
Dangerous Relations in the Arctic Marine Food Web: Interactions between Toxin Producing Pseudo-nitzschia Diatoms and Calanus Copepodites

Abstract
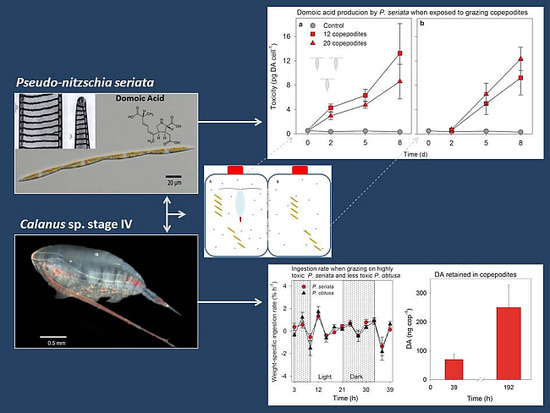
:
1. Introduction
2. Results
2.1. Temporal Grazing Experiment
2.1.1. Toxicity

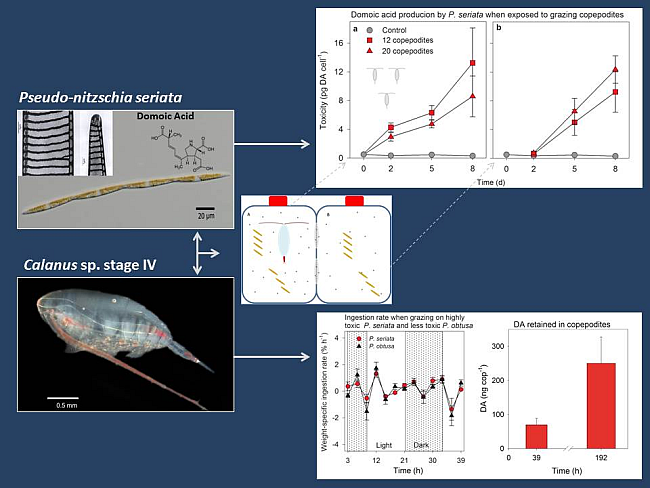{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pseudo-nitzschia | Calanus | |||||||
|---|---|---|---|---|---|---|---|---|
| Number of Cells | DA Cell Quota | Ingested DA | Retained DA | |||||
| (Cells mL−1) | (pg DA Cell−1) | (ng DA cop−1) | (ng DA cop−1) | (µg DA g DW cop−1) | % of Ingested DA | |||
| Initial | End | Initial | End | |||||
| Controls | ||||||||
| P. seriata | 3964 ± 40 | 5048 ± 102 | 0.1 ± 0.0 | 0.1 ± 0.0 | ||||
| P. obtusa | 5061 ± 32 | 8907 ± 683 | <LOD | <LOD | ||||
| P. seriata + P. obtusa | 7144 ± 103 | 8617 ± 122 | 0.1 ± 0.0 | 0.2 ± 0.0 | ||||
| Treatments | ||||||||
| P. seriata + copepodites | 3951 ± 52 | 1901 ± 187 | 0.1 ± 0.0 | 2.6 ± 0.6 *, ** | 530 ± 21 | 70 ± 18 | 410 ± 112 | 13 ± 4 |
| P. obtusa + copepodites | 5951 ± 285 | 4988 ± 570 | <LOD | 0.2 ± 0.1 *, ** | 137 ± 37 | 8 ± 2 | 48 ± 10 | 6 ± 2 |
| P. seriata + P. obtusa + copepodites | 7033 ± 97 | 4262 ± 249 | 0.1 ± 0.0 | 3.7 ± 0.6 *, ** | 275 ± 70 | 42 ± 8 | 246 ± 51 | 16 ± 4 |
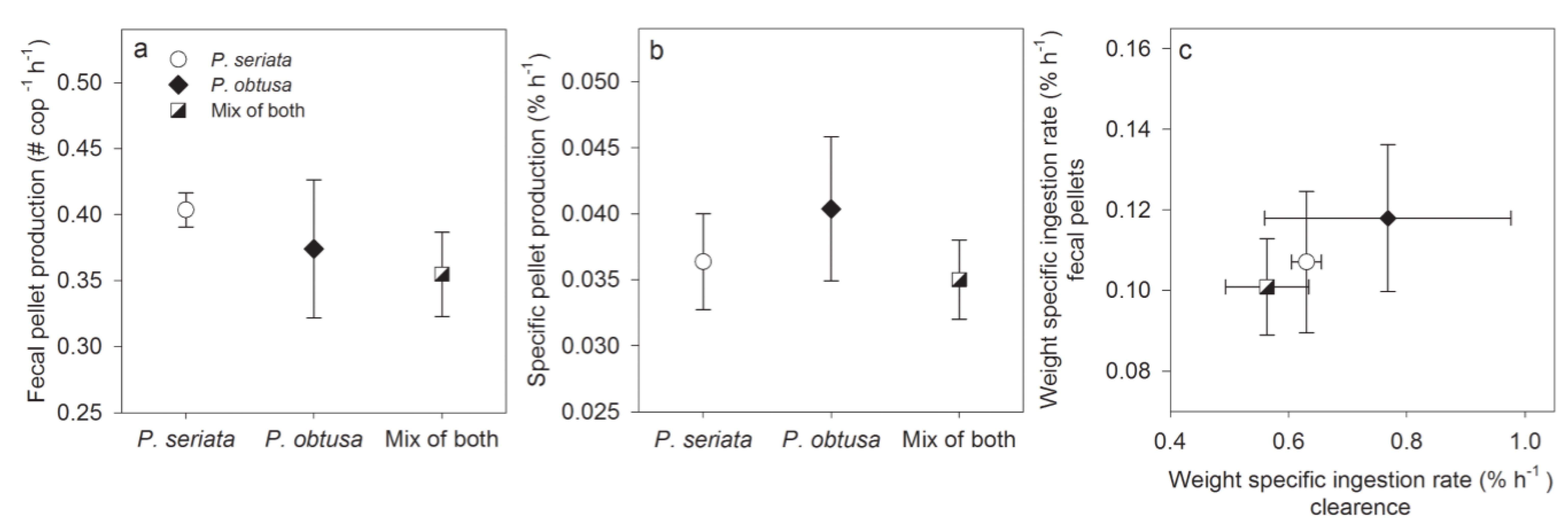
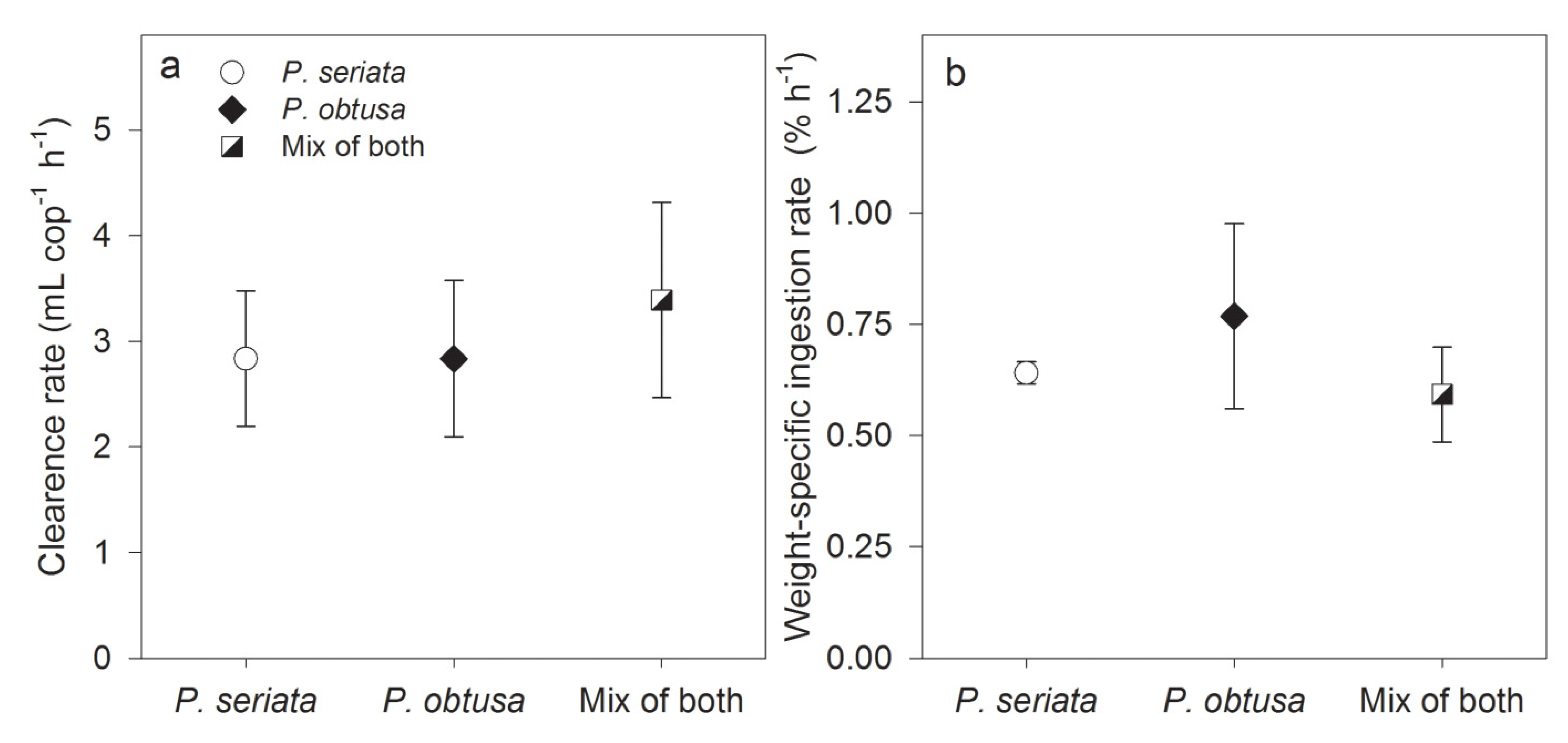
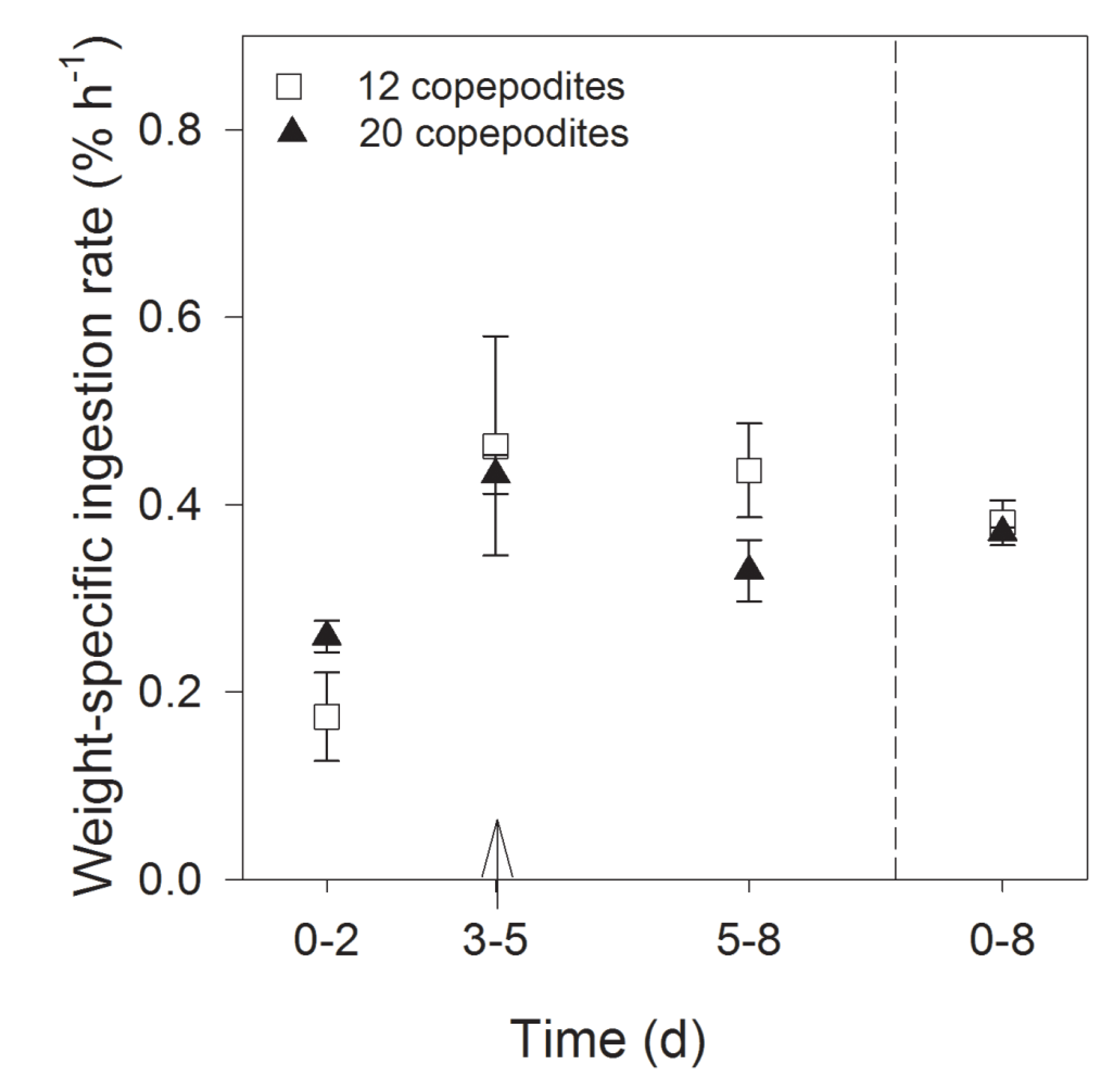
2.1.2. Grazing and Fecal Pellet Production
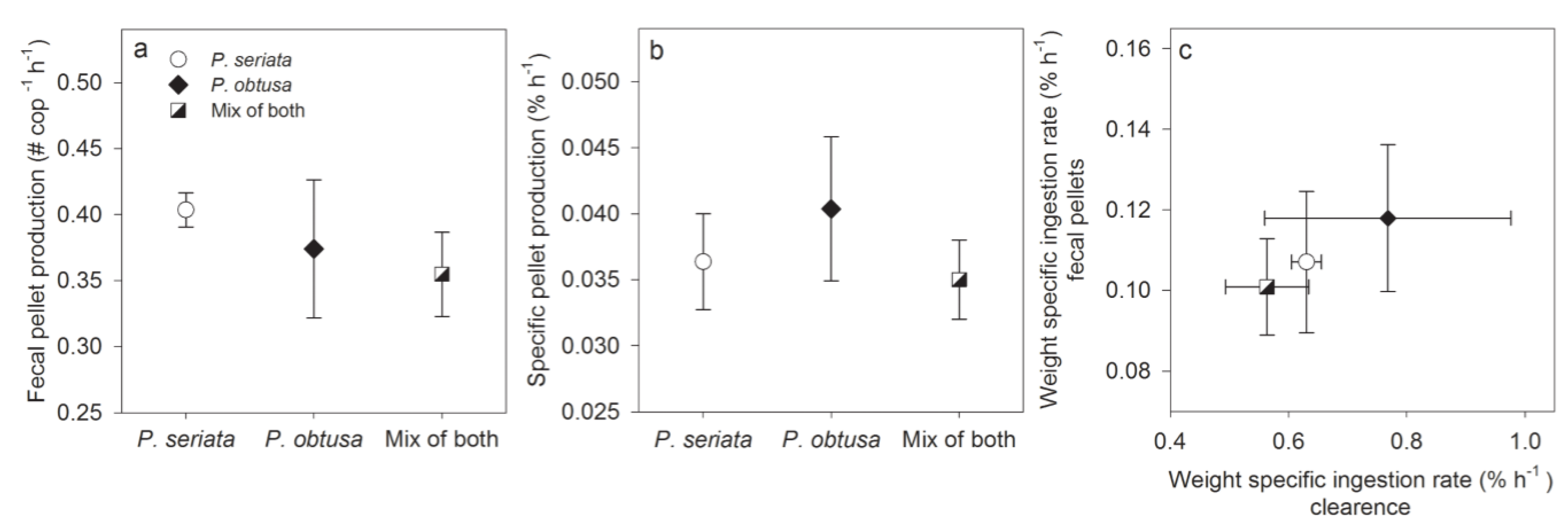
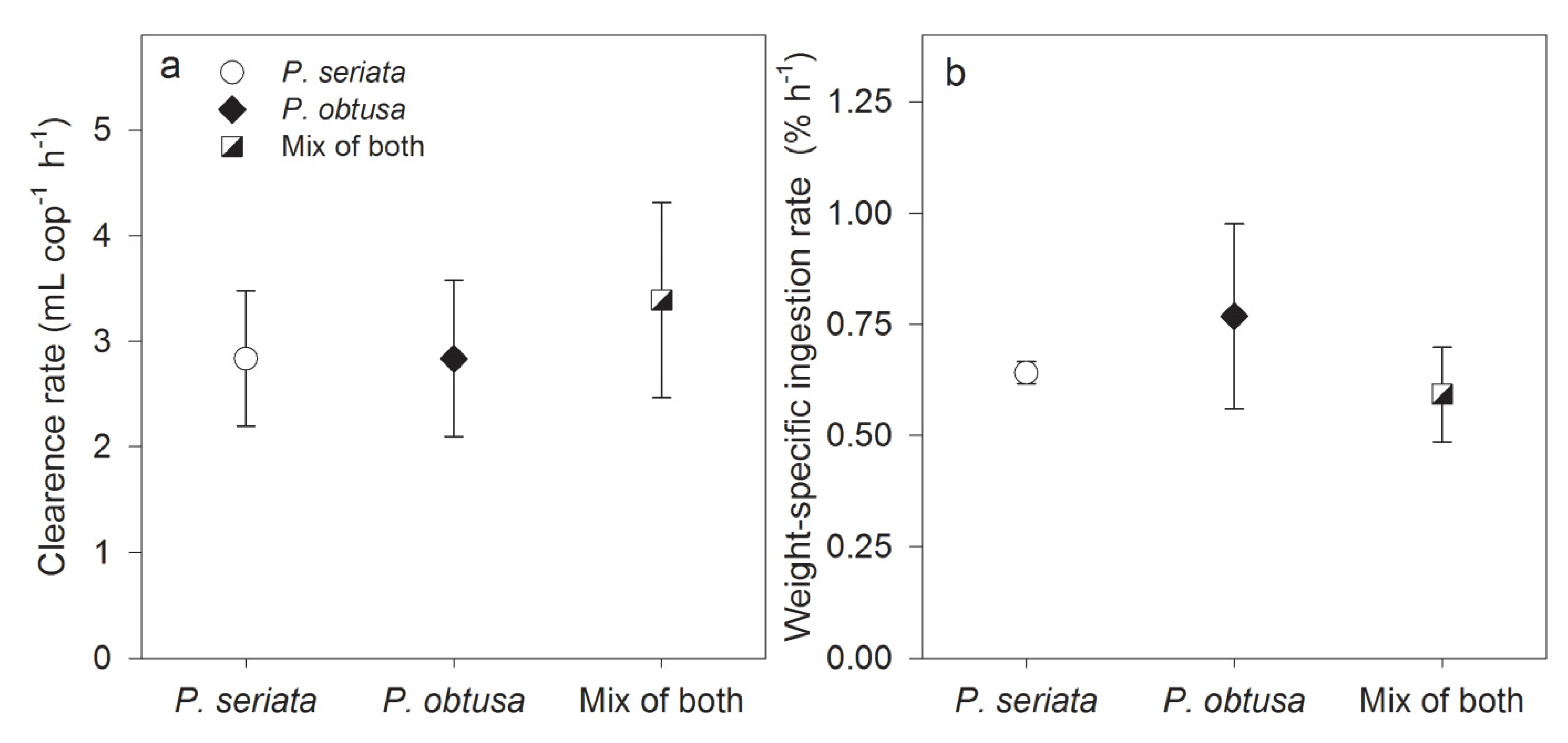
2.1.3. Growth Conditions
2.2. Induction Experiments
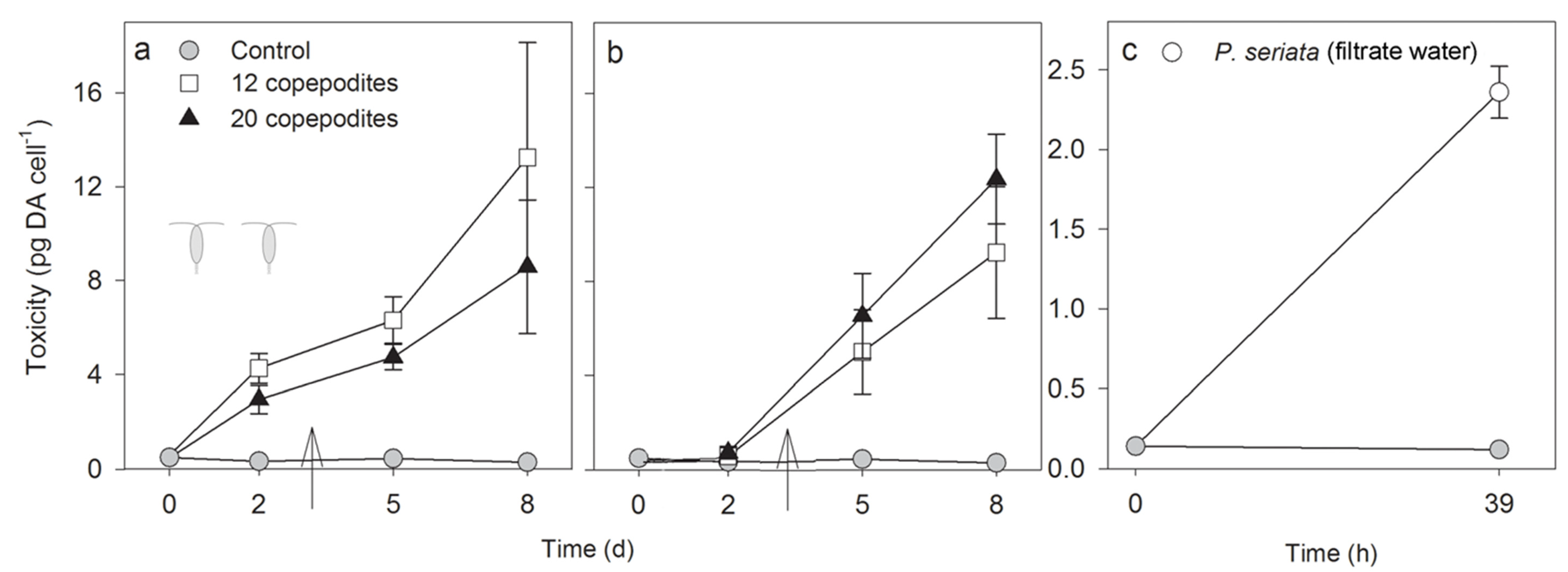
2.2.1. Toxicity
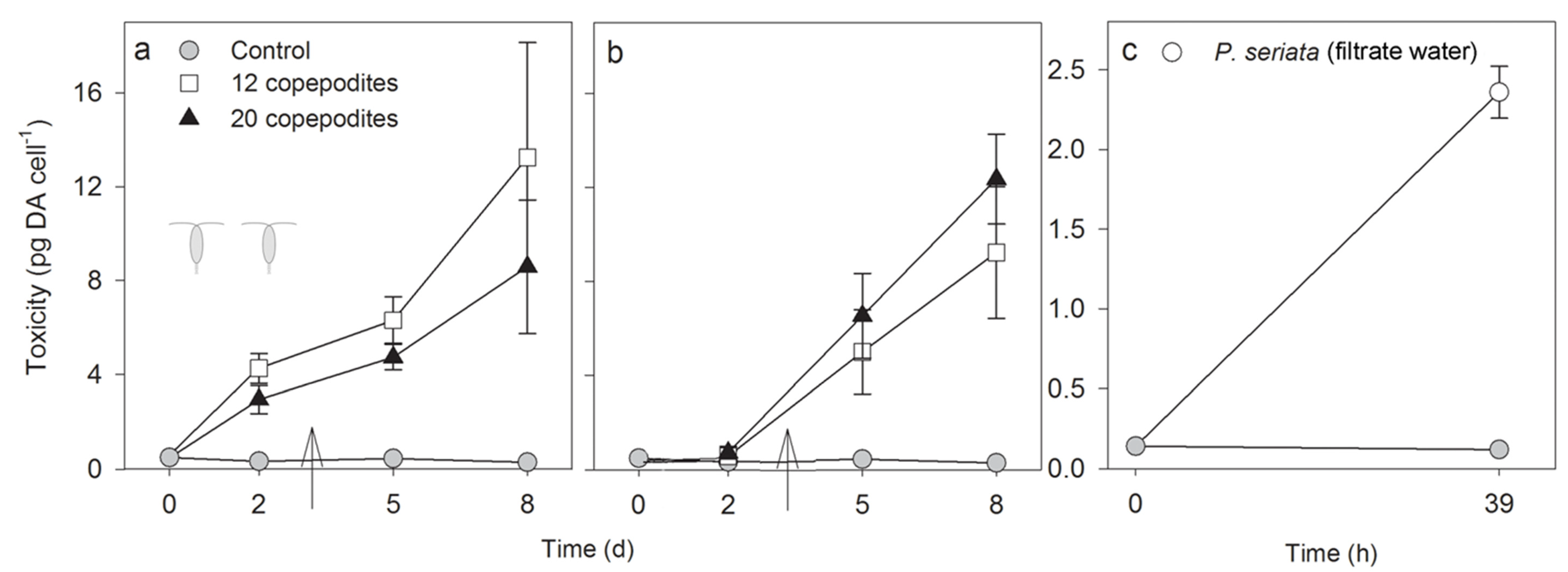
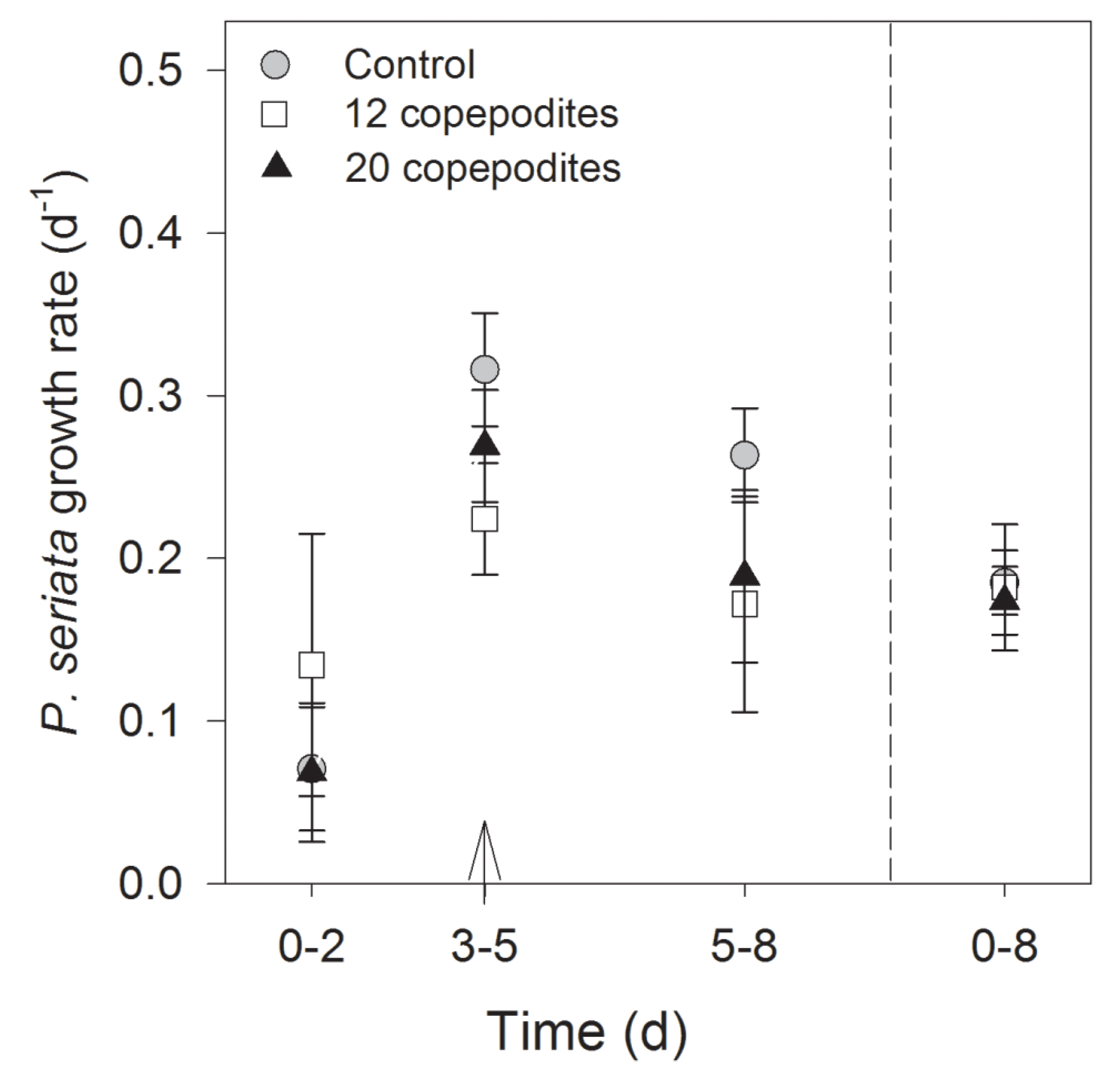
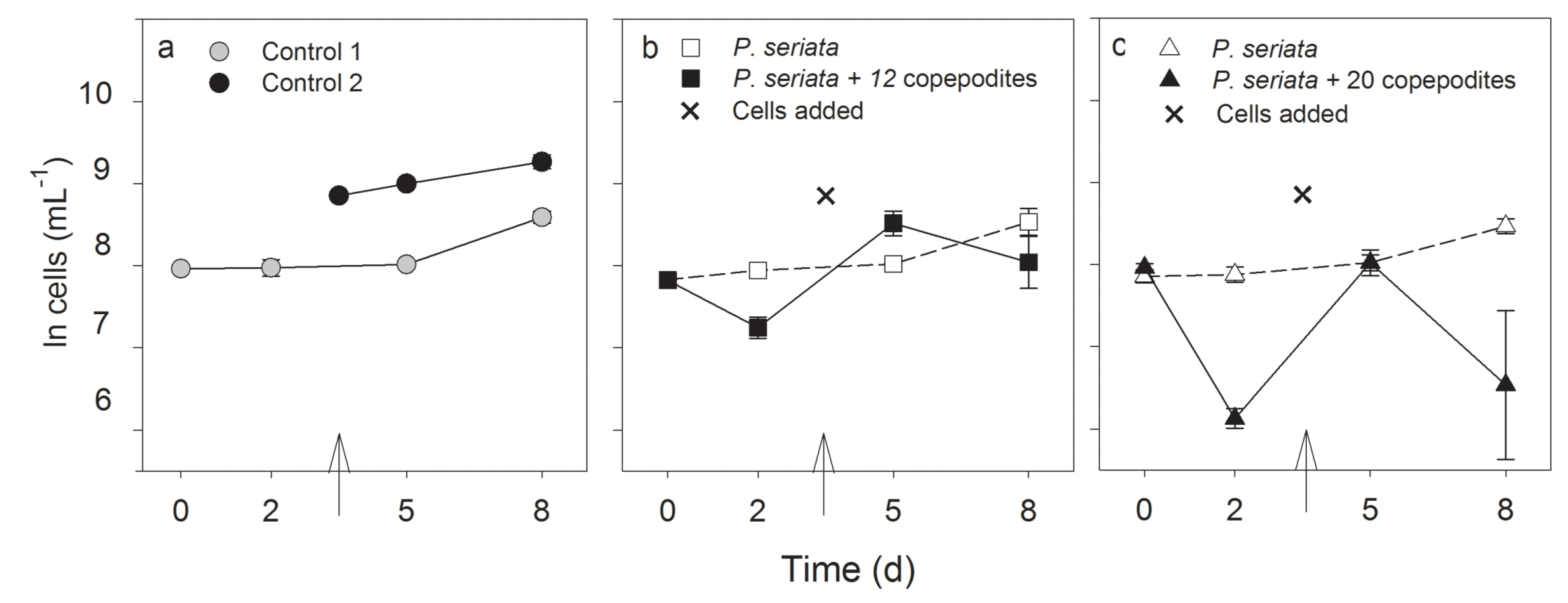
2.2.2. Growth Rate, Cell Density and Growth Condition
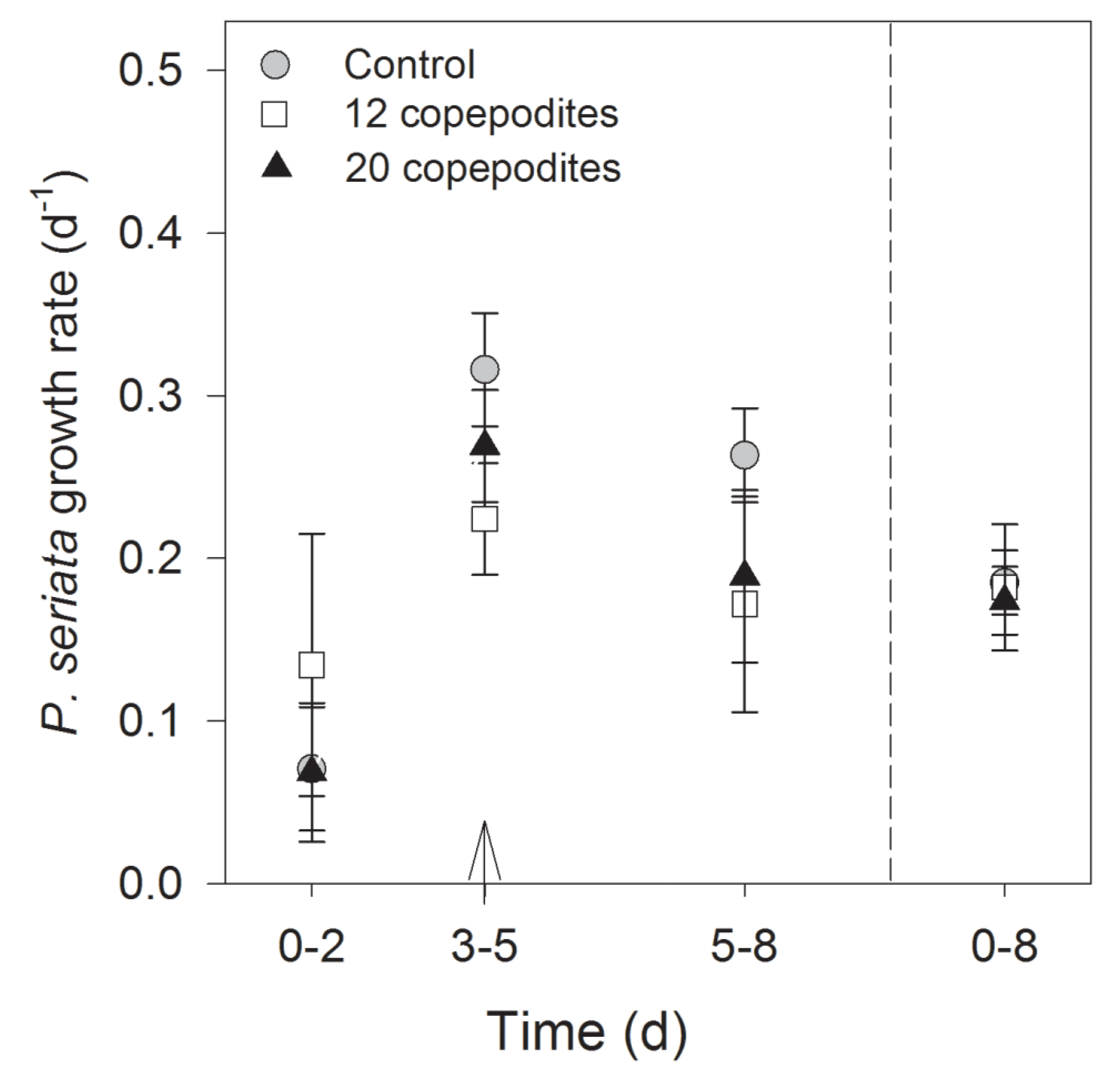
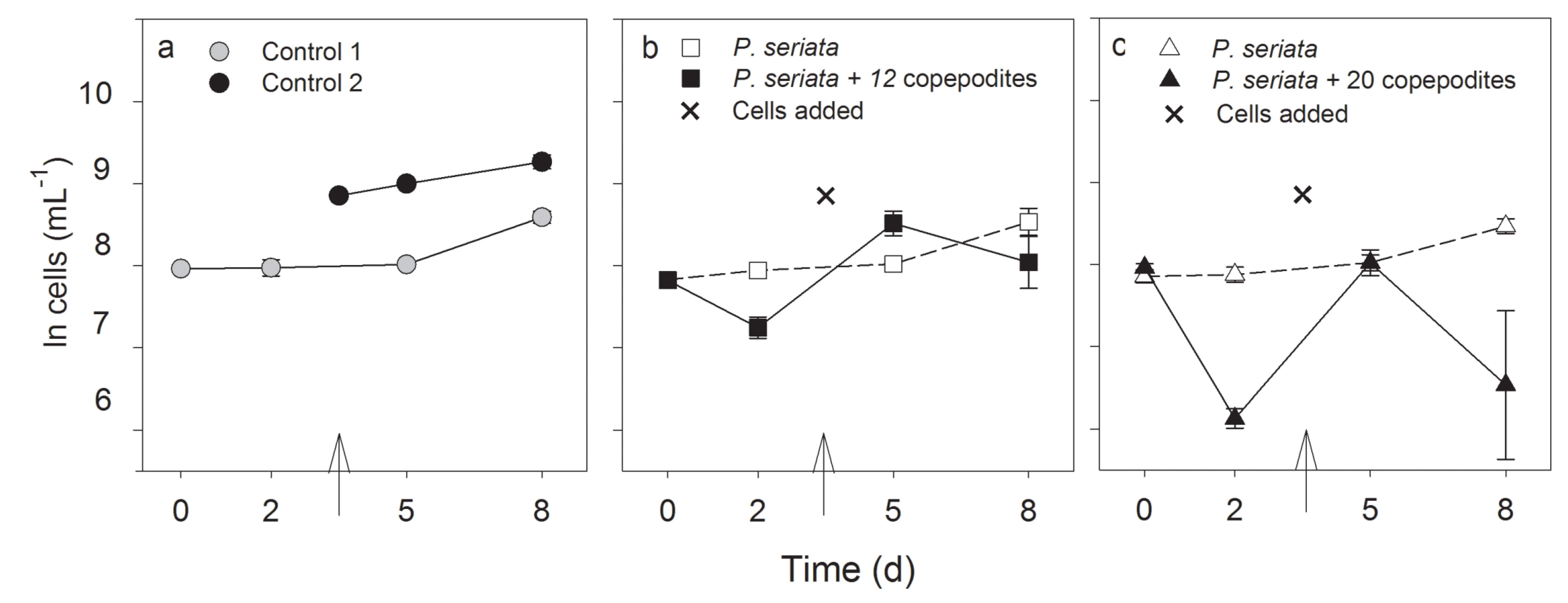
| Time (day)/(h) | Treatment | Si(OH)4 (µmol L−1) | PO43− (µmol L−1) | NH4+ (µmol L−1) | NO3− (µmol L−1) | pH |
|---|---|---|---|---|---|---|
| 0 | Initial | 5.7 ± 0.6 | 5.6 ± 0.6 | 38.3 ± 1.6 | 131.6 ± 5.6 | 8.10 |
| 8 | Control | 0.0 * | 3.5 ± 0.3 * | 25.6 ± 1.0 * | 99.3 ± 6.3 | 8.12 ± 0.02 |
| 8 | Flask A, n = 12 | 1.3 ± 1.0 * | 6.6 ± 1.2 ** | 28.8 ± 2.3 | 182.6 ± 20.7 ** | 8.16 ± 0.02 |
| 8 | Flask A, n = 20 | 3.6 ± 0.7 *, ** | 7.1 ± 0.5 *, ** | 33.9 ± 0.7 ** | 191.2 ± 4.2 ** | 8.12 ± 0.02 |
| 8 | Flask B, n = 12 | 0.4 ± 0.7 * | 5.5 ± 1.1 | 25.9 ± 1.4 | 164.2 ± 17.7 ** | 8.18 ± 0.00 |
| 8 | Flask B, n = 20 | 0.4 ± 0.3 * | 5.1 ± 0.1 | 27.9 ± 1.5 | 161.4 ± 10.5 ** | 8.11 ± 0.01 |
| 0 | Initial | 6.7 ± 0.6 | 7.0 ± 0.2 | 38.8 ± 1.7 | 164.8 ± 12.5 | 8.06 ± 0.00 |
| 39 h | Control | 3.6 ± 0.3 * | 7.4 ± 0.1 * | 39.9 ± 1.0 | 177.7 ± 1.0 | 8.09 ± 0.01 |
| 39 h | End | 3.7 ± 0.31 * | 8.0 ± 0.4 * | 43.3 ± 0.7 *, ** | 197.7 ± 1.1 *, ** | 8.10 ± 0.01 |
2.2.3. Grazing and Retained Domoic Acid
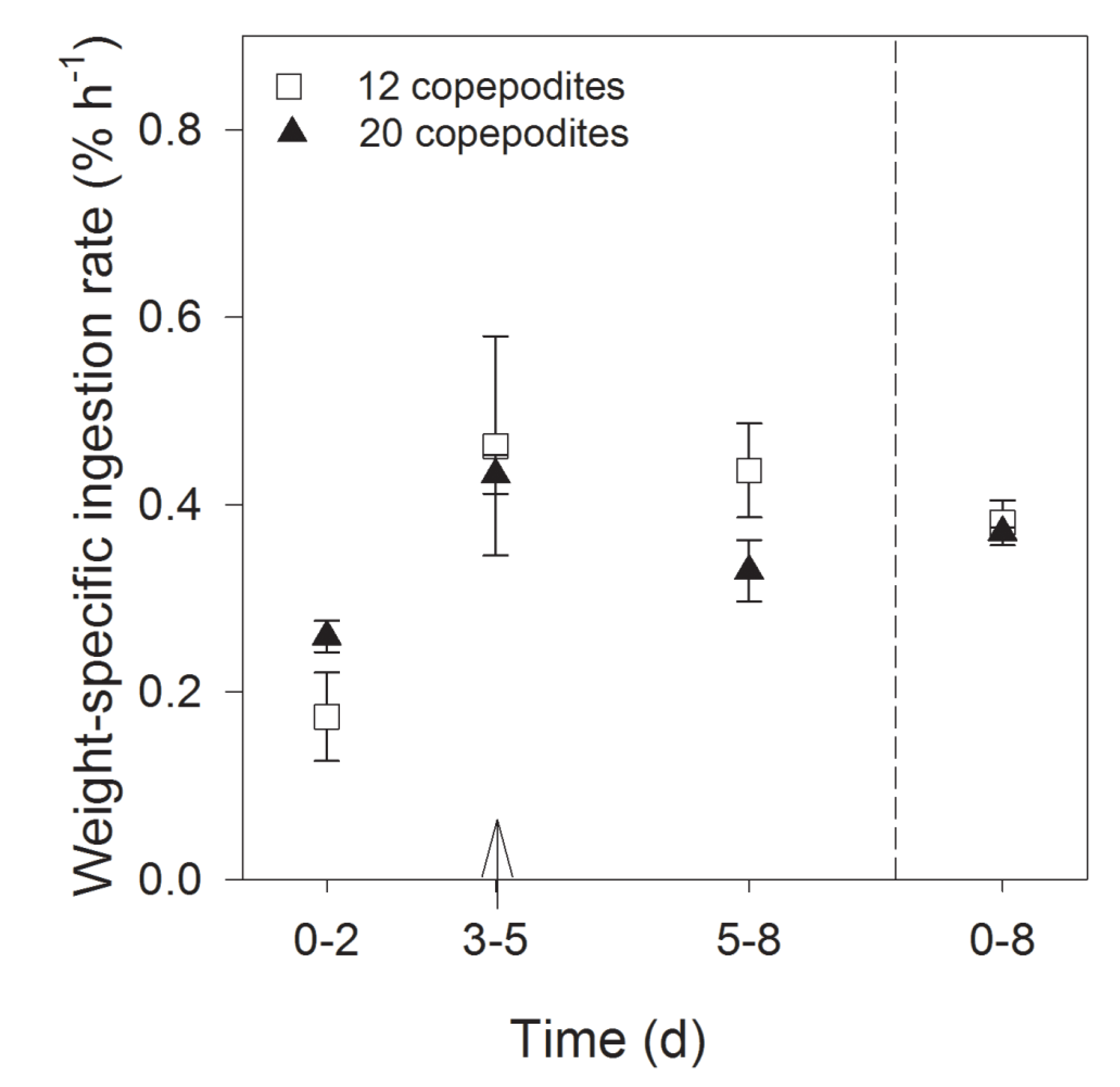
3. Discussion and Conclusions
3.1. Calanus Copepodites Grazing on Pseudo-nitzschia
3.1.1. Grazing
3.1.2. Copepodites as Vectors in the Arctic Food Web
3.2. Induction of Domoic Acid Production in Pseudo-nitzschia by Grazing Copepodites
3.3. Toxin Production in the Previously Non-Toxic P. obtusa
3.4. Overall Conclusion
4. Materials and Methods
4.1. Study Organisms
4.1.1. Zooplankton
| Pseudo-nitzschia | Cell Length (µm) | Cell Width (µm) | Cell Volume (µm3) | Carbon Content (pg C cell−1) |
| P. obtusa n = 23 | 33.5 ± 3.2 | 4.2 ± 0.5 | 489 ± 149 | 57.3 |
| P. seriata n = 23 | 49.0 ± 3.4 | 5.4 ± 0.8 | 1163 ± 383 | 137.1 |
| Calanus | Prosome length (mm) | Carbon content per individual (mg cop−1) | ||
| n = 21 | 2.5 ± 0.5 | 0.11 ± 0.1 |
4.1.2. Phytoplankton
4.2. Experiment Preparation, Nutrient and DA Analyses
4.3. Grazing Experiment
4.4. Induction Experiment
4.5. Induction Experiment in Filtrate Water
4.6. Microscopy
4.7. Statistical Analyses
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Quilliam, M.A.; Wright, J.L.C. The amnesic shellfish poisoning mystery. Anal. Chem. 1989, 61, 1053A–1060A. [Google Scholar] [CrossRef] [PubMed]
- Perl, T.M.; Bedard, L.; Kosatsky, T.; Hockin, J.C.; Todd, E.C.D.; Remis, R.S. An outbreak of toxic encephalopathy caused by eating mussels contaminated with domoic acid. N. Engl. J. Med. 1990, 322, 1775–1780. [Google Scholar] [CrossRef] [PubMed]
- Fritz, L.; Quilliam, M.A.; Wright, J.L.C.; Beale, A.M.; Work, T.M. An outbreak of domoic acid poisoning attributed to the pennate diatom Pseudo-nitzschia australis. J. Phycol. 1992, 28, 439–442. [Google Scholar] [CrossRef]
- Fire, S.E.; Wang, Z.H.; Berman, M.; Langlois, G.W.; Morton, S.L.; Sekula-Wood, E.; Benitez-Nelson, C.R. Trophic transfer of the harmful algal toxin domoic acid as a cause of death in a Minke whale (Balaenoptera acutorostrata) stranding in Southern California. Aquat. Mamm. 2010, 36, 342–350. [Google Scholar] [CrossRef]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef]
- Smida, D.B.; Lundholm, N.; Kooistra, W.; Sahraoui, I.; Ruggiero, M.V.; Kotaki, Y.; Ellegaard, M.; Lambert, C.; Mabrouk, H.H.; Hlaili, A.S. Morphology and molecular phylogeny of Nitzschia bizertensis sp. Nov. A new domoic acid producer. Harmful Algae 2014, 32, 49–63. [Google Scholar] [CrossRef]
- Costa, P.R.; Garrido, S. Domoic acid accumulation in the sardine Sardina pilchardus and its relationship to Pseudo-nitzschia diatom ingestion. Mar. Ecol. Prog. Ser. 2004, 284, 261–268. [Google Scholar] [CrossRef]
- Wohlgeschaffen, G.D.; Mann, K.H.; Rao, D.V.S.; Pocklington, R. Dynamics of the phycotoxin domoic acid accumulation and excretion in 2 commercially important bivalves. J. Appl. Phycol. 1992, 4, 297–310. [Google Scholar] [CrossRef]
- Maneiro, I.; Iglesias, P.; Guisande, C.; Riveiro, I.; Barreiro, A.; Zervoudaki, S.; Graneli, E. Fate of domoic acid ingested by the copepod Acartia clausi. Mar. Biol. 2005, 148, 123–130. [Google Scholar] [CrossRef]
- Turner, J.T. Planktonic marine copepods and harmful algae. Harmful Algae 2014, 32, 81–93. [Google Scholar] [CrossRef]
- Lelong, A.; Hegaret, H.; Soudant, P.; Bates, S.S. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: Revisiting previous paradigms. Phycologia 2012, 51, 168–216. [Google Scholar] [CrossRef]
- Hansen, L.R.; Soylu, S.Í.; Kotaki, Y.; Moestrup, Ø.; Lundholm, N. Toxin production and temperature-induced morphological variation of the diatom Pseudo-nitzschia seriata from the Arctic. Harmful Algae 2011, 10, 689–696. [Google Scholar] [CrossRef]
- Lundholm, N.; Skov, J.; Pocklington, R.; Moestrup, Ø. Domoic acid, the toxic amino-acid responsible for amnesic shellfish poisoning, now in Pseudonitzschia seriata (Bacillariophyceae) in Europe. Phycologia 1994, 33, 475–478. [Google Scholar] [CrossRef]
- Lincoln, J.A.; Turner, J.T.; Bates, S.S.; Leger, C.; Gauthier, D.A. Feeding, egg production, and egg hatching success of the copepods Acartia tonsa and Temora longicornis on diets of the toxic diatom Pseudo-nitzschia multiseries and the non-toxic diatom Pseudo-nitzschia pungens. Hydrobiologia 2001, 453, 107–120. [Google Scholar] [CrossRef]
- Leandro, L.F.; Teegarden, G.J.; Roth, P.B.; Wang, Z.; Doucette, G.J. The copepod Calanus finmarchicus: A potential vector for trophic transfer of the marine algal biotoxin, domoic acid. J. Exp. Mar. Biol. Ecol. 2010, 382, 88–95. [Google Scholar] [CrossRef]
- Tester, P.A.; Pan, Y.; Douchette, G.J. Accumulation of domoic acid activity in copepods. In Harmful and Toxic Algal Blooms 2000; Hallegraeff, G.M., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2001; pp. 401–404. [Google Scholar]
- Windust, A. The Responses of Bacteria, Microalgae, and Zooplankton to the Diatom Nitzschia pungens f. Multiseries and Its Toxic Metabolite Domoic Acid. M.S. Thesis, Dalhousie University, Halifax, Canada, 1992. [Google Scholar]
- Tammilehto, A.; Nielsen, T.G.; Krock, B.; Møller, E.F.; Lundholm, N. Calanus spp.-vectors for the biotoxin, domoic acid, in the Arctic marine ecosystem? Harmful Algae 2012, 20, 165–174. [Google Scholar] [CrossRef]
- Bargu, S.; Marinovic, B.; Mansergh, S.; Silver, M.W. Feeding responses of krill to the toxin-producing diatom Pseudo-nitzschia. J. Exp. Mar. Biol. Ecol. 2003, 284, 87–104. [Google Scholar] [CrossRef]
- Tammilehto, A.; Nielsen, T.G.; Krock, B.; Møller, E.F.; Lundholm, N. Induction of domoic acid production in the toxic diatom Pseudo-nitzschia seriata by calanoid copepods. Aquat. Toxicol. 2015, 159, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehling, J.; Davidson, K.; Bolch, C.J.; Bates, S.S. Growth and domoic acid production by Pseudo-nitzschia seriata (Bacillariophyceae) under phosphate and silicate limitation. J. Phycol. 2004, 40, 674–683. [Google Scholar] [CrossRef]
- Bates, S.S.; Worms, J.; Smith, J.C. Effects of ammonium and nitrate on growth and domoic acid production by Nitzschia pungens in batch culture. Can. J. Fish. Aquat. Sci. 1993, 50, 1248–1254. [Google Scholar] [CrossRef]
- Lundholm, N.; Hansen, P.J.; Kotaki, Y. Effect of pH on growth and domoic acid production by potentially toxic diatoms of the genera Pseudo-nitzschia and Nitzschia. Mar. Ecol. Prog. Ser. 2004, 273, 1–15. [Google Scholar] [CrossRef]
- Selander, E.; Thor, P.; Toth, G.; Pavia, H. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc. R. Soc. B 2006, 273, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Selander, E.; Cervin, G.; Pavia, H. Effects of nitrate and phosphate on grazer-induced toxin production in Alexandrium minutum. Limnol. Oceanogr. 2008, 53, 523–530. [Google Scholar] [CrossRef]
- Long, J.D.; Smalley, G.W.; Barsby, T.; Anderson, J.T.; Hay, M.E. Chemical cues induce consumer-specific defenses in a bloom-forming marine phytoplankton. Proc. Natl. Acad. Sci. USA 2007, 104, 10512–10517. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, V.; Granéli, E. Grazer-induced defence in Phaeocystis globosa (Prymnesiophyceae): Influence of different nutrient conditions. Limnol. Oceanogr. 2010, 55, 1965–1976. [Google Scholar] [CrossRef]
- Tande, K.S. Ecological investigations on the zooplankton community of Balsfjorden, northern Norway: Generation cycles, and variations in body-weight and body content of carbon and nitrogen related to overwintering and reproduction in the copepod Calanus finmarchicus (Gunnerus). J. Exp. Mar. Biol. Ecol. 1982, 62, 129–142. [Google Scholar]
- Hansen, B.W.; Nielsen, T.G.; Levinsen, H. Plankton community structure and carbon cycling on the western coast of Greenland during the stratified summer situation. III. Mesozooplankton. Aquat. Microb. Ecol. 1999, 16, 233–249. [Google Scholar] [CrossRef]
- Nielsen, T.G.; Hansen, B. Plankton community structure and carbon cycling on the western coast of Greenland during and after the sedimentation of a diatom bloom. Mar. Ecol. Prog. Ser. 1995, 125, 239–257. [Google Scholar] [CrossRef]
- Madsen, S.D.; Nielsen, T.G.; Hansen, B.W. Annual population development and production by Calanus finmarchicus, C. glacialis and C. hyperboreus in Disko bay, western Greenland. Mar. Biol. 2001, 139, 75–93. [Google Scholar]
- Wohlrab, S.; Iversen, M.H.; John, U. A molecular and co-evolutionary context for grazer induced toxin production in Alexandrium tamarense. PLoS ONE 2010, 5, e15039. [Google Scholar] [CrossRef] [PubMed]
- Whyte, J.N.C.; Townsend, L.D.; Ginther, N.G. Fecundity, toxin and trophic levels of the rotifer Brachionus plicatilis fed Pseudo-nitzschia pungens f. Multiseries. Available online: http://diatom.myspecies.info/node/331 (accessed on 23 May 2015).
- Meyer, B.; Irigoien, X.; Graeve, M.; Head, R.N.; Harris, R.P. Feeding rates and selectivity among nauplii, copepodites and adult females of Calanus finmarchicus and Calanus helgolandicus. Helgoland Mar. Res. 2002, 56, 169–176. [Google Scholar]
- Koski, M.; Wexels Riser, C. Post-bloom feeding of Calanus finmarchicus copepodites: Selection for autotrophic versus heterotrophic prey. Mar. Biol. Res. 2006, 2, 109–119. [Google Scholar] [CrossRef]
- Huntley, M.; Sykes, P.; Rohan, S.; Marin, V. Chemically-mediated rejection of dinoflagellate prey by the copepods Calanus pacificus and Paracalanus parvus–mechanism, occurrence and significance. Mar. Ecol. Prog. Ser. 1986, 28, 105–120. [Google Scholar] [CrossRef]
- Sykes, P.F.; Huntley, M.E. Acute physiological reactions of Calanus pacificus to selected dinoflagellates—direct observations. Mar. Biol. 1987, 94, 19–24. [Google Scholar] [CrossRef]
- Bergkvist, J.; Selander, E.; Pavia, H. Induction of toxin production in dinoflagellates: The grazer makes a difference. Oecologia 2008, 156, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Frost, B.W. Effects of size and concentration of food particles on feeding behavior of marine planktonic copepod Calanus pacificus. Limnol. Oceanogr. 1972, 17, 805–815. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology & Morphology of the Genera; Cambridge University Press: London, UK, 1990. [Google Scholar]
- Levinsen, H.; Turner, J.T.; Nielsen, T.G.; Hansen, B.W. On the trophic coupling between protists and copepods in arctic marine ecosystems. Mar. Ecol. Prog. Ser. 2000, 204, 65–77. [Google Scholar] [CrossRef]
- Shaw, B.A.; Andersen, R.J.; Harrison, P.J. Feeding deterrent and toxicity effects of apo-fucoxanthinoids and phycotoxins on a marine copepod (Tigriopus californicus). Mar. Biol. 1997, 128, 273–280. [Google Scholar] [CrossRef]
- Bargu, S.; Lefebvre, K.; Silver, M.W. Effect of dissolved domoic acid on the grazing rate of krill Euphausia pacifica. Mar. Ecol. Prog. Ser. 2006, 312, 169–175. [Google Scholar] [CrossRef]
- Colin, S.P.; Dam, H.G. Comparison of the functional and numerical responses of resistant versus non-resistant populations of the copepod Acartia hudsonica fed the toxic dinofiagellate Alexandrium tamarense. Harmful Algae 2007, 6, 875–882. [Google Scholar] [CrossRef]
- Colin, S.P.; Dam, H.G. Latitudinal differentiation in the effects of the toxic dinoflagellate Alexandrium spp. On the feeding and reproduction of populations of the copepod Acartia hudsonica. Harmful Algae 2002, 1, 113–125. [Google Scholar] [CrossRef]
- Colin, S.P.; Dam, H.G. Testing for resistance of pelagic marine copepods to a toxic dinoflagellate. Evol. Ecol. 2004, 18, 355–377. [Google Scholar] [CrossRef]
- Jiang, X.; Lonsdale, D.J.; Gobler, C.J. Rapid gain and loss of evolutionary resistance to the harmful dinoflagellate Cochlodinium polykrikoides in the copepod Acartia tonsa. Limnol. Oceanogr. 2011, 56, 947–954. [Google Scholar] [CrossRef]
- Zheng, Y.; Dam, H.G.; Avery, D.E. Differential responses of populations of the copepod Acartia hudsonica to toxic and nutritionally insufficient food algae. Harmful Algae 2011, 10, 723–731. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Connell, L.; Konoki, K.; MacQuarrie, S.P.; Scheuer, T.; Catterall, W.A.; Trainer, V.L. Sodium channel mutation leading to saxitoxin resistance in clams increases risk of PSP. Nature 2005, 434, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Hygum, B.H.; Rey, C.; Hansen, B.W.; Tande, K. Importance of food quantity to structural growth rate and neutral lipid reserves accumulated in Calanus finmarchicus. Mar. Biol. 2000, 136, 1057–1073. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H. Lipid content of Calanus finmarchicus during overwintering in the Faroe-Shetland Channel. Fish. Oceanogr. 1999, 8, 61–72. [Google Scholar] [CrossRef]
- Niehoff, B. The effect of food limitation on gonad development and egg production of the planktonic copepod Calanus finmarchicus. J. Exp. Mar. Biol. Ecol. 2004, 307, 237–259. [Google Scholar] [CrossRef]
- Cushing, D.H.; Humphrey, G.F.; Banse, K.; Laevastu, T. Report of the committee on terms and equivalents. Rapp. P.-V. Reun. Cons. Int. Explor. Mer. 1958, 144, 15–16. [Google Scholar]
- Swalethorp, R.; Kjellerup, S.; Dünweber, M.; Nielsen, T.G.; Møller, E.F.; Rysgaard, S.; Hansen, B.W. Grazing, egg production, and biochemical evidence of differences in the life strategies of Calanus finmarchicus, C. glacialis and C. hyperboreus in Disko bay, western Greenland. Mar. Ecol. Prog. Ser. 2011, 429, 125–144. [Google Scholar] [CrossRef]
- Harris, R.P.; Wiebe, P.H.; Lenz, J.; Skjoldal, H.R.; Huntley, M. Zooplankton Methodology Manual; Elsevier Academic Press: Beijing, China, 2005. [Google Scholar]
- Thor, P.; Wendt, I. Functional response of carbon absorption efficiency in the pelagic calanoid copepod Acartia tonsa Dana. Limnol. Oceanogr. 2010, 55, 1779–1789. [Google Scholar] [CrossRef]
- Selander, E.; Kubanek, J.; Hamberg, M.; Andersson, M.X.; Cervin, G.; Pavia, H. Predator lipids induce paralytic shellfish toxins in bloom-forming algae. Proc. Natl. Acad. Sci. USA 2015, 112, 6395–6400. [Google Scholar] [CrossRef] [PubMed]
- Trimborn, S.; Lundholm, N.; Thoms, S.; Richter, K.U.; Krock, B.; Hansen, P.J.; Rost, B. Inorganic carbon acquisition in potentially toxic and non-toxic diatoms: The effect of pH-induced changes in seawater carbonate chemistry. Physiol. Plant. 2008, 133, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, M.D.A.; Cochlan, W.P.; Ladizinsky, N.; Kudela, R.M. Nitrogenous preference of toxigenic Pseudo-nitzschia australis (Bacillariophyceae) from field and laboratory experiments. Harmful Algae 2007, 6, 206–217. [Google Scholar] [CrossRef]
- Heisterkamp, I.M.; Schramm, A.; de Beer, D.; Stief, P. Nitrous oxide production associated with coastal marine invertebrates. Mar. Ecol. Prog. Ser. 2010, 415, 1–9. [Google Scholar] [CrossRef]
- Koops, H.-P.; Purkhold, U.; Pommerening-Roeser, A.; Timmermann, G.; Wagner, M. The Lithoautotrophic Ammonia-Oxidizing Bacteria; Springer: New York, NY, USA, 2006; pp. 778–811. [Google Scholar]
- Stief, P.; University of Southern Denmark, Odense, Denmark. Personal communication, 2015.
- Anderson, C.R.; Brzezinski, M.A.; Washburn, L.; Kudela, R. Circulation and environmental conditions during a toxigenic Pseudo-nitzschia australis bloom in the Santa Barbara Channel, california. Mar. Ecol. Prog. Ser. 2006, 327, 119–133. [Google Scholar] [CrossRef]
- Schnetzer, A.; Miller, P.E.; Schaffner, R.A.; Stauffer, B.A.; Jones, B.H.; Weisberg, S.B.; DiGiacomo, P.M.; Berelson, W.M.; Caron, D.A. Blooms of Pseudo-nitzschia and domoic acid in the San Pedro Channel and Los Angeles harbor areas of the Southern California Bight, 2003–2004. Harmful Algae 2007, 6, 372–387. [Google Scholar] [CrossRef]
- Trainer, V.L.; Hickey, B.M.; Lessard, E.J.; Cochlan, W.P.; Trick, C.G.; Wells, M.L.; MacFadyen, A.; Moore, S.K. Variability of Pseudo-nitzschia and domoic acid in the Juan de Fuca eddy region and its adjacent shelves. Limnol. Oceanogr. 2009, 54, 289–308. [Google Scholar] [CrossRef]
- Trainer, V.L.; Wells, M.L.; Cochlan, W.P.; Trick, C.G.; Bill, B.D.; Baugh, K.A.; Beall, B.F.; Herndon, J.; Lundholm, N. An ecological study of a massive bloom of toxigenic Pseudo-nitzschia cuspidata off the Washington state coast. Limnol. Oceanogr. 2009, 54, 1461–1474. [Google Scholar] [CrossRef]
- Marchetti, A.; Trainer, V.L.; Harrison, P.J. Environmental conditions and phytoplankton dynamics associated with Pseudo-nitzschia abundance and domoic acid in the Juan de Fuca eddy. Mar. Ecol. Prog. Ser. 2004, 281, 1–12. [Google Scholar] [CrossRef]
- Lundholm, N.; Andersen, P.; Jorgensen, K.; Thorbjornsen, B.R.; Cembella, A.; Krock, B. Domoic acid in danish blue mussels due to a bloom of Pseudo-nitzschia seriata. Harmful Algae News 2005, 29, 8–10. [Google Scholar]
- Fehling, J.; Green, D.H.; Davidson, K.; Bolch, C.J.; Bates, S.S. Domoic acid production by Pseudo-nitzschia seriata (Bacillariophyceae) in Scottish waters. J. Phycol. 2004, 40, 622–630. [Google Scholar] [CrossRef]
- Bates, S.S.; Léger, C.; White, J.M.; MacNair, N.; Ehrman, J.M.; Levasseur, M.; Couture, J.Y.; Gagnon, R.; Bonneau, F.; Michaud, S.; et al. Pennate diatom Pseudo-nitzschia seriata; domoic acid production causes spring closures of shellfish harvesting for the first time in the southern gulf of St. Lawrence, eastern Canada. In Proceedings of the 17th International Diatom Symposium, Ottawa, Canada, 25–31 August 2002.
- Trainer, V.L.; Adams, N.G.; Bill, B.D.; Stehr, C.M.; Wekell, J.C.; Moeller, P.; Busman, M.; Woodruff, D. Domoic acid production near California coastal upwelling zones, June 1998. Limnol. Oceanogr. 2000, 45, 1818–1833. [Google Scholar] [CrossRef]
- Scholin, C.A.; Gulland, F.; Doucette, G.J.; Benson, S.; Busman, M.; Chavez, F.P.; Cordaro, J.; DeLong, R.; De Vogelaere, A.; Harvey, J.; et al. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 403, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Hasle, G.R.; Lundholm, N. Pseudo-nitzschia seriata f. obtusa (Bacillariophyceae) raised in rank based on morphological, phylogenetic and distributional data. Phycologia 2005, 44, 608–619. [Google Scholar]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a diatom, not a chyrsophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Hansen, P.J.; Bjornsen, P.K.; Hansen, B.W.; Bjørnsen, P.K. Zooplankton grazing and growth: Scaling within the 2–2000 µm body size range. Limnol. Oceanogr. 1997, 42, 687–704. [Google Scholar] [CrossRef]
- Corner, E.D.S.; Newell, B.S. On nutrition and metabolism of zooplankton 4 forms of nitrogen excreted by Calanus. J. Mar. Biol. Assoc. UK 1967, 47, 113–120. [Google Scholar] [CrossRef]
- Hansen, H.P.; Koroleff, F. Determination of nutrients. In Methods of Seawater Analysis; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2007; pp. 159–228. [Google Scholar]
- Tang, K.W. Grazing and colony size development in Phaeocystis globosa (Prymnesiophyceae): The role of a chemical signal. J. Plankton Res. 2003, 25, 831–842. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harðardóttir, S.; Pančić, M.; Tammilehto, A.; Krock, B.; Møller, E.F.; Nielsen, T.G.; Lundholm, N. Dangerous Relations in the Arctic Marine Food Web: Interactions between Toxin Producing Pseudo-nitzschia Diatoms and Calanus Copepodites. Mar. Drugs 2015, 13, 3809-3835. https://doi.org/10.3390/md13063809
Harðardóttir S, Pančić M, Tammilehto A, Krock B, Møller EF, Nielsen TG, Lundholm N. Dangerous Relations in the Arctic Marine Food Web: Interactions between Toxin Producing Pseudo-nitzschia Diatoms and Calanus Copepodites. Marine Drugs. 2015; 13(6):3809-3835. https://doi.org/10.3390/md13063809
Chicago/Turabian StyleHarðardóttir, Sara, Marina Pančić, Anna Tammilehto, Bernd Krock, Eva Friis Møller, Torkel Gissel Nielsen, and Nina Lundholm. 2015. "Dangerous Relations in the Arctic Marine Food Web: Interactions between Toxin Producing Pseudo-nitzschia Diatoms and Calanus Copepodites" Marine Drugs 13, no. 6: 3809-3835. https://doi.org/10.3390/md13063809