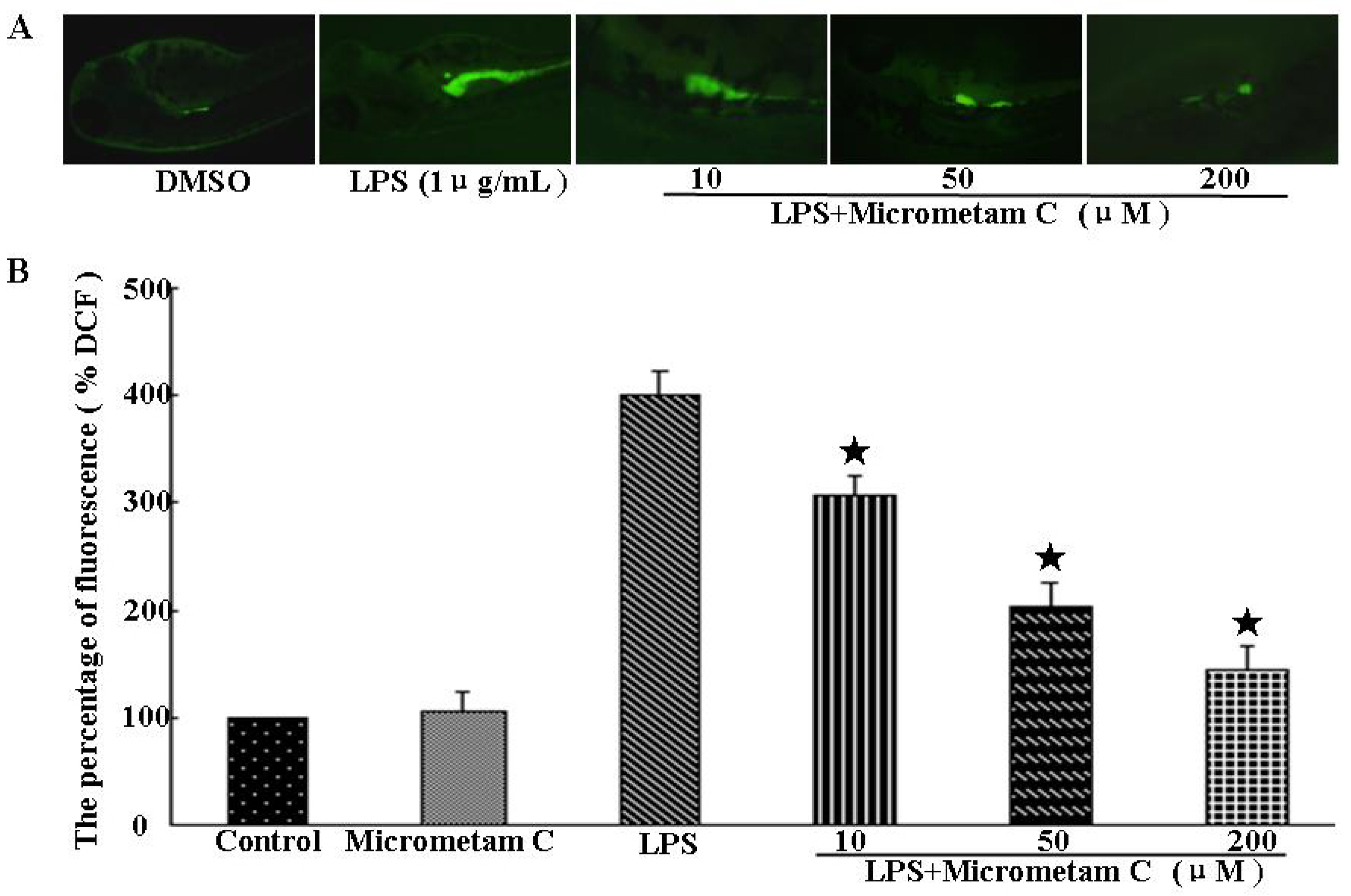
2.4. Effect of Micrometam C on LPS-Induced Oxidative Stress in Zebrafish Embryo

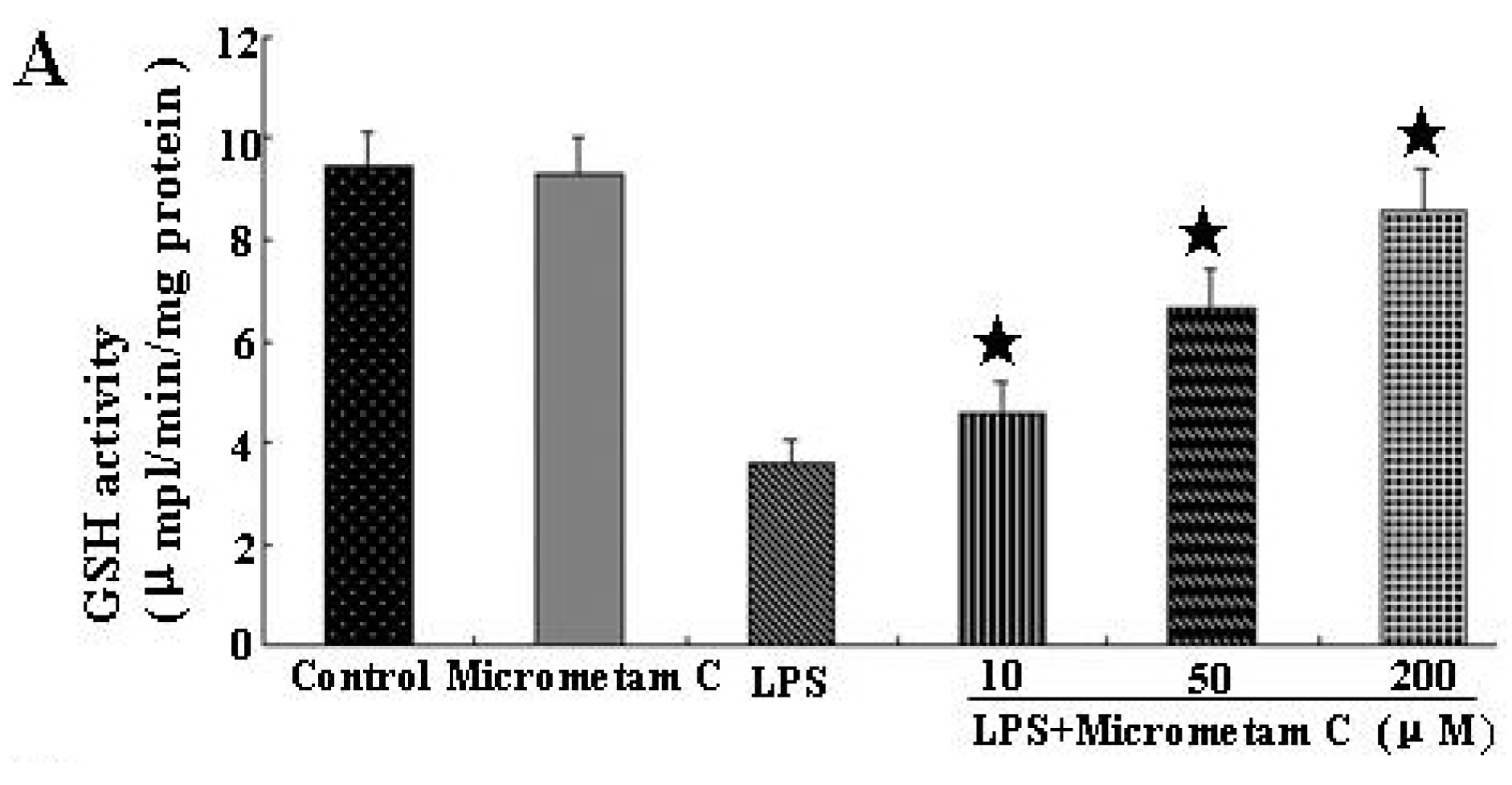
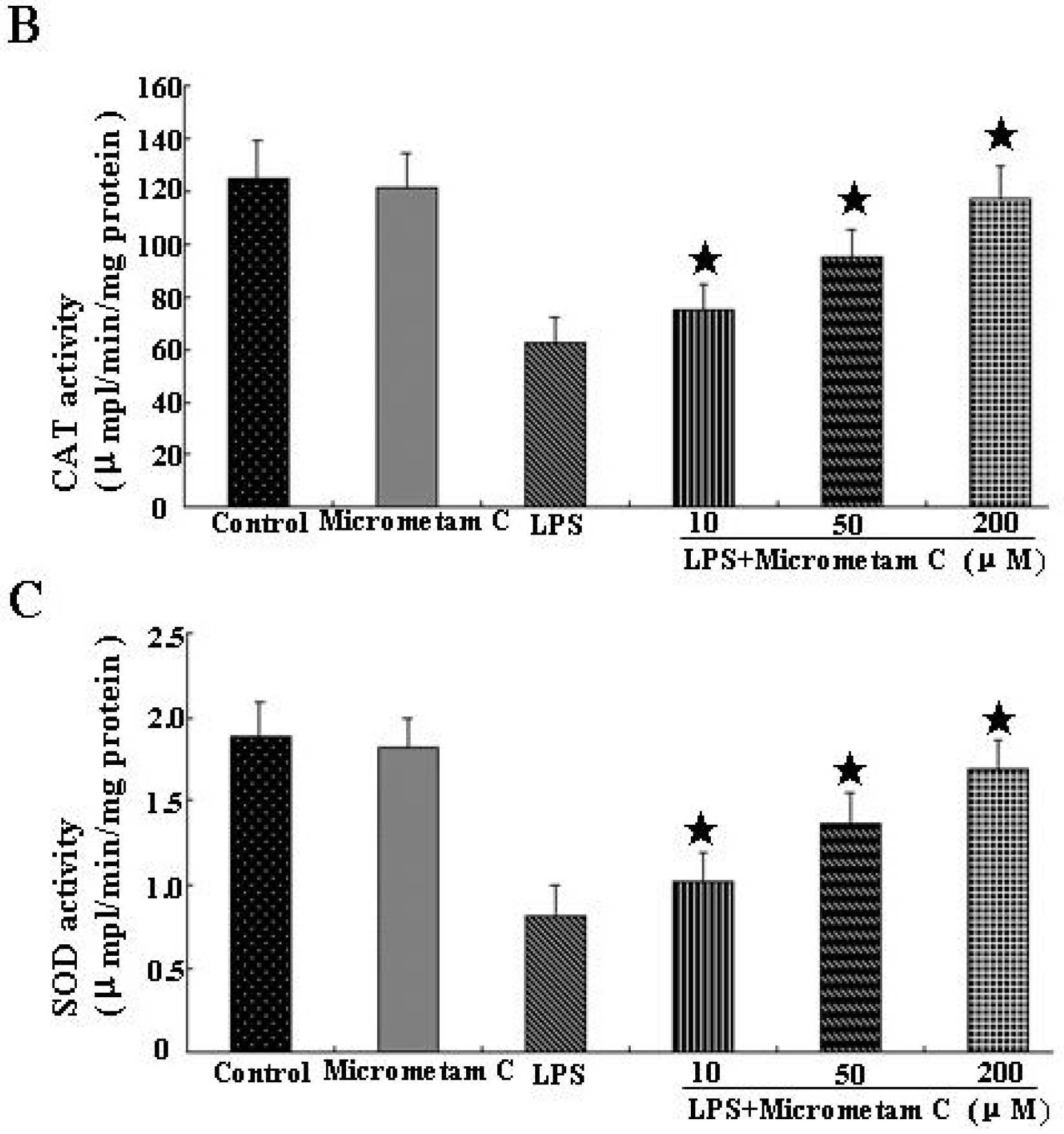
2.5. Micrometam C Restored LPS-Induced Depletion of CAT, GSH and SOD


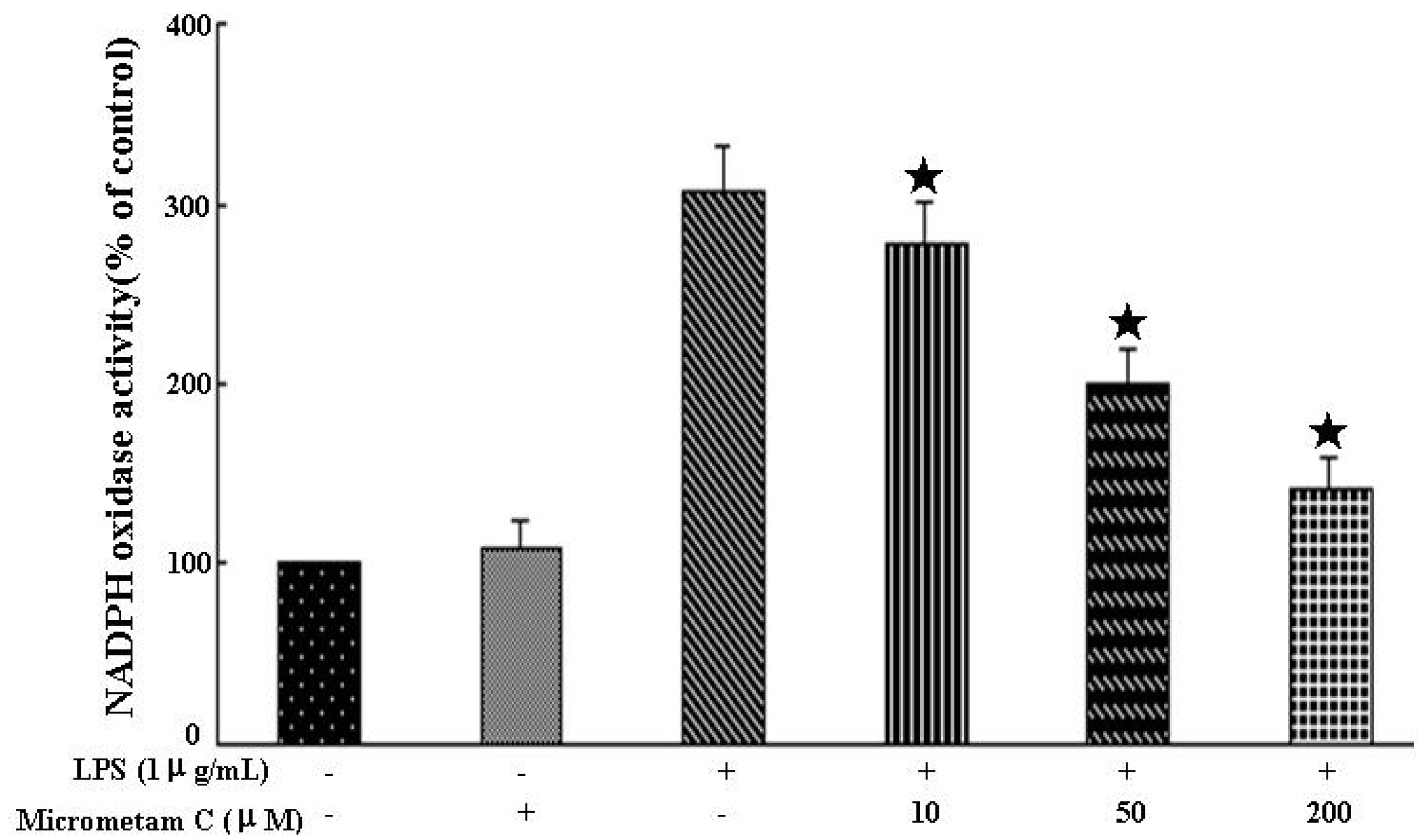
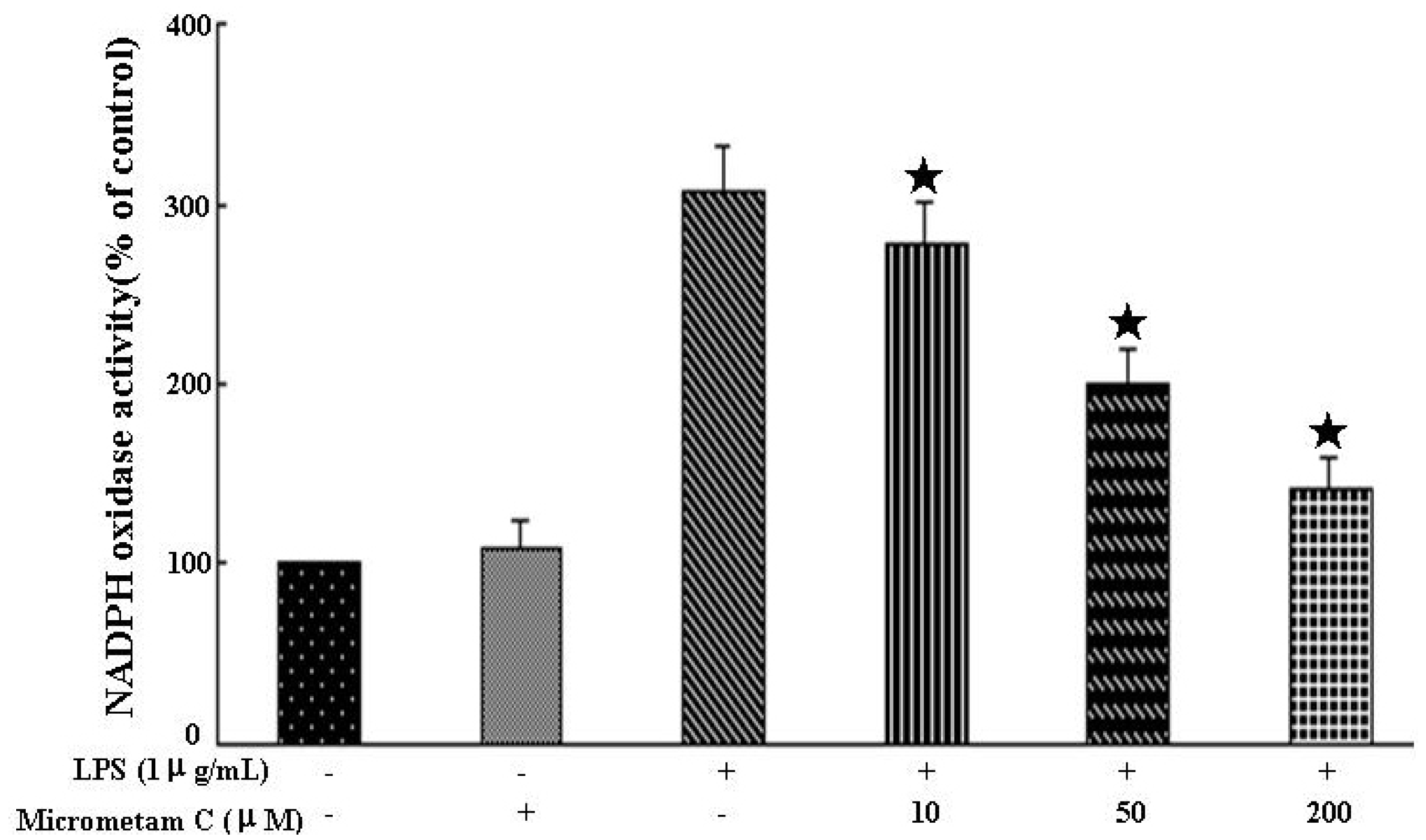
2.6. Effect of Micrometam C on LPS-Stimulated NADPH Oxidase Activity in Macrophages
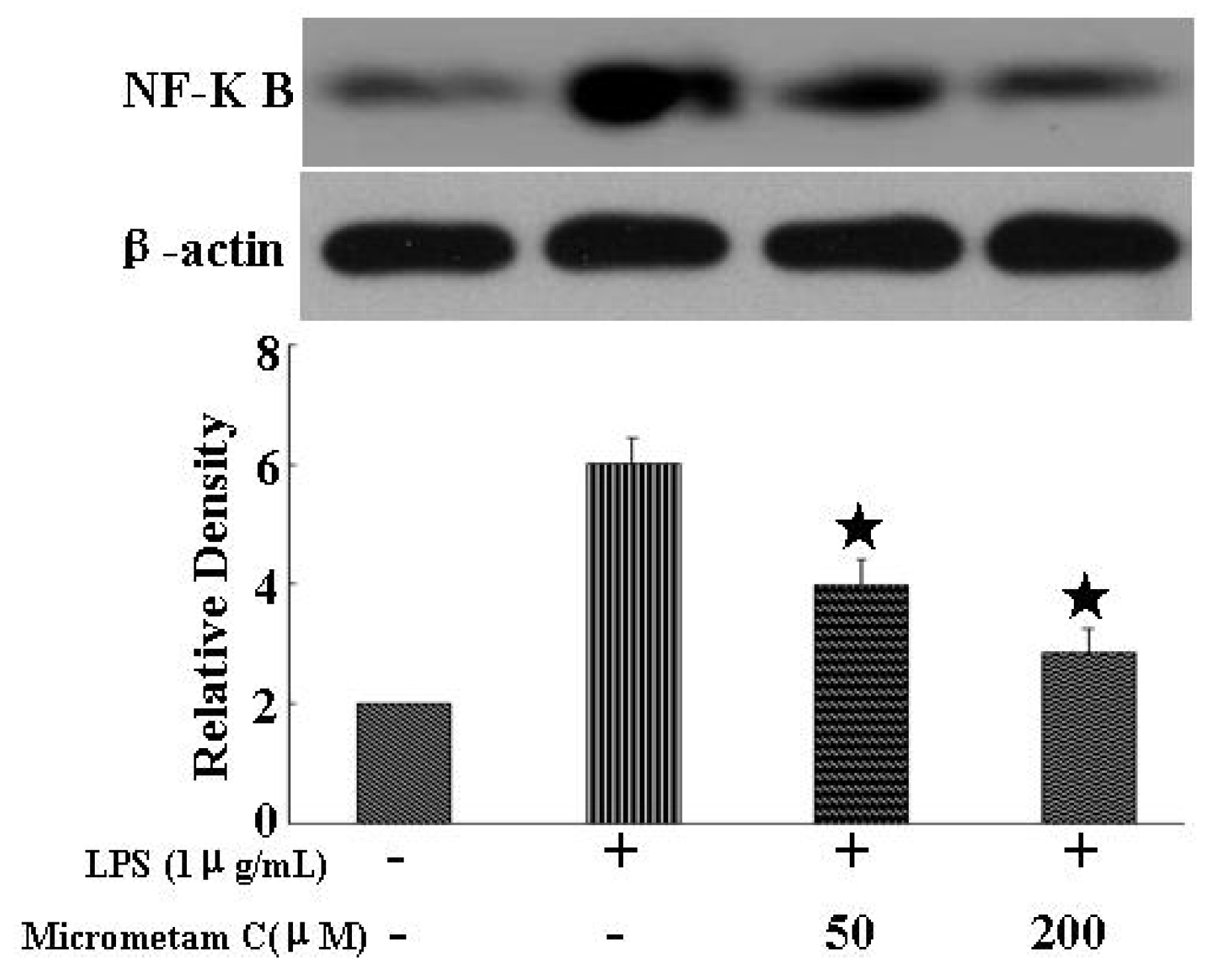
2.7. Anti-Inflammatory Effects of Micrometam C and Its Action on the NF-κB Signaling in Mouse Macrophage Cells in Vitro

3. Experimental Section
3.1. Preparation of Micrometam B and Micrometam C
3.2. The Isolation of Micrometam B and Micrometam C
3.3. Preparation of Inflammation-Induced Zebrafish Model by Tail-Cutting and Application of Micrometam C
3.4. Measurement of the Toxicity of the Isolated Compound
3.5. Respiratory Burst Assay in Zebrafish
3.6. Endogenous Antioxidant Assay
3.7. Cell Culture
3.8. NADPH Oxidase Activity Assay
3.9. Western Immunoblot Analyses
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Dewanjee, S.; Dua, T.K.; Sahu, R. Potential anti-inflammatory effect of Leea macrophylla Roxb. leaves: A wild edible plant. Food Chem. Toxicol. 2013, 59, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Terao, H.; Rikihisa, W.; Urabe, K.; Kinukawa, N.; Nose, Y.; Koga, T. Clinical dose and adverse effects of topical steroids in daily management of atopic dermatitis. Br. J. Dermatol. 2003, 148, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Süleyman, H.; Demircan, B.; Karagöz, Y. Anti-inflammatory and side effects of cyclooxygenase inhibitors. Pharmacol. Rep. 2007, 59, 247–258. [Google Scholar] [PubMed]
- Gaddi, A.; Cicero, A.F.; Pedro, E.J. Clinical perspectives of anti-inflammatory therapy in the elderly: The lipoxigenase (LOX)/cycloxigenase (COX) inhibition concept. Arch. Gerontol. Geriatr. 2004, 38, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Swain, P.; Nayak, S.K.; Nanda, P.K.; Dash, S. Biological effects of bacterial lipopolysaccharide (endotoxin) in fish: A review. Fish Shellfish Immunol. 2008, 25, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Chatter, R.; Ben Othman, R.; Rabhi, S.; Kladi, M.; Tarhouni, S.; Vagias, C.; Roussis, V.; Guizani-Tabbane, L.; Kharrat, R. In vivo and in vitro anti-inflammatory activity of neorogioltriol, a new diterpene extracted from the red algae Laurencia glandulifera. Mar. Drugs 2011, 9, 1293–1306. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.T.; Yang, C.M. Role of NADPH oxidase/ROS in pro-inflammatory mediators-induced airway and pulmonary diseases. Biochem. Pharmacol. 2012, 84, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Phelps, H.A.; Neely, M.N. Evolution of the zebrafish model: From development to immunity and infectious disease. Zebrafish 2005, 2, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Van der Sar, A.M.; Appelmelk, B.J.; Vandenbroucke-Grauls, C.M.; Bitter, W. A star with stripes: Zebrafish as an infection model. Trends Microbiol. 2004, 12, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.K.; Xu, Z.-R. Biomedical compounds from Marine organisms. Mar. Drugs 2004, 2, 123–146. [Google Scholar] [CrossRef]
- Mayer, A.M.; Rodriguez, A.D.; Berlinck, R.G.; Hamann, M.T. Marine pharmacology in 2005–6: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim. Biophys. Acta 2009, 1790, 283–308. [Google Scholar] [PubMed]
- Luo, X.; Huang, Y.; Zhang, S.; Yin, H.; Li, C.; Li, Q. Five new phenethyl cinnamides from the mangrove associates Micromelum falcatum. Biochem. Syst. Ecol. 2014, 56, 191–195. [Google Scholar] [CrossRef]
- Jin, H.; Sood, R.; Xu, J.; Zhen, F.; English, M.A.; Liu, P.P.; Wen, Z. Definitive hematopoietic stem/progenitor cells manifest distinct differentiation output in the zebrafish VDA and PBI. Development 2009, 136, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yan, B.; Shi, Y.Q.; Zhang, W.Q.; Wen, Z.L. Live imaging reveals differing roles of macrophages and neutrophils during zebrafish tail fin regeneration. J. Biol. Chem. 2012, 287, 25353–25360. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Oates, A.C.; Crowhurst, M.O.; Ward, A.C.; Layton, J.E. Morphologic and functional characterization of granulocytes and macrophages in embryonic and adult zebrafish. Blood 2001, 98, 3087–3096. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.; Flores, M.V.; Storm, T.; Crosier, K.; Crosier, P. The zebrafish lysozyme C promoter drives myeloid-specific expression in transgenic fish. BMC Dev. Biol. 2007, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.H.; Ko, S.C.; Kim, D.K.; Jeon, Y.J. Screening of marine algae for potential tyrosinase inhibitor: Those inhibitors reduced tyrosinase activity and melanin synthesis in zebrafish. J. Dermatol. 2011, 38, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.L.; Liu, J.X.; Wu, Q.; Long, S.M.; Zheng, M.Y.; Yao, X.L.; Ren, H.; Wang, Y.G.; Su, W.W.; Fai Cheung, R.T.; et al. Protective effects of puerarin against Aβ40-induced vascular dysfunction in zebrafish and human endothelial cells. Eur. J. Pharmacol. 2014, 732, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.X.; Chen, J.W.; Yuan, F.; Huang, Y.Y.; Zhao, L.Y.; Li, J.; Su, H.X.; Liu, J.; Pang, J.Y.; Lin, Y.C.; et al. Xyloketal B exhibits its antioxidant activity through induction of HO-1 in vascular endothelial cells and zebrafish. Mar. Drugs 2013, 11, 504–522. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Lu, X.L.; Yao, X.L.; Liu, Z.; Zhang, H.; Li, W.; Li, Z.; Wang, G.L.; Pang, J.; Lin, Y.; Xu, Z.; et al. Protective effects of xyloketal B against MPP+-induced neurotoxicity in Caenorhabditis elegans and PC12 cells. Brain Res. 2010, 1332, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Bannister, J.V.; Calabrese, L. Assays for superoxide dismutase. In Methods of Biochemical Analysis; Wiley: New York, NY, USA, 1987; Volume 32, pp. 279–312. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, H.; Ge, H.; Chen, Z.-B.; Luo, X.-M.; Su, F.-J.; Liang, Y.-B.; Li, Z.-Y.; Wu, J.-G.; Yang, Q.; Zeng, L.-J.; et al. Micrometam C Protects against Oxidative Stress in Inflammation Models in Zebrafish and RAW264.7 Macrophages. Mar. Drugs 2015, 13, 5593-5605. https://doi.org/10.3390/md13095593
Tang H, Ge H, Chen Z-B, Luo X-M, Su F-J, Liang Y-B, Li Z-Y, Wu J-G, Yang Q, Zeng L-J, et al. Micrometam C Protects against Oxidative Stress in Inflammation Models in Zebrafish and RAW264.7 Macrophages. Marine Drugs. 2015; 13(9):5593-5605. https://doi.org/10.3390/md13095593
Chicago/Turabian StyleTang, Hao, Hui Ge, Zhi-Bin Chen, Xiong-Ming Luo, Feng-Juan Su, Yan-Bing Liang, Zhen-Yu Li, Jing-Guo Wu, Qing Yang, Li-Jin Zeng, and et al. 2015. "Micrometam C Protects against Oxidative Stress in Inflammation Models in Zebrafish and RAW264.7 Macrophages" Marine Drugs 13, no. 9: 5593-5605. https://doi.org/10.3390/md13095593