
Are the Traditional Medical Uses of Muricidae Molluscs Substantiated by Their Pharmacological Properties and Bioactive Compounds?

 , ,
, ,
Abstract
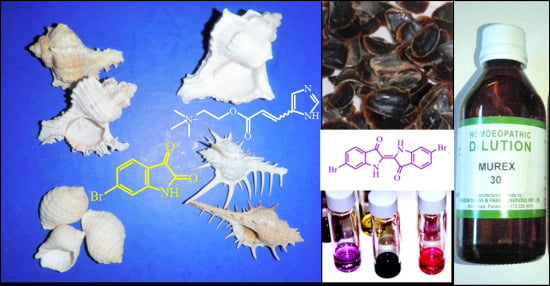
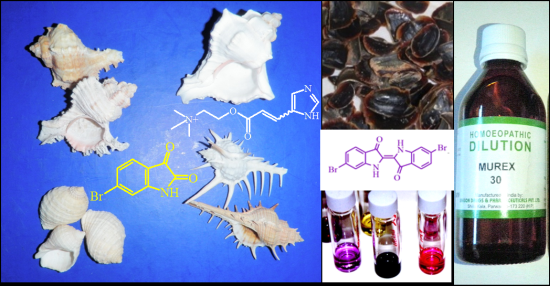
:
1. Introduction
2. Traditional Muricidae Bioresources

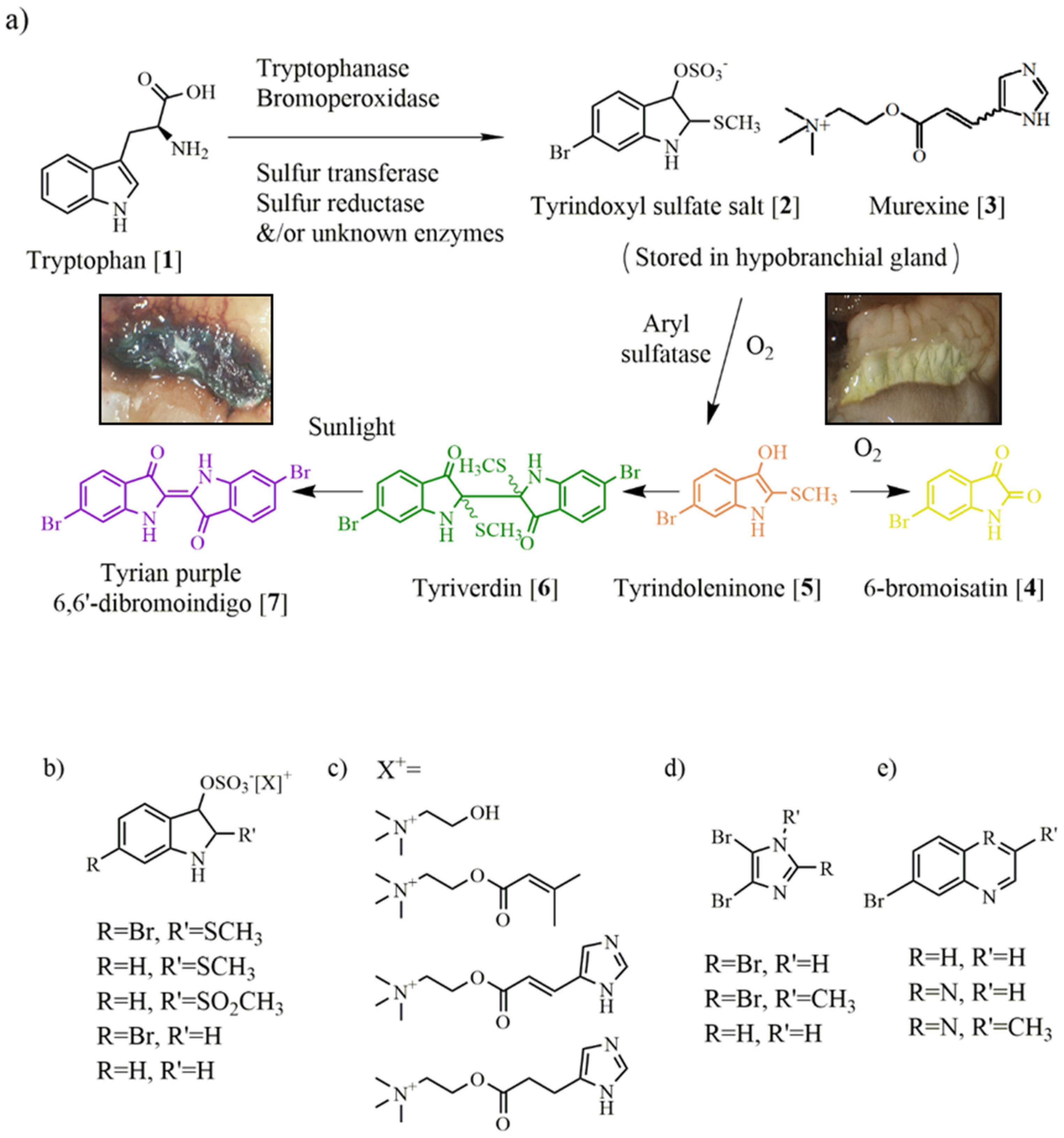
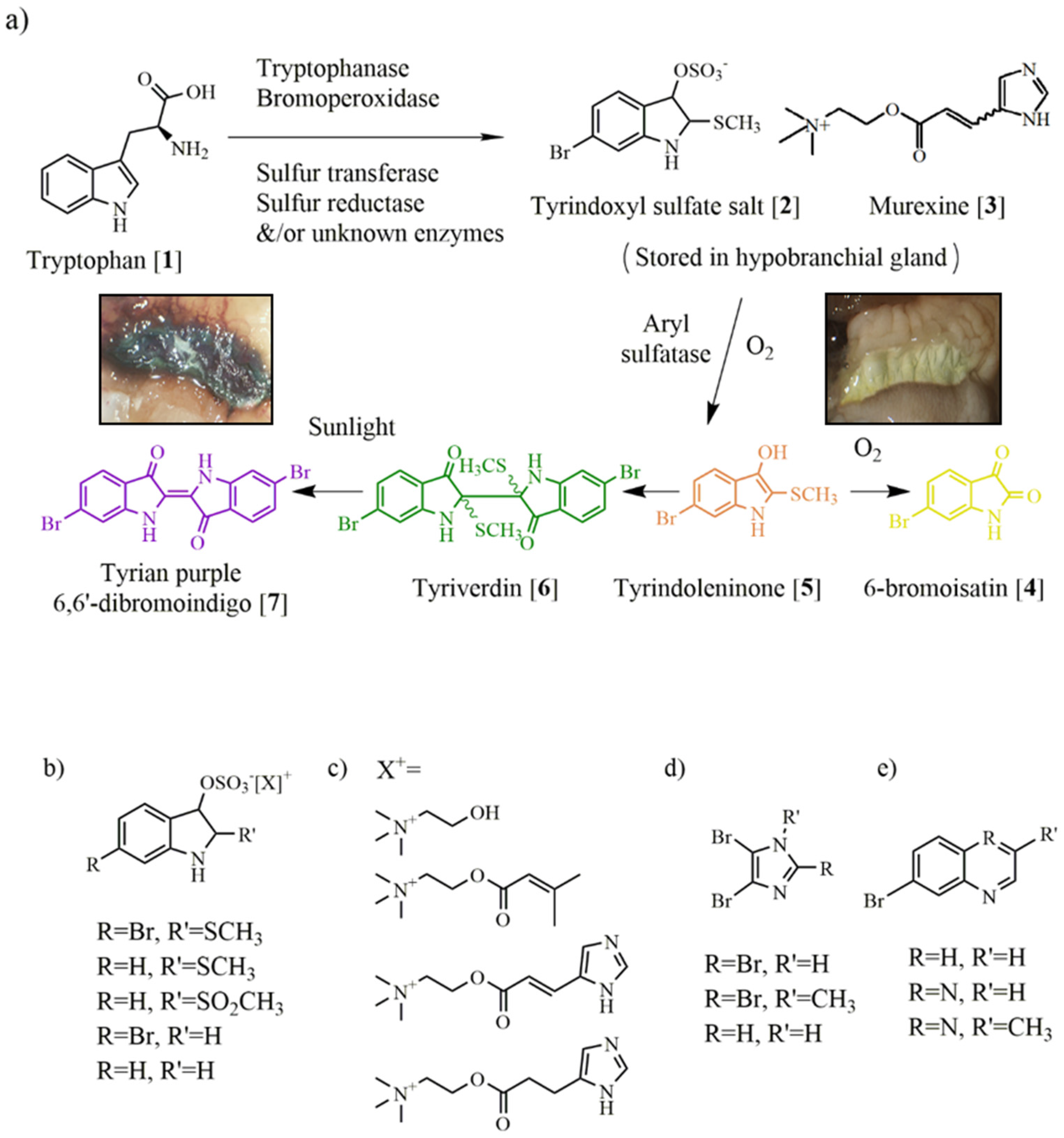
3. Muricidae Chemistry
3.1. Brominated Indoles and Related Compounds
3.2. Choline Esters
4. Bioactivity of Muricidae Extracts and Compounds
4.1. Antimicrobial and Antiviral Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source species 1 | Source Tissue | Extract or Compound | Activity Profile |
|---|---|---|---|
| Dicathais orbita [58,83,84] | egg mass | CHCl3 | Inhibits Gram +ve and Gram -ve human and marine pathogenic bacteria and C. albicans in the range of 0.1–1 mg/mL |
| Diethyl ether | Inhibits Gram +ve and Gram -ve human pathogens at 10 mg/mL | ||
| EtOH | Inhibits Gram +ve and Gram -ve human pathogens at 0.1 mg/mL | ||
| Tyriverdin | Inhibits human Gram +ve and Gram -ve pathogens at 0.0005 mg/mL, active against C. albicans and marine pathogens at 0.001 mg/mL | ||
| Tyrindoleninone | Inhibits human pathogens at 0.5–1 mg/mL, C. albicans and marine pathogens at 0.1 mg/mL | ||
| 6 Bromoisatin | Inhibits Gram +ve and Gram -ve human pathogenic bacteria in the range of 0.1–1 mg/mL, but >1 mg/mL for C. albicans and marine pathogens | ||
| Agnewia tritoniformis [83,84] | egg mass | CHCl3 | Inhibits human pathogens at 10 mg/mL |
| Bedeva paivae (Trophon hanleyi) [83,84] | egg mass | CHCl3 | Inhibits human pathogens at 10 mg/mL |
| Ocenebra (Ceratostoma) erinaceus [63,84] | egg mass | CHCl3 | Inhibits human pathogens at 1 mg/mL |
| 2,4,5-Tribromo-1H-imidazole | Inhibits human pathogens at 0.1 mg/mL | ||
| Chicoreus ramosus [79] | whole body | MeOH | Inhibited 58% of the marine biofilm bacteria tested |
| digestive gland | MeOH, H2O, DCM, Acetone | No activity against biofilm bacteria | |
| egg mass | MeOH | Inhibited 100% of the marine biofilm bacteria tested | |
| Chicoreus virgineus [79] | whole body | MeOH | Inhibited 50% of the marine biofilm bacteria tested |
| digestive gland | H2O, DCM or Acetone | No activity against biofilm bacteria | |
| operculum | MeOH | Inhibited 50% of the marine biofilm bacteria tested | |
| egg mass | MeOH | Inhibited 50% of the marine biofilm bacteria tested | |
| Drupella (Cronia) margariticola [79] | whole body | MeOH/DCM | Inhibited 63% of the marine biofilm bacteria tested |
| egg mass | MeOH | Inhibited 70% of the marine biofilm bacteria tested | |
| Phycothais (Lepsiella) reticulata [83,84] | egg mass | CHCl3 | Inhibits human pathogens at 10 mg/mL |
| Tenguella (Morula) marginalba [83,84] | egg mass | CHCl3 | Inhibits human pathogens at 10 mg/mL |
| Murex tribulus [79] | whole body | Acetone | Inhibited 60% of the marine biofilm bacteria tested |
| Rapana rapiformis [79] | whole body | Acetone | Inhibited 23% of the marine biofilm bacteria tested |
| Rapana venosa [85] | haemolymph | Proline rich peptides | Inhibited Gram +ve (S. aureus) and Gram -ve (Klebsiella pneumoniae) |
| Rapana venosa [86,87] | haemolymph | Haemocyanin | Inhibits the replication of Epstein-Barr virus at 1 μg/mL and Herpes simplex virus type 1 at 200 μg/mL |
| Stramonita (Thais) biserialis [79] | whole body | MeOH | Inhibited 35% of the marine biofilm bacteria tested |
| Purpura (Thais) bufo [79] | whole body | MeOH | Inhibited 25% of the marine biofilm bacteria tested |
| Semiricinula (Thais) tissoti [79] | whole body | MeOH | Inhibited 18% the marine biofilm bacteria tested |
| Hexaplex trunculus [63,84] | egg mass | CHCl3 | Inhibits S. aureus at 1 mg/mL and E. coli at 10 mg/mL |
| 2,4,5-Tribromo-1H-imidazole | Inhibits human pathogens at 0.1 mg/mL |
4.2. Wound Healing and Anti-Inflammatory Activity
4.3. Anticancer and Kinase Receptor Binding Activity
| (a) In Vitro Assays | ||
| Source Species/Compounds | Cell Line or Purified Protein | Assays 1 and Effects Examined |
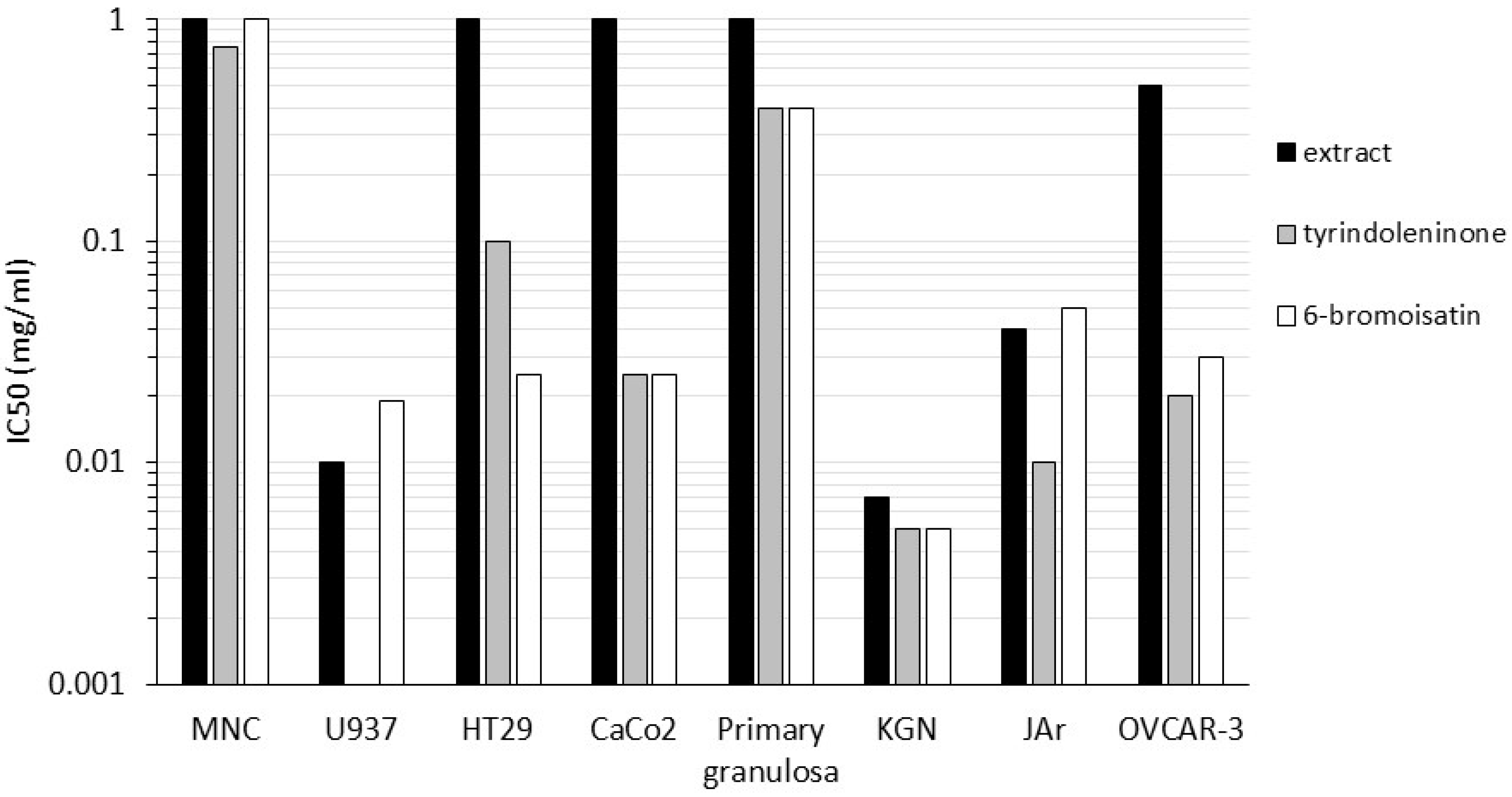
| Dicathais orbita (Chloroform extracts, purified tyrindoleninone & 6 Bromoisatin) [51,68,69,104] | A range of female reproductive, colon and breast tumurs and lymphomas (Figure 3) | MTS/MTT cell viability; Crystal violet; Caspase 3/7 activity for apoptosis; Lactate dehydrogenase for necrosis; Tunnel staining for apoptosis; Flow cytometry for apoptosis, necrosis and cell cycle analysis |
| Dicathais orbita (extracts and compounds) [69,106] | JAr and human granulosa cells | Radioimmunoassays (RIA); Steroidogenesis assays: estradiol (E2) and progesterone (P4) synthesis |
| Hexaplex trunculus (purified bromoindirubins) [107,108] | Recombinant or naturally purified protein kinases | CDK1/Cyclin B, CDK5/p25, GSK-3 and other protein receptor kinase assays |
| Rapana venosa (Ethanol extracts) [109] | Human leukemia HL-60 and human lung cancer A-549 | MTT cell viability and liquid-scintillation radioassay for cell proliferation (3H-TdR) |
| Thais clavigera (Ethanol extracts) [109] | Human leukemia HL-60 and human lung cancer A-549 | MTT cell viability and liquid-scintillation radioassay for cell proliferation (3H-TdR) |
| Rapana thomasiana (Purified haemocyanin) [110] | SiHa-cervical squamous cell carcinoma, CaOV-ovarian adenocarcinoma, MIA PaCa-pancreatic carcinoma, RD 64-rhabdomyosarcoma, EJ-urinary bladder carcinoma and Lep-nontumor human lung cell line. | Cell proliferation assay and apoptosis indicated by DNA degradation and caspase-3 activation |
| Rapana venosa (Haemocyanins) [111] | 647-V, T-24 and CAL_29 bladder cancer cells | MTT AND WST-1 cell viability assays, apoptosis with acridine orange/propidium iodine staining and gene expression profiles for 168 inflammatory cytokines and signal transduction pathways. |
| Synthetic isatin derivatives [105,112] | The human leukemic (U937, monocyte and Jurkat, T cell), breast (MDA-MB-231 and MCF-7), prostate (PC-3), and colorectal (HCT-116) | MTS cell viability, caspase 3/7 for apoptosis, CDK2 inhibition |
| Synthetic indirubin derivatives [107,108,113,114,115,116] | Recombinant or naturally purified protein kinases | CDK1/Cyclin B, CDK5/p25, GSK-3 and other protein receptor kinase assays; affinity chromatography; crystallography and in silico modelling; rt PCR on Xenopus embryos |
| Synthetic indirubin derivatives [117,118] | Human neuroblastoma and breast cancer cell lines | Apoptosis induction pathways |
| Synthetic indirubin derivatives [119,120] | Human melanoma andmyeloid leukemia cell lines | Jak/Stat 3 phosphorylation, FLT3 inhibition |
| Synthetic isatin and indirubin derivatives [69,106,121] | JAr and human granulosa cells for female hormones and H294 adrenal cells for male | RAI; ELISA for E2 and P4; E-screen (xeno-oestrogenic potential) for E2 receptor binding; H294 adrenal cells for cortisol, testosterone, androgen, and didehydroepiandrosterone |
| Synthetic indirubin derivatives [122] | JAr and human granulosa cells | RAI |
| (b) In Vivo Models | ||
| Source Species/Compounds | Cancer Type | Animal Model |
| Dicathais orbita (Chloroform extracts) [123], (purified tyrindoleninone, 6 bromoisatin) [124] | Colon cancer prevention | Apoptotic response to genotoxic damage by azoxymethane (AOM) in mice. Compounds delivered by oral gavage two weeks prior to AOM |
| Concholepas concholepas (haemocyanin subunits CCHA & CCHB) [92] | Bladder carcinoma treatment | MBT-2 heterotopic murine bladder carcinoma model |
| Synthetic 6-bromoistain [112] | Colon cancer prevention | Apoptotic response to genotoxic damage by AOM in mice. Compound delivered by oral gavage two weeks prior to AOM |
| Synthetic 6-bromoindirubin derivatives [119] | Human melanoma treatment | Xenograph model in BALC/c mice, 14 day treatment |
| Synthetic indirubin derivatives [125] | Renal, prostate, lung and colon cancer treatment | Xenograph model in BALC/c mice |

4.4. Steroidogenic Effects of Muricidae Extracts and Analogue Compounds
| Source | Compound | Cell Type | Hormone | Dose Response |
|---|---|---|---|---|
| Synthetic compounds 0, 0.00001, 0.0001, 0.001, 0.01, 0.1 mg/mL | 6,6′ dibromoindirubin | JAr | Basal P4 | N-shaped 1, low doses stimulated, 0 and high doses N/E, 4 h, 6 h, 8 h, 10 h |
| indirubin | Basal P4 | N-shaped, Low doses stimulated, 0 and high doses N/E, 24 h | ||
| 5-bromoisatin | Basal P4 | N-shaped, low doses stimulated, 0 and high doses N/E, 24 h | ||
| indirubin | GC | Basal P4 | U-shaped 2, 48 h, 72 h | |
| 5-bromoisatin | Basal P4 | Dose-dependent stimulation, 48 h | ||
| Hypobranchial gland extract 0, 0.005, 0.01, 0.05, 0.1, 0.5, 1 mg/mL | mixture of 6-bromoisatin, tyrindolinone, tyrindoleninone, tyrindoxyl sulphate | GC | Basal P4 | N/E |
| hCG P4 | N/E | |||
| Basal E2 | 0.05 mg/mL stimulated, 24 h | |||
| hCG E2 | U-shaped, 4 h, 24 h, 48 h | |||
| tyrindoleninone | GC | Basal P4 | N/E | |
| hCG P4 | N/E | |||
| Basal E2 | N-shaped, 24, 48 h | |||
| hCG E2 | N-shaped, 24, inhibition—48 h | |||
| 6-bromoisatin | GC | Basal P4 | U-shaped 4 h, N/E 24 h, 48 h | |
| hCG P4 | N/E | |||
| Basal E2 | U-shaped 4 h, N/E 24 h, 48 h | |||
| hCG E2 | Dose-dependent inhibition 4 h, 48 h | |||
| Egg mass extract 0, 0.005, 0.05, 0.5 mg/mL | mixture of 6-bromoisatin, tyrindolinone, tyrindoleninone, tyriverdin | GC | Basal P4 | N/E |
| hCG P4 | Dose-dependent stimulation, 48 h | |||
| Basal E2 | N/E | |||
| hCG E2 | Dose-dependent stimulation, 48 h | |||
| tyrindoleninone | GC | Basal P4 | N/E | |
| hCG P4 | Dose-dependent inhibition | |||
| Basal E2 | N-shaped 24, 48 h | |||
| hCG E2 | Stimulation 4, 24 h, n-shaped 48 h | |||
| Egg mass extract 0, 0.005, 0.05, 0.5 mg/mL | 6-bromoisatin | GC | Basal P4 | Lowest dose cytotoxic |
| hCG P4 | Dose-dependent inhibition, 48 h | |||
| Basal E2 | Lowest dose cytotoxic | |||
| hCG E2 | Dose-dependent stimulation, 48 h |
4.5. Muscle Relaxing and Nicotinic Activity of Choline Esters
| (a) | ||||||
| Source Compound a | Conc. | Assay/in Vivo Model | Observed Effects | |||
| HG Extract/Synthetic | Neuromuscular block Paralysis of the skeletal musculature after intravenous (i.v.) administration. Murexine stimulates nicotinic acetylcholine receptors opening the monovalent cation channel for depolarization of the motor endplate. Mild or no muscarine like activity was detected in guinea-pig and rabbit intestine, rabbit atrium and isolated frog heart assays. | |||||
| M, MCH, DHM, SCH | various | Frog rectus abdominis muscle assay [74,76,140] | ||||
| M, MCH | 0.75 mg/assay | Neuromuscular block in rat diaphragm assay [140] | ||||
| M, MCH | 100–2000 μg/kg | % Twitch reduction assays on cat, dog and rabbit gastro-cnemius were used to calculate concentration effect [75] | ||||
| M, MCH | 0.65 mg/kg | 50% rabbit head drop after i.v. injection [74] | ||||
| DHM | 0.52 mg/kg | |||||
| M, MCH | 0.35 mg/kg | 50% dog paralyzing dose after i.v. injection [140] | ||||
| DHM | 0.022 mg/kg | |||||
| M, MCH | 1.0–1.2 mg/kg | Relaxing effect in human preliminary clinical trial on 160 patients after a single i.v. injection [140] | ||||
| HG Extract/Synthetic | Nicotinic activity Murexine i.v. at high dose is likely to have nicotinic effects on sympathetic ganglia and adrenal medulla | |||||
| M | 60 μg/kg/min (no effect) | Nicotinic effects of murexine showed a dose dependant rise in blood pressure whilst inducing neuromuscular block in anaesthetised cats and dogs [74,140,141] | ||||
| M | 300 μ/kg | |||||
| (b) | ||||||
| Compound | Conc. | Assay/in Vivo Model and Effects Observed | Method Admin. | Mode of Action/Pathways Identified | Overall Effects | |
| 2,3 dioxoindoline | 15–20 mg/kg | Mice and rats showed anxiogenic behaviour in the open-field and elevated plus maze test and the social interaction test [142] | i.p. | Monoamine oxidase B inhibition as a contribution to stress related tribulin activity. | Anxiogenic at low dose | |
| <50 mg/kg | Mice showed immobility in the forced swim test [143] | i.p. | Inhibits monoamine oxidase affecting monoamine levels. | Sedative at high dose | ||
| indole-2,3-dione | 20 mg/kg | Isatin administered after pentylenetetrazole (PTZ) and 3-mercaptopropionic acid (3MPA) induced seizures in rats [143] | i.p. | Antagonise natriuretic peptide receptor A (NPR-A) and NPR-C signalling at low dose due to the metabolite 5-hydroxyisatin. | Proconvulsant at low dose | |
| 60–80 mg/kg | Effective against PTZ and 3MPA induced clonic convulsions [144] | i.p. | Anticonvulsant at high dose | |||
| indole-2,3-dione-3-oxime or as salt, oxide or hydrate derivative | 10 uM | Intermediate-conductance and small-conductance Ca(2+)-activated potassium channel (IKCa and SKCa)activation in a 15 ul cell chamber on human embryonic kidney 293 cell line [145] | immersion | Ikca and SKca ion channel associated conditions including respiratory conditions, muscle spasms, convulsive conditions, mood disorders and dementia. | Ikca and Skca ion channel activation | |
| 5,7-dinitro-1-methyl-1H-indole-2,3-dione-3-(0-methyloxime) | 0.1–10 mg/kg | Administered to NMRI mice for ATPA rigidity, to DAB/2 mice for quisqualate seizures, to NMRI mice for N-methyl-d-aspartate (NMDA) seizures and to NMRI mice for cocaine hypermotility [146] | i.v. and orally (cocaine hypermotility) | Excitatory amino acid antagonist blocking glycine and glutamate on the quisqualate, 2-amino-3-(3-hydroxy-5-tert-butylisoxazol-4-yl)propionic acid (ATPA), 2-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA), kainate and NMDA receptors. | Anticonvulsant for ATPA quisqualate, NMDA seizures and cocaine hypermotility | |
| 5-bromoisatin | 200 mg/kg | Phenylquinone test for analgesia in mice showing 90% inhibition after 30 min [147] | i.p. | 5-bromoisatin was comparable to acetylsalicylic and showed analgesia with fewer side effects. | Analgesic | |
| 400 mg/kg | Randall and Selitto test for analgesic comparison in rats [147] | orally | ||||
| 90 mg/kg | Overall ED50 after 30 min reaction time [147] | |||||
4.6. Neurotransmitters, Analgesic and Sedative Properties of Isatins
4.7. In Vivo Toxicity
| Compound/Extract | Model | Test | Method Admin | Conc. (mg/kg) | General Side Effects |
|---|---|---|---|---|---|
| murexine | Mouse | LD50 | i.v. | 6.45 | Paralysis of the skeletal musculature preceded by transient stimulation including muscle tension and fasciculation. Death is caused by anoxia after peripheral respiratory arrest [74] |
| LD50 | s.c. | 50 | |||
| LD (ineffective) | oral | <1000 ineffective | |||
| Dog | GE | i.v. | 0.27 | For dogs, murexine additionally caused increased saliva and evacuation of urine and faeces (defecation). Nevertheless they handled 200 times dose with artificial respiration (intubation) [74] | |
| GE | s.c. | 1.35–2.16 | |||
| Pigeon | GE | i.v. | 0.05 | Birds developed contracture, leg cramp and opisthotonus instead of muscular paralysis [74] | |
| LD50 | i.v. | 0.2–0.3 | |||
| Octopus | GE | i.b.h. | 30–40 | Eledone moschata showed brief stimulation and motor agitation with deep respiratory behaviour [74] | |
| Humans | GE | i.v. | 1–1.2 | Muscle relaxation with mild nicotinic effects [140] | |
| dihydromurexine | Mouse | LD50 | i.v. | 5.57 | Similar to murexine but more potent for mice. 12 times more potent for frog rectus abdominis [74] |
| tigloylcholine | Mouse | LD50 | i.v. | 0.92 | Considerably more potent than murexine [77] |
| D. orbita brominated indole extracts | Mouse | GE | oral | 0.5 | Idiosyncratic effects on hepatocytes including nonsteroidal fatty change and necrosis [152] |
| GE | oral | 0.5 | Idosyncratic gastrointestinal inflammation and ulcers [152] | ||
| LD (ineffective) | oral | >1 | No mortality after 4 weeks daily oral gavage, no effects on behaviour or any signs of ill health [153] | ||
| Rat | GE | oral | 1 | No mucositic, inflammation, or negative effects of gastric epithelium or blood cells [103] | |
| LD (ineffective) | oral | >1 | No mortality after 1 week daily oral gavage, no effects on behaviour or any signs of ill health [103] | ||
| 6-bromoisatin | Mouse | GE | oral | 0.25–1 | Diuretic effects evidenced by reduced K/Na ratio in blood no negative effects on blood cells, hemoglobin or serum liver enzymes [112] |
| LD (ineffective) | oral | >1 | No mortality after 2 weeks daily oral gavage, no effects on behaviour or any signs of ill health [112] |
5. Traditional Medical Uses
5.1. Ancient Mediterranean and Middle Eastern Use
| Culture | Part of snail | Source species | Preparation | Pharmaceutical properties (Treatment) |
|---|---|---|---|---|
| Ancient Greco-Roman (Dioscorides, Oribasius and Galen) [21] | Operculum | Hexaplex trunculus, Bolinus brandaris, Thais haemastoma | Crushed and mixed with oil and vinegar | Hearing loss, swollen spleen, depression, menstrual cycle abnormalities, after labour for placenta removal |
| Flesh and ashes of burned shell | Hexaplex trunculus, Bolinus brandaris and Thais haemastoma | Burned flesh along with shell | Wound healing, cleaning teeth, treatment of cracked skin, healing parotid gland swelling, anti-inflammatory properties | |
| Ancient Greece (Dioscorides) [155] | Whole shell with meat | Muricidae (Purpura-Tyrian Purple producing shellfish) | Burnt and dry whole animal | Cleaning teeth, healing warts, boils or tumour |
| Columellae | “Purpurae” (Muricidae) | Burnt and dry | Good for stomach | |
| Ancient Greece (Xenocrates) [21] | Hypobranchial gland | Hexaplex trunculus, Bolinus brandaris and Thais haemastoma | unknown | Laxative, diuretic, increases salivary secretion, perspiration.Excess consumption may cause nausea, vomiting and diarrhoea |
| Ancient Greece (Athenaeus) [21] | Foot | Hexaplex trunculus, Bolinus brandaris and Thais haemastoma | Boiling | Heals stomach disorders |
| Medieval Eastern Mediterranean Genizah [14,155] | Operculum | Muricidae such as Murex anguliferus | Smell the aromatic substance or smoke produced while placing the operculum on slowly burning charcoal | Rheumatism or arthritis Stomach problem (wounds in stomach), skin diseases, teeth problems, eye and ear diseases, tumors, epilepsy , paralysis, purgative, treatment of diseases of the uterus |
| Bahrein Middle Eastern [13,15] | Operculum | Murex inflatus | Fumigation | Atresia of uterus |
| Europe (Aphrodisiacs) [156] | Operculum (Blatta byzantine) | Banded dye Murex | Operculum medicine with vinegar Fumigation | Reduced swollen spleen For women (dislodge the placenta after labour) |
| Operculum | Calcified operculum | Ashes of calcified operculum | Stimulate capillary growth |
5.2. Muricidae Used in Traditional Chinese Medicine (TCM)
| Scientific Name | Method of Use | Traditional Uses and Claims |
|---|---|---|
| Genus Ceratostoma | ||
| C. rorifluum (Adams & Reeve) | Decoct the shell (10–50 g) and ingest. | Tranquilize and sedate the mind; astringe and preserve the essence; Use to treat insomnia, amnesia, spermatorrhea, uterine bleeding and leukorrhagia. |
| Genus Chicoreus | ||
| C. asianus (Kuroda) | Same as C. rorifluum | Same as C. rorifluum |
| C. brunneus (Link) | Decoct the shell (15–25 g) and ingest. | Resolve phlegm, disperse retention, tranquilize liver and wind; Use to treat stomach pain, scrofula and spastic muscles. |
| C. ramosus (Linnaeus) | Decoct the crushed shell (15–50 g); Ustulate (scorch) the shell, ground into powder and apply externally. | Clear heat, expel toxins, soften hard lumps, dispel nodes, reduce flatulence and pain; Use to treat pathopyretic ulcer, scrofula (infection of the lymph nodes), stomach pain, dyspepsia, stomach and duodenal ulcer. |
| Genus Murex | ||
| M. aduncospinosus (Beck) 2 | Decoct the shell (15–25 g) and ingest. Ustulate the shell, ground into powder and apply externally. | Clear heat, expel toxins, invigorate blood circulation. Use to treat pyretic toxicity, carbuncle, furuncle, otitis medium and ulcer of lower limb. |
| M. pecten (Lightfoot) 3 | ||
| M. rectirostris (Sowerby) 4 | ||
| M. ternispina (Lamarck) | ||
| M. trapa Röding | ||
| Genus Nassa | ||
| N. francolinus (Bruguière) 5 | Same as genus Murex | Same as genus Murex |
| Genus Purpura | ||
| P. rudolphi (Lamarck) | Same as T. alouina, etc. | Same as T. alouina, etc. |
| Genus Rapana | ||
| R. bezoar (Linnaeus) | Fresh meat: boil and eat the meat; decoct the shell (30–60 g) and ingest. Combine the juice and with other medicine as eye drops | Fresh meat: Remove heat to brighten vision; Use to treat hepatic heat and red eyes, ophthalmalgia, chest and abdomen heat and pain. |
| R. bezoar (Linnaeus) | Shell: Decoct the shell (15–30 g) and ingest, used as medicinal powder (3–6 g) and ingest; Ustulate the shell, ground into powder, mixed with sesame oil and apply externally. Operculum: Decoct the operculum (10–20 g) and ingest; Ustulate the shell, ground into powder and apply externally. | Shell: Relieve gastric hyperacidity to alleviate stomachache, resolve phlegm, disperse retention, tranquilize liver and wind; Use to treat stomach and duodenal ulcer, panasthenia, spastic hand and foot, chronic osteomyelitis, and scrofula. Operculum: Clear heat, expel toxins, remove dampness through diuresis, free strangury. Use to treat strangury (painful & frequent urination), swelling and ulcer on the body surface, hepatic coma, eye diseases, dysentery. |
| R. rapiformis (Born) | Decoct the shell (15–25 g) and ingest. | Eliminating phlegm and soften indurated mass, relieving convulsion and spasm, relieve gastric hyperacidity to alleviate stomach ache; Use to treat stomach pain, scrofula, spastic hand and foot. |
| R. venosa (Valenciennes) | same as R. bezoar | same as R. bezoar |
| Genus Thais | ||
| T. alouina (Röding) 6 T. armiger (Link) 7 T. bronni (Dunker) 8 T. bufo (Lamarck) 9 T. clavigera (Kuster) 10 T. echinata (Blainville) 11 | Decoct the shell (15–50 g) and ingest; used for making pills or medicinal powder; Ustulate the shell, ground into powder and apply externally. | Soften hard lumps, dispel nodes, clear heat, expel toxins; Use to treat pyogenic infection, swelling and ulcer on the body surface and scrofula. |
| T. gradate (Jonas) 12 | Decoct the crushed shell (15–25 g) | Soften hard lumps, dispel nodes, clear heat, expel toxins, clear expectoration, relieve cough, removing nebula to improve eyesight; Use to treat scrofula, phlegm and cough, scrofula, goitre, nebula, swelling and ulcer on the body surface. |
| T. hippocastanum 13 | Same as T. alouina, etc. | Same as T. alouina, etc. |
| T. luteostoma 14 | Same as T. gradate | Same as T. gradate |
| T. mutabilis 15 | Same as T. alouina, etc. | Same as T. alouina, etc. |
| T. tuberosa 16 | Same as T. alouina, etc. | Same as T. alouina, etc. |
5.3. The Murex Homeopathic Remedy
6. Conclusions
Acknowledgments
Author Contributions
Conflict of Interest
References
- Newman, D.J.; Cragg, G.M. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.L.; Andrianasolo, E.; McPhail, K.; Flatt, P.; Gerwick, W.H. Marine natural products as anticancer drugs. Mol. Can. Ther. 2005, 4, 333–342. [Google Scholar]
- Benkendorff, K. Molluscan biological and chemical diversity: Secondary metabolites and medicinal resources produced by marine molluscs. Biol. Rev. 2010, 85, 757–775. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2012, 29, 144–222. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K. Chemical diversity in molluscan communities: From natural products to chemical ecology. In Neuroecology and Neuroethology in Molluscs: The interface Between Behaviour and Environment; Di Cosmo, A., Winlow, W., Eds.; Nova Scientific Publishers Inc.: New York, NY, USA, 2014; pp. 13–41. [Google Scholar]
- Madden, T.; Tran, H.T.; Beck, D.; Huie, R.; Newman, R.A.; Pusztai, L.; Wright, J.J.; Abbruzzese, J.L. Novel marine-derived anticancer agents: A phase I clinical, pharmacological, and pharmacodynamic study of dolastatin 10 (nsc 376128) in patients with advanced solid tumors. Clin. Cancer. Res. 2000, 6, 1293–1301. [Google Scholar] [PubMed]
- Chin, Y.; Balunas, M.J.; Chai, H.B.; Kinghorn, D. Drug discovery from natural sources. AAPS J. 2006, 8, E239–E253. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, A.K.; Roy, S.P. Ethno-medicinal uses of some shell fishes by people of Kosi River basin of North-Bihar, India. Stud. Ethno-Med. 2009, 3, 1–4. [Google Scholar]
- Herbert, D.G.; Hamer, M.L.; Mander, M.; Mkhize, N.; Prins, F. Invertebrate animals as a component of the traditional medicine trade in Kwazulu- Natal, South Africa. Afr. Invert. 2003, 44, 327–344. [Google Scholar]
- Yang, X.R. Encyclopedic Reference of Traditional Chinese Medicine; Springer-Verlag Berlin: Heidelberg, Germany, 2003; p. 106. [Google Scholar]
- Gopal, R.; Vijayakumaran, M.; Venkatesam, R.; Kathiroli, S. Marine organisms in Indian medicine and their future prospects. Nat. Prod. Rad. 2008, 7, 139–145. [Google Scholar]
- Meyerhof, M.; Sobhy, G.P. The Abridged Version of “The Book of Simple Drugs”, of Ahmad Ibn Muhammad Al-Ghafiqi by Gregorius Abul-Farag; Al Ettemad Printing Press and Publising House: Cairo, Eygpt, 1932; pp. 224–228. [Google Scholar]
- Lev, E.; Zohar, A. Practical Materia Medica of the Medieval Eastern Mediterranean According to the Cairo Genizah; Brill Online Books and Journals: Leiden, The Netherlands, 2008; Volume 7, p. 664. [Google Scholar]
- Levey, M. Ibn Mäsawaih and his treatise on simple aromatic substances: Studies in the history of Arabic pharmacology I. J. His. Med. All. Sci. 1961, 16, 394–410. [Google Scholar] [CrossRef]
- HPUS. Homœopathic Pharmacopoeia of the United States, 1st ed.; Chicago Duncan Brothers Publishers: Chicago, IL, USA, 1878. [Google Scholar]
- Andlauer, W.; Furst, P. Nutraceuticals: A piece of history, present status and outlook. Food Res. Int. 2002, 35, 171–176. [Google Scholar] [CrossRef]
- Barrow, C.; Shahidi, F. Marine Nutraceuticals and Functional Foods; CRC Press: London, UK, 2007; p. 512. [Google Scholar]
- Piscitelli, S.C.; Burstein, A.H.; Chaitt, D.; Alfaro, R.M.; Falloon, J. Indinavir concentrations and St John’s wort. Lancet 2000, 355, 547–548. [Google Scholar] [CrossRef]
- Straus, S.E. Complementary and alternative medicine: Challenges and opportunities for pharmacology and therapeutic research. Pharmacologist 2000, 42, 74–76. [Google Scholar] [CrossRef]
- Voultsiadou, E. Therapeutic properties and uses of marine invertebrates in the ancient greek world and early byzantium. J. Ethnopharm. 2010, 130, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.S.; Wang, S.G. Hai yang wu ji zhui dong wu yao. In Zhong Hua Hai Ben Cao (Chinese Marine Materia Medica); Shanghai Science and Technology Publishing House: Shanghai, China, 2009; Volume 3, pp. 154–171. [Google Scholar]
- China State Administration Traditional Chinese Materia Medica Editorial Board. Zhong Hua Ben Cao (The Chinese Materia Medica); Shanghai Science and technology Publishing House: Shanghai, China, 1999. [Google Scholar]
- Dunham, C. Murex purpurea. Am. Homeo. Rev. 1864, 4, 306–315. [Google Scholar]
- Benkendorff, K. Natural product research in the Australian marine invertebrate Dicathais orbita. Mar. Drugs 2013, 11, 1370–1398. [Google Scholar] [CrossRef] [PubMed]
- Dolashka, P.; Voelter, W. Antiviral activity of hemocyanins. Invertebr. Surviv. J. 2013, 10, 120–127. [Google Scholar]
- Barco, A.; Claremont, M.; Reid, D.G.; Houart, R.; Bouchet, P.; Williams, S.T.; Cruaud, C.; Couloux, A.; Oliverio, M. A molecular phylogenetic framework for the Muricidae, a diverse family of carnivorous gastropods. Mol. Phylogenet. Evol. 2010, 56, 1025–1039. [Google Scholar] [CrossRef] [PubMed]
- Jennings, S.; Kaiser, M.J.; Reynolds, J.D. Marine Fisheries Ecology; Blackwell Science: Oxford, London, 2001; Volume 11. [Google Scholar]
- Leiva, G.E.; Castilla, J.C. A review of the world marine gastropod fishery: Evolution of catches, management and the Chilean experience. Rev. Fish Biol. Fish. 2001, 11, 283–300. [Google Scholar] [CrossRef]
- Vasconcelos, P.; Carvalho, S.; Castro, M.; Gaspar, M. The artisanal fishery for muricid gastropods (banded murex and purple dye murex) in the Ria Formosa Lagoon (Algrave Coast, Southern Portugal). Sci. Mar. 2008, 72, 287–298. [Google Scholar]
- Woodcock, S.H.; Benkendorff, K. The impact of diet in the growth and proximate composition of juvenile whelks, Dicathais orbita (Gastropoda: Mollusca). Aquaculture 2008, 276, 162–170. [Google Scholar] [CrossRef]
- Mchugh, J. Blattes de byzance in India: Mollusk opercula and the history of perfumery. J. Roy. Asia. Soc. 2013, 23, 53–67. [Google Scholar] [CrossRef]
- Nawata, H. An exported item from Badi on the western Red Sea coast in the eighth century: Historical and ethnographical studies on operculum as incense and perfume. In Ethiopia in Broader Perspective: ICES: International Conference of Ethiopian Studies; Fukui, K., Kurimo, E., Eds.; Shokado Book Sellers: Kyoto, Japan, 1997; Volume 1, pp. 307–325. [Google Scholar]
- Cooksey, C.J. Tyrian purple: 6,6′-dibromoindigo and related compounds. Molecules 2001, 6, 736–769. [Google Scholar] [CrossRef]
- Naegel, L.C.A.; Cooksey, C.J. Tyrian purple from marine muricids, especially from Plicopurpura pansa (Gould, 1853). J. Shellfish Res. 2002, 21, 193–200. [Google Scholar]
- Cooksey, C. Tyrian purple: The first four thousand years. Sci. Prog. 2013, 96, 171–186. [Google Scholar] [CrossRef] [PubMed]
- López-Chávez, F.J.; Ríos-Chávez, P.; Oymam, K. Brominated precursors of Tyrian purple (C.I. Natural violet 1) from Plicopupura pansa, Plicopurpura columellaris and Plicopurpura patula. Dye Pigment. 2009, 83, 7–13. [Google Scholar] [CrossRef]
- Baker, J.T. Tyrian purple: An ancient dye, a modern problem. Endeavour 1974, 33, 11–17. [Google Scholar] [CrossRef]
- Jensen, L.B. Royal purple of Tyre. J. Near East. Stud. 1963, 22, 104–118. [Google Scholar] [CrossRef]
- Allan, J.K. Tyrian purple: An ancient industry. Aust Mus. Mag. 1934, 5, 147–151. [Google Scholar]
- Hutchinson, R.W. Prehistoric Crete; Cox & Wyman Ltd: London, UK, 1962; pp. 238–239. [Google Scholar]
- McGovern, P.E.; Michel, R.H. Royal purple dye: The chemical reconstruction of the ancient Mediterranean industry. Am. Chem. Soc. 1990, 23, 152–158. [Google Scholar] [CrossRef]
- Naegel, L.C.A.; Alvarez, J.I.M. Biological and chemical properties of the secretion from the hypobranchial gland of the purple snail Plicopurpura pansa (Gould 1853). J. Shellfish Res. 2005, 24, 421–428. [Google Scholar]
- Sterman, B.; Taubes-Sterman, J. The Rarest Blue; Lyons Press: Guilford, CO, USA, 2012. [Google Scholar]
- Cooksey, C.J. Marine indirubins. In Indirubin, the Red Shade of Indigo; Meijer, L., Guyard, N., Skaltsounis, L., Eisenbrand, G., Eds.; Progress in Life Series: Roscoff, France, 2006; pp. 23–30. [Google Scholar]
- Nawata, H. Coastal resource use by camel pastoralists: A case study of gathering and fishing activities among the Beja in Eastern Sudan. Nilo-Ethiop. Stud. 2001, 7, 23–43. [Google Scholar]
- Pulak, C. The Uluburun shipwreck and late bronze age trade. In Beyond Babylon: Art, Trade, and Diplomacy in the Second Millennium B.C.; Aruz, J., Bensel, K., Evans, J., Eds.; Metropolitan Museum of Art: New York, NY, USA, 2008; pp. 289–375. [Google Scholar]
- Friedlander, P.; Bruckner, S.; Deutsch, G. Bromo-and methoxy derivatives of indigo. Justus Liebigs Annalen der Chemie 1906, 388, 23–49. [Google Scholar] [CrossRef]
- Hamburger, M. Isatis tinctoria—From the rediscovery of an ancient medicinal plant towards a novel anti-inflammatory phytopharmaceutical. Phytochem. Rev. 2002, 1, 333–344. [Google Scholar] [CrossRef]
- Lim, H.; Chung, E.; Kin, J.-C.; Choi, G.; Jang, K.; Chung, Y.; Cho, Y.; Lee, S.-W. Characterization of a forest soil metagenome clone that confers indirubin and indigo production on Escherichia coli. Appl. Envi. Micro. 2005, 71, 7768–7777. [Google Scholar] [CrossRef] [PubMed]
- Westley, C.B.; Vine, K.L.; Benkendorff, K. A proposed functional role for indole derivatives in reproduction and defence of the Muricidae (Neogastropoda: Mollusca). In Indirubin, the Red Shade of Indigo; Meijer, L., Guyard, N., Skaltsounis, L., Eisenbrand, G., Eds.; Life in Progress Editions: Roscoff, France, 2006; pp. 31–44. [Google Scholar]
- Fouquet, H.; Bielig, H.J. Biological precursors and genesis of Tyrian purple. Angew. Chem. 1971, 10, 816–817. [Google Scholar] [CrossRef]
- Baker, J.T.; Duke, C.C. Isolation from the hypobranchial glands of marine molluscs of 6-bromo-2,2-dimethylthioindolin-3-one and 6-bromo-2-methylthioindoleninone as alternative precursors to Tyrian purple. Tetrahedron Lett. 1973, 14, 2481–2482. [Google Scholar] [CrossRef]
- Baker, J.; Duke, C. Isolation of choline and choline ester salts of tyrindoxyl sulphate from the marine molluscs Dicathais orbita and Mancinella keineri. Tetrahedron Lett. 1976, 15, 1233–1234. [Google Scholar] [CrossRef]
- Baker, J.; Duke, C. Chemistry of the indoleninones. Ii. Isolation from the hypobranchial glands of marine molluscs of 6-bromo-2,2-dimethylthioindolin-3-one and 6-bromo-2-methylthioindoleninone as alternative precursors to Tyrian purple. Aust. J. Chem. 1973, 26, 2153–2157. [Google Scholar] [CrossRef]
- Baker, J.T.; Duke, C.C. Precursors of tyrian purple. Food-Drugs Sea Pro. 1974, 1976, 345–353. [Google Scholar]
- Baker, J.; Sutherland, M. Pigments of marine animals VIII. Precursors of 6,6′-dibromoindigotin (tyrian purple) from the mollusc Dicathais orbita (Gmelin). Tetrahedron Lett. 1968, 1, 43–46. [Google Scholar] [CrossRef]
- Benkendorff, K.; Bremner, J.B.; Davis, A.R. Tyrian purple precursors in the egg masses of the Australian muricid, Dicathais orbita: A possible defensive role. J. Chem. Ecol. 2000, 26, 1037–1050. [Google Scholar] [CrossRef]
- Palma, H.; Paredes, J.; Cristi, E. 6,6′-Dibromoindigotin en capsulas de embriones di Concholepas concholepas (Bruguiere, 1789). Medio Ambient. 1999, 11, 93–95. [Google Scholar]
- Benkendorff, K.; Westley, C.B.; Gallardo, C.S. Observations on the production of purple pigments in the egg capsules, hypobranchial and reproductive glands from seven species of Muricidae (Gastropoda: Mollusca). Invert. Reprod. Dev. 2004, 46, 93–102. [Google Scholar] [CrossRef]
- Westley, C.; Benkendorff, K. Sex-specific tyrian purple genesis: Precursor and pigment distribution in the reproductive system of the marine mollusc, Dicathais orbita. J. Chem. Ecol. 2008, 34, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K.; Bremner, J.; Davis, A. Indole derivatives from the egg masses of muricid molluscs. Molecules 2001, 6, 70–78. [Google Scholar] [CrossRef]
- Benkendorff, K.; Pillai, R.; Bremner, J.B. 2,4,5-tribromo-1 h-imidazole in the egg masses of three muricid molluscs. Nat. Prod. Res. 2004, 18, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Ochi, N.; Kataoka, K.; Ariki, S.; Iwatsuki, C.; Kodama, M.; Fukuyama, Y. Antioxidative bromoindole derivatives from the mid-intestinal gland of the muricid gastropod Drupella fragum. J. Nat. Prod. 1998, 61, 1043–1045. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, F.; Iwatsuki, C.; Kodama, M.; Ochi, M.; Kataoka, K.; Shibata, K. Antimicrobial indolequinones from the mid-intestinal gland of the muricid gastropod Drupella fragum. Tetrahedron 1998, 54, 10007–10016. [Google Scholar] [CrossRef]
- Gul, W.; Hamann, M.T. Indole alkaloid marine natural products: An established source of cancer drug leads with considerable promise for hte control of parasitic, neurological and other diseases. Life Sci. 2005, 78, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Vine, K.L.; Matesic, L.; Locke, J.M.; Ranson, M.; Skropeta, D. Cytotoxic and anticancer activities of isatin and its derivatives: A comprehensive review from 2000–2008. Anticancer Agents Med. Chem. 2009, 9, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Esmaeelian, B.; Benkendorff, K.; Johnston, M.; Abbott, C. Purified brominated indole derivatives from Dicathais orbita induce apoptosis and cell cycle arrest in colorectal cancer cell lines. Mar. Drugs 2013, 11, 3802–3822. [Google Scholar] [CrossRef] [PubMed]
- Edwards, V.; Benkendorff, K.; Young, F. Marine compounds selectively induce apoptosis in female reproductive cancer cells but not in primary-derived human reproductive granulosa cells. Mar. Drugs 2012, 10, 64–83. [Google Scholar] [CrossRef] [PubMed]
- Rudd, D.; Benkendorff, K. Supercritical CO2 extraction of bioactive Tyrian purple precursors from the hypobranchial gland of a marine gastropod. J. Supercrit. Fluids 2014, 94, 1–7. [Google Scholar] [CrossRef]
- Ronci, M.; Rudd, D.; Guinan, T.; Benkendorff, K.; Voelcker, N. Mass spectrometry imaging on porous silicon: Investigating the distribution of bioactives in marine mollusc tissues. Anal. Chem. 2012, 84, 8996–9001. [Google Scholar] [CrossRef] [PubMed]
- Rudd, D.; Benkendorff, K.; Voelcker, N.H. Solvent separating secondary metabolites directly from biosynthetic tissue for surface-assisted laser desorption ionisation mass spectrometry. Mar. Drugs 2015, 13, 1410–1431. [Google Scholar] [CrossRef] [PubMed]
- Roseghini, M.; Erspamer, V.; Ramorino, L.; Gutierrez, J.E. Choline esters, their precursors and metabolites in the hypobranchial gland of prosobranchiate molluscs, Concholepas concholepas and Thais chocolata. Eur. J. Biochem. FEBS 1970, 12, 468–473. [Google Scholar] [CrossRef]
- Roseghini, M.; Severini, C.; Erspamer, G.F.; Erspamer, V. Choline esters and biogenic amines in the hypobranchial gland of 55 molluscan species of the neogastropod Muricoidea superfamily. Toxicon 1996, 34, 33–55. [Google Scholar] [CrossRef]
- Erspamer, V.; Benati, O. Identification of murexine as beta-(imidazolyl-(4))-acrylcholine. Science 1953, 117, 161–162. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.A.; Deriemer, K.; Roberts, T.E.; Rushton, R.; Boothe, P.; Mosher, H.S.; Fuhrman, F.A. Choline esters in marine gastropods Nucella emarginata and Acanthina spirata—New choline ester, tentatively identified as N-methylmurexine. General Pharmacol. 1974, 5, 191–198. [Google Scholar] [CrossRef]
- Shiomi, K.; Ishii, M.; Shimakura, K.; Nagashima, Y.; Chino, M. Tigloylcholine: A new choline ester toxin from the hypobranchial gland of two species of muricid gastropods (Thais clavigera and Thais bronni). Toxicon 1998, 36, 795–798. [Google Scholar] [CrossRef]
- Duke, C.C.; Eichholzer, J.V.; Macleod, J.K. N-methylmurexine—Naturally occuring marine compound. Tetrahedron Lett. 1978, 5047–5048. [Google Scholar] [CrossRef]
- Ramasamy, M.S.; Murugan, A. Potential antimicrobial activity of marine molluscs from Tuticorin, southeast coast of India against 40 biofilm bacteria. J. Shellfish Res. 2005, 24, 243. [Google Scholar]
- D’Armas, H.; Yáñez, D.; Reyes, D.; Salazar, G. Composición de ácidos grasos de los caracoles marinos Phyllonotus pomum y Chicoreus brevifrons (gastropoda: Muricidae). Rev. Biol. Trop. 2010, 58, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Zarai, Z.; Frikha, F.; Balti, R.; Miled, N.; Gargouri, Y.; Mejdoub, H. Nutrient composition of the marine snail (Hexaplex trunculus) from the Tunisian Mediterranean coasts. J. Sci. Food Agric. 2011, 91, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K.; Davis, A.R.; Rogers, C.N.; Bremner, J.B. Free fatty acids and sterols in the benthic spawn of aquatic molluscs, and their associated antimicrobial properties. J. Exp. Mar. Biol. Ecol. 2005, 316, 29–44. [Google Scholar] [CrossRef]
- Benkendorff, K.; Davis, A.R.; Bremner, J.B. Chemical defense in the egg masses of benthic invertebrates: An assessment of antibacterial activity in 39 mollusks and 4 polychaetes. J. Invert. Path 2001, 78, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K. Bioactive Molluscan Resources and Their Conservation: Biological and Chemical Studies on the Egg Masses of Marine Molluscs. Ph.D. Thesis, The University of Wollongong, Wollongong, Australia, 1999. [Google Scholar]
- Dolashka, P.; Moshtanska, V.; Borisova, V.; Dolashki, A.; Stevanovic, S.; Dimanov, T.; Voelter, W. Antimicrobial proline-rich peptides from the hemolymph of marine snail Rapana venosa. Peptides 2011, 32, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Dolashka, P.; Nesterova, N.; Zagorodnya, S.; Dolashki, A.; Baranova, G.; Golovan, A.W.V. Antiviral activity of hemocyanins Rapana venosa and its isoforms against Epstein-Barr virus. Glob. J. Pharm. 2014, 8, 206–212. [Google Scholar]
- Velkova, L.; Todorov, D.; Dimitrova, I.; Shishkov, S.; Van Beeumen, J.; Dolashka-Angelova, P. Rapana venosa hemocyanin with antiviral activity. Biotech. Biotech. Equip. 2009, 23, 606–610. [Google Scholar] [CrossRef]
- WoRMS Editorial Board. World Register of Marine Species. Available online: http://www.marinespecies.org (accessed on 24 Apirl 2015).
- Ivanov, M.; Todorovska, E.; Radkova, M.; Georgiev, O.; Dolashki, A.; Dolashka, P. Molecular cloning, characterization and phylogenetic analysis of an actin gene from the marine mollusk Rapana venosa. Int. J. Curr. Microbiol. App. Sci. 2015, 42, 687–700. [Google Scholar]
- Badiu, D.L.; Balu, A.M.; Barbes, L.; Luque, R.; Nita, R.; Radu, M.; Tanase, E.; Rosoiu, N. Physico-chemical characterisation of lipids from Mytilus galloprovincialis (L.) and Rapana venosa and their healing properties on skin burns. Lipids 2008, 43, 829–841. [Google Scholar] [CrossRef] [PubMed]
- Badiu, D.L.; Luque, R.; Dumitrescu, E.; Craciun, A.; Dinca, D. Amino acids from Mytilus galloprovincialis (L.) and Rapana venosa molluscs accelerate skin wounds healing via enhancement of dermal and epidermal neoformation. Protein J. 2010, 29, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.; Fuentes, A.; Del Campo, M.; Manubens, A.; Nova, E.; Oliva, H.; Faunes, F.; Valenzuela, M.; Campos-Vallette, M.; Aliaga, A.; et al. Immunodominant role of CCHA subunit of Concholepas hemocyanin is associated with unique biochemical properties. Int. Immunopharmacol. 2009, 9, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Tchorbanov, A.; Idakieva, K.; Mihaylova, N.; Doumanova, L. Modulation of the immune response using Rapana thomasiana hemocyanin. Int. Immunopharmacol. 2008, 8, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Cantillana, P.; Inestrosa, N.C. Presence of a heparin-binding growth factor in Concholepas concholepas Bruguiere (Mollusca: Gastropoda; Muricidae). J. Exp. Mar. Bio. Ecol. 1991, 171, 239–250. [Google Scholar] [CrossRef]
- Man, Y.; Wang, Y.-X.; ZHu, S.-Y.; Zhao, D.; Hu, F.; Lu, J.-Y. Indirubin inhibits ATP-induced phagocytosis attenuation, ROS production and cell death of macrophages. Acta Pharm. Sin. 2012, 47, 45–50. [Google Scholar]
- Kim, J.K.; Park, G.M. Indirubin-3-monoxime exhibits anti-inflammatory properties by down-regulating NF-kappab and JNK signaling pathways in lipopolysaccharide-treated RAW264.7 cells. Inflamm. Res. 2012, 61, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.-J.; Nam, K.N.; Son, M.-S.; Kang, H.; Hong, J.-W.; Kim, J.W.; Lee, E.H. Indirubin-3′-oxime inhibits inflammatory activation of rat brain microglia. Neurosci. Lett. 2011, 487, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Matheus, M.E.; Flavio de Almeida, V.; Garden, S.; Pinto, J.; Fernamdes, A.C.; Dias, P. Isatins inhibit cyclooxygenase-2 and inducible nitric oxide synthase in a mouse macrophage cell line. Eur. J. Pharmacol. 2007, 556, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Kitaura, Y.; Ito, F.; Stevens, R.W.; Asai, N. Antiallergy and Antiinflamatory Agents. U.S. Patent 4904685, 27 February 1990. [Google Scholar]
- Stevens, R.W.; Morita, H.; Nakane, M. Indole Derivatives as Antiallergy and Antiinflammatory Agents. U.S. Patent 5290788, 1 March 1994. [Google Scholar]
- Olofsson, K.; Suna, E.; Pelcman, B.; Ozola, V.; Kakvins, I.; Schaal, W. Indoles Useful in the Treatment of Inflammation. U.S. Patent US 2006/0160879, July 2006. [Google Scholar]
- Farina, C.; Gagliardi, S.; Novella, P.A. Indole Derivatives and Their Use for the Treatment of Osteoporosis amongst Other Applications. U.S. Patent 6,787,550 B1, 7 September 2004. [Google Scholar]
- Yazback, R.; Lindsay, R.; Abbott, C.A.; Benkendorff, K.; Howarth, G.S. Combined effects of muricid extract and 5-fluorouracil on intestinal toxicity in rats. Evid.-Based Complement. Altern. Med. 2015, 2015, 170859. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K.; McIver, C.M.; Abbott, C.A. Bioactivity of the murex homeopathic remedy and of extracts from an Australian muricid mollusc against human cancer cells. Evid.-Based Complement. Altern. Med. 2011, 2011, 12. [Google Scholar] [CrossRef] [PubMed]
- Vine, K.; Locke, J.; Ranson, M.; Benkendorff, K.; Pyne, S.; Bremner, J. In vitro cytotoxicity evaluation of some substituted isatin derivatives. Bioorg. Med. Chem. 2007, 15, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Edwards, V.; Young, F.; Benkendorff, K. An in vitro high throughput assay for screening reproductive and toxic effects of anticancer compounds. Biotech. Appl. Biochem. 2014, 61, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Meijer, L.; Shearer, J.; Bettayeb, K.; Ferandin, Y. Diversity of intracellular mechanisms underlying the anti-tumor properties of indirubins. In Indirubin, the Red Shade of Indigo; Meijer, L., Guyard, N., Skaltsounis, A.-L., Eisenbrand, G., Eds.; Life in Progress Editions: Roscoff, France, 2006; Chapter 24; pp. 235–246. [Google Scholar]
- Magiatis, P.; Skaltsounis, A.L. From Hexaplex trunculus to new kinase inhibitory indirubins. In Indirubin, the Red shade of Indigo; Meijer, L., Guyard, N., Skaltsounis, A.-L., Eisenbrand, G., Eds.; Life in Progress Editions: Roscoff, France, 2006; pp. 147–156. [Google Scholar]
- Zhang, L.; Fan, X.; Han, L. Antitumor and immune regulation activities of the extracts of some chinese marine invertebrates. Chin. J. Oceanol. Limnol. 2005, 23, 110–117. [Google Scholar]
- Genova-Kalou, P.; Idakieva, K.; Dundarova, D.; Argirova, R.; Alexandrova, R.; Yotovska, K.; Mohmmed, A. Cytotoxic and apoptogenic properties of the hemocyanin derived from the marine mollusk Rapana thomasiana in vitro. Planta Med. 2008, 74, PA319. [Google Scholar] [CrossRef]
- Antonova, O.; Yossifova, L.; Staneva, R.; Stevanovic, S.; Dolashka, P.; Toncheva, D. Changes in the gene expression profile of the bladder cancer cell lines after treatment with Helix lucorum and Rapana venosa hemocyanin. J. Balk. Union Oncol. 2015, 20, 180–187. [Google Scholar]
- Esmaeelian, B.; Abbott, C.; Le Leu, R.; Benkendorff, K. 6-bromoisatin found in muricid mollusc extracts inhibits colon cancer cell proliferation and induces apoptosis, preventing early stage tumor formation in a colorectal cancer rodent model. Mar. Drugs 2014, 12, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Meijer, L.; Skaltsounis, A.L.; Magiatis, P.; Polychronopoulos, P.; Knockaert, M.; Leost, M.; Ryan, X.P.; Vonica, C.A.; Brivanlou, A.; Dajani, R.; et al. GSK-3-selective inhibitors derived from Tyrian purple indirubins. Chem. Biol. 2003, 10, 1255–1266. [Google Scholar] [CrossRef] [PubMed]
- Hoessel, R.; Leclerc, S.; Endicot, J.A.; Nobel, M.E.M.; Lawrie, A.; Tunnah, P.; Leost, M.; Damiens, E.; Marie, D.; Marko, D.; et al. Indirubin, the active constituent of a chinese antileukaemia medicine, inhibits cyclin-dependent kinases. Nat. Cell Biol. 1999, 1, 60–67. [Google Scholar] [PubMed]
- Leclerc, S.; Garnier, M.; Hoessel, R.; Marko, D.; Bibb, J.A.; Snyder, G.L.; Greengard, P.; Jacek Biernati, J.; Wui, Y.; Mandelkowi, E.; et al. Indirubins inhibit glycogen synthase kinase-3b and CDK5/p25, two protein kinases involved in abnormal tau phosphorylation in alzheimer’s disease. J. Biol. Chem. 2001, 276, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Vougogiannopoulou, K.; Skaltsounis, A.L. From Tyrian purple to kinase modulators: Naturally halogenated indirubins and synthetic analogues. Planta Med. 2012, 78, 1515–1528. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Tabata, K.; Hanada, S.; Kanda, Y.; Suzuki, T.; Miyairi, S. Synthesis of methoxy- and bromo-substituted indirubins and their activities on apoptosis induction in human neuroblastoma cells. Bioorg. Med. Chem. Lett. 2011, 21, 5370–5373. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.A.; Liapis, V.; Evdokiou, A.; Constantinou, C.; Magiatis, P.; Skaltsounis, A.L.; Koumas, L.; Costeas, P.A.; Constantinou, A.I. Induction of discrete apoptotic pathways by bromo-substituted indirubin derivatives in invasive breast cancer cells. Biochem. Biophys. Res. Commun. 2012, 425, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Nam, S.; Tian, Y.; Yang, F.; Wu, J.; Wang, Y.; Scuto, A.; Polychronopoulos, P.; Magiatis, P.; Skaltsounis, L.; et al. 6-bromoindirubin-3′-oxime inhibits JAK/STAT3 signaling and induces apoptosis of human melanoma cells. Cancer Res. 2011, 71, 3972–3979. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Moon, M.J.; Lee, S.D.; Choi, S.U.; Han, S.Y.; Kim, Y.C. Indirubin derivatives as potent FLT3 inhibitors with anti-proliferative activity of acute myeloid leukemic cells. Bioorg. Med. Chem. Lett. 2010, 20, 2033–2037. [Google Scholar] [CrossRef] [PubMed]
- Edwards, V.; Young, F.; Benkendorff, K.; Flinders University, Adelaide, Australia. Effects of Murcidae extracts on estrogen-sensitive breast cancer and the steroidogenic pathway. Unpublished work. 2013. [Google Scholar]
- Grubb, G.S.; Fensome, A.; Miller, L.L.; Ullrich, J.W.; Bender, R.H.W.; Zhang, P.; Wrobel, J.E.; Edwards, J.P.; Jones, T.K.; Tegley, C.M.; et al. Cyclic Regimens Utilizing Indoline Derivatives. U.S. Patent 2003/0050288, 13 March 2003. [Google Scholar]
- Westley, C.B.; McIver, C.M.; Abbott, C.A.; Le Leu, R.K.; Benkendorff, K. Enhanced acute apoptotic response to azoxymethane-induced DNA damage in the rodent colonic epithelium by tyrian purple precursors a potential colorectal cancer chemopreventative. Cancer Biol. Ther. 2010, 9, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Esmaeelian, B. In-Vitro and in Vivo Testing of Purified Muricid Mollusc Extract on Colorectal Cancer. Ph.D. Thesis, The Flinders University, Adeliade, SA, Australia, 2014. [Google Scholar]
- Fiebig, H.H.; Schuler, J. In vivo anti-tumour activity of indirubins. In Indirubin, the Red Shade of Indigo; Meijer, L., Guyard, N., Skaltsounis, A.L., Eisenbrand, G., Eds.; Life in Progress Editions: Roscoff, France, 2006. [Google Scholar]
- Wang, R. Effects of Marine Mollusc Extracts on Human Immune Function. Master’s Thesis, Flinders University, Adelaide, Australia, 2009. [Google Scholar]
- Cane, A.; Tournaire, M.-C.; Barritault, D.; Crumeyrolle-Arias, M. The endogenous oxindoles 5-hydroxyoxindole and isatin are antiproliferative and proapoptotic. Biochem. Biophys. Res.Commun. 2000, 276, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Igosheva, N.; Lorz, C.; O’Conner, E.; Glover, V.; Mehmet, H. Isatin, an endogenous monoamine oxidase inhibitor, triggers a dose-and time-dependent switch from apoptosis to necrosis in human neuroblastoma cells. Neurochem. Int. 2005, 47, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Gaboriaud-Kolar, N.; Vougogiannopoulou, K.; Skaltsounis, A.-L. Indirubin derivatives: A patent review (2010-present). Expert Opin. Ther. Pat. 2014, 25, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.; Buettner, R.; Turkson, J.; Kim, D.; Cheng, J.Q.; Muehlbeyer, S.; Hippe, F.; Vatter, S.; Merz, K.H.; Eisenbrand, G.; et al. Indirubin derivatives inhibit STAT3 signaling and induce apoptosis in human cancer cells. PNAS 2005, 102, 5998–6003. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Kim, Y.C.; Kim, S.W.; Lee, S.H.; Min, J.J.; Ahn, S.G.; Yoon, J.H. Antitumor activity of novel indirubin derivatives in rat tumor model. Clin. Cancer Res. 2007, 13, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Karabelas, K.; Lepisto, M.; Sjo, P. Indoles as Protein Kinase Inhibitors. U.S. Patent 6,346,625, 2 February 2002. [Google Scholar]
- Kim, Y.C.; Kim, S.W.; Kim, T.S.; Lee, S.K.; Kim, J.D.; Yoon, J.H. Indirubin Derivatives Having Anticancer Property against Human Cancer Cell Lines. U.S. Patent 7,572,923, 11 August 2009. [Google Scholar]
- Wang, L.; Liu, X.P.; Chen, R. Derivatives of Isoindigo, Indigo and Indirubin and Methods of Treating Cancer. U.S. Patent 6,933,315, 23 August 2005. [Google Scholar]
- Wang, L.; Liu, X.P.; Chen, R. Derivative of Isoindigo and Indirubin for the Treatment of Cancer. U.S. Patent 6,566,341, 20 May 2003. [Google Scholar]
- Carson, D.A.; Leoni, L.M.; Cotton, H.B. Indoles Treatment of Cancer. U.S. Patent 7,151,100, 19 December 2006. [Google Scholar]
- Leung, P.C.K.; Adashi, E.Y. The Ovary, 2nd ed.; Elsevier Academic Press: San Diego, CA, USA, 2004. [Google Scholar]
- Edwards, V. The Effects of Bioactive Compounds from the Marine Mollusc Dicathais Orbita on Human Reproductive Cells and Human Reproductive Cancer Cells. Ph.D. Thesis, The Flinders University, Adelaide, SA, Australia, 2012. [Google Scholar]
- Ohlsson, A.; Cedergreen, N.; Oskarsson, A.; Ullera, E. Mixture effects of imidazole fungicides on cortisol and aldosterone secretion in human adrenocortical H295r cells. Toxicology 2010, 275, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Erspamer, V.; Glasser, A. The pharmacological actions of murexine (urocanylcholine). Br. J. Pharmacol. Chemother. 1957, 12, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Keyl, M.J.; Whittaker, V.P. Some pharmacological properties of murexine (urocanoylcholine). Br. J. Pharmacol. Chemother. 1958, 13, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.K.; Mitra, S.K.; Acharya, S.B. Anxiogenic activity of isatin, a putative biological factor, in rodents. J. Psychopharmacol 1991, 5, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.L. Behavioral effects of isatin on open field activity and immobility in the forced swim test in rats. Physiol. Behav. 1995, 57, 611–613. [Google Scholar] [CrossRef]
- Bhattacharya, S.K.; Chakrabarti, A. Dose-related proconvulsant and anticonvulsant activity of isatin, a putative biological factor, in rats. Indian. J. Exp. Biol. 1998, 36, 118–121. [Google Scholar] [PubMed]
- Ahring, P.K.; Christophersen, P.; Jensen, B.S.; Jorgensen, T.D.; Strobaek, D.; Teuber, L.; Olesen, S.P. Use of Isatin Derivatives as Ion Channel Activating Agents. International Patent WO2000033834, 15 June 2000. [Google Scholar]
- Watjen, F.; Drejer, J.; Jensen, L.H. Isatin Derivatives, Their Preparation and Use. European Patent EP0432648B, 2 August 1995. [Google Scholar]
- Debat, J. Promotion of Analgesic and Sedative Action with 5-Bromoisatin. International Patent 3,659,011, 25 April 1972. [Google Scholar]
- Hey, P. On relationships between structure and nicotine-like stimulant activity in choline esters and ethers. Br. J. Pharmacol. Chemother. 1952, 7, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, A.; Buneeva, O.; Glover, V. Biological targets for isatin and its analogues: Implications for therapy. Biologics 2007, 1, 151–162. [Google Scholar] [PubMed]
- Crumeyrolle-Arias, M.; Medvedev, A.; Cardona, A.; Barritault, D.; Glover, V. In situ imaging of specific binding of [H]isatin in rat brain. J. Neurochem. 2003, 84, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, A.E.; Sandler, M.; Glover, V. Interaction of isatin with type-a natriuretic peptide receptor: Possible mechanism. Life Sci. 1998, 62, 2391–2398. [Google Scholar] [CrossRef]
- Westley, C.B.; Benkendorff, K.; McIver, C.M.; Le Leu, R.K.; Abbott, C.A. Gastrointestinal and hepatotoxicity assessment of an anticancer extract from muricid molluscs. Evid. Based Complement. Altern. Med. 2013, 2013, 837370. [Google Scholar]
- Chahal, C.S. A Study of the Anticancer Properties of the Crude Extract and Synthetic 6-Bromoisatin from Dicathais orbita: An In Vivo Colorectal Cancer Mice Model. Master’s Thesis, Flinders University, Adelaide, Australia, 2014. [Google Scholar]
- Nataraj, K.S.; Rao, J.V.; Jayaveera, K.N. Diuretic Activity of Some Novel Isatin Derivatives. J. Pharm. Res. 2010, 3, 863. [Google Scholar]
- Lev, E. Drugs held and sold by pharmacists of the Jewish community of medieval (11–14th Centuries) Cairo according to lists of Materia Medica found at the Taylor–Schechter Genizah collection, Cambridge. J. Ethnopharmacol. 2007, 110, 275–293. [Google Scholar] [CrossRef] [PubMed]
- Rätsch, C.; Müller-Ebeling, C. The Encyclopedia of Aphrodisiacs: Psychoactive Substances for Use in Sexual Practices; Inner Traditions/Bear & Co: Rochester, VT, USA, 2013. [Google Scholar]
- National Health and Medical Research Council. NHMRC Information Paper: Evidence on the Effectiveness of Homeopathy for Treating Health Conditions; National Health and Medical Research Council: Canberra, Australia, 2015. [Google Scholar]
- World Health Organisation. Saftey Issues in the Preparation of Homeopathic Medicines; WHO Press: Geneva, Switzerland, 2009. [Google Scholar]
- HPUS. Monograph 6116 Murex Purpurea in Homoepathic Pharmacopoeia of the United States. Available online: www.hpus.com/online_database/view_monograph_action.php?id=6116 (accessed on 11 September 2012).
- Zacharia, G.; Government Homeopathic Hospital, Cochin, India. Personal communication, 2012.
- Lee, A. Homeopathic Mind Maps Remedies of the Animal Kingdom, 4th ed.; Moozoonsil Publishing: Auckland, Australia, 2011. [Google Scholar]
- Hering, C. Guiding Symptoms of our Materia Medica; American Homeopathic Publication Society: Philadelphia, PA, USA, 1890. [Google Scholar]
- Lesser, O. The molluscs: Murex and Sepia. Br. Homeopath. J. 1990, 79, 138–146. [Google Scholar] [CrossRef]
- Vermeulen, F. Prisma; Emryss bv: Haarlem, The Netherlands, 2002. [Google Scholar]
- Murphy, R. Homeopathic Clinical Repertory, 3rd ed.; Lotus Health Institute: Blacksburg, VA, USA, 2005. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benkendorff, K.; Rudd, D.; Nongmaithem, B.D.; Liu, L.; Young, F.; Edwards, V.; Avila, C.; Abbott, C.A. Are the Traditional Medical Uses of Muricidae Molluscs Substantiated by Their Pharmacological Properties and Bioactive Compounds? Mar. Drugs 2015, 13, 5237-5275. https://doi.org/10.3390/md13085237
Benkendorff K, Rudd D, Nongmaithem BD, Liu L, Young F, Edwards V, Avila C, Abbott CA. Are the Traditional Medical Uses of Muricidae Molluscs Substantiated by Their Pharmacological Properties and Bioactive Compounds? Marine Drugs. 2015; 13(8):5237-5275. https://doi.org/10.3390/md13085237
Chicago/Turabian StyleBenkendorff, Kirsten, David Rudd, Bijayalakshmi Devi Nongmaithem, Lei Liu, Fiona Young, Vicki Edwards, Cathy Avila, and Catherine A. Abbott. 2015. "Are the Traditional Medical Uses of Muricidae Molluscs Substantiated by Their Pharmacological Properties and Bioactive Compounds?" Marine Drugs 13, no. 8: 5237-5275. https://doi.org/10.3390/md13085237