
Effects of Cylindrospermopsin Producing Cyanobacterium and Its Crude Extracts on a Benthic Green Alga—Competition or Allelopathy?

Abstract
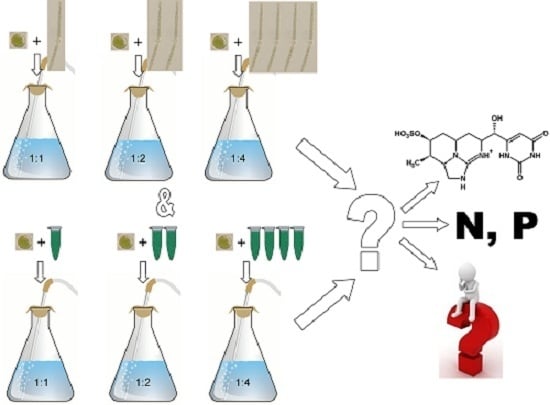
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Coexistence of Chrysosporum ovalisporum and Chlorococcum sp. in Mixed Cultures
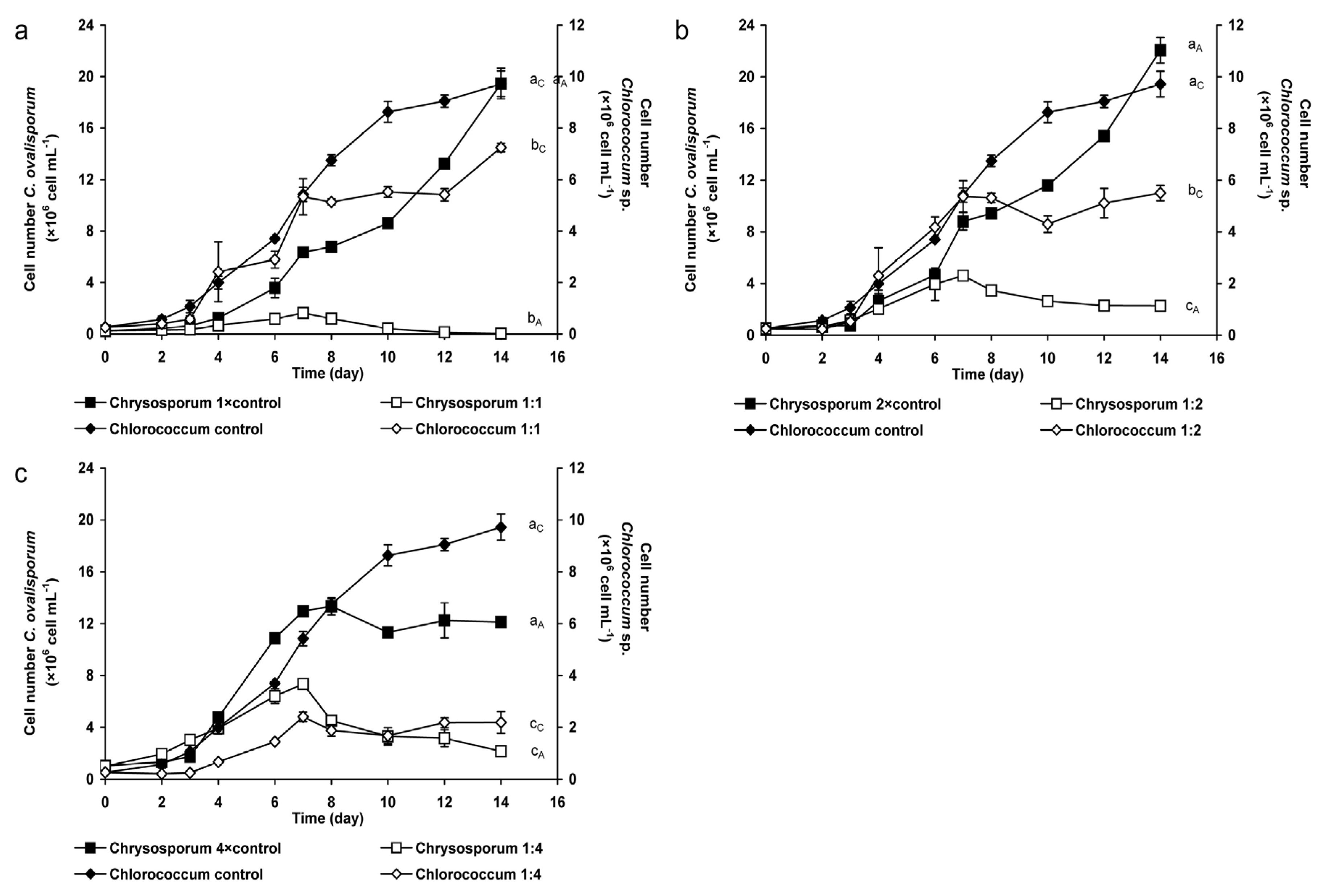
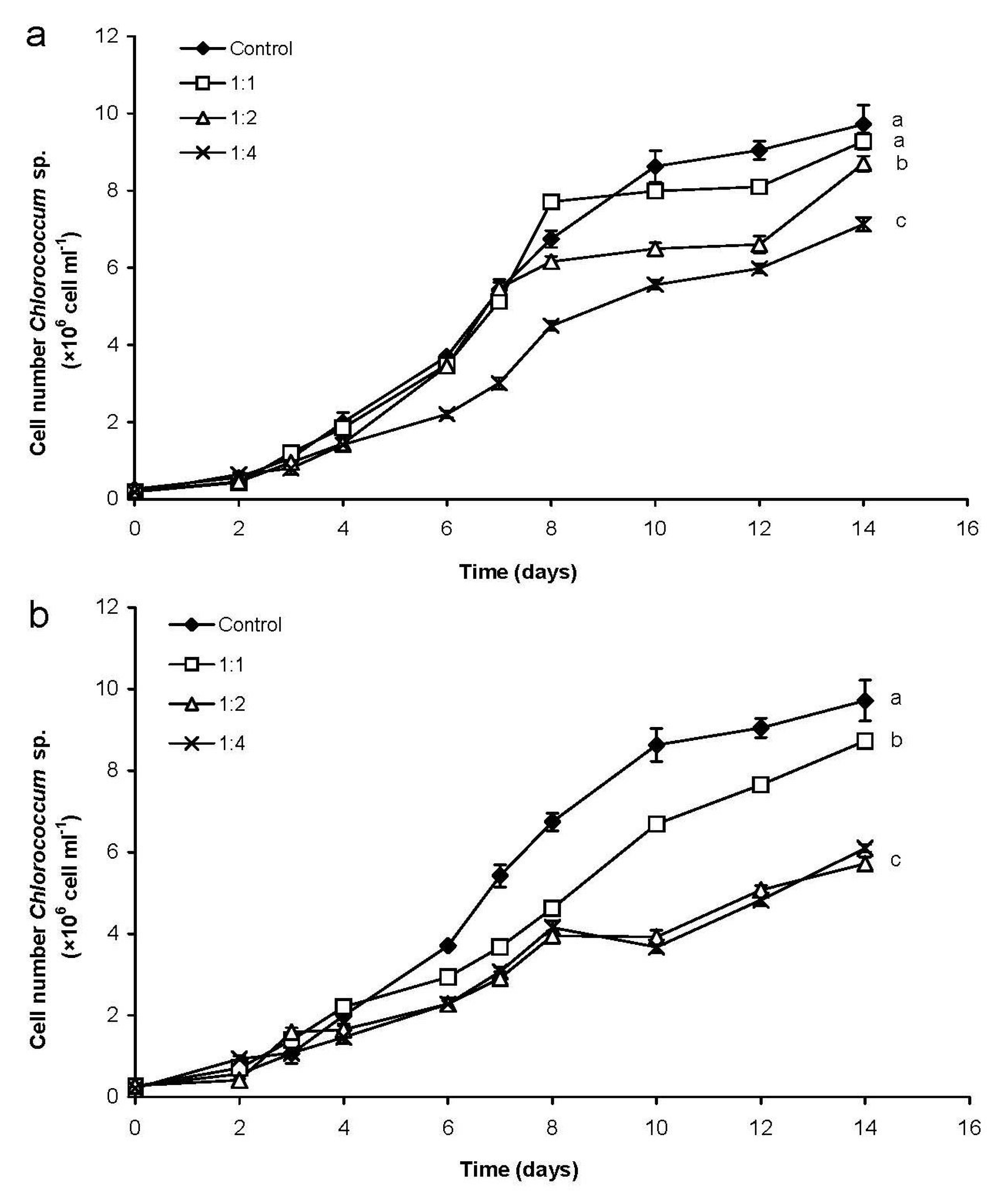
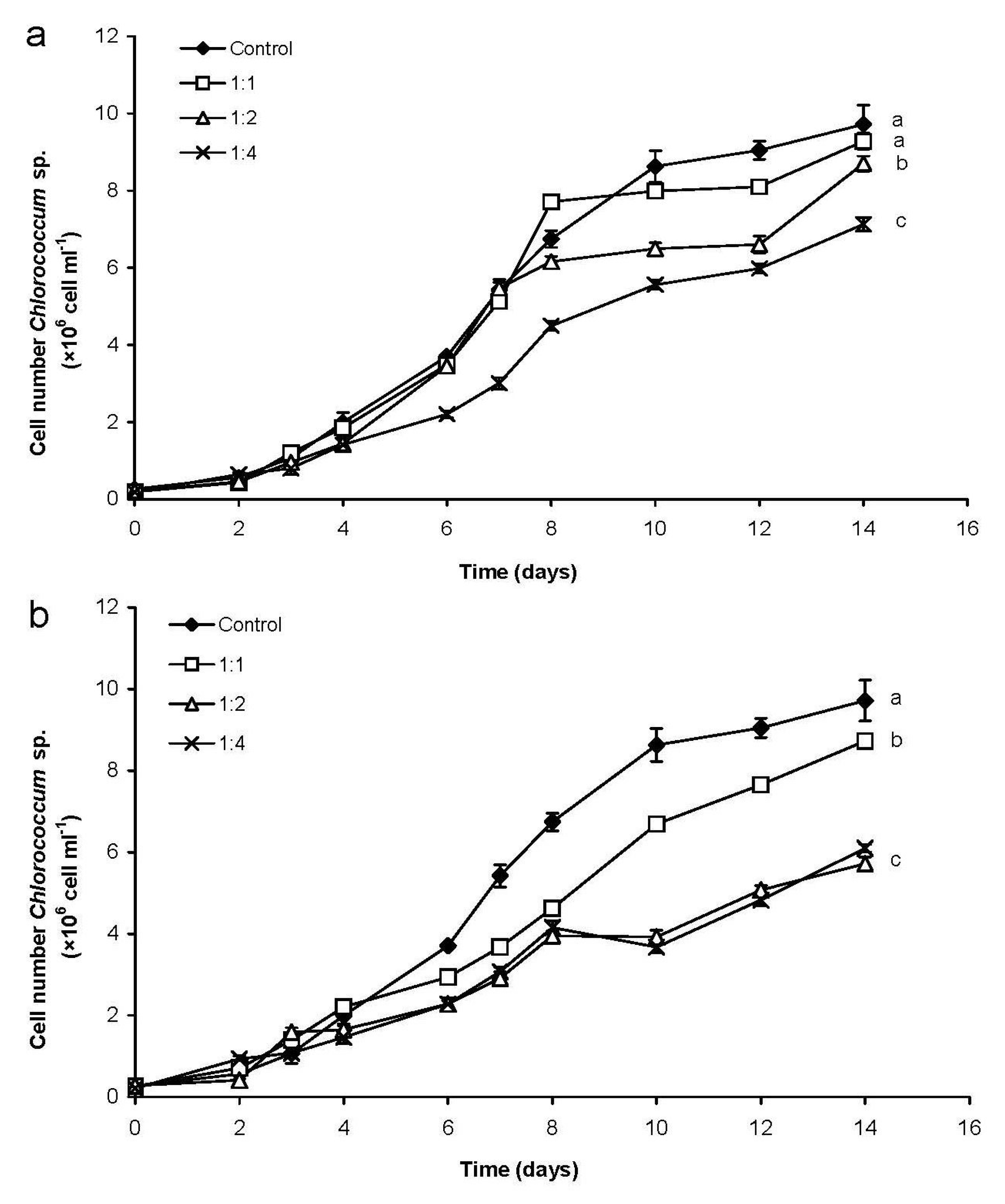
2.1.1. Changes of Eukaryotic Algal and Cyanobacterial Cell Numbers

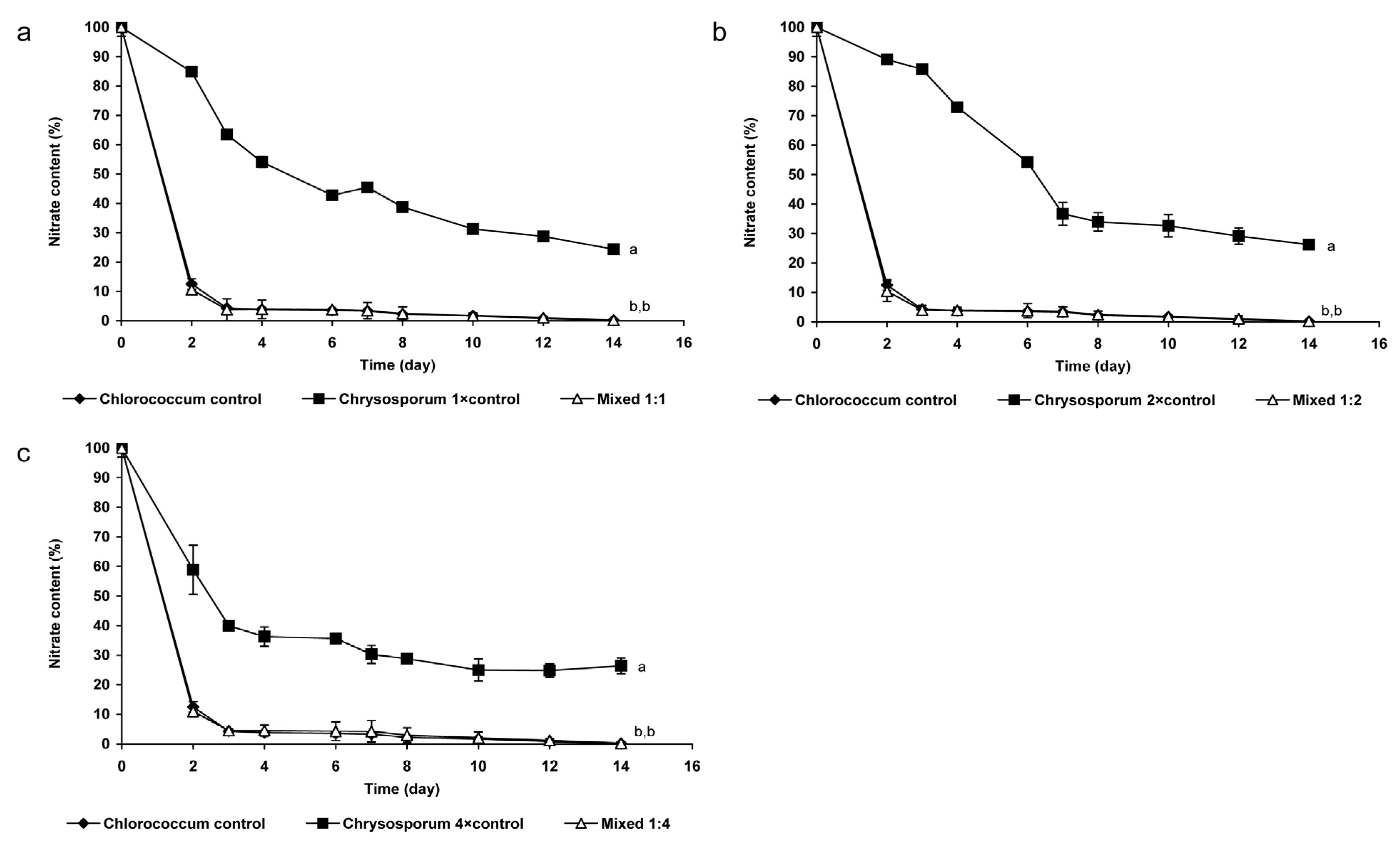
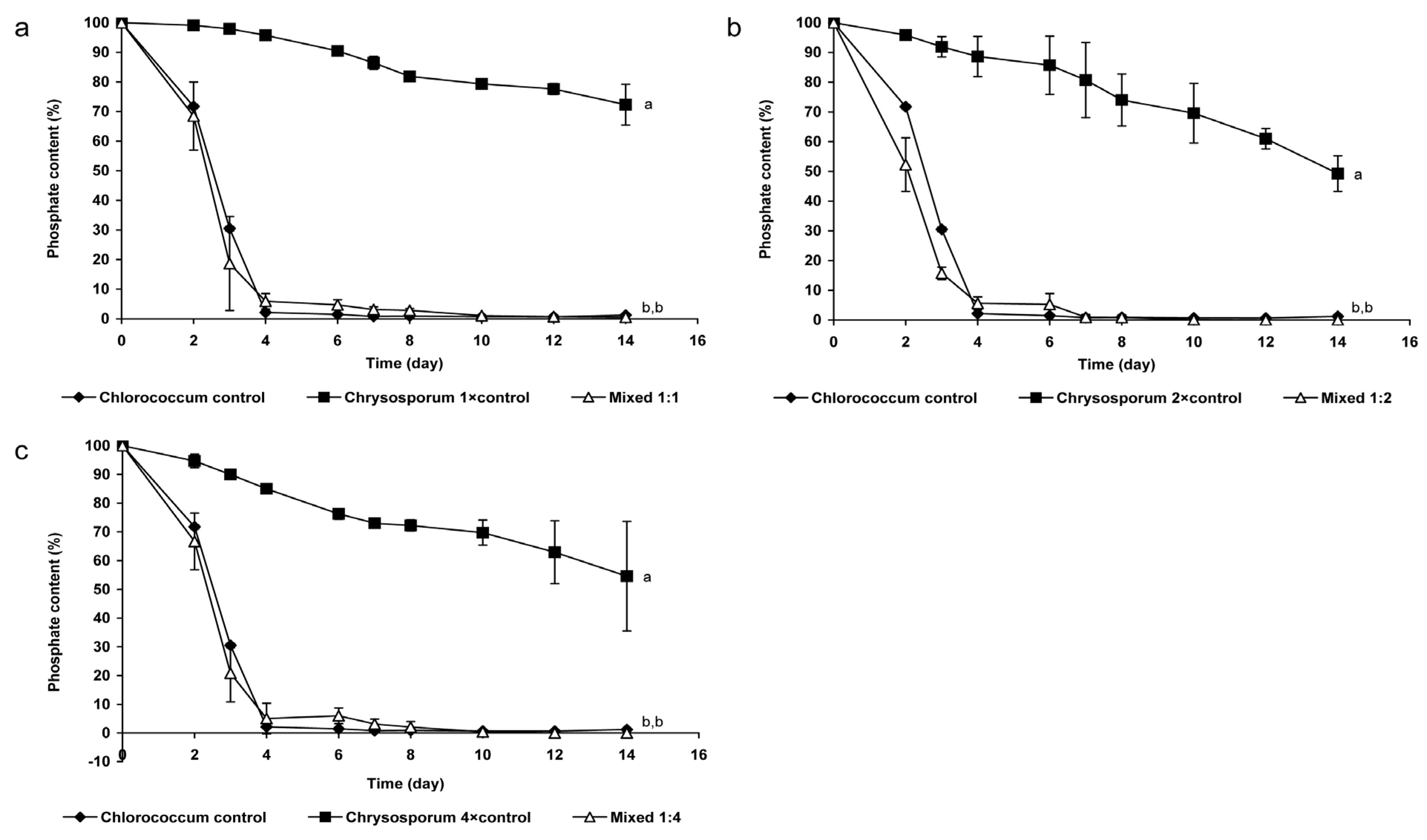
2.1.2. Changes of Nitrate- and Phosphate Content of the Medium
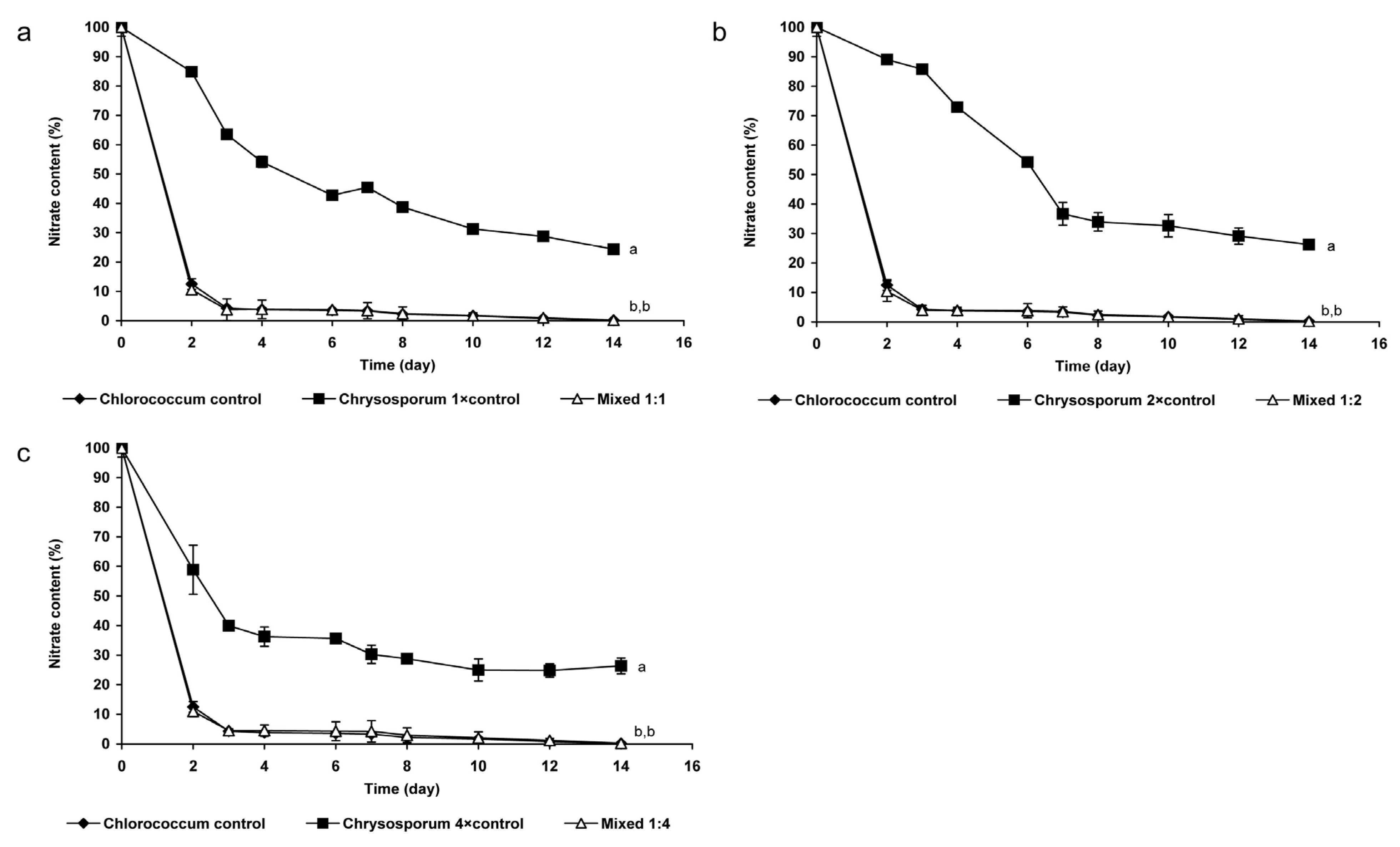

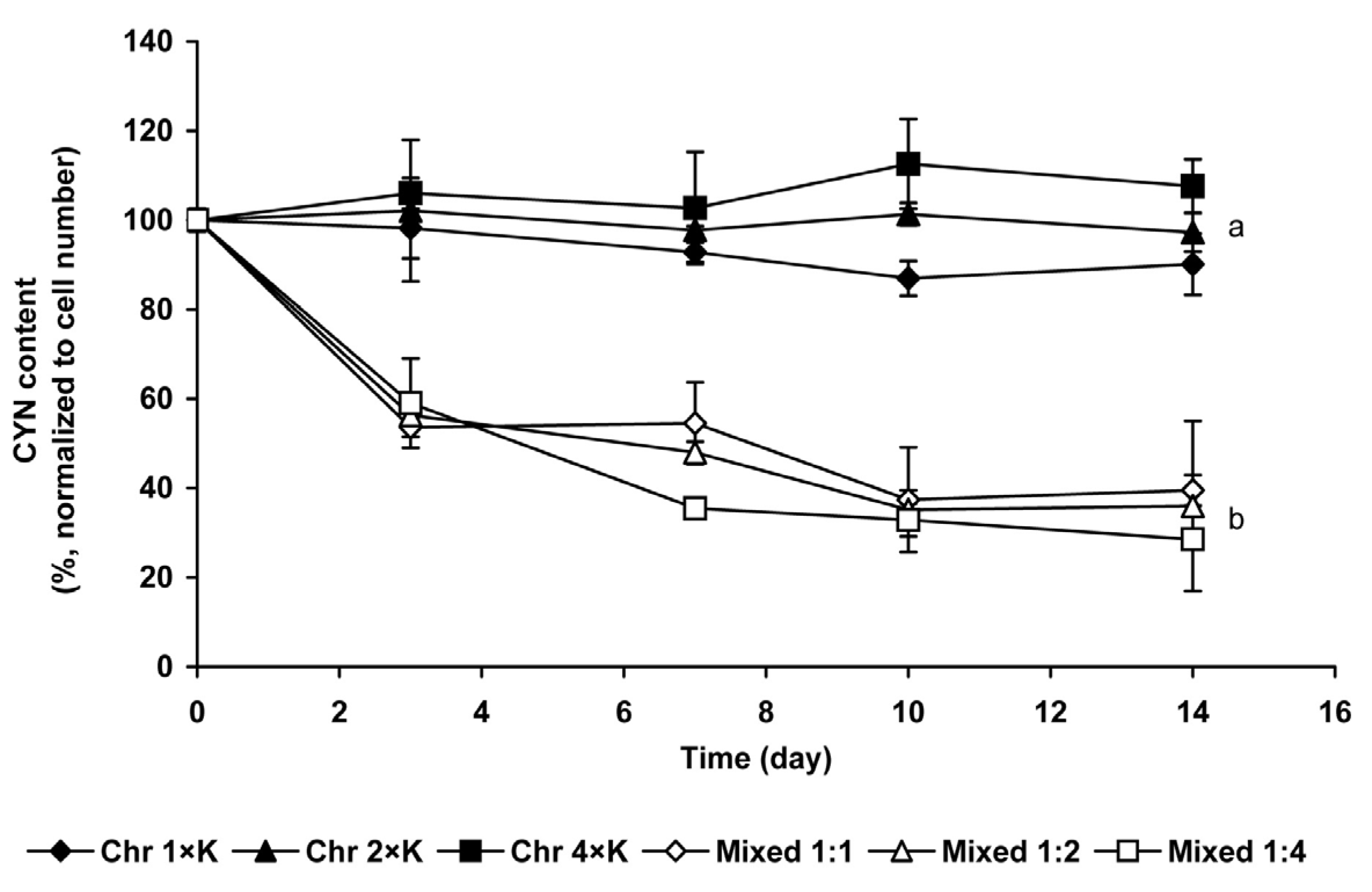
2.1.3. Changes of Extra- and Intracellular CYN Content
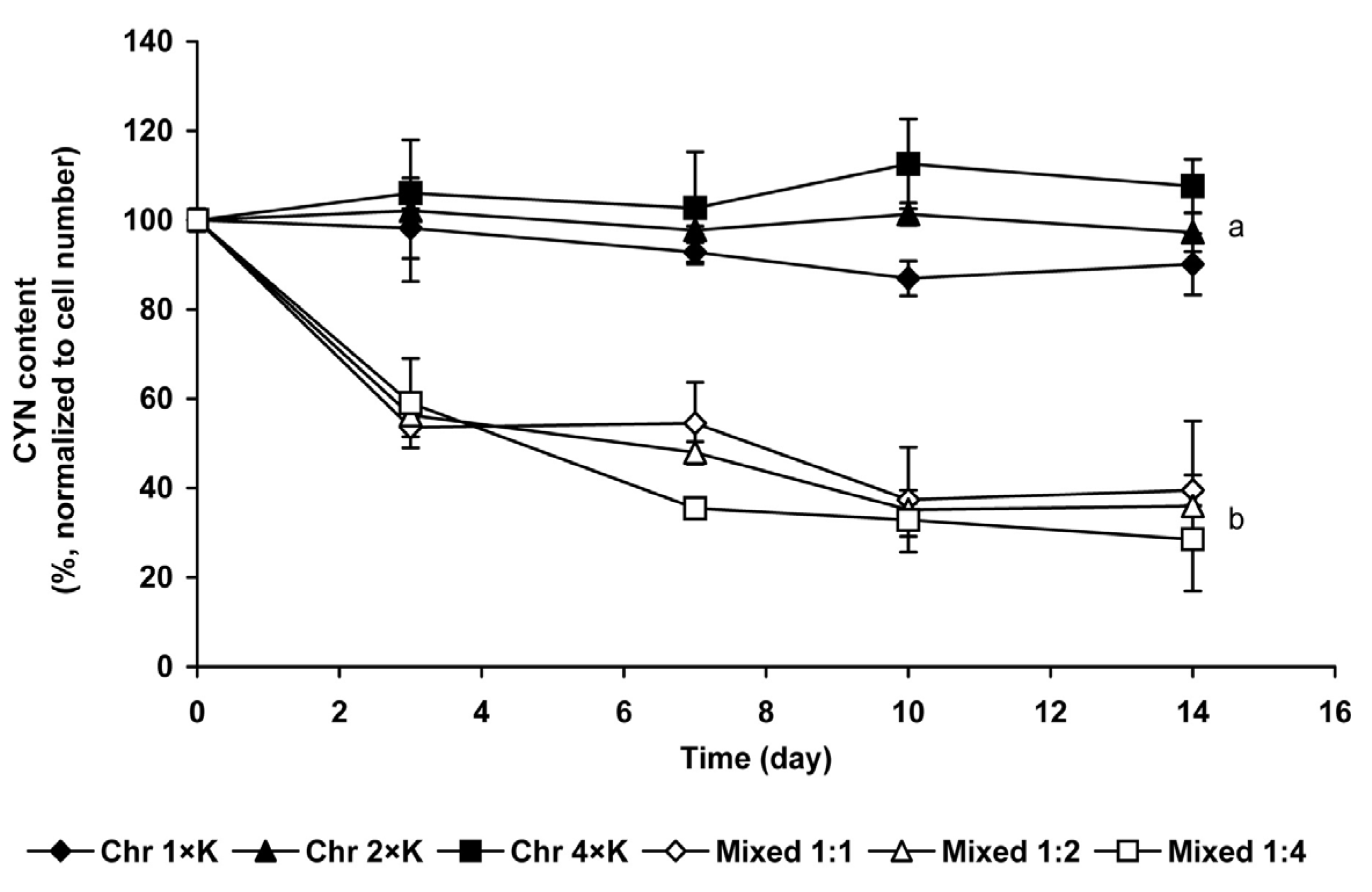
2.2. Chlorococcum sp. Cultures Treated with Cyanobacterial Crude Extract
2.2.1. Changes of Chlorococcum sp. Cell Number
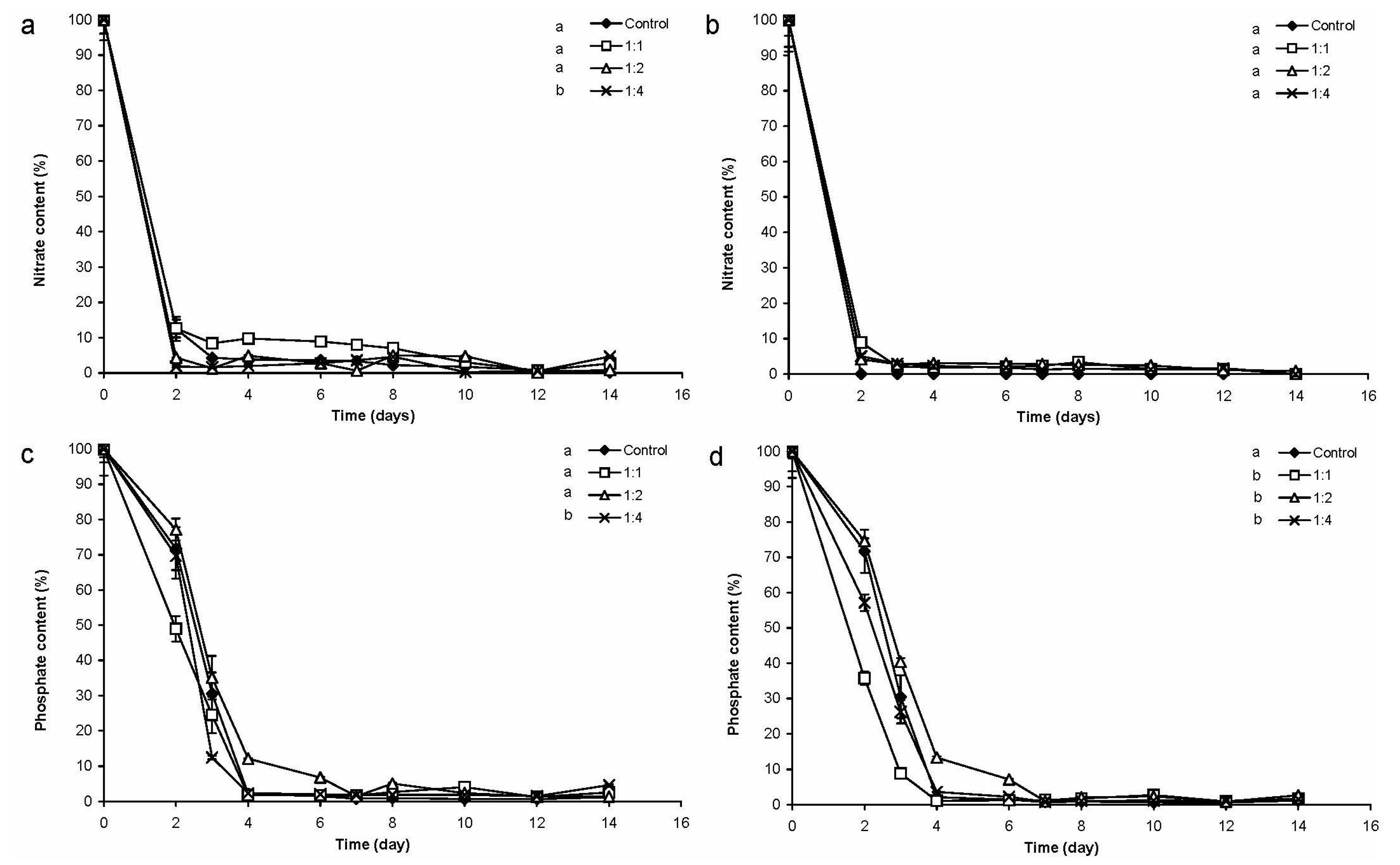
2.2.2. Changes of Nitrate- and Phosphate-Content of the Medium
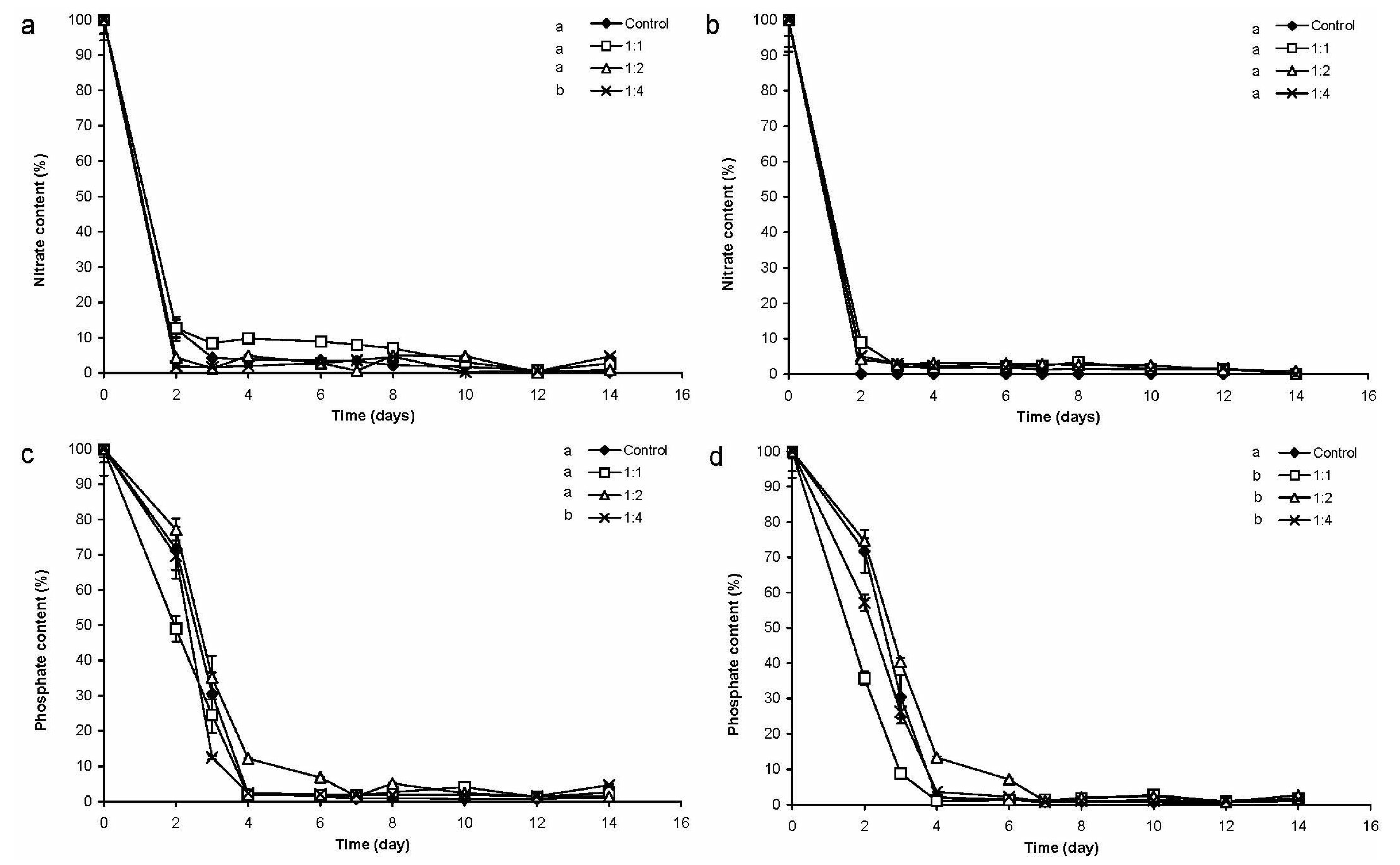
2.2.3. Changes of CYN Concentration
2.3. Comparison the Changes of Different Chlorococcum Cultures
3. Discussion
3.1. Coexistence of Chrysosporum ovalisporum and Chlorococcum sp. in Mixed Cultures
3.2. Chlorococcum sp. Cultures Treated with Cyanobacterial Crude Extract
4. Experimental Section
4.1. Strains and Culturing Conditions
4.1.1. Mixed Cultures
4.1.2. Crude Extract Treated Cultures, Preparation of Cyanobacterial Crude Extract
4.2. Nutrient Measurements
4.3. Quantification of Cylindrospermopsin Content of C. ovalisporum Cells and Culturing Medium
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Leflaive, J.; Ten-Hage, L. Algal and cyanobacterial secondary metabolites in freshwaters, a comparison of allelopathic compounds and toxins. Freshw. Biol. 2007, 52, 199–214. [Google Scholar] [CrossRef]
- Bhagavathy, S.; Sumathi, P.; Sherene, J.B., I. Green algae Chlorococcum humicola—A new source of bioactive compounds with antimicrobial activity. Asian Pac. J. Trop. Biomed. 2011, 1, S1–S7. [Google Scholar] [CrossRef]
- Larned, S.T. A prospectus for periphyton, recent and future ecological research. J. North Am. Benthol. Soc. 2010, 29, 182–206. [Google Scholar] [CrossRef]
- Leflaive, J.; Lacroix, G.; Nicaise, I.; Ten-Hage, L. Colony induction and growth inhibition in Desmodesmus quadrispina (Chlorococcales) by allelochemicals released from the filamentous alga Uronema confervicolum (Ulotrichales). Environ. Microbiol. 2008, 10, 1536–1546. [Google Scholar] [CrossRef] [PubMed]
- El-Sheekh, M.M.; Khairy, H.M.; El-Shenody, R.A. Allelopathic effects of cyanobacterium Microcystis aeruginosa Kützing on the growth and photosynthetic pigments of some algal species. Allelopath. J. 2010, 26, 275–289. [Google Scholar]
- Leao, P.N.; Vasconcelos, M.T.S.D.; Vasconcelos, V.M. Allelopathy in freshwater cyanobacteria. Crit. Rev. Microbiol. 2009, 35, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Pflugmacher, S. Possible allelopathic effects of cyanotoxins, with reference to microcystin-LR, in aquatic ecosystem. Environ. Toxicol. 2002, 17, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, C.; Pflugmacher, S. Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review. Toxicol. Appl. Pharm. 2005, 203, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Babica, P.; Hilscherova, K.; Bartova, K.; Blaha, L.; Marsalek, B. Effects of dissolved microcystins on growth of planktonic photoautotrophs. Phycologia 2007, 46, 137–142. [Google Scholar] [CrossRef]
- Fastner, J.; Heinze, R.; Humpage, A.R.; Mischke, U.; Eaglesham, G.K.; Chorus, I. Cylindrospermopsin occurrence in two German lakes and preliminary assessment of toxicity and toxin production of Cylindrospermopsis raciborskii (Cyanobacteria) isolates. Toxicon 2003, 42, 313–321. [Google Scholar] [CrossRef]
- Rohrlack, T.; Hyenstrand, P. Fate of intracellular microcystins in the cyanobacterium Microcystis aeruginosa (Chroococcales, Cyanophyceae). Phycologia 2007, 46, 277–283. [Google Scholar] [CrossRef]
- Shi, L.; Carmichael, W.W.; Miller, I. Immuno-gold localization of hepatotoxins in cyanobacterial cells. Arch. Microbiol. 1995, 163, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, I.; Moore, R.E.; Runnegar, M.T.C. Cylindrospermopsin: A potent hepatotoxin from the bluegreen alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc. 1992, 114, 7941–7942. [Google Scholar] [CrossRef]
- Harada, K.I.; Ohtani, I.; Iwamoto, K.; Suzuki, M.; Watanabe, M.F.; Watanabe, M.; Terao, K. Isolation of cylindrospermopsin from a cyanobacterium Umezakia natans and its screening method. Toxicon 1994, 32, 73–84. [Google Scholar] [CrossRef]
- Banker, R.; Carmeli, S.; Hadas, O.; Teltsch, B.; Porat, R.; Sukenik, A. Identification of cylindrospermopsin in Aphanizomenon ovalisporum (Cyanophyceae) isolated from Lake Kinneret, Israel. J. Phycol. 1997, 33, 613–616. [Google Scholar] [CrossRef]
- Preussel, K.; Stuken, A.; Wiedner, C.; Chorus, I.; Fastner, J. First report on cylindrospermopsin producing Aphanizomenon flos-aquae (Cyanobacteria) isolated from two German lakes. Toxicon 2006, 47, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Rücker, J.; Stüken, A.; Noxforf, B.; Fastner, J.; Chorus, I.; Wiedner, C. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon-dominated temperate lakes. Toxicon 2007, 50, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Kokociński, M.; Mankiewicz-Boczek, J.; Jurczak, T.; Spoof, L.; Meriluoto, J.; Rejmonczyk, E.; Hautala, H.; Vehniäinen, M.; Pawełczyk, J.; Soininen, J. Aphanizomenon gracile (Nostocales), a cylindrospermopsin-producing cyanobacterium in Polish lakes. Environ. Sci. Pollut. Res. Int. 2013, 20, 5243–5264. [Google Scholar] [CrossRef] [PubMed]
- Bláhová, L.; Babica, P.; Adamovský, O.; Kohoutek, J.; Maršálek, B.; Bláha, L. Analyses of cyanobacterial toxins (microcystins, cylindrospermopsin) in the reservoirs of the Czech Republic and evaluation of health risks. Environ. Chem. Lett. 2008, 6, 223–227. [Google Scholar] [CrossRef]
- Li, R.; Carmichael, W.W.; Brittain, S.; Eaglesham, G.K.; Shaw, G.R.; Liu, Y.; Watanabe, M.M. First report of the cyanotoxins cylindrospermopsin and deoxycylindrospermopsin from Raphidiopsis curvata (Cyanobacteria). J. Phycol. 2001, 37, 1121–1126. [Google Scholar] [CrossRef]
- McGregor, G.B.; Sendall, B.C.; Hunt, L.T.; Eaglesham, G.F. Report of the cyanotoxins cylindrospermopsin and deoxycylindrospermopsin from Raphidiopsis mediterranea Skuja (Cyanobacteria/Nostocales). Harmful Algae 2011, 10, 402–410. [Google Scholar] [CrossRef]
- Schembri, M.A.; Neilan, B.A.; Saint, C.P. Identification of genes implicated in toxin production in the cyanobacterium Cylindrospermopsis raciborskii. Environ. Toxicol. 2001, 16, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Brient, L.; Lengronne, M.; Bormans, M.; Fastner, J. First occurrence of cylindrospermopsin in freshwater in France. Environ. Toxicol. 2009, 24, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Spoof, L.; Berg, K.A.; Rapala, J.; Lahti, K.; Lepistö, L.; Metcalf, J.S.; Codd, G.A.; Meriluoto, J. First observation of cylindrospermopsin in Anabaena lapponica isolated from the boreal environment (Finland). Environ. Toxicol. 2006, 21, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Seifert, M.; McGregor, G.; Eaglesham, G.; Wickramasinghe, W.; Shaw, G. First evidence for the production of cylindrospermopsin and deoxy-cylindrospermopsin by the freshwater benthic cyanobacterium Lyngbya wollei (Farlow ex Gomont) Speziale and Dyck. Harmful Algae 2007, 6, 73–80. [Google Scholar] [CrossRef]
- Mazmouz, R.; Chapuis-Hugon, F.; Mann, S.; Pichon, V.; Mejean, A.; Ploux, O. Biosynthesis of cylindrospermopsin and 7-epicylindrospermopsin in Oscillatoria sp. strain PCC 6506: Identification of the cyr gene cluster and toxin analysis. Appl. Environ. Microbiol. 2010, 76, 4943–4949. [Google Scholar] [CrossRef] [PubMed]
- Akcaalan, R.; Köker, L.; Oğuz, A.; Spoof, L.; Meriluoto, J.; Albay, M. First Report of Cylindrospermopsin Production by Two Cyanobacteria (Dolichospermum mendotae and Chrysosporum ovalisporum) in Lake Iznik, Turkey. Toxins 2014, 6, 3173–3186. [Google Scholar] [CrossRef] [PubMed]
- Bohunická, M.; Mareš, J.; Hrouzek, P.; Urajová, P.; Lukeš, M.; Šmarda, J.; Komárek, J.; Gaysina, L.A.; Sturnecký, O. A combined morphological, ultrastructural, molecular, and biochemical study of the peculiar family Gomontiellaceae (Oscillatoriales) reveals a new cylindrospermopsin-producing clade of cyanobacteria. J. Phycol. 2015. [Google Scholar] [CrossRef]
- Poniedziałek, B.; Rzymski, P.; Kokociński, M. Cylindrospermopsin: Water-linked potential threat to human health in Europe. Environ. Toxicol. Pharmacol. 2012, 34, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Poniedziałek, B. In search of environmental role of cylindrospermopsin: A review on global distribution and ecology of its producers. Water Res. 2014, 66, 320–337. [Google Scholar] [CrossRef] [PubMed]
- Törökné, A.; Asztalos, M.; Bánkiné, M.; Bickel, H.; Borbély, G.; Carmeli, S.; Codd, G.A.; Fastner, J.; Huang, Q.; Humpage, A.; et al. Interlaboratory comparison trial on cylindrospermopsin measurement. Anal. Biochem. 2004, 332, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Vasas, G.; Surányi, G.; Máthé, C.; M-Hamvas, M.; Borbély, G. Investigation of toxin content in Cylindrospermopsis raciborskii (Wołoszyńska) Seenaya and Subba Raju and Aphanizomenon ovalisporum (Forti) strains isolated from shallow lakes of Hungary. Acta Biol. Hung. 2010, 61, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Vehovszky, Á.; Kovács, A.W.; Farkas, A.; Győri, J.; Szabó, H.; Vasas, G. Pharmacological Studies Confirm Neurotoxic Metabolite(s) Produced by the Bloom-Forming Cylindrospermopsis raciborskii in Hungary. Environ. Toxicol. 2013, 30, 501–512. [Google Scholar] [PubMed]
- Rzymski, P.; Poniedziałek, B.; Kokociński, M.; Jurczak, T.; Lipski, D.; Wiktorowicz, K. Interspecific allelopathy in cyanobacteria: Cylindrospermopsin and Cylindrospermopsis raciborskii effect on the growth and metabolism of Microcystis aeruginosa. Harmful Algae 2014, 35, 1–8. [Google Scholar] [CrossRef]
- Bar-Yosef, Y.; Sukenik, A.; Hadas, O.; Viner-Mozzini, Y.; Kaplan, A. Enslavementin the water body by toxic Aphanizomenon ovalisporum, inducing alkalinephosphatase in phytoplanktons. Curr. Biol. 2010, 20, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Araújó, P.; Pinheiro, C.; Azvedo, J.; Osório, H.; Vasconcelos, V. Effects on growth, antioxidant enzyme activity and levels of extracellular proteins in the green alga Chlorella vulgaris exposed to crude cyanobacterial extracts and pure microcystin and cylindrospermopsin. Ecotox. Environ. Saf. 2013, 94, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Figueredo, C.C.; Giani, A.; Bird, D.F. Does allelopathy contribute to Cylindrospermopsis raciborskii (Cyanobacteria) bloom occurrence and geographic expansion? J. Phycol. 2007, 43, 256–265. [Google Scholar] [CrossRef]
- Pinheiro, C.; Azvedo, J.; Campos, A.; Loureiro, S.; Vasconcelos, V. Absence of negative allelopathic effects of cylindrospermopsin and microcystin-LR on selected marine and freshwater phytoplankton species. Hydrobiologia 2013, 705, 27–42. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 26 September 2015).
- Padisák, J. Cylindrospermopsis raciborskii (Woloszyńska) Seenayya et Subba Raju, an expanding highly adaptive blue-green algal species, worldwide distribution and review of its ecology. Arch. Hydrobiol. 1997, 107, 563–593. [Google Scholar]
- Briand, J.F.; Robillot, C.; Quiblier-Lioberas, C.; Humbert, J.F.; Coute, A.; Bernard, C. Environmental context of Cylindrospermopsis raciborskii (Cyanobacteria) blooms in shallow pond in France. Water Res. 2002, 36, 3183–3192. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Al-Shehri, A.M. Assessment of cylindrospermopsin toxin in an arid Saudi lake containing dense cyanobacterial bloom. Environ. Monit. Assess. 2013, 185, 2157–2166. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.L.; Griffith, D.J. Effects of temperature on growth and cylindrospermopsin content of seven isolates of Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) from water bodies in northern Australia. Phycologia 2000, 39, 349–354. [Google Scholar] [CrossRef]
- Bácsi, I.; Vasas, G.; Surányi, G.; M-Havas, M.; Máthé, C.; Tóth, E.; Grigorszky, I.; Gáspár, A.; Tóth, S.; Borbély, G. Alteration of cylindrospermopsin production in sulfate- or phosphate-starved cyanobacterium Aphanizomenon ovalisporum. FEMS Microbiol. Lett. 2006, 259, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Vasas, G.; Surányi, G.; Bácsi, I.; M-Hamvas, M.; Máthé, C.; Gonda, S.; Borbély, G. Alteration of cylindrospermopsin content of Aphanizomenon ovalisporum (Cyanobacteria, Nostocales) due to step-down from combined nitrogen to dinitrogen. Adv. Microbiol. 2013, 3, 557–564. [Google Scholar] [CrossRef]
- Cirés, S.; Wörmer, L.; Timón, J.; Wiedner, C.; Quesada, A. Cylindrospermopsin production and release by the potentially invasive cyanobacterium Aphanizomenon ovalisporum under temperature and light gradients. Harmful Algae 2011, 10, 668–675. [Google Scholar] [CrossRef]
- Preussel, K.; Wessel, G.; Fastner, J.; Chorus, I. Response of cylindrospermopsin production, release in Aphanizomenon flos-aquae (Cyanobacteria) to varying light and temperature conditions. Harmful Algae 2009, 8, 645–650. [Google Scholar] [CrossRef]
- Saker, M.L.; Eaglesham, G.K. The accumulation of cylindrospermopsin from the cyanobacterium Cylindrospermopsis raciborskii in tissues of the redclaw crayfish Cherax quadricarinatus. Toxicon 1999, 37, 1065–1077. [Google Scholar] [CrossRef]
- Shaw, G.R.; Sukenik, A.; Livne, A.; Chiswellm, R.K.; Smith, M.J.; Seawright, A.A.; Norris, R.L.; Eaglesham, G.K.; Moore, M.R. Blooms of the hepatotoxic cyanobacterium, Aphanizomenon ovalisporum (Forti) in newly constructed lakes, Queensland, Australia. Environ. Toxicol. 1999, 14, 167–177. [Google Scholar] [CrossRef]
- Jámbrik, K.; Máthé, C.; Vasas, G.; Bácsi, I.; Surányi, G.; Gonda, S.; Borbély, G.; M-Hamvas, M. Cylindrospermopsin inhibits growth and modulates protease activity int he aquatic plants Lemna minor L. and Wolffia arrhiza (L.) Horkel. Acta Biol. Hung. 2010, 61, 77–94. [Google Scholar] [CrossRef] [PubMed]
- B-Béres, V.; Grigorszky, I.; Vasas, G.; Borics, G.; Várbíró, G.; Nagy, S.A.; Borbély, G.; Bácsi, I. The effects of Microcystis aeruginosa (cyanobacterium) on Cryptomonas ovata (Cryptophyta) in laboratory cultures, why these organisms do not coexist in steady-state assemblages? Hydrobiologia 2012, 691, 97–107. [Google Scholar] [CrossRef]
- Vardi, A.; Schatz, D.; Beeri, K.; Motro, U.; Sukenik, A.; Levine, A.; Kaplan, A. Dinoflagellate-Cyanobacterium communication may determine the composition of phytoplankton assemblage in a mesotrophic lake. Curr. Biol. 2002, 12, 1767–1772. [Google Scholar] [CrossRef]
- Kroes, H.W. Extracellular Products from Chlorococcum ellipsoideum and Chlamydomonas globosa. Arch. Mikrobiol. 1972, 84, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Ohta, S.; Shiomi, Y.; Kawashima, A.; Aozasa, O.; Nakao, T.; Nagate, T.; Kitamura, K.; Miyata, H. Antibiotic effect of linolenic acid from Chlorococcum strain HS-101 and Dunaliella perimolecta on methicillin-resistant Staphylococcus aureus. B. Environ. Contam. Toxin 1995, 50, 171–178. [Google Scholar]
- Ohta, S.; Chang, T.; Ikegami, N.; Kondo, M.; Miyata, H. Antibiotic Substance Produced by a Newly Isolated Marine Microalga, Chlorococcum HS-101. B. Environ. Contam. Toxin 1993, 50, 171–178. [Google Scholar] [CrossRef]
- Bartova, K.; Hilscherova, K.; Babica, P.; Marsalek, B.; Blaha, L. Effects of microcystin and complex cyanobacterial samples on the growth and oxidative stress parameters in green alga Pseudokirchneriella subcapitata and comparison with the model oxidative stressor—Herbicide paraquat. Environ. Toxicol. 2011, 26, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Krischke, M.; Roitsch, T.; Hasnain, S. Rapid determination of cytokinins and auxin in cyanobacteria. Curr. Microbiol. 2010, 61, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Sergeeva, E.; Liaimer, A.; Bergman, B. Evidence for production of the phytohormone indole-3-acetic acid by cyanobacteria. Planta 2002, 215, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Stirk, W.A.; Ördög, V.; Van Staden, J.; Jäger, K. Cytokinin- and auxin-like activity in Cyanophyta and microalgae. J. Appl. Phycol. 2002, 14, 215–221. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Klimova, S.Y.; Cherdyntseva, T.A.; Netrusov, A.I. Microbial producers of plant growth stimulators and their practical use, a review. Appl. Biochem. Microbiol. 2006, 42, 117–126. [Google Scholar] [CrossRef]
- CCAP Media Recipes. Available online: http://www.ccap.ac.uk/media/documents/JM.pdf (accessed on 28 October 2015).
- MSZ 1484-13, Hungarian Standard. Spectrophotometric Determination of Nitrate Ion; Hungarian Standard Institution: Budapest, Hungary, 2009.
- MSZ 448-20, Hungarian Standard. Spectrophotometric Determination of Dissolved Inorganic Phosphate; Hungarian Standard Institution: Budapest, Hungary, 1990.
- Vasas, G.; Gáspár, A.; Surányi, G.; Batta, G.; Gyémánt, G.; M-Hamvas, M.; Máthé, M.; Grigorszky, I.; Molnár, E.; Borbély, G. Capillary Electrophoretic Assay and Purification of Cylindrospermopsin, a Cyanobacterial Toxin from Aphanizomenon ovalisporum, by Plant Test (Blue-Green Sinapis Test). Anal. Biochem. 2002, 302, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Hammer, R.; Harper, D.A.T.; Ryan, P.D. PAST, paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 3rd ed.; Prentice-Hall Inc.: Upper Saddle River, NJ, USA, 1996. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
B-Béres, V.; Vasas, G.; Dobronoki, D.; Gonda, S.; Nagy, S.A.; Bácsi, I. Effects of Cylindrospermopsin Producing Cyanobacterium and Its Crude Extracts on a Benthic Green Alga—Competition or Allelopathy? Mar. Drugs 2015, 13, 6703-6722. https://doi.org/10.3390/md13116703
B-Béres V, Vasas G, Dobronoki D, Gonda S, Nagy SA, Bácsi I. Effects of Cylindrospermopsin Producing Cyanobacterium and Its Crude Extracts on a Benthic Green Alga—Competition or Allelopathy? Marine Drugs. 2015; 13(11):6703-6722. https://doi.org/10.3390/md13116703
Chicago/Turabian StyleB-Béres, Viktória, Gábor Vasas, Dalma Dobronoki, Sándor Gonda, Sándor Alex Nagy, and István Bácsi. 2015. "Effects of Cylindrospermopsin Producing Cyanobacterium and Its Crude Extracts on a Benthic Green Alga—Competition or Allelopathy?" Marine Drugs 13, no. 11: 6703-6722. https://doi.org/10.3390/md13116703