2.1. Aeroplysinin-1 Inhibits the Growth of Endothelial and Tumor Cells
The effect of aeroplysinin-1 on endothelial and tumor cell growth was investigated by cell count and by MTT assay. As shown in
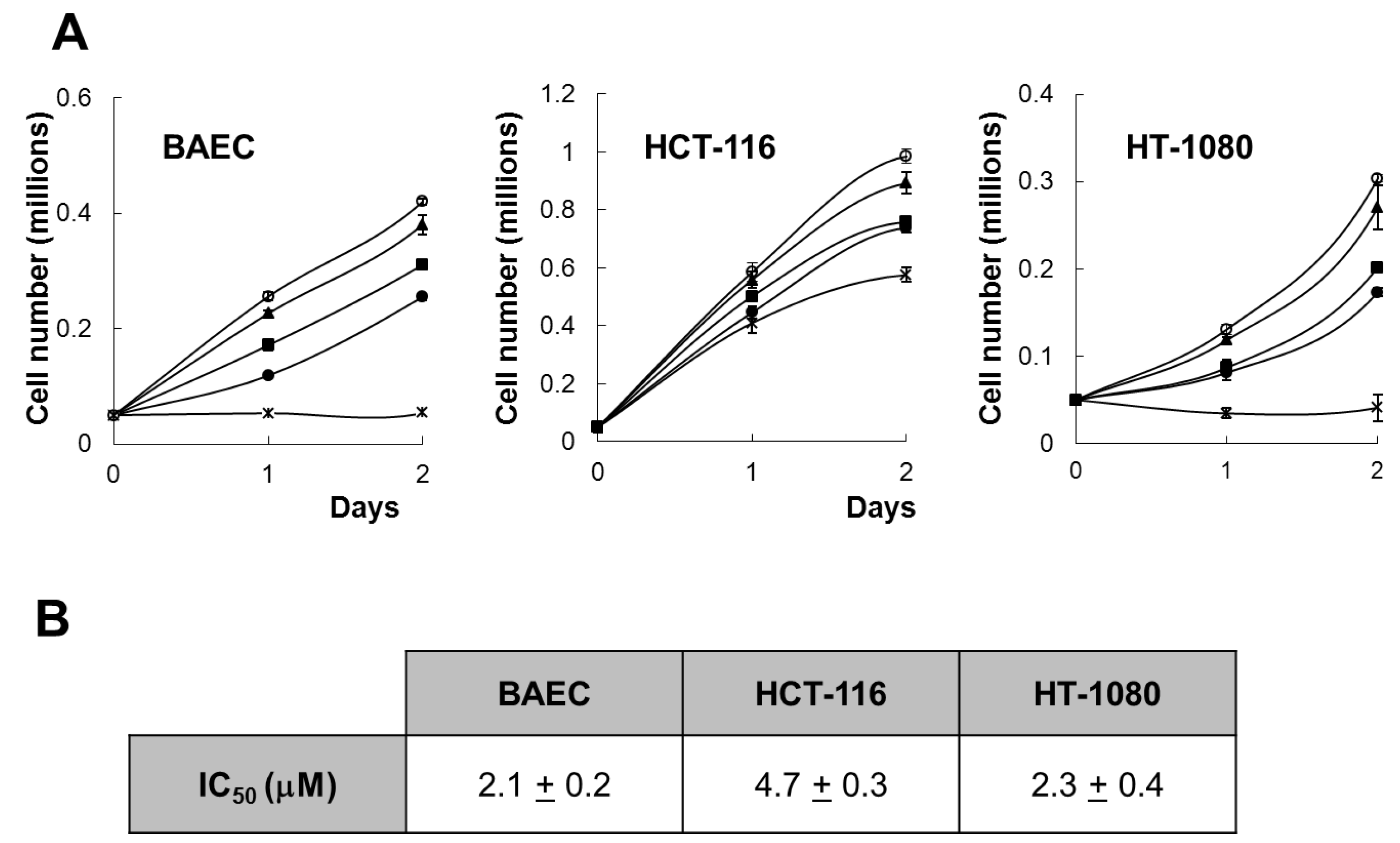
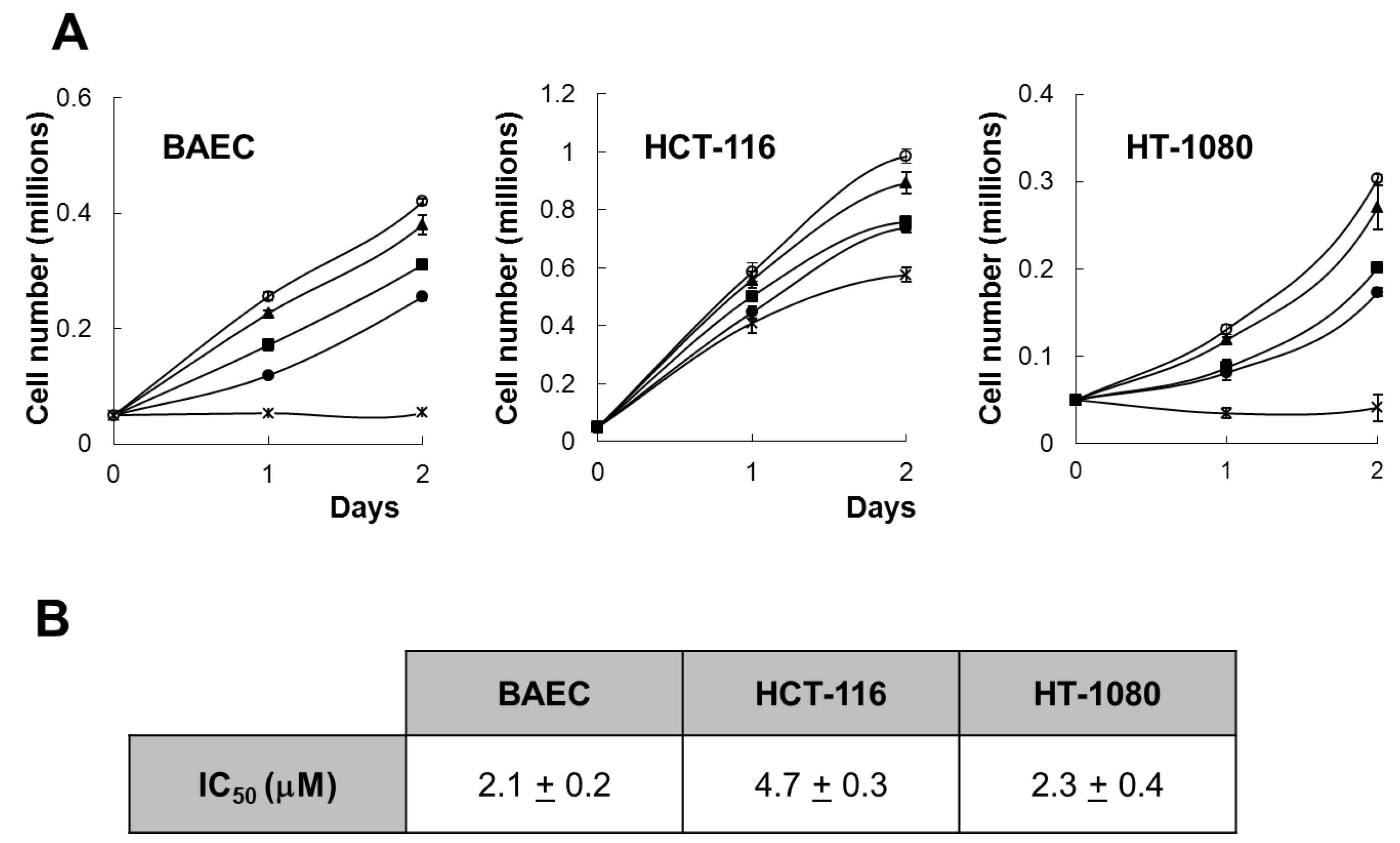
Figure 2A, aeroplysinin-1 inhibited, in a concentration-dependent manner, the growth of BAEC, HCT-116 and HT-1080, with a complete inhibition of BAEC and HCT-116 cell proliferation, exerted at a concentration of 10 µM. This is in agreement with the antiproliferative effect of aeroplysinin-1 on endothelial cells, previously described by us [
5]. Data obtained with the colorimetric MTT assay (
Figure 2B) show that aeroplysinin-1 is not a specific inhibitor of endothelial cell growth, as the half-maximal inhibitory concentration (IC50) value of this effect on endothelial cells (BAECs) is in the same range as those obtained with tumor cells (HCT-116 and HT-1080).
Figure 2.
Effect of aeroplysinin-1 on endothelial and tumor cell growth. (A) Cell counts of BAEC, HCT-116 and HT1080 grown either in the absence or the presence of 1 µM, 3 µM, 5 µM or 10 µM aeroplysinin-1 (respectively marked as o, ▲, ■, ● and x). Values are means ± SD of quadruplicate samples. (B) Half-maximal inhibitory concentration (IC50) values were calculated from MTT dose-response curves as the concentration of aeroplysinin-1 yielding 50% of control cell survival. They are expressed as means ± SD of three independent experiments with quadruplicate samples each.
Figure 2.
Effect of aeroplysinin-1 on endothelial and tumor cell growth. (A) Cell counts of BAEC, HCT-116 and HT1080 grown either in the absence or the presence of 1 µM, 3 µM, 5 µM or 10 µM aeroplysinin-1 (respectively marked as o, ▲, ■, ● and x). Values are means ± SD of quadruplicate samples. (B) Half-maximal inhibitory concentration (IC50) values were calculated from MTT dose-response curves as the concentration of aeroplysinin-1 yielding 50% of control cell survival. They are expressed as means ± SD of three independent experiments with quadruplicate samples each.
2.2. Aeroplysinin-1 Induces Apoptosis in Endothelial Cells
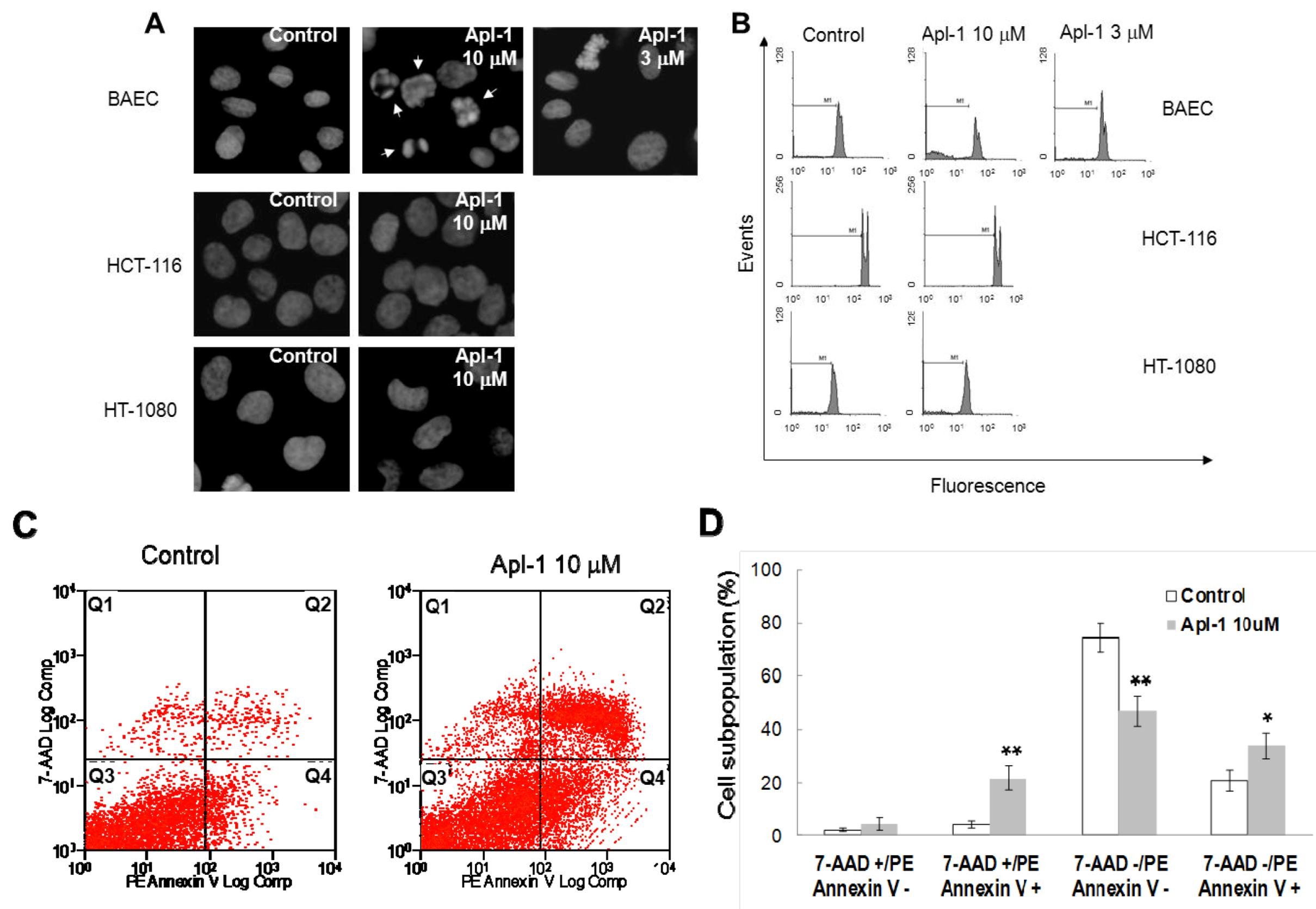
As a first approach to study the effect of aeroplysinin-1 in endothelial and tumor cells, the nuclear morphology of BAE, HT1080 and HCT116 cells was analyzed by Hoechst staining after 14 h treatment with this compound. As shown in
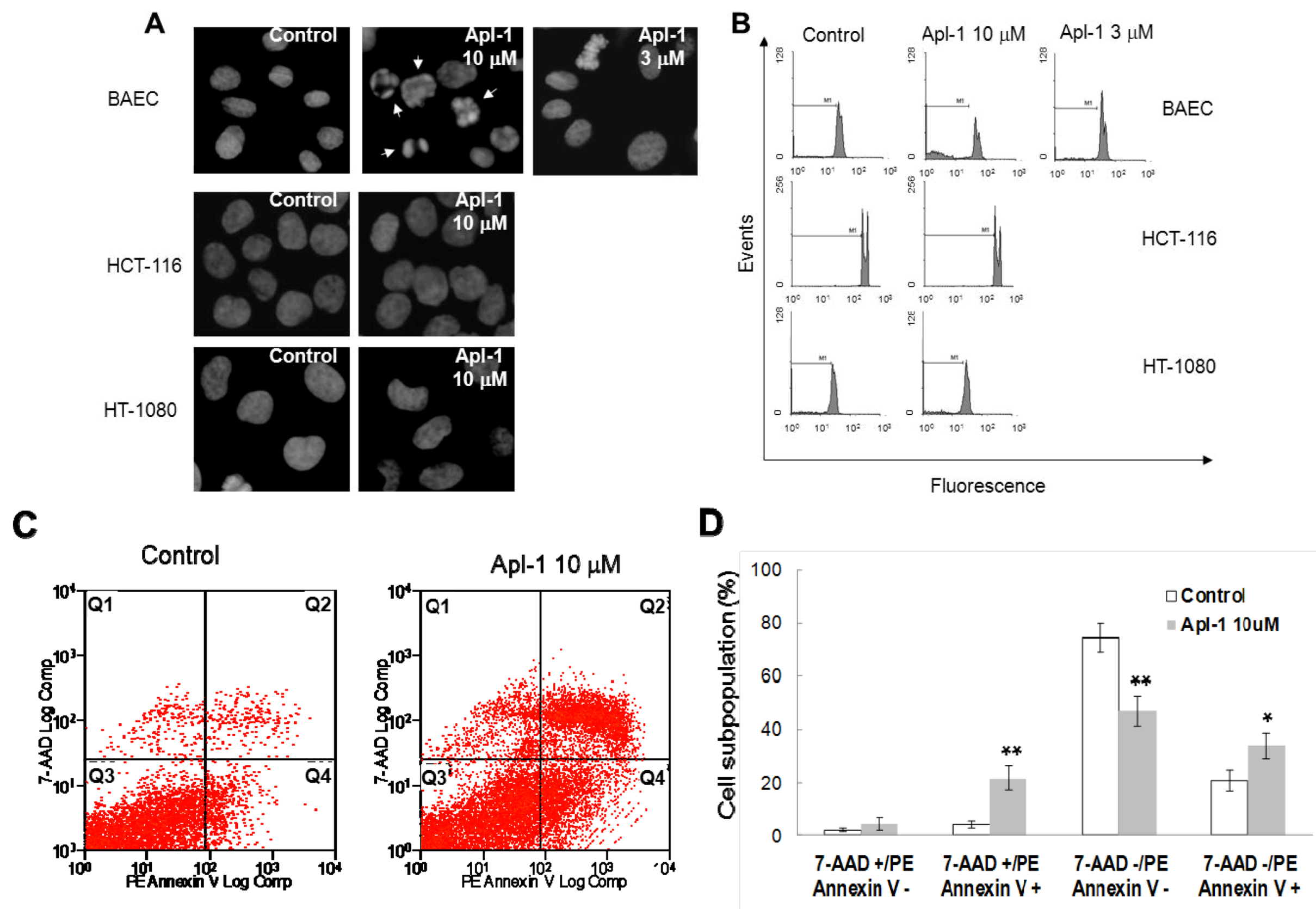
Figure 3A, treatment with 10 μM aeroplysinin-1 induced chromatin condensation and nuclear fragmentation in endothelial (BAE) cells, but not in colon carcinoma (HCT-116) or fibrosarcoma (HT-1080) cells. Nuclei of BAE cells treated with 3 μM aeroplysinin-1 did not show morphological changes compared to nuclei of non-treated cells, showing a dose dependence of this effect that is in agreement with that observed in the cell growth assay, where 10 μM aeroplysinin-1 was required to completely inhibit proliferation of BAEC. To confirm these results, the cell cycle distribution of propidium iodide-stained cells was analyzed by flow-cytometric analysis.
Figure 3B shows that a significant increase (6-fold) in the sub-G1 population was observed in BAE cells treated with 10 μM aeroplysinin-1 when compared to untreated cells. Nevertheless, no significant increases of sub-diploid population were observed in either endothelial cells treated with 3 μM aeroplysinin-1, or in HCT-116 or HT-1080 cells treated with 10 μM aeroplysinin-1 (
Figure 3B). These results suggested that aeroplysinin-1 might be a selective apoptosis trigger in endothelial cells. Endothelial cell apoptosis induced by aeroplysinin-1 was also confirmed by flow cytometry analysis after PE-Annexin V and 7-aminoactinomycin D (7AAD) staining (
Figure 3C,D), showing that 10 µM aeroplysinin-1 induced a significant increase in the percentage of cells in both early and late apoptotic cell subpopulations.
Figure 3.
Aeroplysinin-1 potentiates apoptosis in endothelial cells. (A) Nuclear morphology of endothelial and tumor cells after treatment with aeroplysinin-1, assessed under a fluorescence microscope. (B) Cell cycle distribution of endothelial and tumor cells after treatment with aeroplysinin-1, analyzed by FACS. M1 indicates the subG1 population. (C) Determination of endothelial cell (BAEC) apoptosis by flow cytometry analysis after PE-Annexin V and 7-aminoactinomycin D (7AAD) staining. (D) 7AAD−/PE-Annexin V−, 7AAD−/PE-Annexin V+, 7AAD+/PE-Annexin V+ and 7AAD+/PE-Annexin V− populations, corresponding to viable (Q3), early apoptotic (Q4), late apoptotic (Q2) and necrotic (Q1) cells respectively, were evaluated as in (C). Values are expressed as means ± SD of three independent experiments. * p < 0.05; ** p < 0.005 versus control.
Figure 3.
Aeroplysinin-1 potentiates apoptosis in endothelial cells. (A) Nuclear morphology of endothelial and tumor cells after treatment with aeroplysinin-1, assessed under a fluorescence microscope. (B) Cell cycle distribution of endothelial and tumor cells after treatment with aeroplysinin-1, analyzed by FACS. M1 indicates the subG1 population. (C) Determination of endothelial cell (BAEC) apoptosis by flow cytometry analysis after PE-Annexin V and 7-aminoactinomycin D (7AAD) staining. (D) 7AAD−/PE-Annexin V−, 7AAD−/PE-Annexin V+, 7AAD+/PE-Annexin V+ and 7AAD+/PE-Annexin V− populations, corresponding to viable (Q3), early apoptotic (Q4), late apoptotic (Q2) and necrotic (Q1) cells respectively, were evaluated as in (C). Values are expressed as means ± SD of three independent experiments. * p < 0.05; ** p < 0.005 versus control.
2.3. Aeroplysinin-1 Induces Activation of Caspases and Cleavage of PARP and Lamin-A in Endothelial Cells
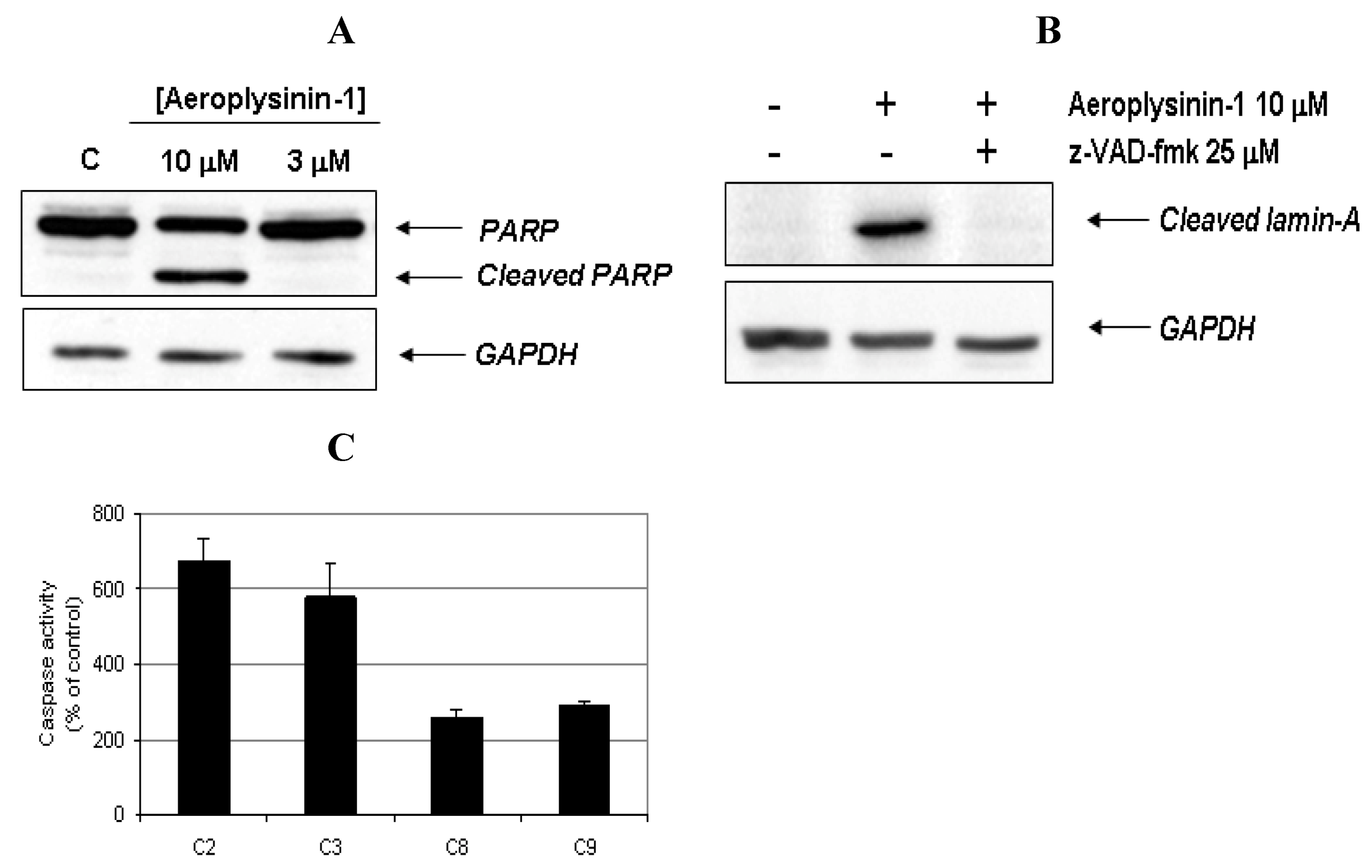
In order to provide biochemical evidence for the induction of apoptosis in BAE cells treated with aeroplysinin-1, the cleavage of poly (ADP-ribose) polymerase (PARP) and lamin-A, was studied. PARP cleavage by activated caspase-3 is a key event in the process of apoptosis and is used as an early marker for apoptosis induction [
12]. As shown in
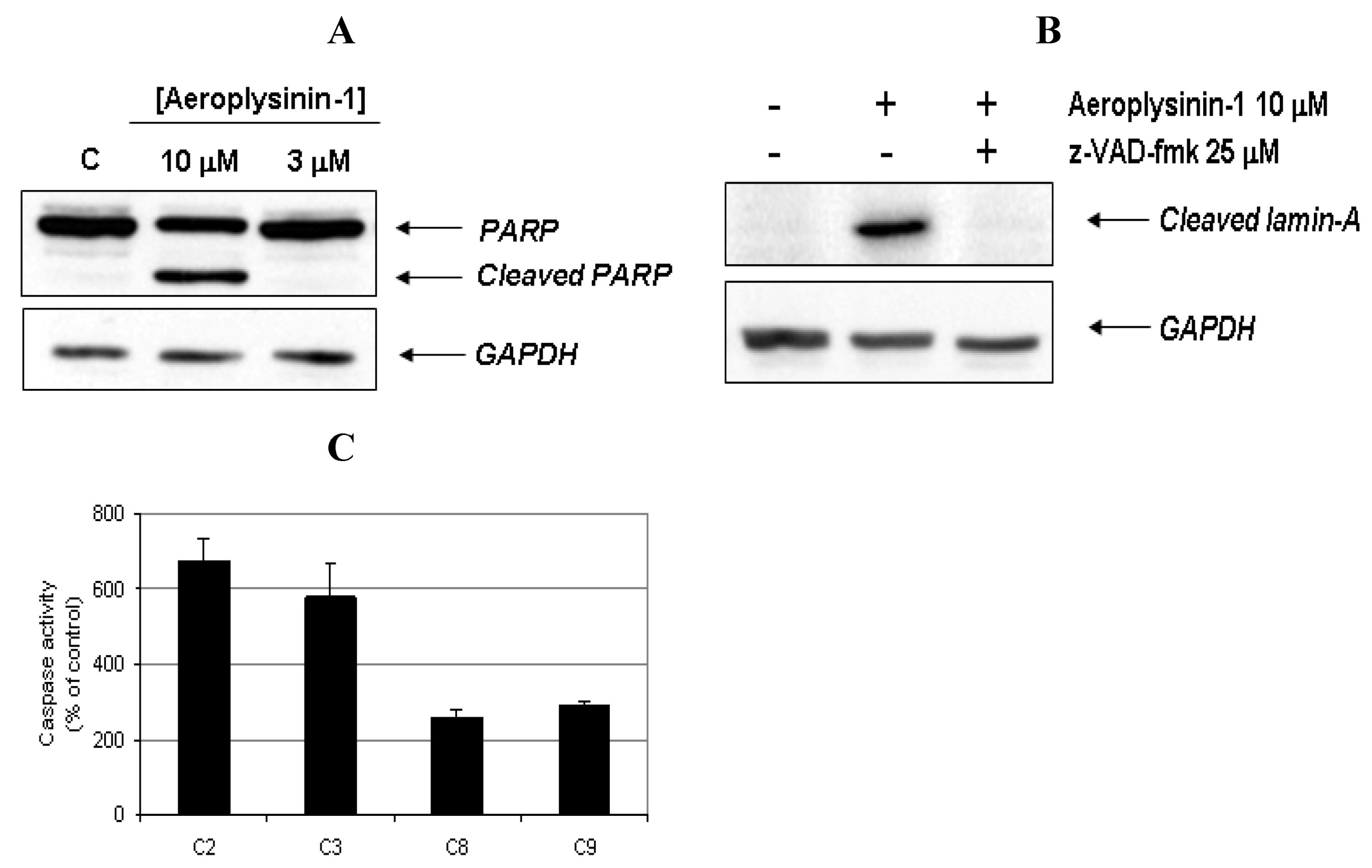
Figure 4A, PARP was cleaved from 116 kDa intact form into 85 kDa fragment after BAE cell treatment with 10 μM of aeroplysinin-1. Lamin-A is a structural protein belonging to the intermediate filament family that, together with other lamin species, constitutes the scaffolding of the nuclear envelope. During apoptosis, lamin A is cleaved at Asp230 by caspase-6 [
13] causing the characteristic collapse of the nucleus observed during apoptosis. Treatment of BAEC with 10 µM aeroplysinin-1 caused lamin A cleavage, generating a fragment of 28 kDa. Based on those results, we reasoned whether aeroplysinin-1 induced apoptosis could be mediated by caspases, and used a pharmacological inhibitor of caspases to gain insight into this question. As shown in
Figure 4B, the addition of 25 µM of the pan-caspase inhibitor
N-benzyloxycabonyl-Val-Ala-Asp-fluoromethylketone (ZVAD) inhibited lamin A cleavage, indicating that aeroplysinin-1 induces apoptosis in BAE cells through a caspase-dependent pathway.
To further confirm that aeroplysinin-1 induced cell death is a caspase-dependent process, the activities of different caspases were measured. Fluorometric assays using specific substrates showed that 14 h treatment of BAE cells with 10 µM aeroplysinin-1 induced a 7 fold increase of caspase-2 activity, a 6 fold increase of caspase-3 and a 3 fold increase of caspases-8 and -9, as compared to the activity of untreated cells (
Figure 4C). Inhibition of caspase-9 after the addition of the specific caspase-9 inhibitor LEHD-fmk significantly abrogated aeroplysinin-1-induced activation of caspase-2 and caspase-3 (results not shown), indicating the relevant role of caspase-9 as an upstream caspase in the mitochondria-mediated apoptosis pathway.
Figure 4.
Activation of caspases and cleavage of apoptotic substrates induced by aeroplysinin-1. (A) Western-blot detection of PARP cleavage in aeroplysinin-1-treated BAE cells. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as loading control. (B) Western-blot detection of cleaved lamin-A in aeroplysinin-1-treated BAE cells with or without pre-incubation of cells with the pan-caspase inhibitor z-VAD 25 μM. (C) Effect of 10 µM aeroplysinin-1 in BAE cells caspase-2, -3, -8 and -9 activation. Activity values from treated cells are expressed as percentage of untreated (control) cells. Bars represent the standard deviation from duplicated samples in the same assay. Similar results were obtained in three independent experiments.
Figure 4.
Activation of caspases and cleavage of apoptotic substrates induced by aeroplysinin-1. (A) Western-blot detection of PARP cleavage in aeroplysinin-1-treated BAE cells. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as loading control. (B) Western-blot detection of cleaved lamin-A in aeroplysinin-1-treated BAE cells with or without pre-incubation of cells with the pan-caspase inhibitor z-VAD 25 μM. (C) Effect of 10 µM aeroplysinin-1 in BAE cells caspase-2, -3, -8 and -9 activation. Activity values from treated cells are expressed as percentage of untreated (control) cells. Bars represent the standard deviation from duplicated samples in the same assay. Similar results were obtained in three independent experiments.
2.4. Aeroplysinin-1 Induces Cytochrome c Release in Endothelial Cells
Our data, suggesting a relevant role of caspase-9 activation in the aeroplysinin-1 apoptogenic activity, prompted us to investigate alterations in the physiology of the mitochondria caused by this drug. The intrinsic or mitochondrial apoptotic pathway is characterized by a permeabilization of the outer mitochondrial membrane induced by the pro-apoptotic stimuli, leading to the release of cytochrome
c from the intermembrane space into the cytosol. Cytochrome
c is a crucial component of the mitochondrial pathway of apoptosis, since by binding to the cytosolic apoptotic protease-activating factor 1 (Apaf-1), it contributes to the apoptosome assembly, which acts as an activation platform for procaspase-9 [
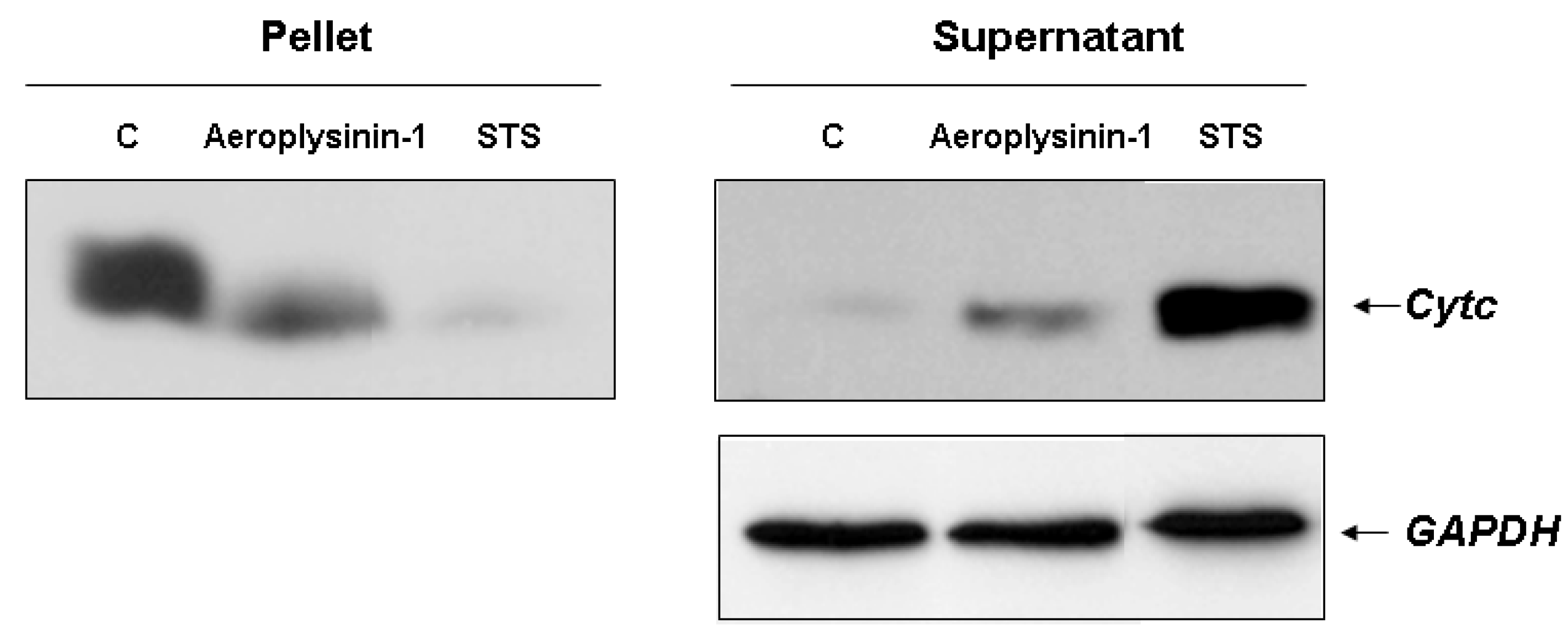
14]. This feature was studied in digitonin-permeabilized BAE cells treated with 10 µM aeroplysinin-1 as described in
Experimental Section. Treatment with aeroplysinin-1 elicited the redistribution of cytochrome
c from mitochondria to cytosol, although the release seemed to be incomplete, as shown by the presence of cytochrome
c in the pellet (
Figure 5).
Figure 5.
Detection of cytochrome c release in 10 µM aeroplysinin-1-treated BAE cells. Treatment of BAEC with staurosporine (STS) 2 µM was used as internal control of mitochondrial cytochrome c release. Representative blots from two different experiments are shown.
Figure 5.
Detection of cytochrome c release in 10 µM aeroplysinin-1-treated BAE cells. Treatment of BAEC with staurosporine (STS) 2 µM was used as internal control of mitochondrial cytochrome c release. Representative blots from two different experiments are shown.
2.5. Aeroplysinin-1 Inhibits BAD Phosphorylation in Endothelial Cells
The BH3-only pro-apoptotic protein Bad is regulated by phosphorylation-dephosphorylation in response to extracellular stimuli. The phosphorylation status of Bad determines its pro-apoptotic activity at the mitochondrial level [
15], facilitating its depolarization by interacting and restraining the anti-apoptotic protein Bcl-XL. Therefore, we decided to study the effect of aeroplysinin-1 on the phosphorylation status of Bad in endothelial cells by serum deprivation and re-stimulation, with or without addition of aeroplysinin-1. As shown in
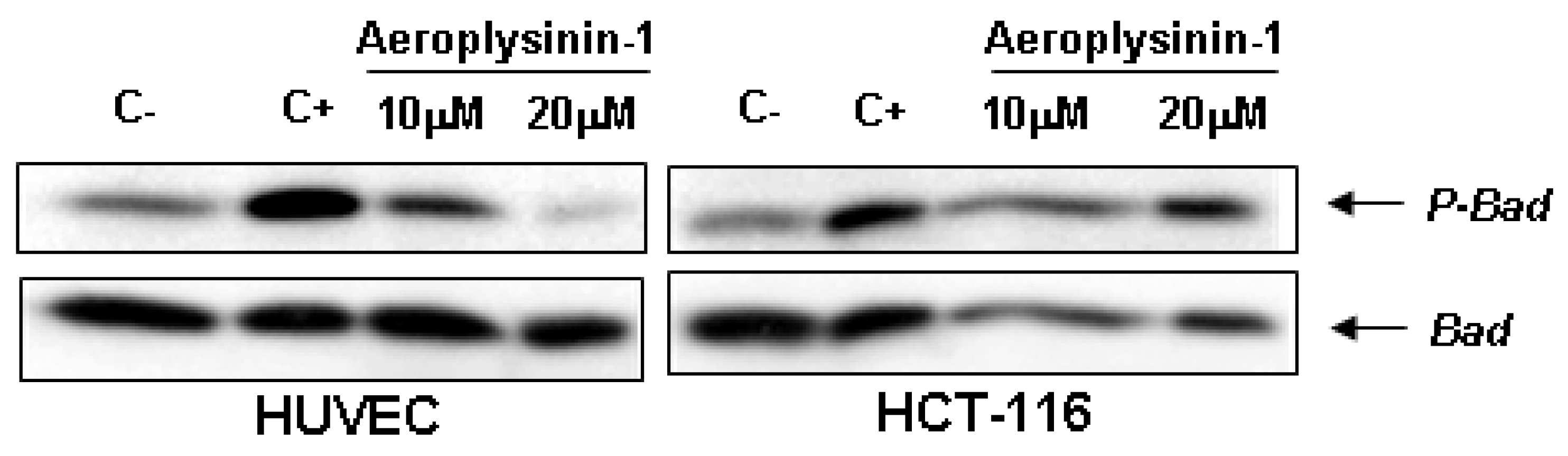
Figure 6, aeroplysinin-1 prevented phosphorylation of Bad in HUVE cells in a dose-dependent manner, but not in HCT-116 cells, reinforcing the previously shown data regarding the endothelial selectivity of aeroplysinin-1 apoptogenic activity (
Figure 3 and
Figure 6).
Figure 6.
Effects of aeroplysinin-1 on Bad phosphorylation in human umbilical vein endothelial cells (HUVEC) and human HCT-116 colon carcinoma cells. Negative control (C−) corresponds to starved but not induced cells; positive control (C+) corresponds to starved and serum-induced cells. Identical results were obtained in two independent experiments.
Figure 6.
Effects of aeroplysinin-1 on Bad phosphorylation in human umbilical vein endothelial cells (HUVEC) and human HCT-116 colon carcinoma cells. Negative control (C−) corresponds to starved but not induced cells; positive control (C+) corresponds to starved and serum-induced cells. Identical results were obtained in two independent experiments.
2.6. Global Discussion
Aeroplysinin-1 has been previously described by us as a potent antiangiogenic compound, showing inhibitory effects
in vitro and
in vivo. In addition, it was suggested to trigger apoptosis in endothelial cells, although no results characterizing this process have been provided to date [
5]. The present work provides evidence showing that aeroplysinin-1 effectively induces apoptosis in endothelial cells via the apoptogenic mitochondrial pathway, with inhibition of Bad phosphorylation, cytochrome
c release from mitochondria and activation of caspases.
Our previous results indicated that aeroplysinin-1 inhibits the growth of highly proliferant BAE cells, and it exhibits a significantly lower activity on low-proliferant cells [
5]. Results shown in this paper confirm the antiproliferative activity of this compound on endothelial and tumor cells. Moreover, our previous suggestion that aeroplysinin-1 could induce cell death by activation of apoptosis is now confirmed by new experimental data, revealing that incubation with this compound induces several changes on endothelial cells which are indicative of apoptosis, including chromatin condensation and nuclear fragmentation, an increase in the percentage of cells with sub-diploid DNA content, and an increase in the Annexin V+ subpopulations. Interestingly, the same doses of aeroplysinin-1 did not cause any observable effect in the human HCT-116 and HT-1080 tumor cell lines, which suggests a selective induction of apoptosis in endothelial cells.
Activation of caspases is considered to be the prerequisite to define apoptotic cell death [
16]. Here, we demonstrate that aeroplysinin-1 induces activation of caspase-2, -3, -8 and -9 in endothelial cells, in parallel with the cleavage of hallmark apoptotic substrates, such as PARP and lamin-A. Selective inhibition of caspase-9 by LEHD-fmk abrogated activation of caspases-2 and -3 by aeroplysinin-1, suggesting it to be dependent on caspase-9 activation upstream of caspase-2 and -3. Caspase-2 is a putative initiator caspase characterized by the presence of a caspase activation recruitment domain (CARD) and structurally related to the initiator caspase-9. The precise mechanism of caspase-2 activation during apoptosis is the subject of intense research [
17] and seems to be dependent on the nature of the stimulus. The activation of caspase-2 in response to genotoxic stress has been shown to occur within a PIDDosome complex [
18] and this process is dependent on p53 function [
19]. The role of caspase-2 remains controversial as to whether it is either an essential initiator caspase acting upstream of mitochondria [
20], or an executioner caspase activated downstream of mitochondria [
21]. Our results suggest that aeroplysinin-1-induced caspase-2 activation in endothelial cells depends on caspase-9 activity and hence, it occurs downstream of mitochondria.
The linked observations of the presence of caspase-9 activity and the induction of cytochrome
c release in aeroplysinin-1-treated endothelial cells suggest an implication of mitochondria in this model. Mitochondria plays a central role in the transmission of the apoptotic cascade [
14]. Depolarization of the outer mitochondrial membrane causes the release of different pro-apoptotic proteins from the mitochondrial intermembrane space to the cytosol, such as cytochrome
c, which interacts with the adaptor protein Apaf-1 and constitutes the platform for the activation of caspase-9 and downstream executioner caspases [
22].
The Bcl-2-related BH3-only pro-apoptotic protein Bad is an upstream sensor of cellular damage that is activated through phosphorylation and dephosphorylation processes in response to extracellular stimuli [
23]. Dephosphorylated Bad heterodimerises with Bcl-XL in the outer mitochondrial membrane, leading to changes in mitochondria permeability and promoting apoptosis [
15]. Phosphorylation of Bad in serine residues results in its association with the scaffold protein 14-3-3, which sequester Bad in the cytosol, preventing its pro-apoptotic effect. Although Bad can be phosphorylated in some serine residues, Ser112 and Ser136 are the residues implicated in the association of this protein with 14-3-3 [
24]. In fact, the phosphorylation state of Bad has been proposed to decrease the vulnerability of the mitochondria to release cytochrome
c and induce apoptosis in response to death signals [
25].
Little is known about aeroplysinin-1 and how it can selectively target endothelial cells. Several reports provide evidence concerning the inhibitory effect of aeroplysinin-1 on EGF receptor
in vitro and
in vivo [
26,
27], although this effect has been questioned by others [
28]. No data is available about the inhibitory capacity of aeroplysinin-1 on key steps of the signal transduction pathways controlling angiogenesis. Taken together, our results led us to suggest a mechanism by which aeroplysinin-1, through inhibition of Bad phosphorylation, could orchestrate the decision to undergo endothelial apoptosis by mitochondria permeabilization, cytochrome
c release and further activation of the caspases proteolytic cascade (
Figure 7). Undoubtedly, more experimental efforts will be required to elucidate this issue.
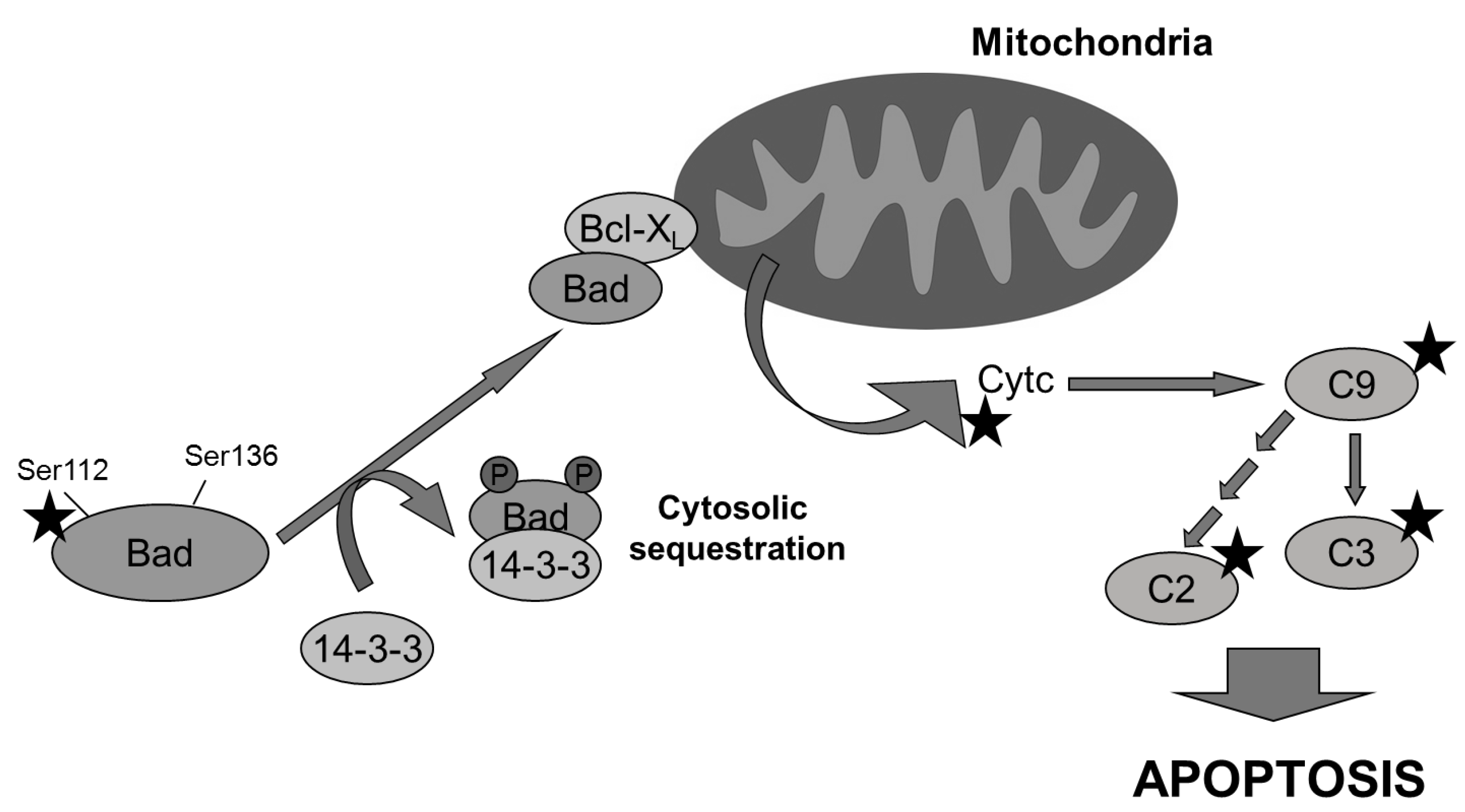
Figure 7.
Multiple molecular targets (indicated with a star) are involved in the pro-apoptotic effect of aeroplysinin-1 observed in endothelial cells. Bad is dephosphorylated and it translocates from cytosol to mitochondria, where it dimerizes with Bcl-XL and promotes the release of cytochrome c, activating caspase-9 (C9) and other downstream caspases.
Figure 7.
Multiple molecular targets (indicated with a star) are involved in the pro-apoptotic effect of aeroplysinin-1 observed in endothelial cells. Bad is dephosphorylated and it translocates from cytosol to mitochondria, where it dimerizes with Bcl-XL and promotes the release of cytochrome c, activating caspase-9 (C9) and other downstream caspases.

 and
and 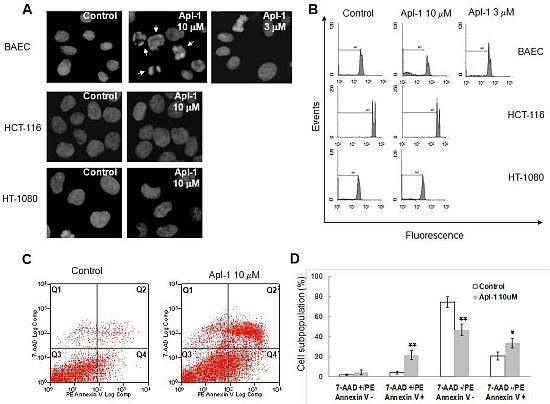
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}