Cytotoxic Effect of Clerosterol Isolated from Codium fragile on A2058 Human Melanoma Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
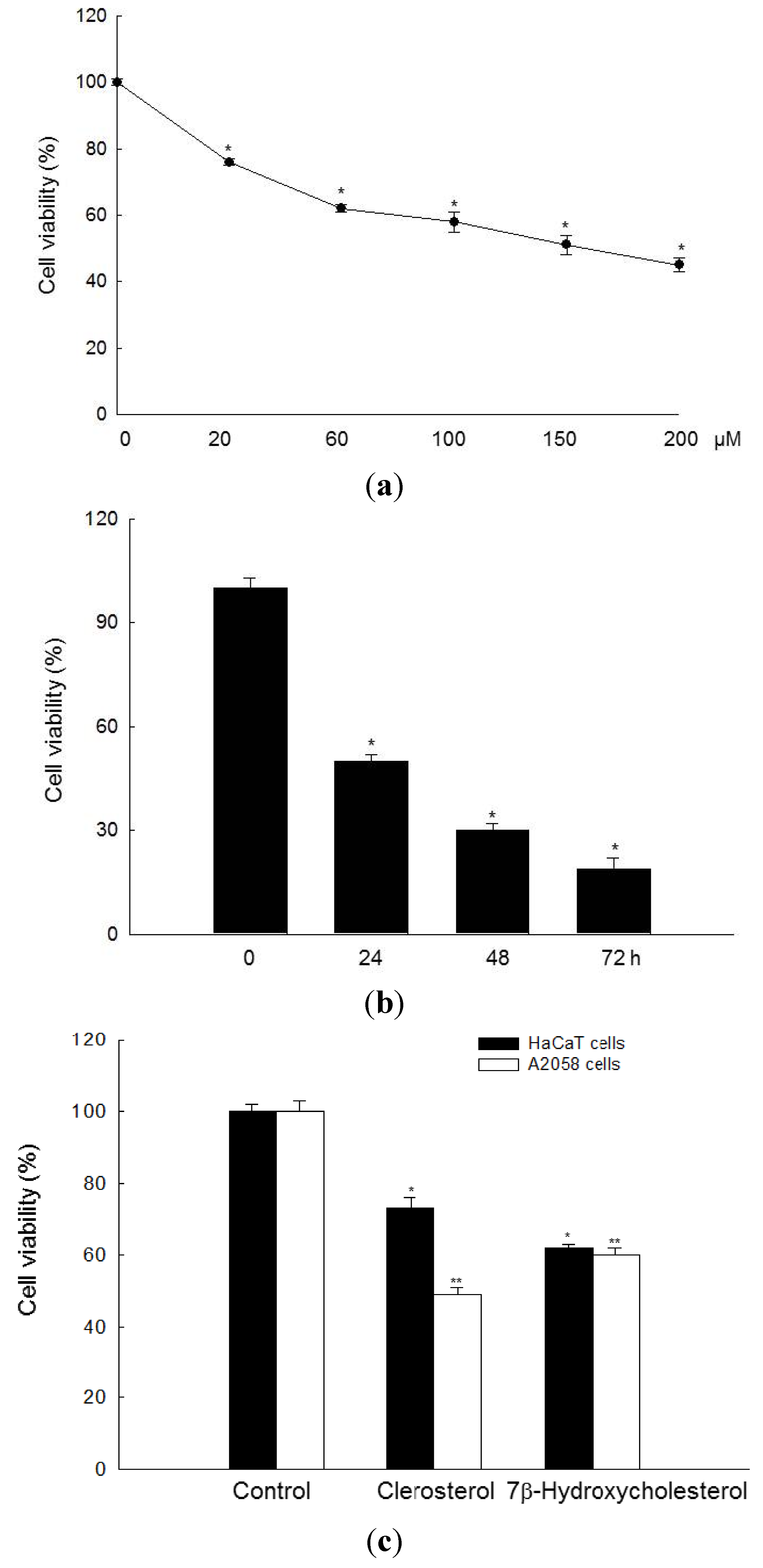
2.1. Clerosterol Inhibits the Growth of A2058 Human Melanoma Cells

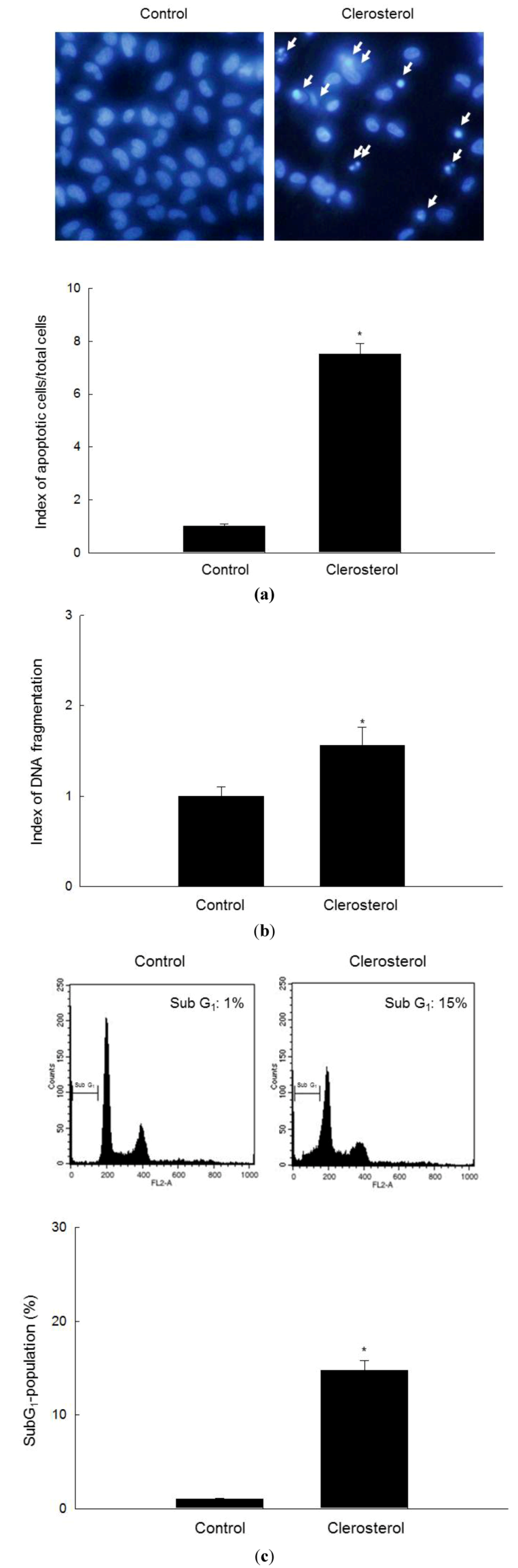
2.2. Clerosterol Induces Apoptotic Cell Death in A2058 Cells

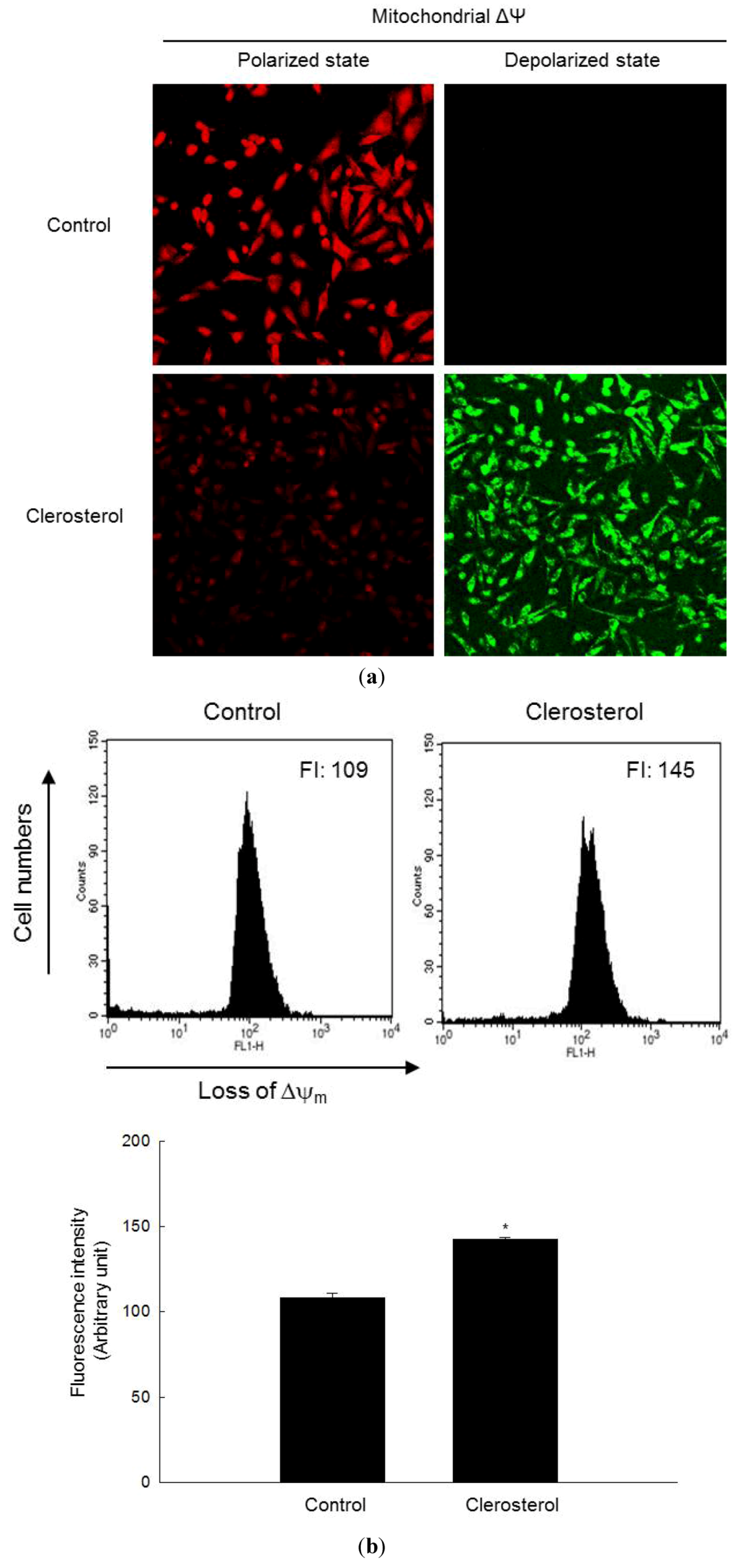
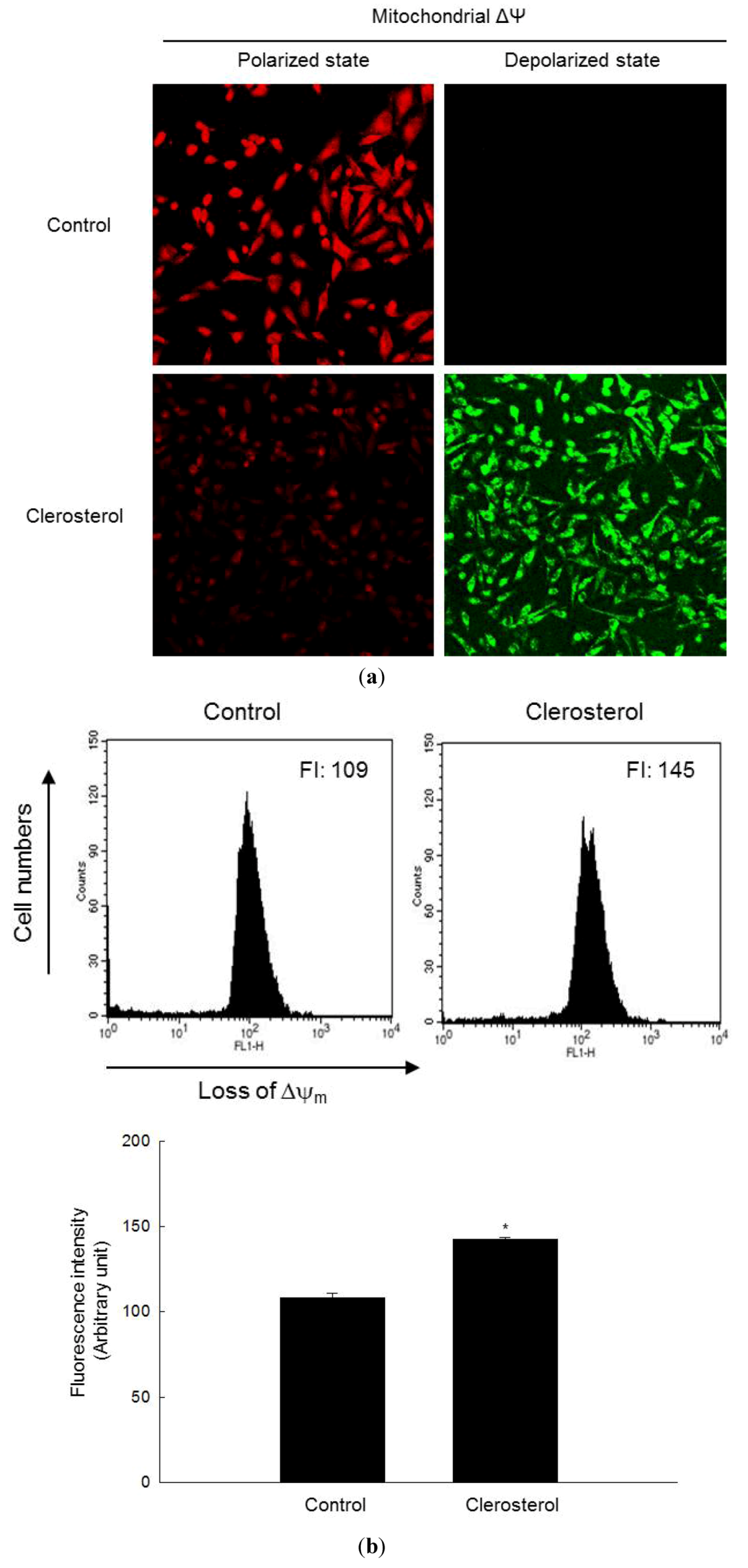
2.3. Clerosterol-Induced Apoptosis Occurs through a Mitochondrial Pathway
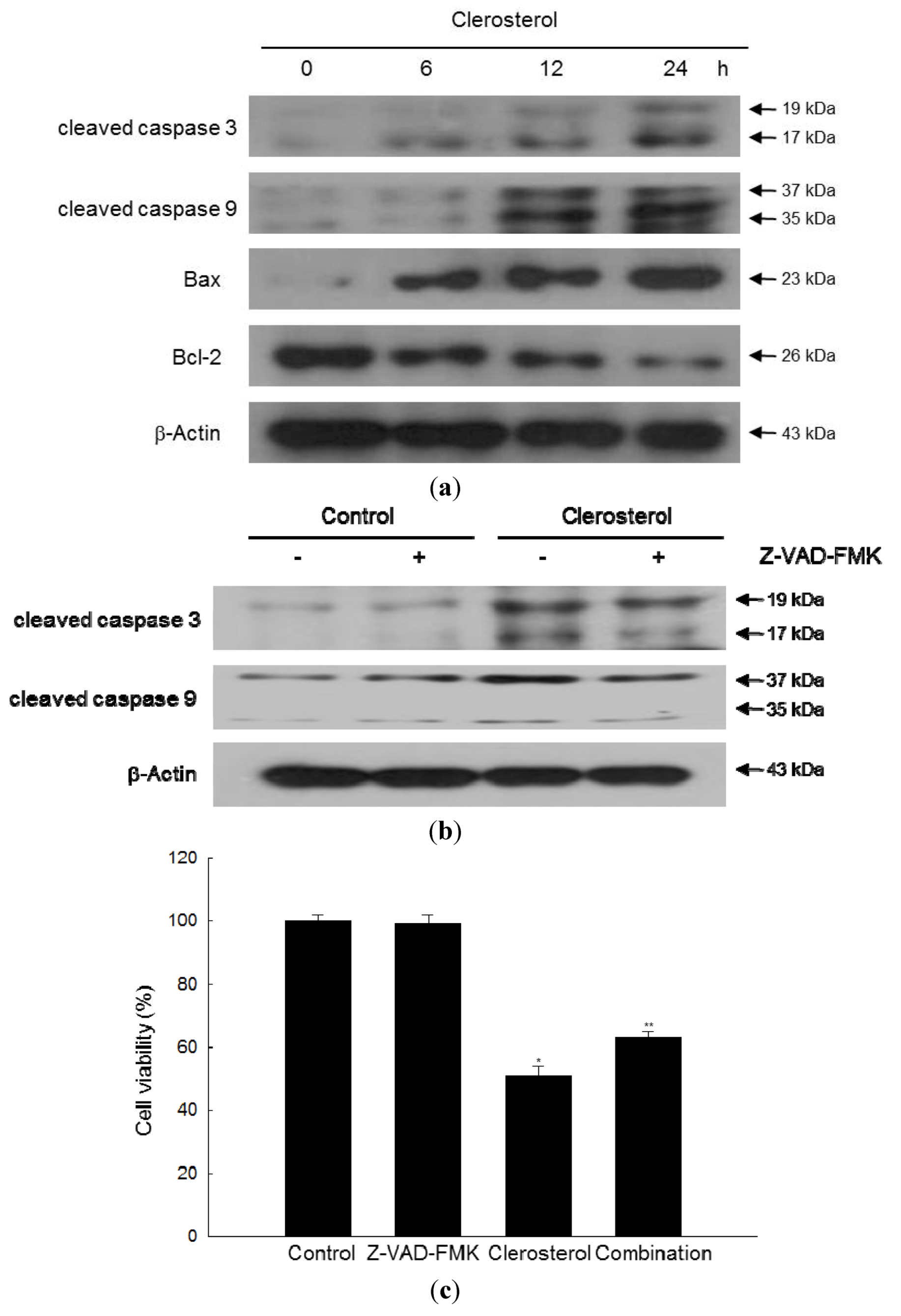
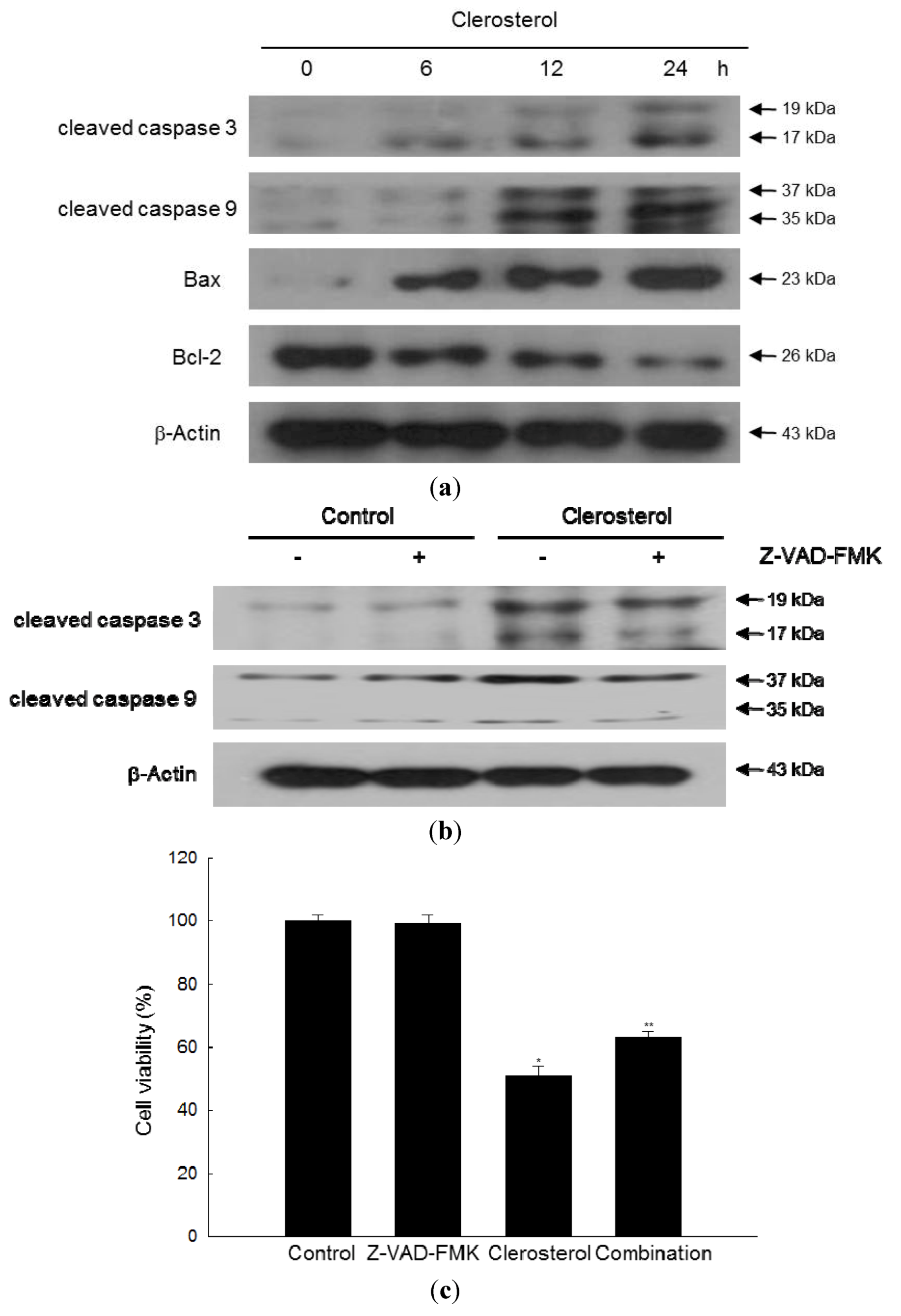
2.4. Clerosterol Induces Apoptosis via Caspase Activation
3. Discussion
4. Experimental Section
4.1. Reagents
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Nuclear Staining with Hoechst 33342
4.5. DNA Fragmentation
4.6. Detection of Sub-G1 Hypodiploid Cells
4.7. Analysis of Mitochondrial Membrane Potential (Δψm)
4.8. Western Blot Analysis
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Hoang, M.T.; Eichenfield, L.F. The rising incidence of melanoma in children and adolescents. Dermatol. Nurs. 2000, 121, 192–193. [Google Scholar]
- Drukala, J.; Rajwa, B.; Pietrzkowski, Z.; Korohoda, W. Comparison of Daunomycin Effects on Human Keratinocytes and Melanoma HTB 140 Cells. Image Cytometry Study. Anticancer Res. 2003, 23, 419–426. [Google Scholar]
- Kaufman, S.H. Induction of endonucleolytic DNA cleavage in human acute myelogenous leukemia cells by etoposide, camptothecin and other cytotoxic anticancer drugs: A cautionary note. Cancer Res. 1989, 49, 5870–5878. [Google Scholar]
- Zhang, X.H.; Zhang, N.; Lu, J.M.; Kong, Q.Z.; Zha, Y.F. Tetrazolium Violet Induced Apoptosis and Cell Cycle Arrest in Human Lung Cancer A549 Cells. Biomol. Ther. 2012, 20, 177–182. [Google Scholar] [CrossRef]
- Huschtscha, L.I.; Bartier, W.A.; Ross, C.E.A.; Tattersall, M.H. Characteristics in cancer death after exposure to cytotoxic drugs in vitro. Br. J. Cancer 1996, 73, 54–60. [Google Scholar]
- Cimino, G.; Ghiselin, M.T. Marine natural products chemistry as an evolutionary narrative. In Marine Chemical Ecology; McClintock, J.B., Baker, B.J., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 115–154. [Google Scholar]
- Kim, S.K.; Karagozlu, M.Z. Marine algae: Natural product source for gastrointestinal cancer treatment. Adv. Food Nutr. Res. 2011, 64, 225–233. [Google Scholar] [CrossRef]
- Kim, S.K.; Ta, Q.V. Potential beneficial effects of marine algal sterols on human health. Adv. Food Nutr. Res. 2011, 64, 191–198. [Google Scholar] [CrossRef]
- Ganesan, P.; Noda, K.; Manabe, Y.; Ohkubo, T.; Tanaka, Y.; Maoka, T.; Sugawara, T.; Hirata, T. Siphonaxanthin, a marine carotenoid from green algae, effectively induces apoptosis in human leukemia (HL-60) cells. Biochim. Biophys. Acta 2011, 1810, 497–503. [Google Scholar] [CrossRef]
- Kim, Y.M.; Kim, I.H.; Nam, T.J. Induction of apoptosis signaling by glycoprotein of Capsosiphon fulvescens in human gastric cancer (AGS) cells. Nutr. Cancer 2012, 64, 761–769. [Google Scholar] [CrossRef]
- Pardhasaradhi, B.V.; Ali, A.M.; Kumari, A.L.; Reddanna, P.; Khar, A. Phycocyanin-mediated apoptosis in AK-5 tumor cells involves down-regulation of Bcl-2 and generation of ROS. Mol. Cancer Ther. 2003, 2, 1165–1170. [Google Scholar]
- Rubinstein, I.; Goad, L.J. Sterols of the siphonous marine alga Codium fragile. Phytochemistry 1974, 13, 481–484. [Google Scholar] [CrossRef]
- Ahmad, V.U.; Aliya, R.; Perveen, S.; Shameel, M. A sterol glycoside from marine green algae Codium iyengarii. Phytochemistry 1992, 31, 1429–1431. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T. Antioxidant Effects of Tannins and Related Polyphenols. ACS Symp. Ser. 1992, 507, 87–97. [Google Scholar]
- Heiduschka, G.; Erovic, B.M.; Vormittag, L.; Skoda, C.; Martinek, H.; Brunner, M.; Ehrenberger, K.; Thurnher, D. 7beta-hydroxycholesterol induces apoptosis and regulates cyclooxygenase 2 in head and neck squamous cell carcinoma. Arch. Otolaryngol. Head Neck Surg. 2009, 135, 261–267. [Google Scholar] [CrossRef]
- Clarion, L.; Schindler, M.; de Weille, J.; Lolmède, K.; Laroche-Clary, A.; Uro-Coste, E.; Robert, J.; Mersel, M.; Bakalara, N. 7β-Hydroxycholesterol-induced energy stress leads to sequential opposing signaling responses and to death of C6 glioblastoma cells. Biochem. Pharmacol. 2012, 83, 37–46. [Google Scholar]
- Green, D.R.; Reed, J.C. Mitochondria and apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef]
- Jerant, A.F.; Johnson, J.T.; Sheridan, C.D.; Caffrey, T.J. Early detection and treatment of skin cancer. Am. Fam. Physician 2000, 62, 357–368. [Google Scholar]
- Koopman, G.; Reutelingsperger, C.P.; Kuijten, G.A.; Keehnen, R.M.; Pals, S.T.; van Oers, M.H. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood 1994, 84, 1415–1420. [Google Scholar]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Costantini, P.; Jacotot, E.; Decaudin, D.; Kroemer, G. Mitochondrion as a novel target of anticancer chemotherapy. J. Natl. Cancer Inst. 2000, 92, 1042–1053. [Google Scholar]
- Debatin, K.M.; Poncet, D.; Kroemer, G. Chemotherapy: Targeting the mitochondrial cell death pathway. Oncogene 2002, 21, 8786–8803. [Google Scholar] [CrossRef]
- Deveraux, Q.L.; Reed, J.C. IAP family proteins—suppressors of apoptosis. Genes Dev. 1999, 13, 239–252. [Google Scholar] [CrossRef]
- Park, C.; Moon, D.O.; Rhu, C.H.; Choi, B.T.; Lee, W.H.; Kim, G.Y.; Choi, Y.H. Beta-sitosterol induces anti-proliferation and apoptosis in human leukemic U937 cells through activation of caspase-3 and induction of Bax/Bcl-2 ratio. Biol. Pharm. Bull. 2007, 30, 1317–1323. [Google Scholar] [CrossRef]
- Oh, K.J.; Barbuto, S.; Pitter, K.; Morash, J.; Walensky, L.D.; Korsmeyer, S.J. A membrane-targeted BID BCL-2 homology 3 peptide is sufficient for high potency activation of BAX in vitro. J. Biol. Chem. 2006, 281, 36999–37008. [Google Scholar]
- Paris, C.; Bertoglio, J.; Breard, J. Lysosomal and mitochondrial pathways in miltefosine-induced apoptosis in U937 cells. Apoptosis 2007, 12, 1257–1267. [Google Scholar] [CrossRef]
- Harada, H.; Noro, T.; Kamei, Y. Selective antitumor activity in vitro from marine algae from Japan coasts. Biol. Pharm. Bull. 1997, 20, 541–546. [Google Scholar]
- Noda, K.; Tanaka, K.; Yamada, A.; Ogata, J.; Tanaka, H.; Shoyama, Y. Simple assay for antitumor immunoactive glycoprotein derived from Chlorella vulgaris strain CK22 using ELISA. Phytother. Res. 2002, 16, 581–585. [Google Scholar] [CrossRef]
- Yusof, Y.A.; Saad, S.M.; Makpol, S.; Shamaan, N.A.; Ngah, W.Z. Hot water extract of Chlorella vulgaris induced DNA damage and apoptosis. Clinics (Sao Paulo) 2010, 65, 1371–1377. [Google Scholar] [CrossRef]
- Sano, T.; Tanaka, Y. Effect of dried, powdered Chlorella vulgaris on experimental atherosclerosis and alimentary hypercholesterolemia in cholesterol-fed rabbits. Artery 1987, 14, 76–84. [Google Scholar]
- Hasegawa, T.; Noda, K.; Kumamoto, S.; Ando, Y.; Yamada, A.; Yoshikai, Y. Chlorella vulgaris culture supernatant (CVS) reduces psychological stress-induced apoptosis in thymocytes of mice. Int. J. Immunopharmacol. 2000, 22, 877–885. [Google Scholar] [CrossRef]
- Mohd Azamai, E.S.; Sulaiman, S.; Mohd Habib, S.H.; Looi, M.L.; Das, S.; Abdul Hamid, N.A.; Wan Ngah, W.Z.; Mohd Yusof, Y.A. Chlorella vulgaris triggers apoptosis in hepatocarcinogenesis-induced rats. J. Zhejiang. Univ. Sci. B 2009, 10, 14–21. [Google Scholar] [CrossRef]
- Kim, K.N.; Kim, S.H.; Kim, W.S.; Kang, S.M.; Lee, K.W.; Lee, W.J.; Park, S.Y.; Kim, S.K.; Jeon, Y.J. Antitumor Activities of Sea Staghorn (Codium fragile) against CT-26 Cells. Food Sci. Biotechnol. 2008, 17, 976–982. [Google Scholar]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a tetrazolium-based semiautomated colorimetric assay: Assessment of chemosensitivity testing. Cancer Res. 1987, 47, 936–942. [Google Scholar]
- Nicoletti, I.; Migliorati, G.; Pagliacci, M.C.; Grignani, F.; Riccardi, C. A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J. Immunol. Methods 1991, 139, 271–279. [Google Scholar] [CrossRef]
- Troiano, L.; Ferraresi, R.; Lugli, E.; Nemes, E.; Roat, E.; Nasi, M.; Pinti, M.; Cossarizza, A. Multiparametric analysis of cells with different mitochondrial membrane potential during apoptosis by polychromatic flow cytometry. Nat. Protoc. 2007, 2, 2719–2727. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, A.D.; Lee, Y.; Kang, S.-H.; Kim, G.Y.; Kim, H.S.; Hyun, J.W. Cytotoxic Effect of Clerosterol Isolated from Codium fragile on A2058 Human Melanoma Cells. Mar. Drugs 2013, 11, 418-430. https://doi.org/10.3390/md11020418
Kim AD, Lee Y, Kang S-H, Kim GY, Kim HS, Hyun JW. Cytotoxic Effect of Clerosterol Isolated from Codium fragile on A2058 Human Melanoma Cells. Marine Drugs. 2013; 11(2):418-430. https://doi.org/10.3390/md11020418
Chicago/Turabian StyleKim, Areum Daseul, Youngki Lee, Sang-Hyuck Kang, Gi Young Kim, Hye Sun Kim, and Jin Won Hyun. 2013. "Cytotoxic Effect of Clerosterol Isolated from Codium fragile on A2058 Human Melanoma Cells" Marine Drugs 11, no. 2: 418-430. https://doi.org/10.3390/md11020418