Activation of the Nuclear Factor E2-Related Factor 2 Pathway by Novel Natural Products Halomadurones A–D and a Synthetic Analogue

Abstract
:1. Introduction
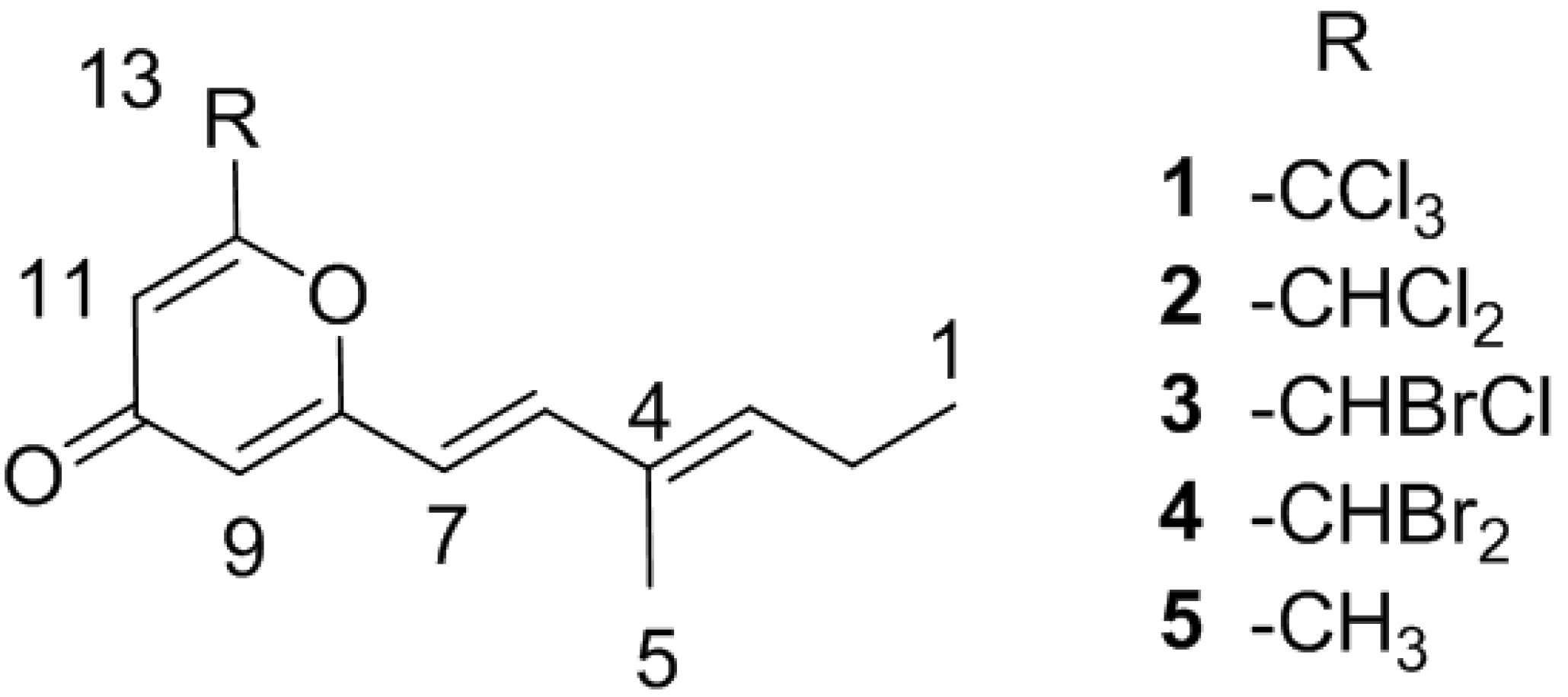
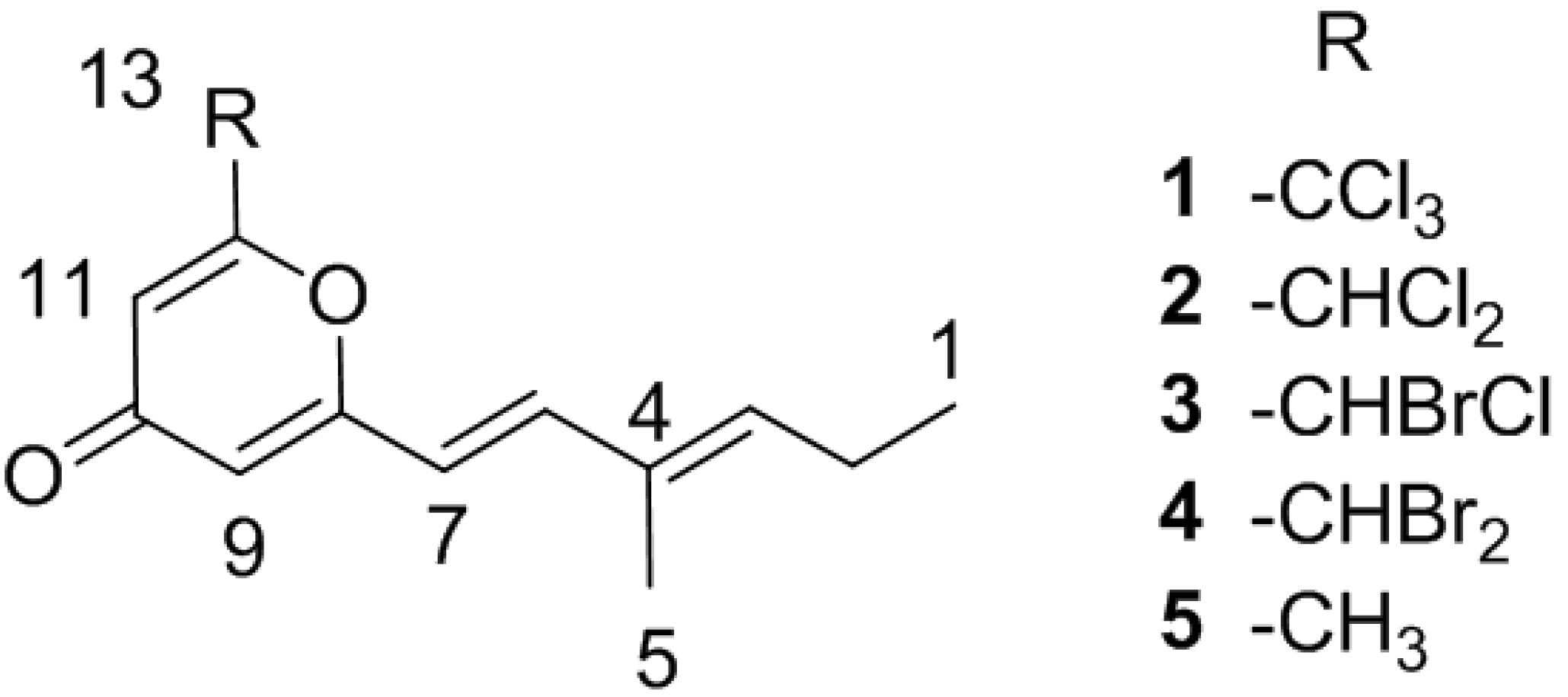
2. Results and Discussion
2.1. Bacterial Strain Selection and Structure Elucidation

{kind=link}
{kind=link}
{kind=link}
| No. | 1 | 2 | 3 | 4 | ||||
|---|---|---|---|---|---|---|---|---|
| δC, mult. | δH (J in Hz) | δC, mult. | δH (J in Hz) | δC, mult. | δH (J in Hz) | δC, mult. | δH (J in Hz) | |
| 1 | 13.8, CH3 | 1.04, t (7.3) | 13.8, CH3 | 1.04, t (7.3) | 13.8, CH3 | 1.04, t (7.5) | 13.8, CH3 | 1.05, t (7.5) |
| 2 | 22.4, CH2 | 2.24, qn (7.3) | 22.4, CH2 | 2.24, qn (7.3) | 22.4, CH2 | 2.24, qn (7.5) | 22.5, CH2 | 2.24, qn (7.5) |
| 3 | 144.3, CH | 5.93, t (7.3) | 144.0, CH | 5.92, t (7.3) | 144.0, CH | 5.93, t (7.2) | 144.0, CH | 5.94, t (7.5) |
| 4 | 132.6, C | 132.6, C | 132.7, C | 132.7, C | ||||
| 5 | 12.2, CH3 | 1.82, s | 12.2, CH3 | 1.82, s | 12.2, CH3 | 1.82, s | 12.2, CH3 | 1.82, s |
| 6 | 143.3, CH | 7.14, d (15.6) | 142.9, CH | 7.14, d (15.6) | 142.9, CH | 7.14, d (15.8) | 142.9, CH | 7.17, d (15.8) |
| 7 | 115.8, CH | 6.07, d (15.6) | 116.1, CH | 6.07, d (15.6) | 116.2, CH | 6.04, d (15.8) | 116.3, CH | 6.05, d (15.8) |
| 8 | 163.2, C | 162.9, C | 163.4, C | 163.4, C | ||||
| 9 | 112.3, CH | 6.17, d (1.8) | 112.7, CH | 6.17, d (2.0) | 112.2, CH | 6.14, d (2.3) | 113.1, CH | 6.13, d (2.3) |
| 10 | 179.5, C | 179.5, C | 179.6, C | 179.5, C | ||||
| 11 | 111.8, CH | 6.85, d (1.8) | 112.7, CH | 6.42, d (2.0) | 112.1, CH | 6.39, d (2.3) | 111.4, CH | 6.32, d (2.3) |
| 12 | 160.5, C | 160.2, C | 160.7, C | 160.8, C | ||||
| 13 | 90.3, C | 65.8, CH | 6.35, s | 50.1, CH | 6.35, s | 32.5, CH | 6.24, s | |
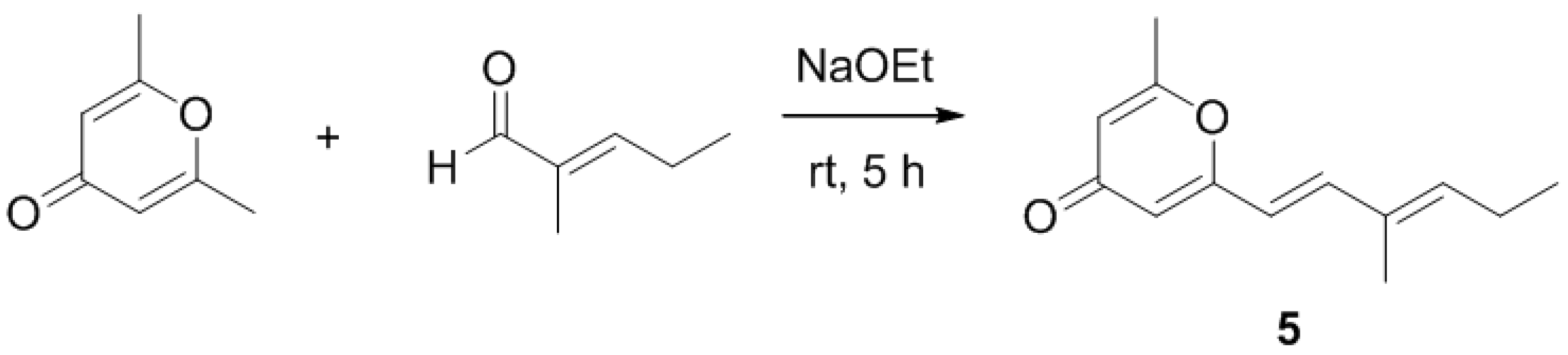
2.2. Synthetic Analogue
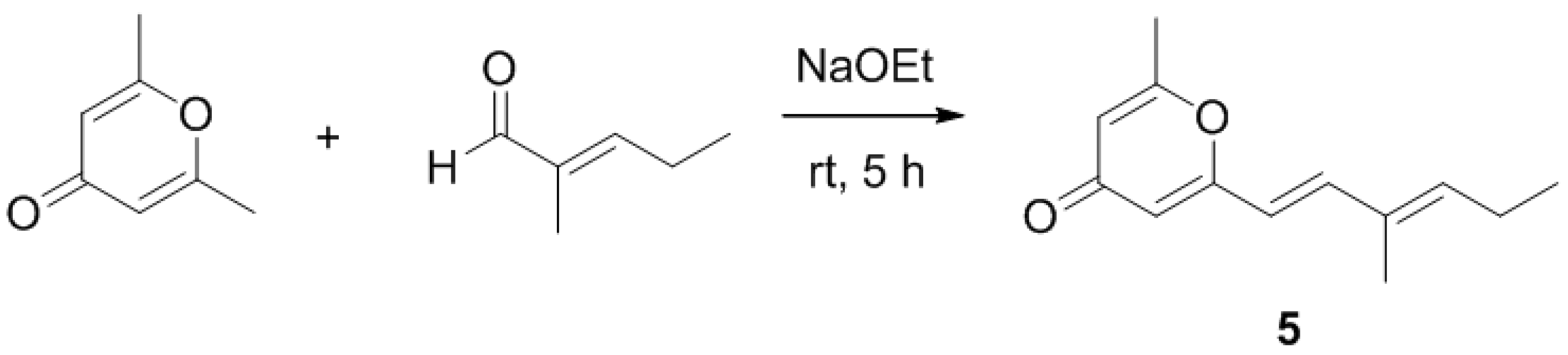
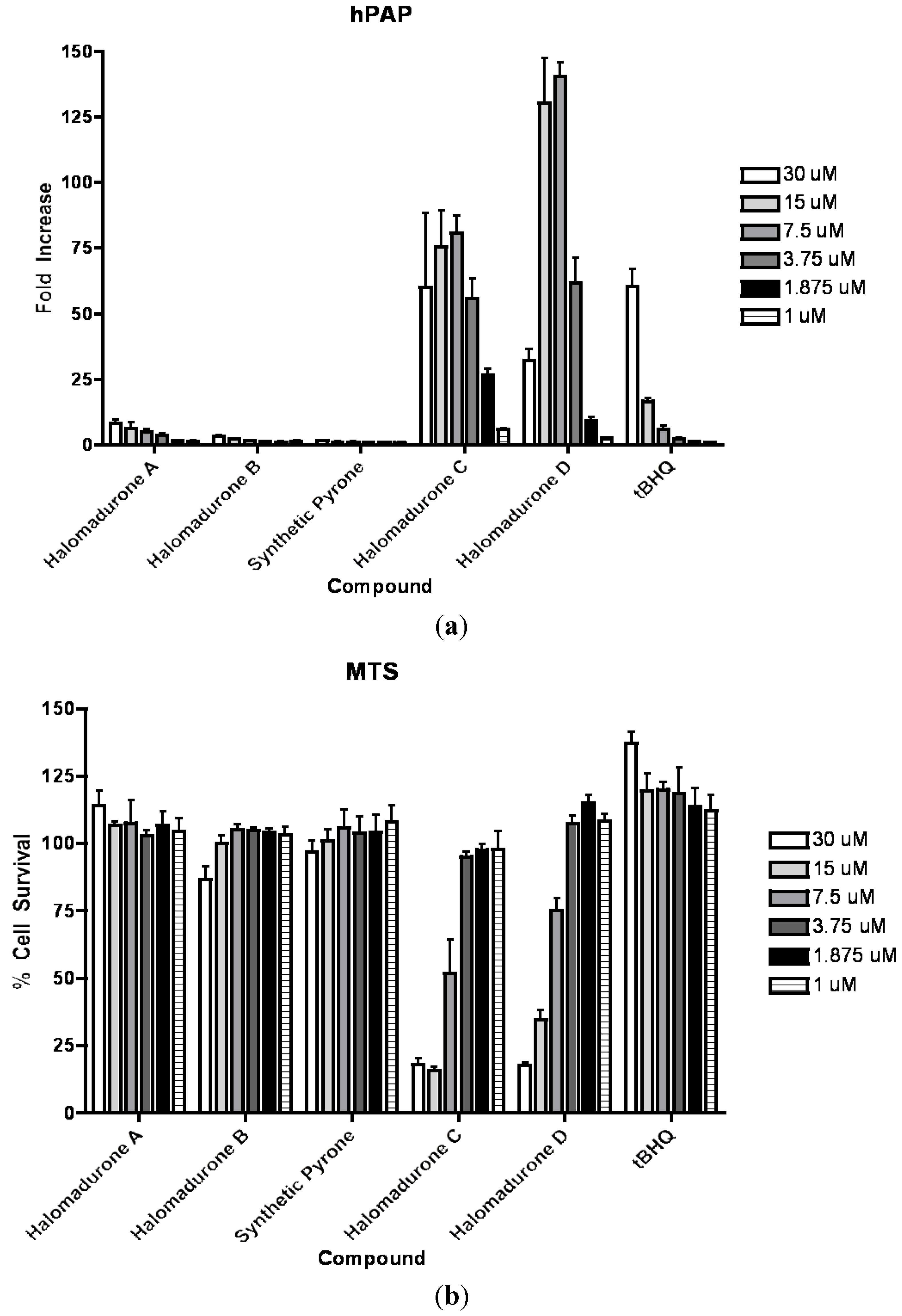
2.3. Biological Activity
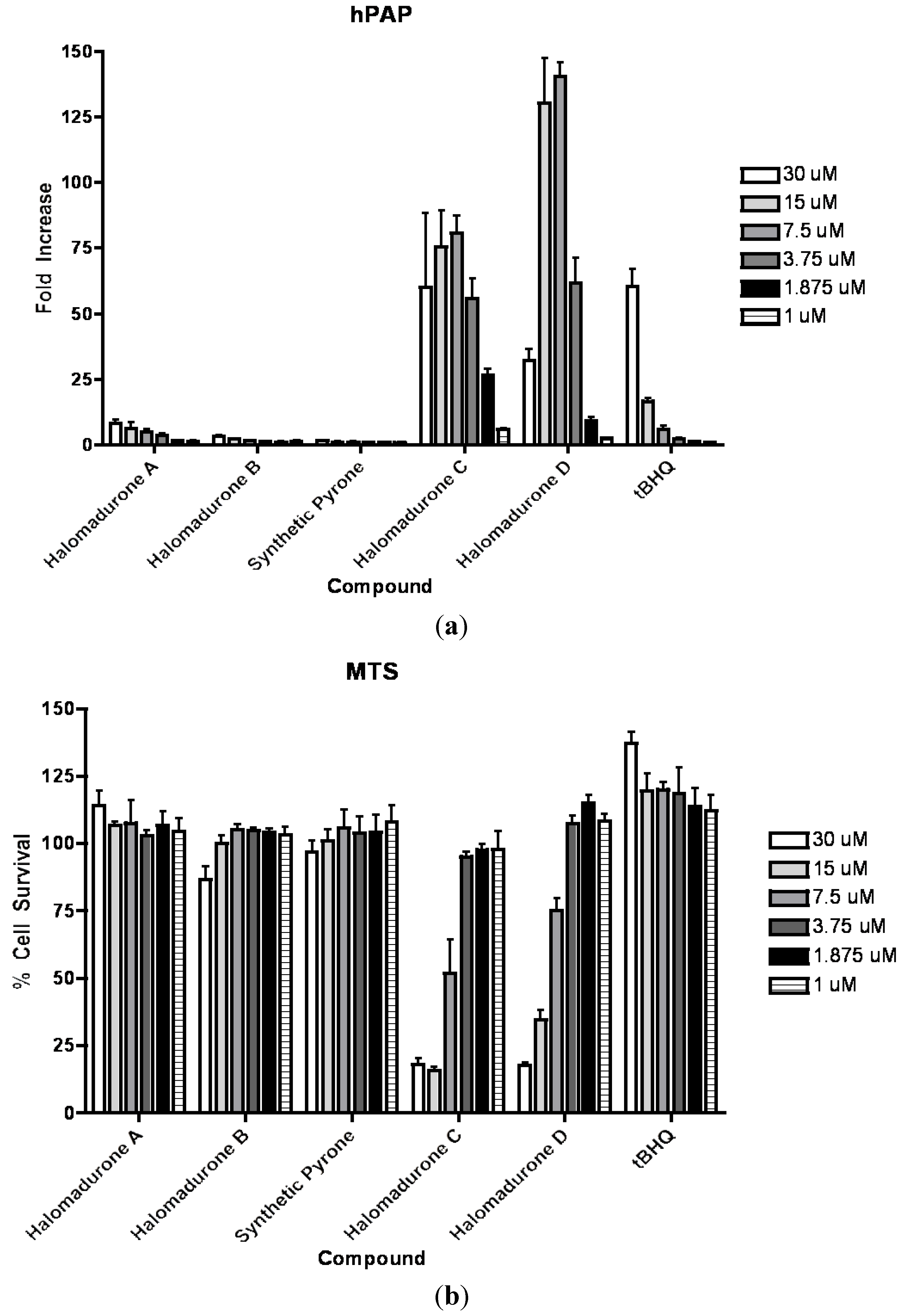
3. Experimental Section
3.1. General Experimental Procedures
3.2. Biological Material
3.3. Sequencing
3.4. Fermentation, Extraction, and Isolation
 ±0 UV (MeOH) λmax (log ε) 206 (4.02), 253 (3.40), 324 (3.50) nm; IR (ATR) νmax 1650, 1601, 1396, 1215, 934 cm−1; 1H and 13C NMR (See Table 1); HRMS [M + H]+ m/z 316.9937 (calcd. for C13H15BrClO2, 316.9938).
±0 UV (MeOH) λmax (log ε) 206 (4.02), 253 (3.40), 324 (3.50) nm; IR (ATR) νmax 1650, 1601, 1396, 1215, 934 cm−1; 1H and 13C NMR (See Table 1); HRMS [M + H]+ m/z 316.9937 (calcd. for C13H15BrClO2, 316.9938).3.5. Bioasssay hPAP and MTS
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Menken, M.; Munsat, T.L.; Toole, J.F. The global burden of disease study: Implications for neurology. Arch. Neurol. 2000, 57, 418–420. [Google Scholar] [CrossRef]
- Hebert, L.E.; Weuve, J.; Scherr, P.A.; Evans, D.A. Alzheimer disease in the United States (2010–2050) estimated during the 2010 census. Neurology 2013, 80, 1778–1783. [Google Scholar] [CrossRef]
- Alzheimer’s Disease Internationa. World Alzheimer Report 2009; Prince, M., Jackson, J., Eds.; Alzheimer’s Disease International: London, UK, 2009. [Google Scholar]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef]
- De Vries, H.E.; Witte, M.; Hondius, D.; Rozemuller, A.J.M.; Drukarch, B.; Hoozemans, J.; van Horssen, J. Nrf2-induced antioxidant protection: A promising target to conteract ROS-mediated damage neurodegenerative disease? Free Radic. Biol. Med. 2008, 45, 1375–1383. [Google Scholar] [CrossRef]
- Calkins, M.J.; Johnson, D.A.; Townsend, J.A.; Vargas, M.R.; Dowell, J.A.; Williamson, T.P.; Kraft, A.D.; Lee, J.-M.; Li, J.; Johnson, J.A. The Nrf2/ARE pathway as a potential therapeutic target in neurodegenerative disease. Antioxid. Redox Signal. 2009, 11, 497–508. [Google Scholar] [CrossRef]
- Vanugopal, R.; Jaiswal, A.K. Nrf1 and Nrf2 positively and c-Fos and Fra1 negatively regulate the human antioxidant response element-mediated expression of NAD(P)H:quinone oxidoreductase1 gene. Proc. Natl. Acad. Sci. USA 1996, 93, 14960–14965. [Google Scholar] [CrossRef]
- Itoh, K.; Ishii, T.; Wakabayashi, N.; Yamamoto, M. Regulatory mechanism of cellular response to oxidative stress. Free Radical Res. 1999, 31, 319–324. [Google Scholar] [CrossRef]
- Wild, A.C.; Moinova, H.R.; Mulcahy, R.T. Regulation of γ-glutamylcysteine synthetase subunit gene expression by the transription factor Nrf2. J. Biol. Chem. 1999, 274, 33627–33636. [Google Scholar] [CrossRef]
- Kraft, A.D.; Johnson, D.A.; Johnson, J.A. Nuclear factor E2-related factor 2-dependent antioxidant response element activation by tert-butylhydroquinone and sulforaphane occurring preferentially in astrocytes conditions neurons against oxidative insult. J. Neurosci. 2004, 24, 1101–1112. [Google Scholar] [CrossRef]
- Johnson, D.A.; Andrews, G.K.; Xu, W.; Johnson, J.A. Activation of the antioxidant response element in primary cortical neuronal cultures derived from transgenic reporter mice. J. Neurochem. 2002, 81, 1233–1241. [Google Scholar] [CrossRef]
- Balogun, E.; Hoque, M.; Gong, P.; Killeen, E.; Green, C.J.; Foresti, R.; Alam, J.; Motterlini, R. Curcumin activates the haem oxygenase-1 gene via regulation of Nrf2 and the antioxidant-responsive element. Biochem. J. 2003, 371, 887–895. [Google Scholar] [CrossRef]
- Haridas, V.; Hanausek, M.; Nishimura, G.; Soehnge, H.; Gaikwad, A.; Narog, M.; Spears, E.; Zoltaszek, R.; Walaszek, Z.; Gutterman, J.U. Triterpenoid electrophiles (avicins) activate the innate stress response by redox regulation of a gene battery. J. Clin. Invest. 2004, 113, 65–73. [Google Scholar]
- Shibuya, A.; Onda, K.; Kawahara, H.; Uchiyama, Y.; Nakayama, H.; Omi, T.; Nagaoka, M.; Matsui, H.; Hirano, T. Sofalcone, a gastric mucosa protective agent, increases vascular endothelial growth factor via the Nrf2-heme-oxygenase-1 dependent pathway in gastric epithelial cells. Biochem. Biophys. Res. Commun. 2010, 398, 581–584. [Google Scholar] [CrossRef]
- Lee, Y.M.; Jeong, G.S.; Lim, H.D.; An, R.B.; Kim, Y.C.; Kim, E.C. Isoliquiritigenin 2′-methyl ether induces growth inhibition and apoptosis in oral cancer cells via heme oxygenase-1. Toxicol. Vitro 2010, 24, 776–782. [Google Scholar] [CrossRef]
- Wang, R.; Mason, D.E.; Choe, K.P.; Lewin, A.S.; Peters, E.C.; Luesch, H. In vitro and in vivo characterization of a tunable dual-reactivity probe of the Nrf2-ARE Pathway. ACS Chem. Biol. 2013, 8, 1764–1774. [Google Scholar] [CrossRef]
- Wang, R.; Paul, V.; Luesch, H. Seaweed extracts and unsaturated fatty acid constituents from the green alga Ulva lactuca as activators of the cytoprotective Nrf2-ARE pathway. Free Radic. Biol. Med. 2013, 57, 141–153. [Google Scholar] [CrossRef]
- Orjala, J.; Gerwick, W.H. Barbamide, a chlorinated metabolite with molluscicidal activity from the Caribbean cyanobacterium Lyngbya majuscula. J. Nat. Prod. 1996, 59, 427–430. [Google Scholar] [CrossRef]
- Jiménez, J.I.; Scheuer, P.J. New lipopeptides from the Caribbean cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2001, 64, 200–203. [Google Scholar] [CrossRef]
- Nozoe, S.; Ishii, N.; Kusano, G.; Kikuchi, K.; Ohta, T. Neocarzilins A and B, novel polyenones from Streptomyces carzinostaticus. Tetrahedron Lett. 1992, 33, 7547–7550. [Google Scholar] [CrossRef]
- Nozoe, S.; Kikuchi, K.; Ishii, N.; Ohta, T. Synthesis of neocarzilin A: An absolute sterochemistry. Tetrahedron Lett. 1992, 33, 7551–7552. [Google Scholar] [CrossRef]
- Otsuka, M.; Ichinose, K.; Fujii, I.; Ebizuka, Y. Cloning, sequencing, and functional analysis of an iterative type I polyketide synthase gene cluster for biosynthesis of the antitumor chlorinated polyenone neocarzilin in “Streptomyces carzinostaticus”. Antimicrob. Agents Chemother. 2004, 48, 3468–3476. [Google Scholar] [CrossRef]
- Hou, Y.; Braun, D.R.; Michel, C.R.; Klassen, J.L.; Adnani, N.; Wyche, T.P.; Bugni, T.S. Microbial strain prioritization using metabolomics tools for the discovery of the natural products. Anal. Chem. 2012, 84, 4277–4283. [Google Scholar] [CrossRef]
- Zeng, J.; Zhan, J. A novel fungal flavin-dependent halogenase for natural product biosynthesis. ChemBioChem 2010, 11, 2119–2123. [Google Scholar] [CrossRef]
- Vaillancourt, F.H.; Vosburg, D.A.; Walsh, C.T. Dichlorination and bromination of a Threonyl-S-carrier protein by the non-heme FeII halogenase SYRB2. Chembiochem 2006, 7, 748–752. [Google Scholar] [CrossRef]
- Williams, D.H.; Fleming, I. Spectroscopic Methods in Organic Chemistry, 4th ed.; McGraw-Hill Book Company Limited: London, UK, 1989. [Google Scholar]
- Eustaquio, A.S.; Pojer, F.; Noel, J.P.; Moore, B.S. Discovery and characterization of a marine bacterial SAM-dependent chlorinase. Nat. Chem. Biol. 2008, 4, 69–74. [Google Scholar] [CrossRef]
- Wagner, C.; El Omari, M.; König, G.M. Biohalogenation: Nature’s way to synthesize halogenated metabolites. J. Nat. Prod. 2009, 72, 540–553. [Google Scholar]
- Ghandi, M.; Bayat, Y.; Teimuri-mofrad, R. A novel method for the synthesis of formyl and hydroxymethyl derivatives of 4H-pyran-4-one. Org. Prep. Proced. Int. 2002, 34, 525–530. [Google Scholar] [CrossRef]
- Satoh, T.; McKercher, S.R.; Lipton, S.A. Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs. Free Radical Biol. Med. 2013, 65, 645–657. [Google Scholar] [CrossRef]
- Wyche, T.P.; Hou, Y.; Braun, D.; Cohen, H.C.; Xiong, M.P.; Bugni, T.S. First natural analogs of the cytotoxic thiodepsipeptide thiocoraline A from a marine Verrucosispora sp. J. Org. Chem. 2011, 76, 6542–6547. [Google Scholar] [CrossRef]
- Fischedick, J.T.; Standiford, M.; Johnson, D.A.; Johnson, J.A. Structure activity relationship of phenolic diterpenes from Salvia officinalis as activators of the nuclear factor E2-related factor 2 pathway. Bioorg. Med. Chem. 2013, 21, 2618–2622. [Google Scholar] [CrossRef]
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wyche, T.P.; Standiford, M.; Hou, Y.; Braun, D.; Johnson, D.A.; Johnson, J.A.; Bugni, T.S. Activation of the Nuclear Factor E2-Related Factor 2 Pathway by Novel Natural Products Halomadurones A–D and a Synthetic Analogue. Mar. Drugs 2013, 11, 5089-5099. https://doi.org/10.3390/md11125089
Wyche TP, Standiford M, Hou Y, Braun D, Johnson DA, Johnson JA, Bugni TS. Activation of the Nuclear Factor E2-Related Factor 2 Pathway by Novel Natural Products Halomadurones A–D and a Synthetic Analogue. Marine Drugs. 2013; 11(12):5089-5099. https://doi.org/10.3390/md11125089
Chicago/Turabian StyleWyche, Thomas P., Miranda Standiford, Yanpeng Hou, Doug Braun, Delinda A. Johnson, Jeffrey A. Johnson, and Tim S. Bugni. 2013. "Activation of the Nuclear Factor E2-Related Factor 2 Pathway by Novel Natural Products Halomadurones A–D and a Synthetic Analogue" Marine Drugs 11, no. 12: 5089-5099. https://doi.org/10.3390/md11125089