
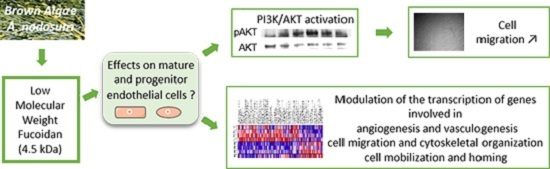
Low-Molecular-Weight Fucoidan Induces Endothelial Cell Migration via the PI3K/AKT Pathway and Modulates the Transcription of Genes Involved in Angiogenesis

 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
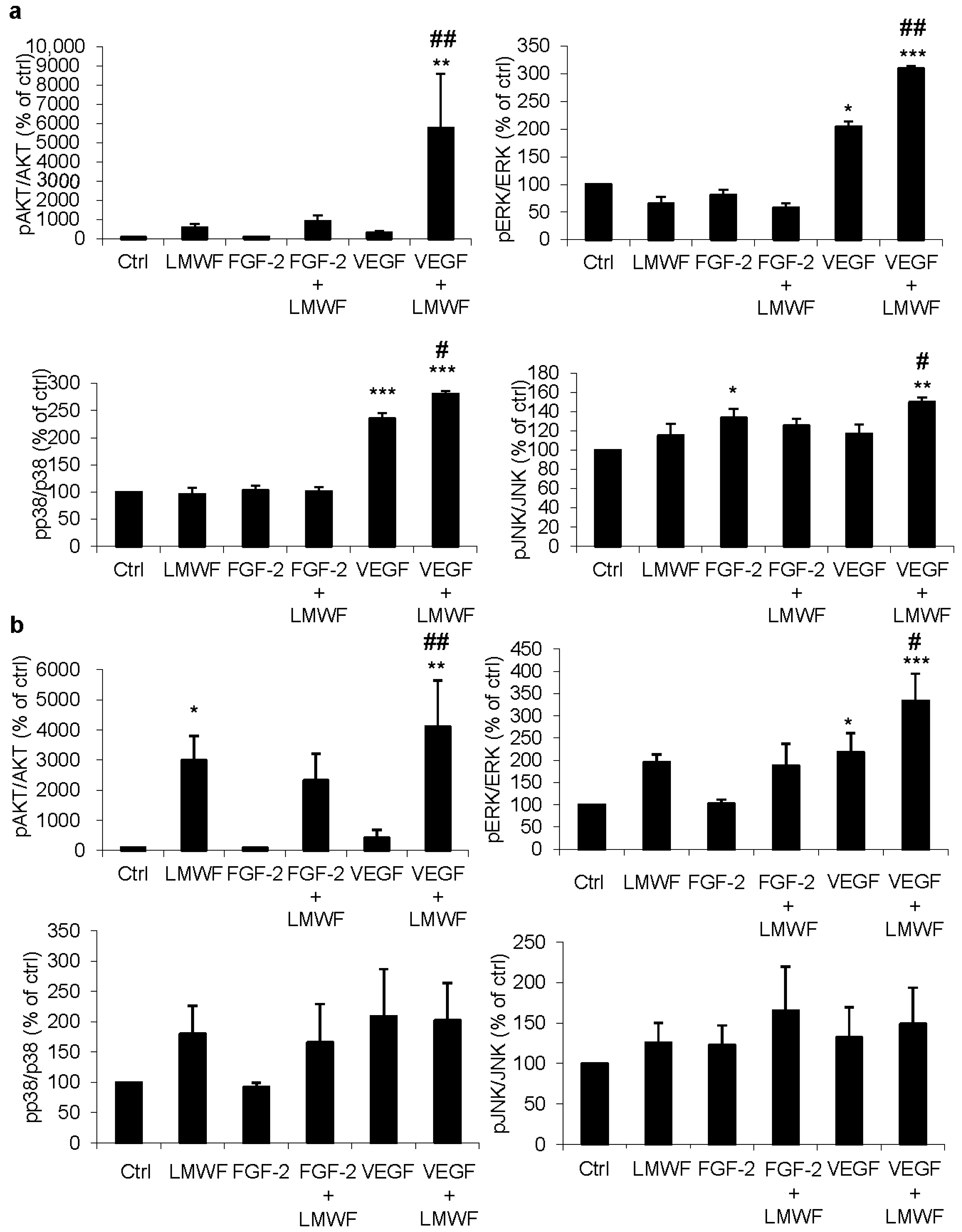
2.1. LMWF Enhances VEGF-Induced Activation of Signaling Pathways
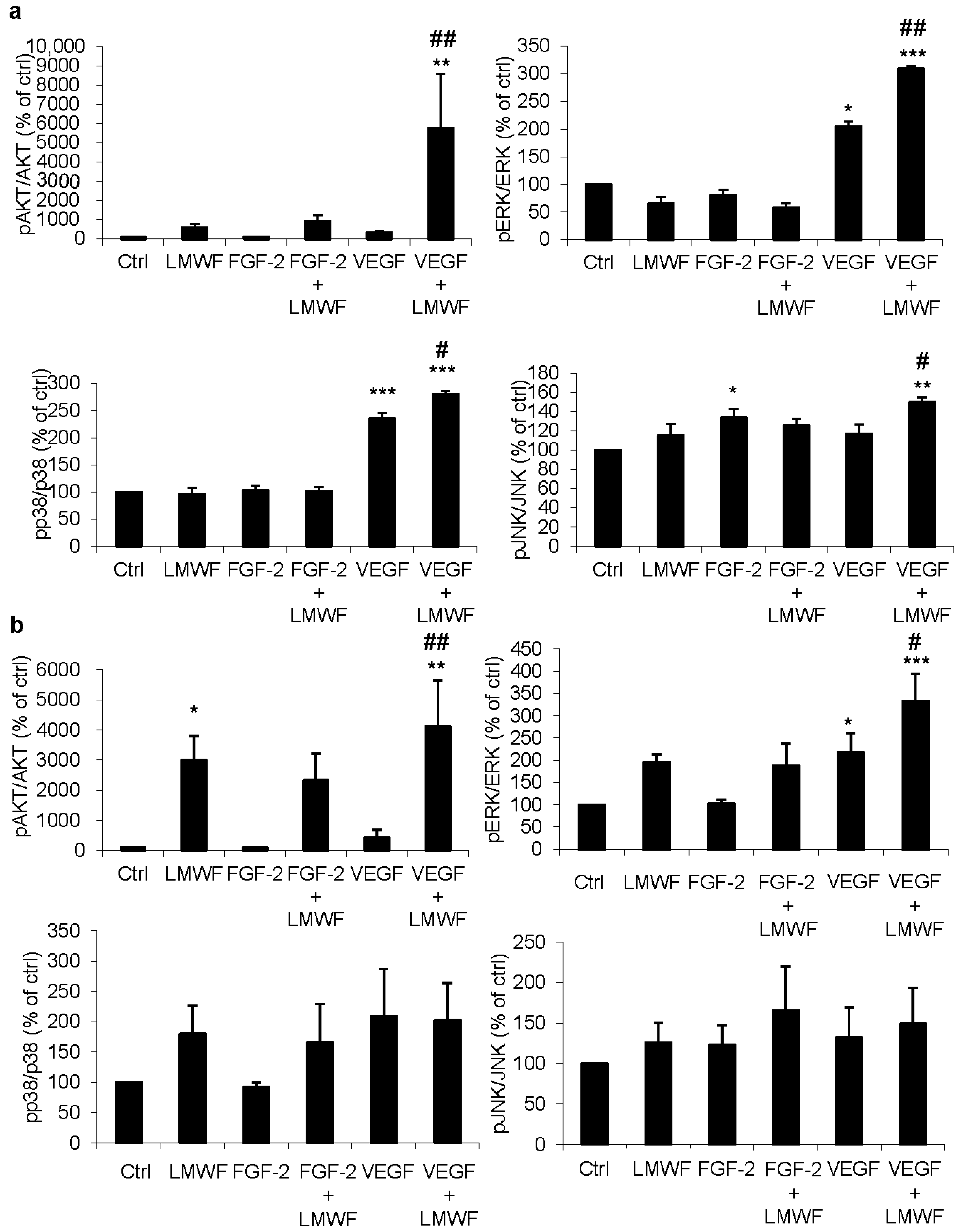
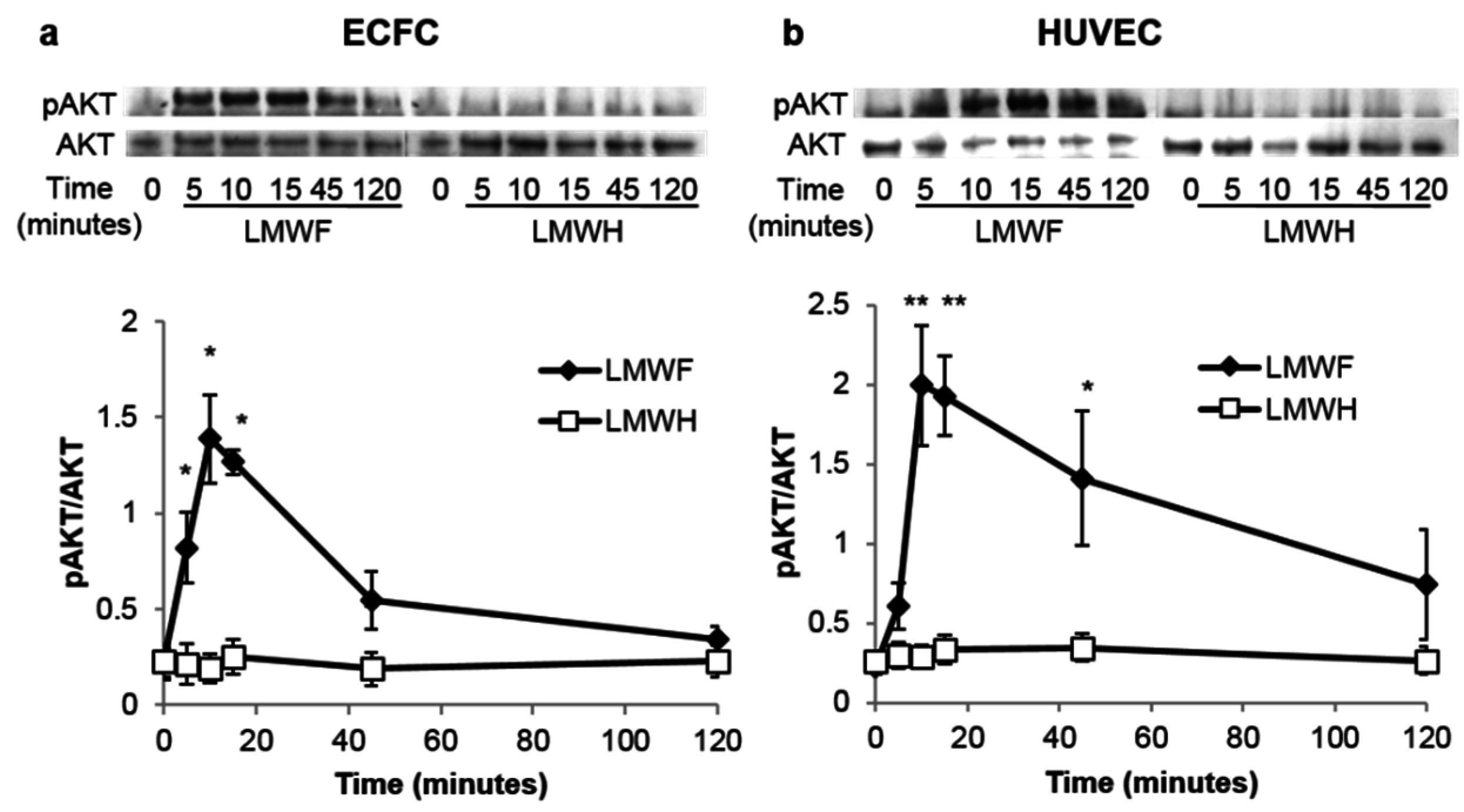
2.2. LMWF but Not LMWH Induces AKT Phosphorylation in ECFCs and HUVECs
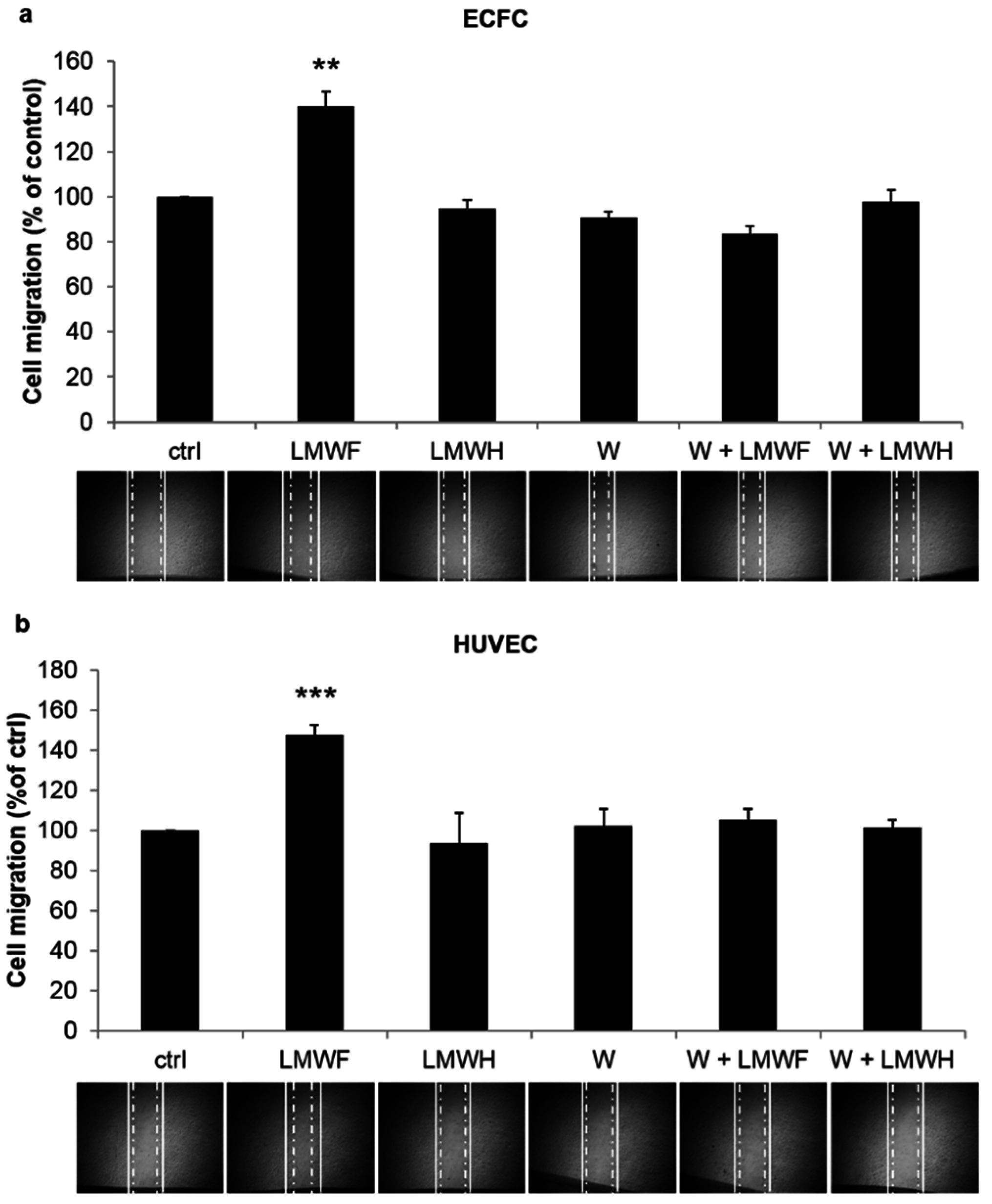
2.3. LMWF Enhances ECFC and HUVEC Cell Migration in a PI3K-Dependent Manner

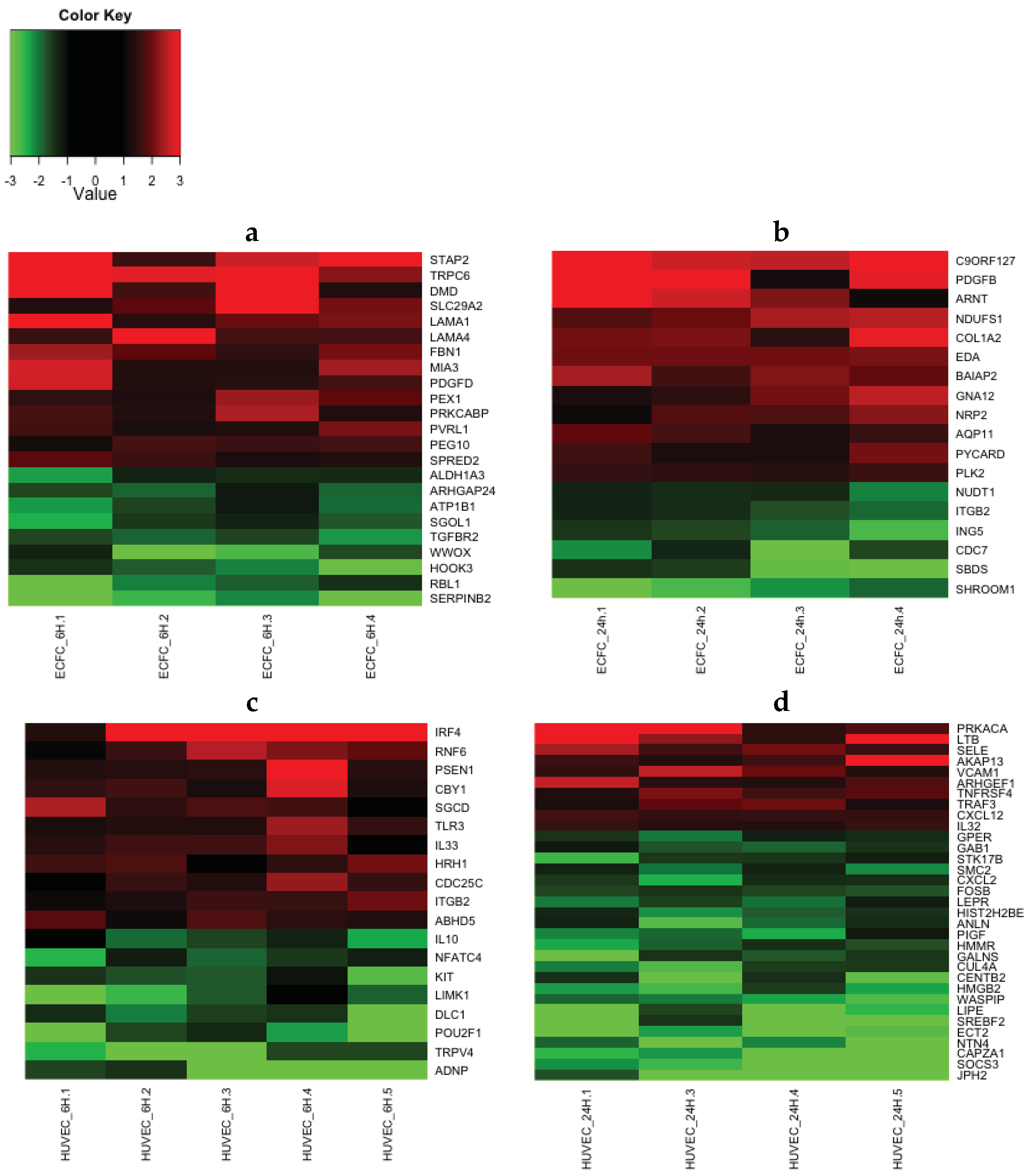
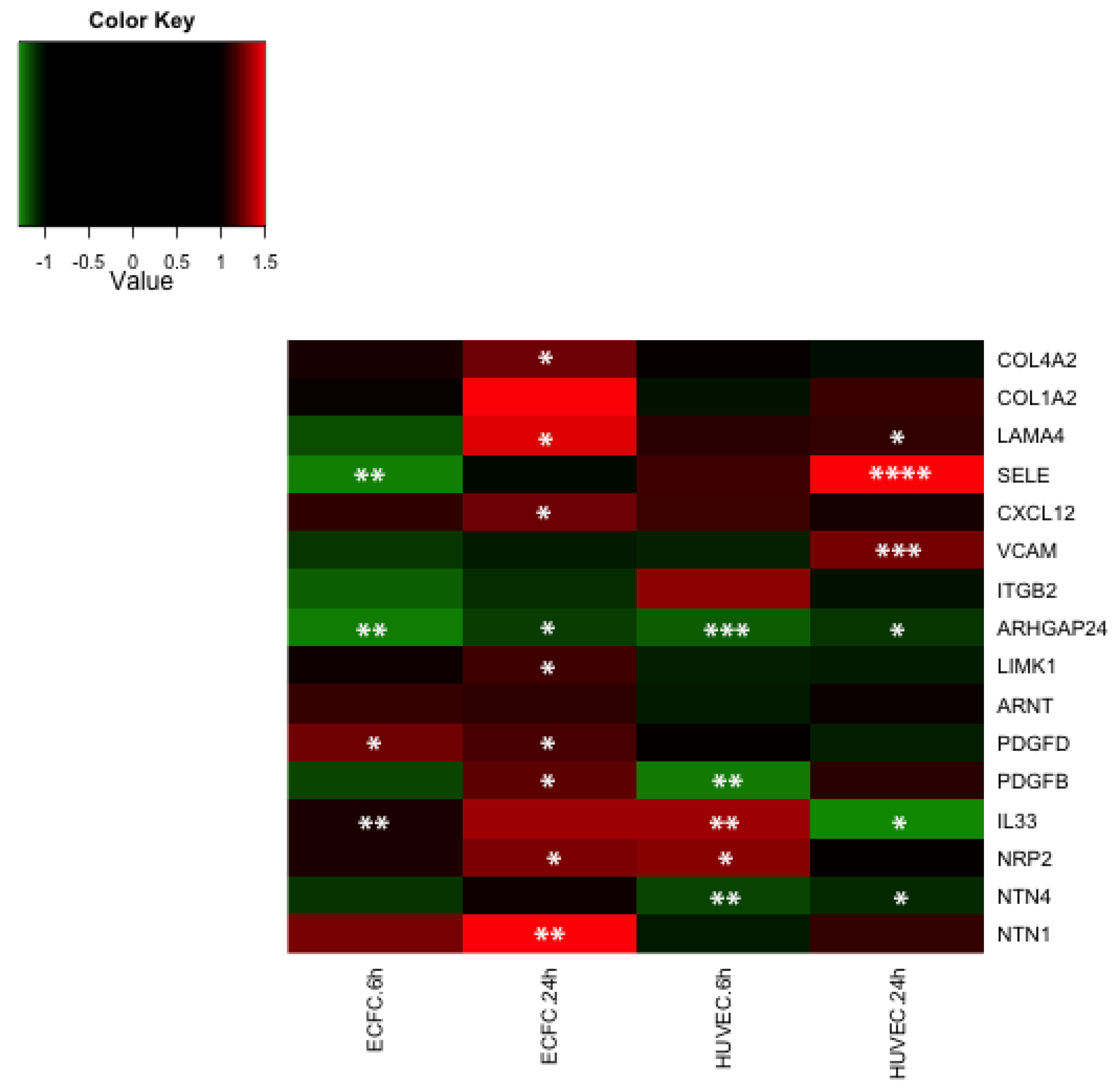
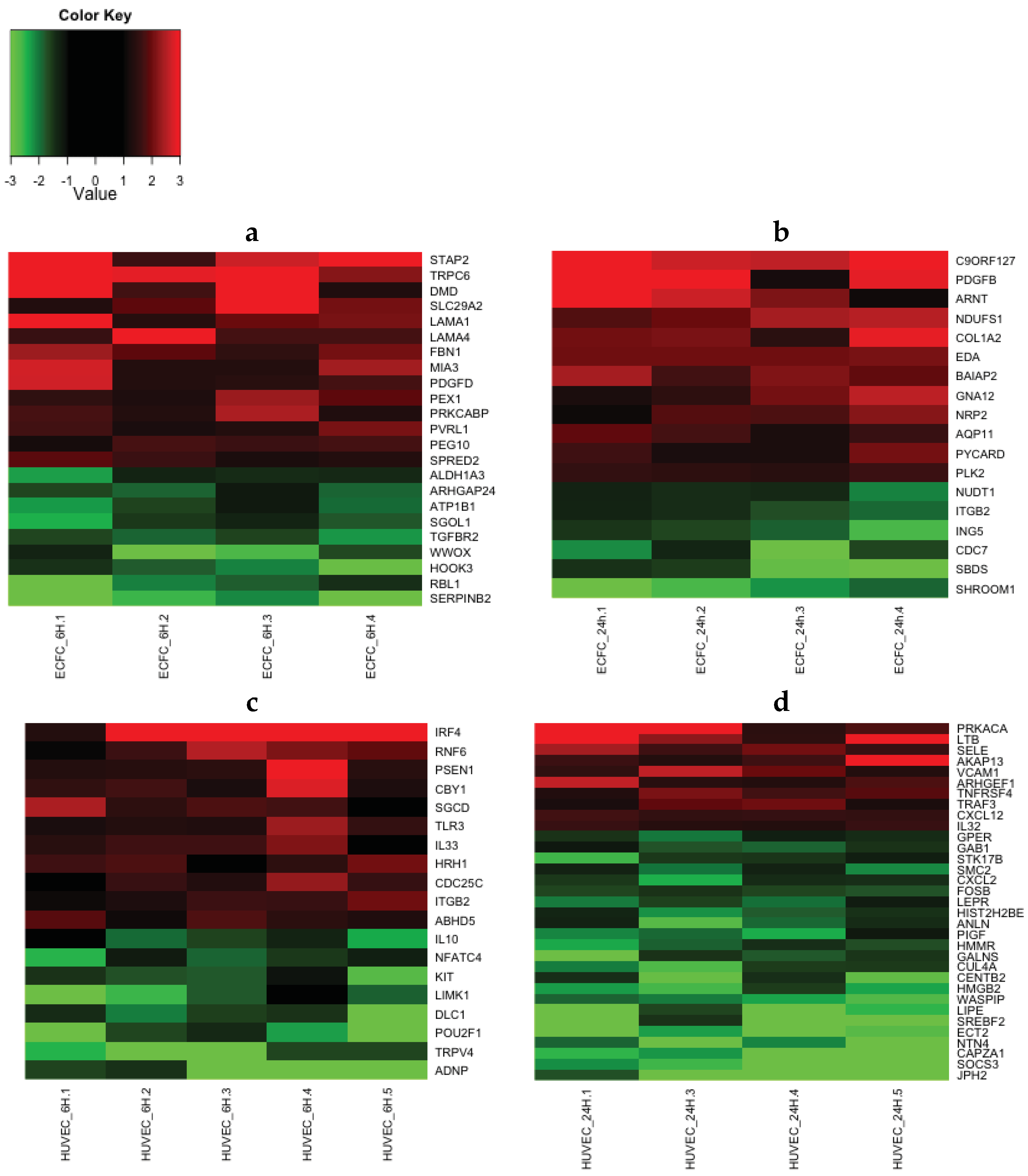
2.4. Transcriptomic Analysis Reveals that LMWF Modulates the Expression of Genes Involved in Angiogenesis, Vasculogenesis and Cell Migration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Genes | ECFC 6 h | ECFC 24 h | HUVEC 6 h | HUVEC 24 h |
|---|---|---|---|---|
| upregulated | 110 | 66 | 74 | 65 |
| downregulated | 41 | 50 | 91 | 128 |
| Total | 151 | 116 | 165 | 193 |
| Number of Genes (n) by Function | ECFC 6 h | ECFC 24 h | HUVEC 6 h | HUVEC 24 h | ||||
|---|---|---|---|---|---|---|---|---|
| n | p-value | n | p-value | n | p-value | n | p-value | |
| Cardiovascular System Development and Function | 5 | 4.64 × 10−3–1.38 × 10−2 | 5 | 5.73 × 10−4–3.24 × 10−2 | 4 | 5.78 × 10−3–3.98 × 10−2 | 11 | 2.93 × 10−4–4.84 × 10−2 |
| Cell Morphology | 4 | 4.64 × 10−3–2.3 × 10−2 | 6 | 3.29 × 10−3–4.51 × 10−2 | 11 | 1.58 × 10−3–3.98 × 10−2 | 14 | 1.33 × 10−3–4.84 × 10−2 |
| Cell-To-Cell Signaling and Interaction | 7 | 4.64 × 10−3–3.65 × 10−2 | 8 | 3.03 × 10−3–4.83 × 10−2 | 10 | 9.03 × 10−4–4.99 × 10−2 | 19 | 1.47 × 10−4–4.84 × 10−2 |
| Cellular Assembly and Organization | 8 | 4.64 × 10−3–4.98 × 10−2 | 10 | 1.4 × 10−3–4.76 × 10−2 | 7 | 1.58 × 10−3–3.98 × 10−2 | 15 | 4.93 × 10−5–4.84 × 10−2 |
| Cellular Growth and Proliferation | 4 | 4.64 × 10−3–4.1 × 10−2 | 4 | 3.29 × 10−3–4.51 × 10−2 | 8 | 3.3 × 10−3–4.53 × 10−2 | 30 | 1.82 × 10−3–4.51 × 10−2 |
| Cellular Movement | 2 | 1.84 × 10−2–2.75 × 10−2 | 6 | 4.7 × 10−4–4.83 × 10−2 | 7 | 4.88 × 10−4–4.53 × 10−2 | 15 | 1.38 × 10−5–4.84 × 10−2 |
| Hematological System Development and Function | 4 | 4.64 × 10−3–4.1 × 10−2 | 4 | 3.29 × 10−3–4.83 × 10−2 | 10 | 4.88 × 10−4–4.99 × 10−2 | 15 | 4.93 × 10−5–4.84 × 10−2 |
| Hematopoiesis | 2 | 4.64 × 10−3–4.1 × 10−2 | 2 | 9.84 × 10−3–3.88 × 10−2 | 7 | 4.88 × 10−4–4.57 × 10−2 | 7 | 1.46 × 10−4–4.84 × 10−2 |
| Immune Cell Trafficking | 0 | n/a | 3 | 3.29 × 10−3–4.83 × 10−2 | 7 | 4.88 × 10−4–4.99 × 10−2 | 7 | 4.93 × 10−5–4.84 × 10−2 |
| Inflammatory Response | 2 | 1.84 × 10−2–4.1 × 10−2 | 4 | 3.29 × 10−3–4.83 × 10−2 | 8 | 4.88 × 10−4–4.57 × 10−2 | 8 | 4.93 × 10−5–4.55 × 10−2 |
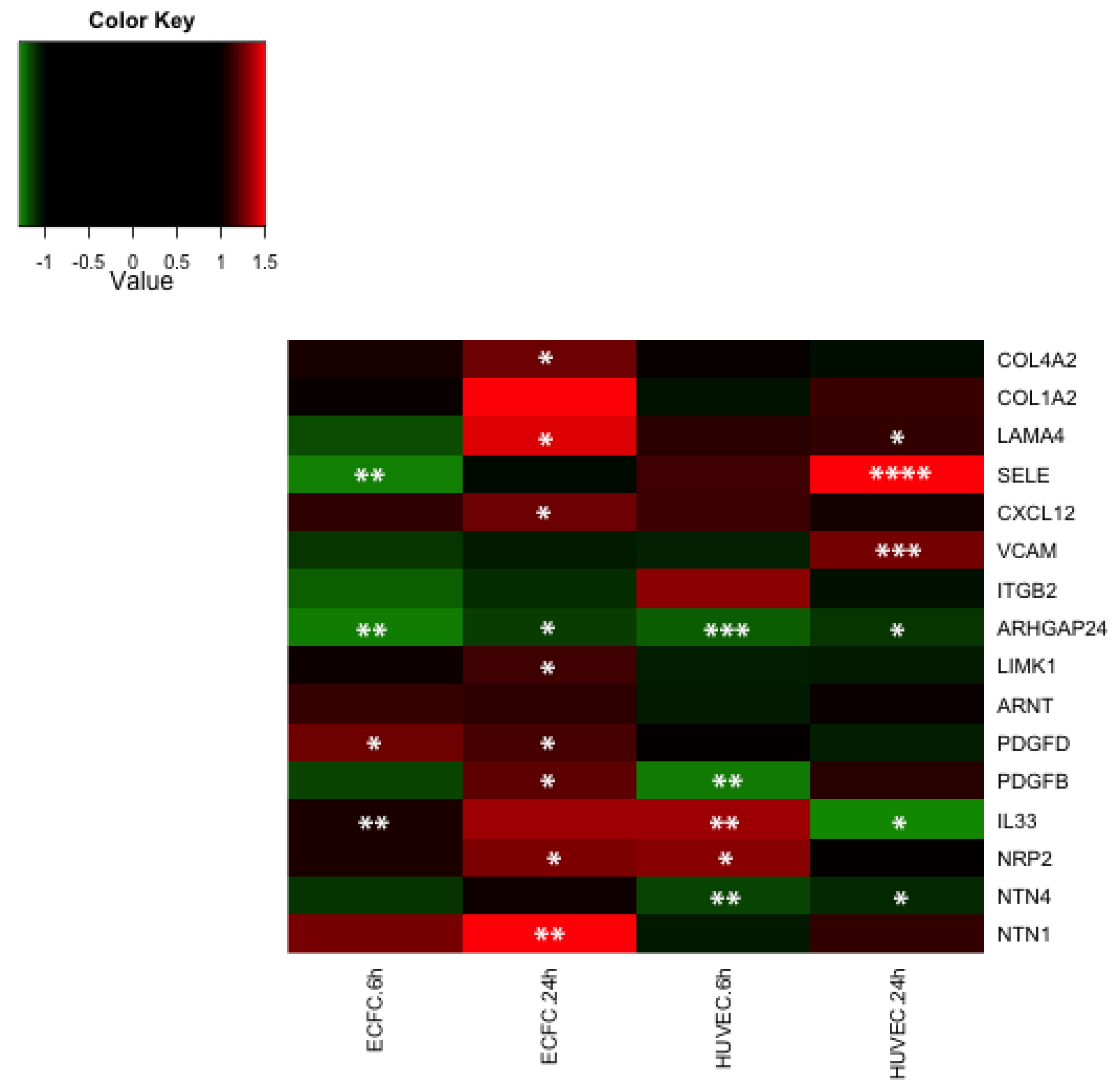
2.5. Discussion
3. Experimental Section
3.1. Reagents
3.2. ECFC Isolation and Culture
3.3. HUVEC Isolation and Culture
3.4. Bio-Plex® Analysis
3.5. Western Blot Analysis
3.6. In Vitro Wound Healing Assay
3.7. Transcriptome Analysis by Microarray
3.8. Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Colliec-Jouault, S.; Millet, J.; Helley, D.; Sinquin, C.; Fischer, A.M. Effect of low-molecular-weight fucoidan on experimental arterial thrombosis in the rabbit and rat. J. Thromb. Haemost. 2003, 1, 1114–1115. [Google Scholar] [CrossRef] [PubMed]
- Durand, E.; Helley, D.; Al Haj Zen, A.; Dujols, C.; Bruneval, P.; Colliec-Jouault, S.; Fischer, A.M.; Lafont, A. Effect of low molecular weight fucoidan and low molecular weight heparin in a rabbit model of arterial thrombosis. J. Vasc. Res. 2008, 45, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Deux, J.F.; Meddahi-Pelle, A.; Bree, F.; Bataille, I.; Michel, J.B.; Letourneur, D. Comparative studies on the mechanisms of action of four polysaccharides on arterial restenosis. J. Biomater. Sci. Polym. Ed. 2009, 20, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Deux, J.F.; Meddahi-Pelle, A.; le Blanche, A.F.; Feldman, L.J.; Colliec-Jouault, S.; Bree, F.; Boudghene, F.; Michel, J.B.; Letourneur, D. Low molecular weight fucoidan prevents neointimal hyperplasia in rabbit iliac artery in-stent restenosis model. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Luyt, C.E.; Meddahi-Pelle, A.; Ho-Tin-Noe, B.; Colliec-Jouault, S.; Guezennec, J.; Louedec, L.; Prats, H.; Jacob, M.P.; Osborne-Pellegrin, M.; Letourneur, D.; et al. Low-molecular-weight fucoidan promotes therapeutic revascularization in a rat model of critical hindlimb ischemia. J. Pharmacol. Exp. Ther. 2003, 305, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Chabut, D.; Fischer, A.M.; Colliec-Jouault, S.; Laurendeau, I.; Matou, S.; le Bonniec, B.; Helley, D. Low molecular weight fucoidan and heparin enhance the basic fibroblast growth factor-induced tube formation of endothelial cells through heparan sulfate-dependent alpha6 overexpression. Mol. Pharmacol. 2003, 64, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Zemani, F.; Benisvy, D.; Galy-Fauroux, I.; Lokajczyk, A.; Colliec-Jouault, S.; Uzan, G.; Fischer, A.M.; Boisson-Vidal, C. Low-molecular-weight fucoidan enhances the proangiogenic phenotype of endothelial progenitor cells. Biochem. Pharmacol. 2005, 70, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Lake, A.C.; Vassy, R.; di Benedetto, M.; Lavigne, D.; le Visage, C.; Perret, G.Y.; Letourneur, D. Low molecular weight fucoidan increases VEGF165-induced endothelial cell migration by enhancing VEGF165 binding to VEGFR-2 and NRP1. J. Biol. Chem. 2006, 281, 37844–37852. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, A.; Asahara, T. Role of progenitor endothelial cells in cardiovascular disease and upcoming therapies. Catheter. Cardiovasc. Interv. 2007, 70, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Bouvard, C.; Gafsou, B.; Dizier, B.; Galy-Fauroux, I.; Lokajczyk, A.; Boisson-Vidal, C.; Fischer, A.M.; Helley, D. Alpha6-integrin subunit plays a major role in the proangiogenic properties of endothelial progenitor cells. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1569–1575. [Google Scholar] [CrossRef] [PubMed]
- Sarlon, G.; Zemani, F.; David, L.; Duong van Huyen, J.P.; Dizier, B.; Grelac, F.; Colliec-Jouault, S.; Galy-Fauroux, I.; Bruneval, P.; Fischer, A.M.; et al. Therapeutic effect of fucoidan-stimulated endothelial colony-forming cells in peripheral ischemia. J. Thromb. Haemost. 2012, 10, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Park, J.Y.; Kang, H.J.; Kim, H.J.; Lee, J. Fucoidan/FGF-2 induces angiogenesis through JNK- and p38-mediated activation of AKT/MMP-2 signalling. Biochem. Biophys. Res. Commun. 2014, 450, 1333–1338. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ruiz, M.; Fulton, D.; Sowa, G.; Languino, L.R.; Fujio, Y.; Walsh, K.; Sessa, W.C. Vascular endothelial growth factor-stimulated actin reorganization and migration of endothelial cells is regulated via the serine/threonine kinase Akt. Circ. Res. 2000, 86, 892–896. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Luciano, A.K.; Ackah, E.; Rodriguez-Vita, J.; Bancroft, T.A.; Eichmann, A.; Simons, M.; Kyriakides, T.R.; Morales-Ruiz, M.; Sessa, W.C. Endothelial Akt1 mediates angiogenesis by phosphorylating multiple angiogenic substrates. Proc. Natl. Acad. Sci. USA 2014, 111, 12865–12870. [Google Scholar] [CrossRef] [PubMed]
- Shiojima, I. Role of Akt signaling in vascular homeostasis and angiogenesis. Circu. Res. 2002, 90, 1243–1250. [Google Scholar] [CrossRef]
- Mena, H.A.; Lokajczyk, A.; Dizier, B.; Strier, S.E.; Voto, L.S.; Boisson-Vidal, C.; Schattner, M.; Negrotto, S. Acidic preconditioning improves the proangiogenic responses of endothelial colony forming cells. Angiogenesis 2014, 17, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Dimmeler, S.; Aicher, A.; Vasa, M.; Mildner-Rihm, C.; Adler, K.; Tiemann, M.; Rutten, H.; Fichtlscherer, S.; Martin, H.; Zeiher, A.M. HMG-CoA reductase inhibitors (statins) increase endothelial progenitor cells via the PI 3-kinase/Akt pathway. J. Clin. Investig. 2001, 108, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.; Yoon, C.H.; Lee, C.S.; Kim, T.Y.; Oh, I.Y.; Park, K.W.; Kim, J.H.; Lee, H.S.; Kang, H.J.; Chae, I.H.; et al. Akt is a key modulator of endothelial progenitor cell trafficking in ischemic muscle. Stem Cells 2007, 25, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Madeddu, P.; Kraenkel, N.; Barcelos, L.S.; Siragusa, M.; Campagnolo, P.; Oikawa, A.; Caporali, A.; Herman, A.; Azzolino, O.; Barberis, L.; et al. Phosphoinositide 3-kinase gamma gene knockout impairs postischemic neovascularization and endothelial progenitor cell functions. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Lavelin, I.; Geiger, B. Characterization of a novel GTPase-activating protein associated with focal adhesions and the actin cytoskeleton. J. Biol. Chem. 2005, 280, 7178–7185. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Hartwig, J.H.; Stossel, T.P. FilGAP, a Rho- and ROCK-regulated GAP for Rac binds filamin A to control actin remodelling. Nat. Cell Biol. 2006, 8, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.J.; Hahn, C.N.; Goodall, G.J.; Reck, N.M.; Leske, A.F.; Davy, A.; Kremmidiotis, G.; Vadas, M.A.; Gamble, J.R. A vascular cell-restricted RhoGAP, p73RhoGAP, is a key regulator of angiogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 12212–12217. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Y.; Xing, F.; Wang, J.; Wang, Y.; Wang, H.; Yang, Y.; Gao, Z. Overexpression of LIMK1 promotes migration ability of multidrug-resistant osteosarcoma cells. Oncol. Res. 2011, 19, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Masuyama, N.; Fukui, Y.; Suzuki, A.; Gotoh, Y. Akt mediates Rac/Cdc42-regulated cell motility in growth factor-stimulated cells and in invasive PTEN knockout cells. Curr. Biol. 2001, 11, 1958–1962. [Google Scholar] [CrossRef]
- Yang, H.W.; Shin, M.G.; Lee, S.; Kim, J.R.; Park, W.S.; Cho, K.H.; Meyer, T.; Heo, W.D. Cooperative activation of PI3K by Ras and Rho family small GTPases. Mol. Cell 2012, 47, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Newey, S.E.; Tsaknakis, G.; Khoo, C.P.; Athanassopoulos, T.; Camicia, R.; Zhang, Y.; Grabowska, R.; Harris, A.L.; Roubelakis, M.G.; Watt, S.M. The hematopoietic chemokine CXCL12 promotes integration of human endothelial colony forming cell-derived cells into immature vessel networks. Stem Cells Dev. 2014, 23, 2730–2743. [Google Scholar] [CrossRef] [PubMed]
- Zemani, F.; Silvestre, J.S.; Fauvel-Lafeve, F.; Bruel, A.; Vilar, J.; Bieche, I.; Laurendeau, I.; Galy-Fauroux, I.; Fischer, A.M.; Boisson-Vidal, C. Ex vivo priming of endothelial progenitor cells with SDF-1 before transplantation could increase their proangiogenic potential. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, J.; Kusano, K.F.; Masuo, O.; Kawamoto, A.; Silver, M.; Murasawa, S.; Bosch-Marce, M.; Masuda, H.; Losordo, D.W.; Isner, J.M.; et al. Stromal cell-derived factor-1 effects on ex vivo expanded endothelial progenitor cell recruitment for ischemic neovascularization. Circulation 2003, 107, 1322–1328. [Google Scholar] [CrossRef] [PubMed]
- Pello, O.M.; Moreno-Ortiz Mdel, C.; Rodriguez-Frade, J.M.; Martinez-Munoz, L.; Lucas, D.; Gomez, L.; Lucas, P.; Samper, E.; Aracil, M.; Martinez, C.; et al. SOCS up-regulation mobilizes autologous stem cells through CXCR4 blockade. Blood 2006, 108, 3928–3937. [Google Scholar] [CrossRef] [PubMed]
- Oh, I.Y.; Yoon, C.H.; Hur, J.; Kim, J.H.; Kim, T.Y.; Lee, C.S.; Park, K.W.; Chae, I.H.; Oh, B.H.; Park, Y.B.; et al. Involvement of E-selectin in recruitment of endothelial progenitor cells and angiogenesis in ischemic muscle. Blood 2007, 110, 3891–3899. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, Y.; Graziani, G.M.; Filion, L.; Allan, D.S. E-selectin mediated adhesion and migration of endothelial colony forming cells is enhanced by SDF-1α/CXCR4. PLoS ONE 2013, 8, e60890. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.E.; Halloran, M.M.; Haskell, C.J.; Shah, M.R.; Polverini, P.J. Angiogenesis mediated by soluble forms of E-selectin and vascular cell adhesion molecule-1. Nature 1995, 376, 517–519. [Google Scholar] [CrossRef] [PubMed]
- Hellstrom, M.; Kalen, M.; Lindahl, P.; Abramsson, A.; Betsholtz, C. Role of PDGF-B and PDGFR-beta in recruitment of vascular smooth muscle cells and pericytes during embryonic blood vessel formation in the mouse. Development 1999, 126, 3047–3055. [Google Scholar] [PubMed]
- Lin, R.Z.; Moreno-Luna, R.; Li, D.; Jaminet, S.C.; Greene, A.K.; Melero-Martin, J.M. Human endothelial colony-forming cells serve as trophic mediators for mesenchymal stem cell engraftment via paracrine signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 10137–10142. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.; Portron, S.; Dizier, B.; Vinatier, C.; Masson, M.; Sourice, S.; Galy-Fauroux, I.; Corre, P.; Weiss, P.; Fischer, A.M.; et al. The in vitro and in vivo effects of a low-molecular-weight fucoidan on the osteogenic capacity of human adipose-derived stromal cells. Tissue Eng. A 2014, 20, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Uutela, M.; Wirzenius, M.; Paavonen, K.; Rajantie, I.; He, Y.; Karpanen, T.; Lohela, M.; Wiig, H.; Salven, P.; Pajusola, K.; et al. PDGF-D induces macrophage recruitment, increased interstitial pressure, and blood vessel maturation during angiogenesis. Blood 2004, 104, 3198–3204. [Google Scholar] [CrossRef] [PubMed]
- Thyboll, J.; Kortesmaa, J.; Cao, R.; Soininen, R.; Wang, L.; Iivanainen, A.; Sorokin, L.; Risling, M.; Cao, Y.; Tryggvason, K. Deletion of the laminin alpha4 chain leads to impaired microvessel maturation. Mol. Cell. Biol. 2002, 22, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Seano, G.; Chiaverina, G.; Gagliardi, P.A.; di Blasio, L.; Puliafito, A.; Bouvard, C.; Sessa, R.; Tarone, G.; Sorokin, L.; Helley, D.; et al. Endothelial podosome rosettes regulate vascular branching in tumour angiogenesis. Nat. Cell Biol. 2014, 16, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.H.; Eichmann, A. Axon guidance molecules in vascular patterning. Cold Spring Harb. Perspect. Biol. 2010, 2, a001875. [Google Scholar] [CrossRef] [PubMed]
- Favier, B.; Alam, A.; Barron, P.; Bonnin, J.; Laboudie, P.; Fons, P.; Mandron, M.; Herault, J.P.; Neufeld, G.; Savi, P.; et al. Neuropilin-2 interacts with VEGFR-2 and VEGFR-3 and promotes human endothelial cell survival and migration. Blood 2006, 108, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.D.; Ii, M.; Park, K.W.; Suli, A.; Sorensen, L.K.; Larrieu-Lahargue, F.; Urness, L.D.; Suh, W.; Asai, J.; Kock, G.A.; et al. Netrins promote developmental and therapeutic angiogenesis. Science 2006, 313, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Larrivee, B.; Freitas, C.; Trombe, M.; Lv, X.; Delafarge, B.; Yuan, L.; Bouvree, K.; Breant, C.; Del Toro, R.; Brechot, N.; et al. Activation of the UNC5B receptor by Netrin-1 inhibits sprouting angiogenesis. Genes Dev. 2007, 21, 2433–2447. [Google Scholar] [CrossRef] [PubMed]
- Lejmi, E.; Leconte, L.; Pedron-Mazoyer, S.; Ropert, S.; Raoul, W.; Lavalette, S.; Bouras, I.; Feron, J.G.; Maitre-Boube, M.; Assayag, F.; et al. Netrin-4 inhibits angiogenesis via binding to neogenin and recruitment of UNC5B. Proc. Natl. Acad. Sci. USA 2008, 105, 12491–12496. [Google Scholar] [CrossRef] [PubMed]
- Uebelhoer, M.; Natynki, M.; Kangas, J.; Mendola, A.; Nguyen, H.L.; Soblet, J.; Godfraind, C.; Boon, L.M.; Eklund, L.; Limaye, N.; et al. Venous malformation-causative TIE2 mutations mediate an AKT-dependent decrease in PDGFB. Hum. Mol. Genet. 2013, 22, 3438–3448. [Google Scholar] [CrossRef] [PubMed]
- Tsoyi, K.; Jang, H.J.; Nizamutdinova, I.T.; Park, K.; Kim, Y.M.; Kim, H.J.; Seo, H.G.; Lee, J.H.; Chang, K.C. PTEN differentially regulates expressions of ICAM-1 and VCAM-1 through PI3K/Akt/GSK-3beta/GATA-6 signaling pathways in TNF-alpha-activated human endothelial cells. Atherosclerosis 2010, 213, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Morello, F.; Perino, A.; Hirsch, E. Phosphoinositide 3-kinase signalling in the vascular system. Cardiovasc. Res. 2009, 82, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Potente, M.; Urbich, C.; Sasaki, K.; Hofmann, W.K.; Heeschen, C.; Aicher, A.; Kollipara, R.; DePinho, R.A.; Zeiher, A.M.; Dimmeler, S. Involvement of Foxo transcription factors in angiogenesis and postnatal neovascularization. J. Clin. Investig. 2005, 115, 2382–2392. [Google Scholar] [CrossRef] [PubMed]
- Ustyuzhanina, N.E.; Bilan, M.I.; Ushakova, N.A.; Usov, A.I.; Kiselevskiy, M.V.; Nifantiev, N.E. Fucoidans: Pro- or antiangiogenic agents? Glycobiology 2014, 24, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Nardella, A.; Chaubet, F.; Boisson-Vidal, C.; Blondin, C.; Durand, P.; Jozefonvicz, J. Anticoagulant low molecular weight fucans produced by radical process and ion exchange chromatography of high molecular weight fucans extracted from the brown seaweed Ascophyllum nodosum. Carbohydr. Res. 1996, 289, 201–208. [Google Scholar] [CrossRef]
- Chevolot, L.; Foucault, A.; Chaubet, F.; Kervarec, N.; Sinquin, C.; Fisher, A.M.; Boisson-Vidal, C. Further data on the structure of brown seaweed fucans: Relationships with anticoagulant activity. Carbohydr. Res. 1999, 319, 154–165. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouvard, C.; Galy-Fauroux, I.; Grelac, F.; Carpentier, W.; Lokajczyk, A.; Gandrille, S.; Colliec-Jouault, S.; Fischer, A.-M.; Helley, D. Low-Molecular-Weight Fucoidan Induces Endothelial Cell Migration via the PI3K/AKT Pathway and Modulates the Transcription of Genes Involved in Angiogenesis. Mar. Drugs 2015, 13, 7446-7462. https://doi.org/10.3390/md13127075
Bouvard C, Galy-Fauroux I, Grelac F, Carpentier W, Lokajczyk A, Gandrille S, Colliec-Jouault S, Fischer A-M, Helley D. Low-Molecular-Weight Fucoidan Induces Endothelial Cell Migration via the PI3K/AKT Pathway and Modulates the Transcription of Genes Involved in Angiogenesis. Marine Drugs. 2015; 13(12):7446-7462. https://doi.org/10.3390/md13127075
Chicago/Turabian StyleBouvard, Claire, Isabelle Galy-Fauroux, Françoise Grelac, Wassila Carpentier, Anna Lokajczyk, Sophie Gandrille, Sylvia Colliec-Jouault, Anne-Marie Fischer, and Dominique Helley. 2015. "Low-Molecular-Weight Fucoidan Induces Endothelial Cell Migration via the PI3K/AKT Pathway and Modulates the Transcription of Genes Involved in Angiogenesis" Marine Drugs 13, no. 12: 7446-7462. https://doi.org/10.3390/md13127075