Mitochondrial Genome-Knockout Cells Demonstrate a Dual Mechanism of Action for the Electron Transport Complex I Inhibitor Mycothiazole

 ,
,
Abstract
:
1. Introduction
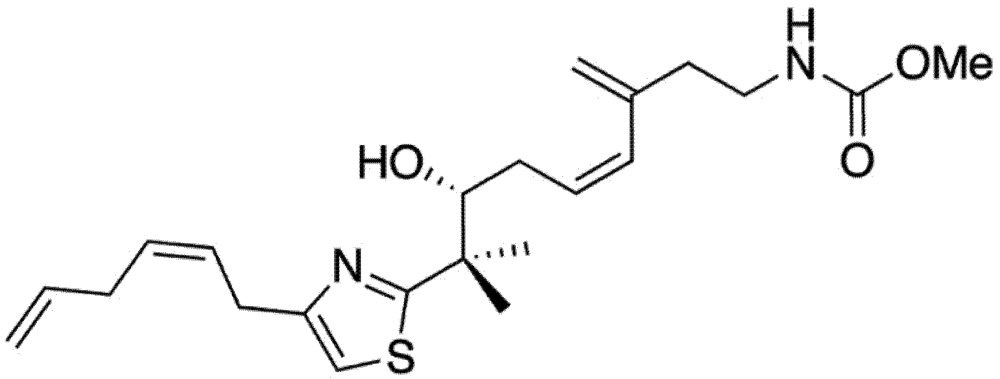
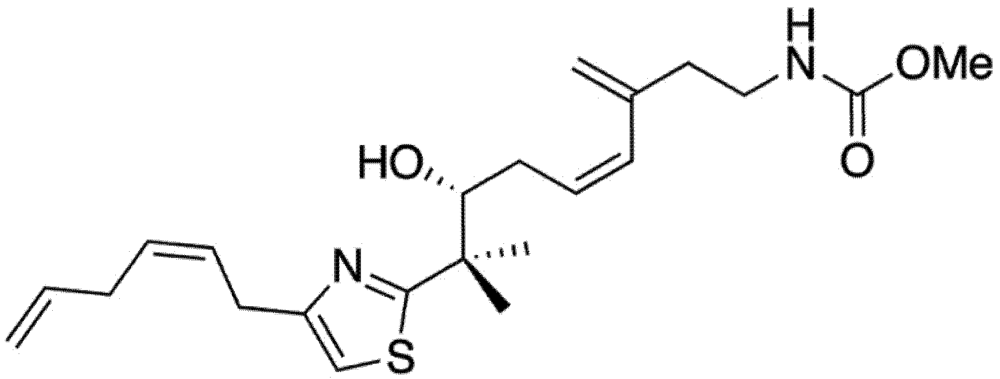
2. Results and Discussion
2.1. Mycothiazole Isolation and Purification
2.2. Effects of MYZ on Cell Metabolism and the Proliferative Response

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
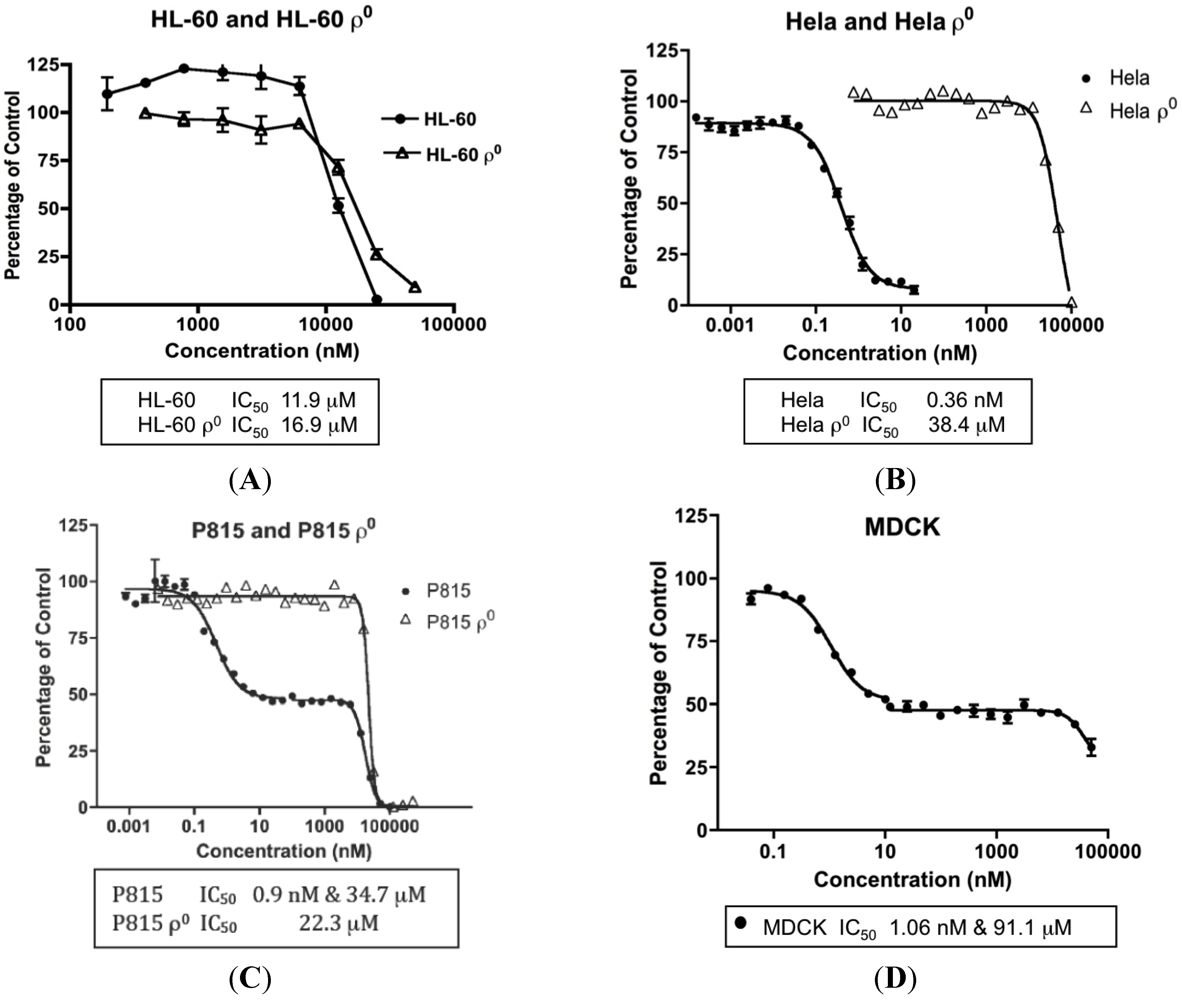
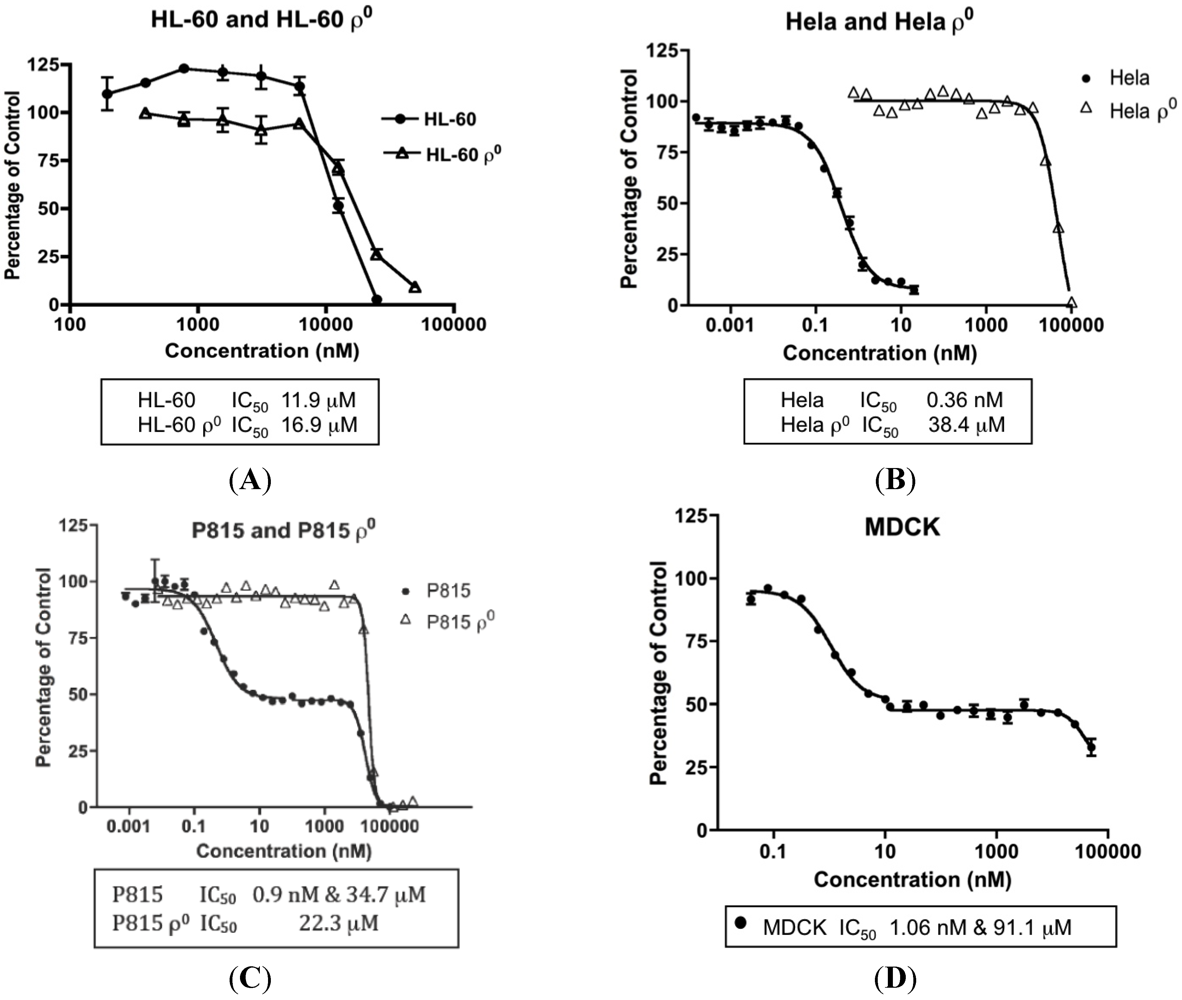
| Parental Cell Lines | IC50 (Mean ± SEM) | n | ρ0 Cell Lines | IC50 (Mean ± SEM) | n |
|---|---|---|---|---|---|
| Sensitive | |||||
| HeLa | 0.36 ± 0.09 nM | 4 | HeLaρ0 | 78.4 ± 20.2 μM | 3 |
| HeLa S3 | 1.84 nM | 2 | |||
| 143B | 3.24 ± 1.6 nM | 4 | 143Bρ0 | 50.7 μM | 1 |
| B16 | 13.8 ± 3.6 nM | 3 | B16ρ0 | 34.7 μM | 1 |
| 4T1 | 12.1 ± 2.5 nM | 3 | |||
| RAW 264/7 | 1.54 nM | 2 | |||
| PrimaryT Cells | CD4 2.04 ± 0.37 nMCD8 7.20 ± 0.52 nM | 55 | |||
| Biphasic | |||||
| P815 | (a) 1.02 ± 0.22 nM (b) 30.3 ± 5.1 μM | 45 | P815ρ0 | 36.4 ± 9.2 μM | 3 |
| MDCK | (a) 1.47 ± 0.79 nM (b) 91.1 μM | 31 | |||
| Insensitive | |||||
| HL-60 | 12.2 ± 2.2 μM | 5 | HL-60ρ0 | 26.1 ± 9.2 μM | 2 |
| LN18 | 26.5 ± 3.1 μM | 3 | |||
| Jurkat | 26.5 μM |

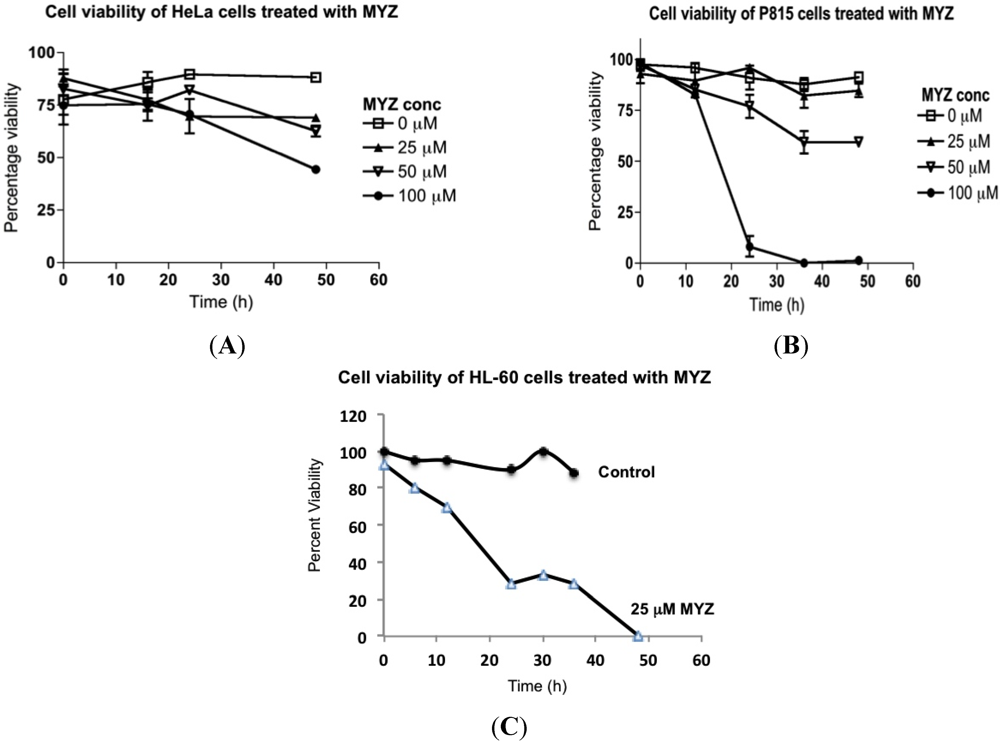
2.3. Effects of MYZ on Cell Viability

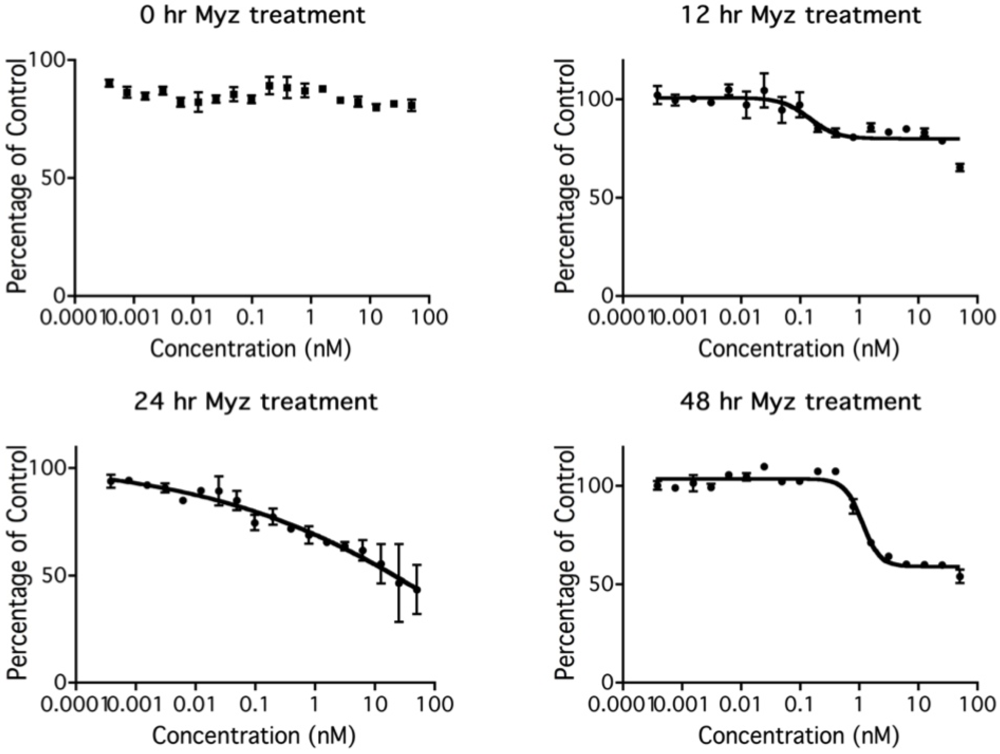
2.4. Lag before MYZ Exerts Its Effects on Cell Metabolism
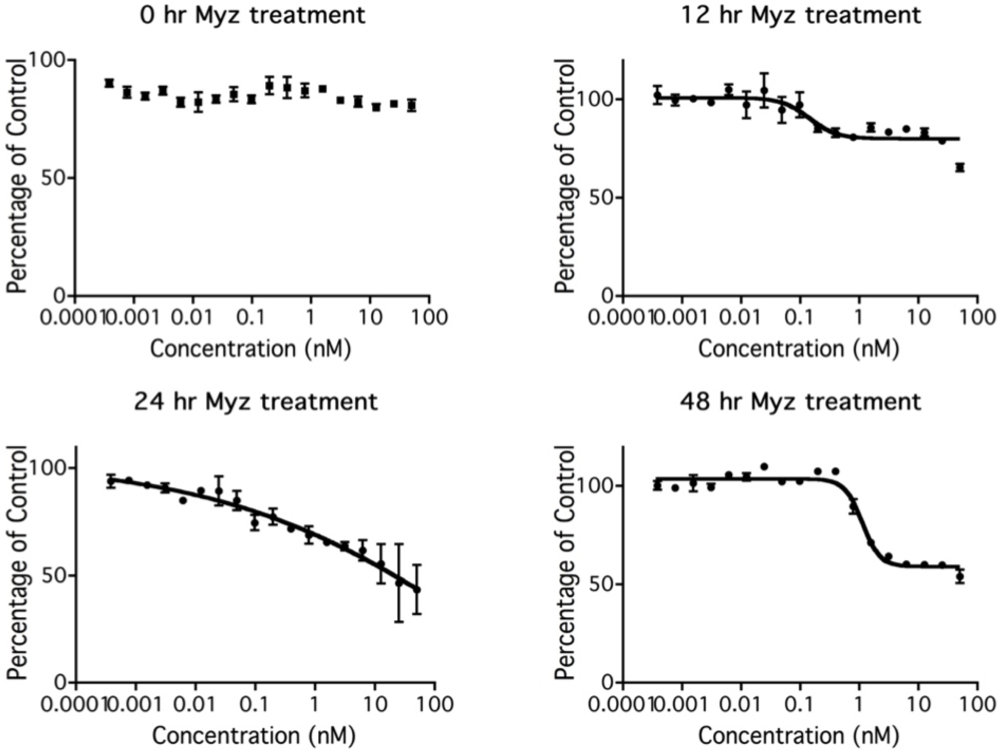
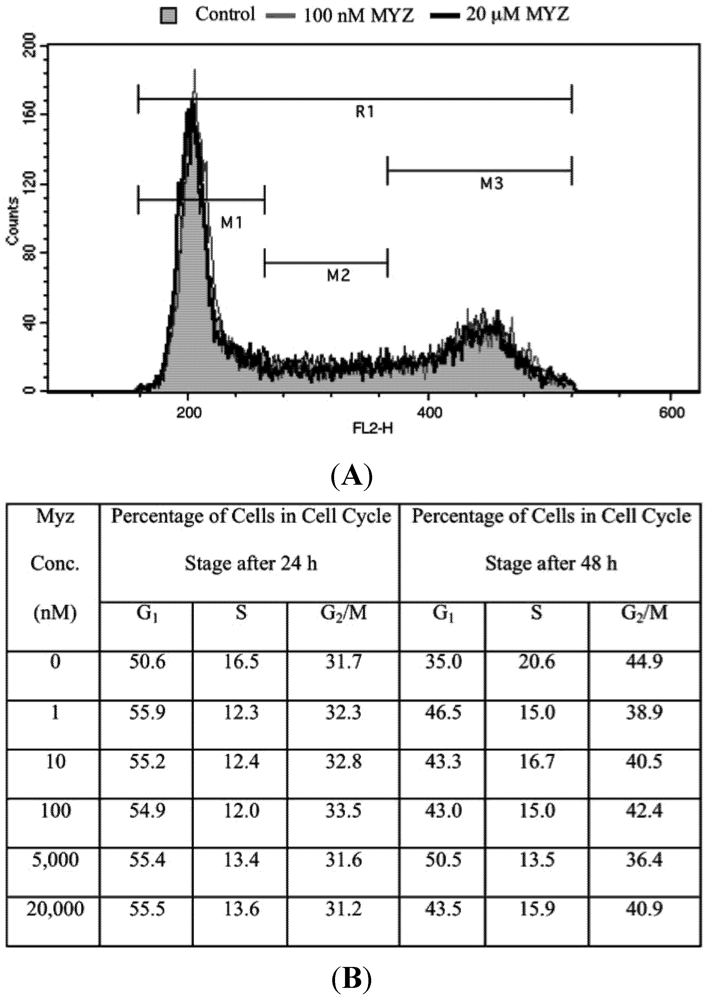
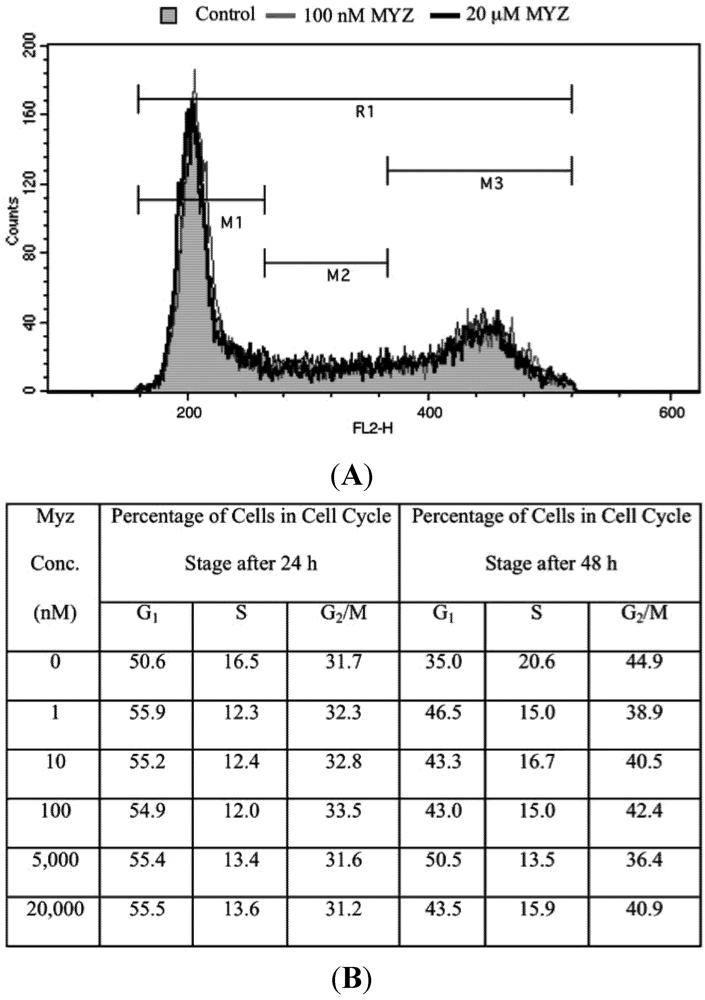
2.5. Effects of MYZ on the Cell Cycle
| Concentration MYZ (nM) | G1 (%) | S (%) | G2/M (%) | n |
|---|---|---|---|---|
| 0 | 70.4 ± 2.2 | 15.5 ± 0.6 | 13.6 ± 2.4 | 7 |
| 0.1 | 68.8 ± 3.8 | 17.4 ± 2.1 | 14.2 ± 2.8 | 4 |
| 1 | 70.4 ± 3.6 | 15.9 ± 1.4 | 14.0 ± 2.6 | 4 |
| 10 | 69.6 ± 2.1 | 15.5 ± 1.2 | 15.2 ± 1.1 | 5 |
| 100 | 69.9 ± 2.9 | 15.4 ± 1.8 | 15.0 ± 1.3 | 5 |
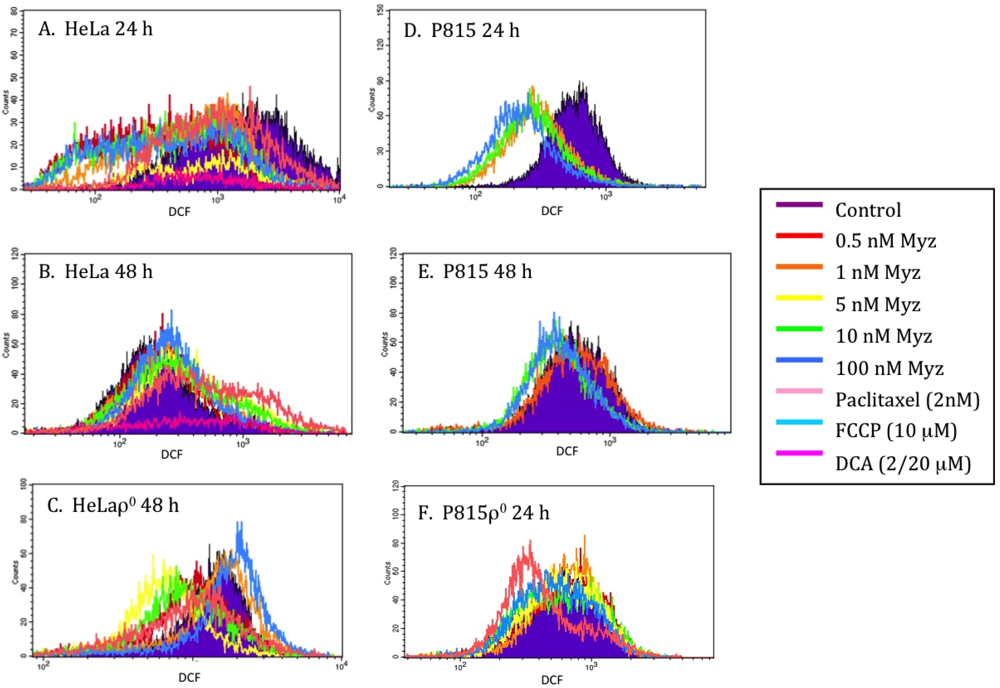
2.6. Effects of MYZ on ROS

2.7. MYZ Activity Range in Different Cell Lines and Dependence on Mitochondrial Function
2.8. MYZ Action on Metabolism, Cell Cycle, and Cell Viability
2.9. Possible Mechanisms of MYZ Action
2.9.1. Lack of Correlation to NQO1 Reductase
2.9.2. Differences between MYZ and Rotenone
2.9.3. Dual Target Mechanism and Future Directions
3. Experimental Section
3.1. MYZ Extraction and Purification
3.2. Cell Culture
3.3. Cell Metabolism Assay
3.4. 3H-Thymidine Uptake
3.5. CFSE Cell Proliferation Assay
3.6. Trypan Blue Assay for Cell Viability
3.7. Cell Cycle Analysis
3.8. Measurement of Cellular ROS
4. Conclusions
Acknowledgments
Supplementary Files
References
- Crews, P.; Kakou, Y.; Quinoa, E. Mycothiazole, a polyketide heterocycle from a marine sponge. J. Am. Chem. Soc. 1988, 110, 4365–4368. [Google Scholar]
- Mahler, G.; Serra, G.; Dematteis, S.; Saldana, J.; Dominguez, L.; Manta, E. Synthesis and biological evaluation of simplified mycothiazole analogues. Bioorg. Med. Chem. Lett. 2006, 16, 1309–1311. [Google Scholar]
- Sonnenschein, R.N.; Johnson, T.A.; Tenney, K.; Valeriote, F.A.; Crews, P. A reassignment of (−)-mycothiazole and the isolation of a related diol. J. Nat. Prod. 2006, 69, 145–147. [Google Scholar]
- Developmental Therapeutics Program (DTP) web site. Available online: http://dtp.nci.nih.gov (accessed on 10 November 2011). NIH NSC number 647640.
- Morgan, J.B.; Liu, Y.; Zhou, Y.D.; Nagle, D.G. Mycothiazole Potently Inhibits Hypoxia-inducible Factor-1 (HIF-1) Activation in Breast and Prostate Tumor Cells. In Proceedings of the 48th Annual Meeting of the American Society Pharmacognosy, Portland, ME, USA, 14–18 July 2007. Abstract P007M.
- Morgan, J.B.; Mahdi, F.; Liu, Y.; Coothankandaswamy, V.; Jekabsons, M.B.; Johnson, T.A.; Sashidhara, K.V.; Crews, P.; Nagle, D.G.; Zhou, Y.D. The marine sponge metabolite mycothiazole: A novel prototype mitochondrial complex I inhibitor. Bioorg. Med. Chem. 2010, 18, 5988–5994. [Google Scholar]
- Scatena, R.; Bottoni, P.; Botta, G.; Martorana, G.E.; Giardina, B. The role of mitochondria in pharmacotoxicology: A reevaluation of an old, newly emerging topic. Am. J. Physiol. Cell Physiol. 2007, 293, C12–C21. [Google Scholar]
- Petrosillo, G.; di Venosa, N.; Ruggiero, F.M.; Pistolese, M.; D’Agostino, D.; Tiravanti, E.; Fiore, T.; Paradies, G. Mitochondrial dysfunction associated with cardiac ischemia/reperfusion can be attenuated by oxygen tension control. Role of oxygen-free radicals and cardiolipin. Biochim. Biophys. Acta 2005, 1710, 78–86. [Google Scholar] [CrossRef]
- Ralph, S.J.; Moreno-Sánchez, R.; Neuzil, J.; Rodríguez-Enríquez, S. Inhibitors of succinate: Quinone reductase/Complex II regulate production of mitochondrial reactive oxygen species and protect normal cells from ischemic damage but induce specific cancer cell death. Pharm. Res. 2011, 28, 2695–2730. [Google Scholar]
- Glover, C.J.; Rabow, A.A.; Isgor, Y.G.; Shoemaker, R.H.; Covell, D.G. Data mining of NCI’s anticancer screening database reveals mitochondrial Complex I inhibitors cytotoxic to leukemia cell lines. Biochem. Pharmacol. 2007, 73, 331–340. [Google Scholar]
- McLaughlin, J.L. Paw paw and cancer: Annonaceous acetogenins from discovery to commercial products. J. Nat. Prod. 2008, 71, 1311–1321. [Google Scholar]
- Ly, J.D.; Lawen, A. Transplasma membrane electron transport: Enzymes involved and biological function. Redox Rep. 2003, 8, 3–21. [Google Scholar]
- Herst, P.; Tan, A.; Scarlett, D.; Berridge, M. Cell surface oxygen consumption by mitochondrial gene knockout cells. Biochim. Biophys. Acta 2004, 1656, 79–87. [Google Scholar]
- King, M.; Attardi, G. Human cells lacking mtDNA: Repopulation with exogenous mitochondria by complementation. Science 1989, 246, 500–504. [Google Scholar]
- Curbo, S.; Johansson, M.; Balzarini, J.; Lewis, L.; Karlsson, A. Acute cytotoxicity of arabinofuranosyl nucleoside analogs is not dependent on mitochondrial DNA. Exp. Cell Res. 2009, 315, 2539–2543. [Google Scholar]
- Field, J.J.; Singh, A.J.; Kanakkanthara, A.; Halafihi, T.; Northcote, P.T.; Miller, J.H. Microtubule-stabilizing activity of Zampanolide, a potent macrolide isolated from the Tongan marine sponge Cacospongia mycofijiensis. J. Med. Chem. 2009, 52, 7328–7332. [Google Scholar] [CrossRef]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS using oxidized DCFDA and flow-cytometry. Meth. Mol. Biol. 2010, 594, 57–72. [Google Scholar]
- Sashidhara, K.; White, K.; Crews, P.A. Selective account of effective paradigms and significant outcomes in the discovery of inspirational marine natural products. J. Nat. Prod. 2009, 72, 588–603. [Google Scholar]
- Xu, R.H.; Pelicano, H.; Zhou, Y.; Carew, J.S.; Feng, L.; Bhalla, K.N.; Keating, M.J.; Huang, P. Inhibition of glycolysis in cancer cells: A novel strategy to overcome drug resistance associated with mitochondrial respiratory defect and hypoxia. Cancer Res. 2005, 65, 613–621. [Google Scholar]
- Berridge, M.V.; Tan, A.S. Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT): Subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar] [CrossRef]
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction. Biotechnol. Annu. Rev. 2005, 11, 127–152. [Google Scholar]
- Herst, P.; Peterson, T.; Jeram, P.; Baty, J.; Berridge, M. The antiproliferative effects of phenoxodiol are associated with inhibition of plasma membrane electron transport in tumour cell lines and primary immune cells. Biochem. Pharmacol. 2007, 74, 1587–1595. [Google Scholar]
- Armstrong, J.S.; Hornung, B.; Lecane, P.; Jones, D.P.; Knox, S.J. Rotenone-induced G2/M cell cycle arrest and apoptosis in a human B lymphoma cell line PW. Biochem. Biophys. Res. Commun. 2001, 289, 973–978. [Google Scholar]
- Gonçalves, A.P.; Máximo, V.; Lima, J.; Singh, K.K.; Soares, P.; Videira, A. Involvement of p53 in cell death following cell cycle arrest and mitotic catastrophe induced by rotenone. Biochim. Biophys. Acta 2011, 1813, 492–499. [Google Scholar]
- Ross, D.; Kepa, J.; Winski, S.; Beall, H.; Anwar, A.; Siegel, D. NAD(P)H:Quinone oxidoreductase 1 (NQO1): Chemoprotection, bioactivation, gene regulation and genetic polymorphisms. Chem. Biol. Interact. 2000, 129, 77–97. [Google Scholar] [CrossRef]
- Ross, D. Quinone reductases multitasking in the metabolic world. Drug Metab. Rev. 2004, 36, 639–654. [Google Scholar]
- Bello, R.; Gomez-Diaz, C.; Navas, P.; Villalba, J. NAD(P)H:Quinone oxidoreductase 1 expression, hydrogen peroxide levels, and growth phase in HeLa cells. Meth. Enzymol. 2004, 382, 234–244. [Google Scholar]
- Tan, A.S.; Berridge, M.V. Evidence for NAD(P)H:Quinone oxidoreductase 1 (NQO1)-mediated quinone-dependent redox cycling via plasma membrane electron transport: A sensitive cellular assay for NQO1. Free Radic. Biol. Med. 2010, 48, 421–429. [Google Scholar]
- Srivastava, P.; Panda, D. Rotenone inhibits mammalian cell proliferation by inhibiting microtubule assembly through tubulin binding. FEBS J. 2007, 274, 4788–4801. [Google Scholar]
- Herst, P.; Berridge, M. Cell surface oxygen consumption: A major contributor to cellular oxygen consumption in glycolytic cell lines. Biochim. Biophys. Acta 2007, 1767, 170–177. [Google Scholar]
- Crume, K.P.; O’Sullivan, D.; Miller, J.H.; Northcote, P.T.; La Flamme, A.C. Delaying the onset of experimental autoimmune encephalomyelitis with the microtubule-stabilizing compounds, paclitaxel and peloruside A. J. Leuk. Biol. 2009, 86, 949–958. [Google Scholar]
- Samples Availability: Available from the authors.
Supplementary Files
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Meyer, K.J.; Singh, A.J.; Cameron, A.; Tan, A.S.; Leahy, D.C.; O’Sullivan, D.; Joshi, P.; La Flamme, A.C.; Northcote, P.T.; Berridge, M.V.; et al. Mitochondrial Genome-Knockout Cells Demonstrate a Dual Mechanism of Action for the Electron Transport Complex I Inhibitor Mycothiazole. Mar. Drugs 2012, 10, 900-917. https://doi.org/10.3390/md10040900
Meyer KJ, Singh AJ, Cameron A, Tan AS, Leahy DC, O’Sullivan D, Joshi P, La Flamme AC, Northcote PT, Berridge MV, et al. Mitochondrial Genome-Knockout Cells Demonstrate a Dual Mechanism of Action for the Electron Transport Complex I Inhibitor Mycothiazole. Marine Drugs. 2012; 10(4):900-917. https://doi.org/10.3390/md10040900
Chicago/Turabian StyleMeyer, Kirsten J., A. Jonathan Singh, Alanna Cameron, An S. Tan, Dora C. Leahy, David O’Sullivan, Praneta Joshi, Anne C. La Flamme, Peter T. Northcote, Michael V. Berridge, and et al. 2012. "Mitochondrial Genome-Knockout Cells Demonstrate a Dual Mechanism of Action for the Electron Transport Complex I Inhibitor Mycothiazole" Marine Drugs 10, no. 4: 900-917. https://doi.org/10.3390/md10040900