Anti-HCV RNA Aptamers Targeting the Genomic cis-Acting Replication Element

 and
and
Abstract
:1. Introduction
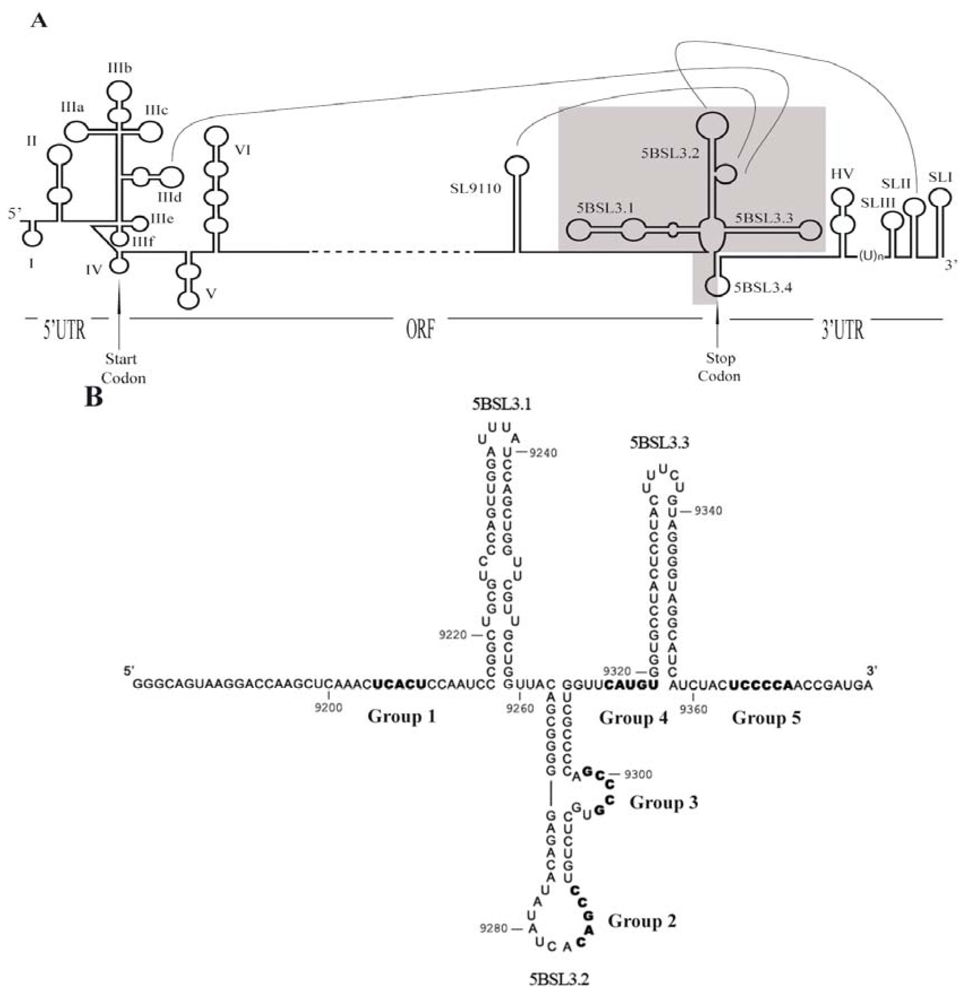
2. Experimental Section
2.1. Construction of the Initial RNA Population
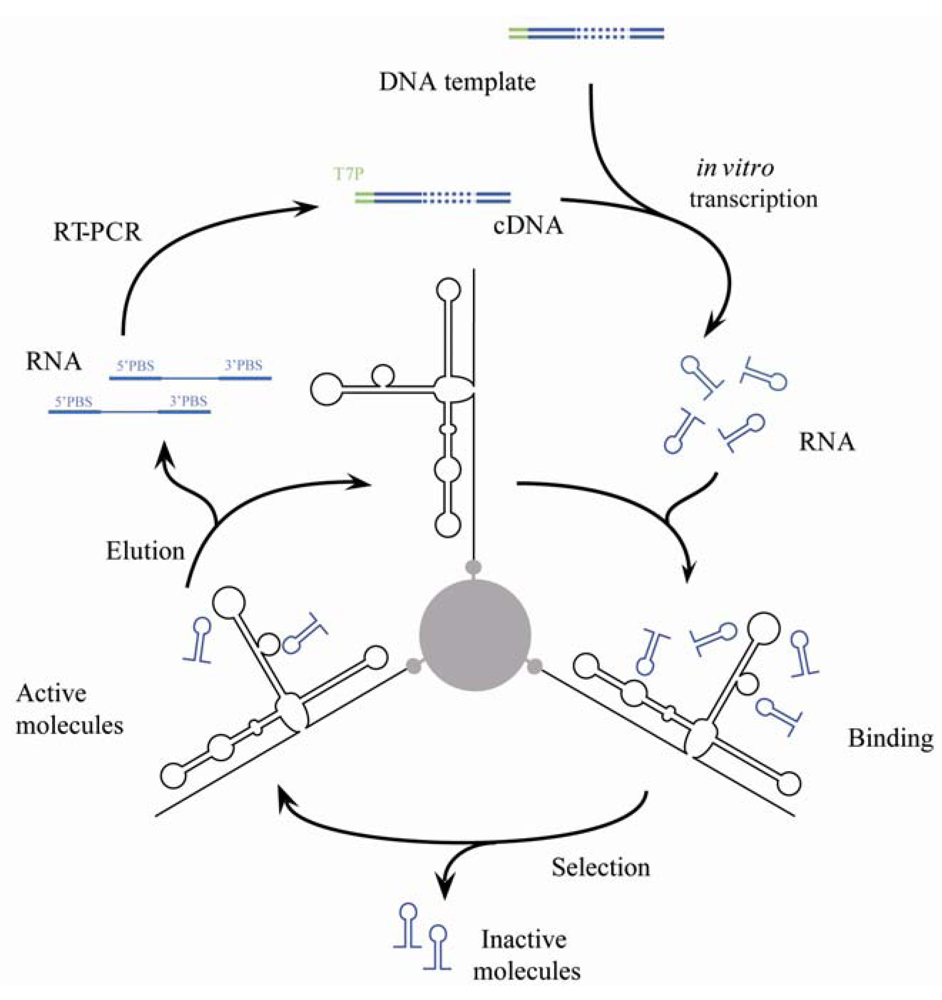
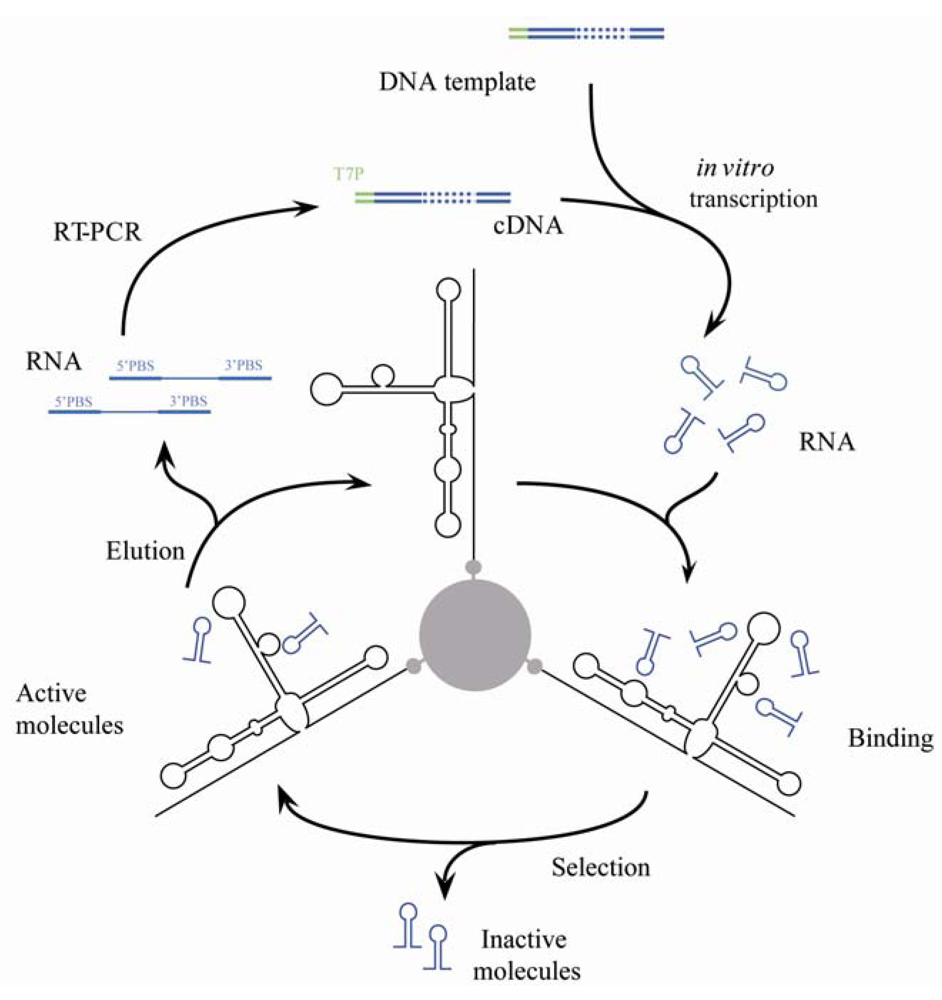
2.2. Selection of Aptamers
2.3. Amplification of Selected Molecules
2.4. Inhibition Assays of the HCV Replication
3. Results and Discussion
3.1. In Vitro Selection for Aptamers Targeting the HCV-CRE
3.2. Sequence Analysis of Selected RNA Molecules
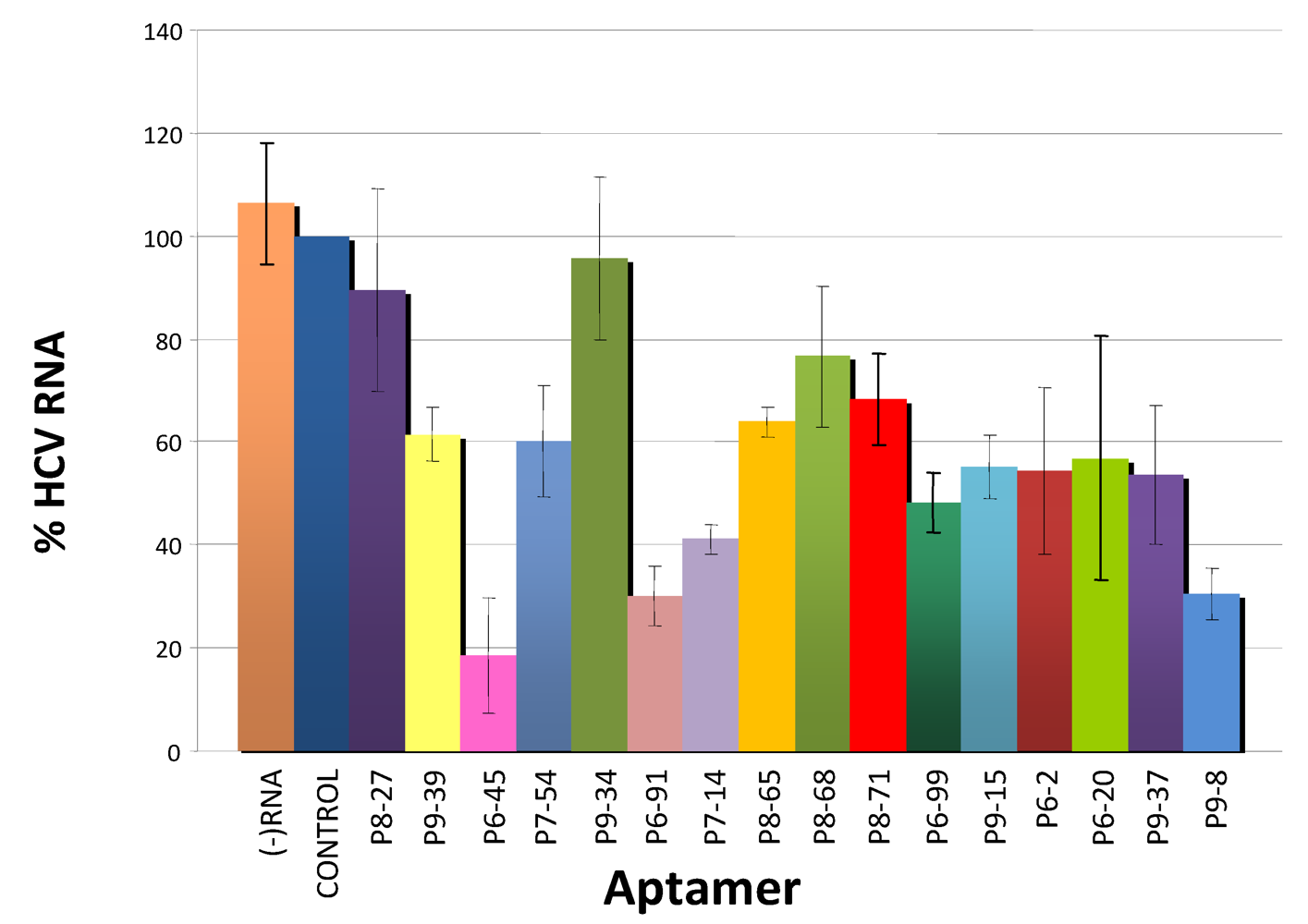
3.3. Inhibition of HCV Replication by Selected Aptamers in Cell Culture


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Aptamer | Sequence * |
|---|---|---|
| I (5'-RGUGRR-3' | P8-27 |  |
| P9-39 |  | |
| I y IV | P6-45 |  |
| II 5'-GGYUGUG-3' | P7-54 |  |
| P9-34 |  | |
| II y IV | P6-91 |  |
| III 5'-YGGGYR-3' | P7-14 |  |
| P8-65 |  | |
| P8-68 |  | |
| P8-71 |  | |
| III y IV | P6-99 |  |
| P9-15 |  | |
| IV 5'-YGUGUG-3' | P6-2 |  |
| P6-20 |  | |
| P9-37 |  | |
| V 5'-UGGGGR-3' | P9-8 |  |
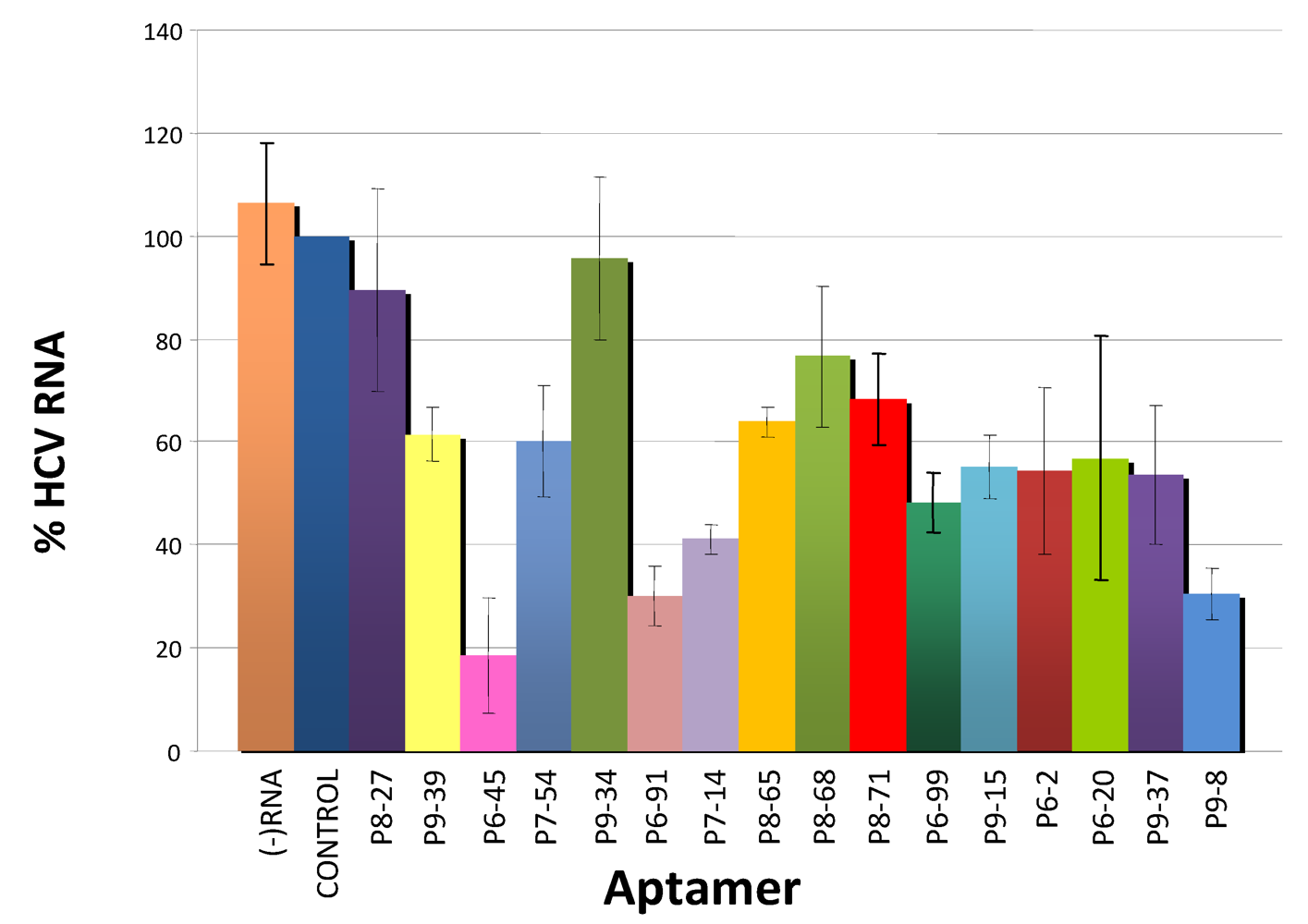
4. Conclusions
Acknowledgements
References and Notes
- Choo, Q.L.; Kuo, G.; Weiner, A.J.; Overby, L.R.; Bradley, D.W.; Houghton, M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science 1989, 244, 359–362. [Google Scholar]
- Poynard, T.; Yuen, M.F.; Ratziu, V.; Lai, C.L. Viral hepatitis C. Lancet 2003, 362, 2095–2100. [Google Scholar]
- Fried, M.W.; Shiffman, M.L.; Reddy, K.R.; Smith, C.; Marinos, G.; Goncales, F.L., Jr.; Haussinger, D.; Diago, M.; Carosi, G.; Dhumeaux, D.; et al. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. N. Engl. J. Med. 2002, 347, 975–982. [Google Scholar] [CrossRef]
- Domingo, E.; Gómez, J. Quasispecies and its impact on viral hepatitis. Virus Res. 2007, 127, 131–150. [Google Scholar] [CrossRef]
- Bukh, J.; Purcell, R.H.; Miller, R.H. Sequence analysis of the 5' noncoding region of hepatitis C virus. Proc. Natl. Acad. Sci. USA 1992, 89, 4942–4946. [Google Scholar]
- Friebe, P.; Bartenschlager, R. Genetic analysis of sequences in the 3' nontranslated region of hepatitis C virus that are important for RNA replication. J. Virol. 2002, 76, 5326–5338. [Google Scholar] [CrossRef]
- Friebe, P.; Lohmann, V.; Krieger, N.; Bartenschlager, R. Sequences in the 5' nontranslated region of hepatitis C virus required for RNA replication. J. Virol. 2001, 75, 12047–12057. [Google Scholar]
- Kolykhalov, A.A.; Mihalik, K.; Feinstone, S.M.; Rice, C.M. Hepatitis C virus-encoded enzymatic activities and conserved RNA elements in the 3' nontranslated region are essential for virus replication in vivo. J. Virol. 2000, 74, 2046–2051. [Google Scholar]
- Tsukiyama-Kohara, K.; Iizuka, N.; Kohara, M.; Nomoto, A. Internal ribosome entry site within hepatitis C virus RNA. J. Virol. 1992, 66, 1476–1483. [Google Scholar]
- Wang, C.; Sarnow, P.; Siddiqui, A. Translation of human hepatitis C virus RNA in cultured cells is mediated by an internal ribosome-binding mechanism. J. Virol. 1993, 67, 3338–3344. [Google Scholar]
- Yi, M.; Lemon, S.M. 3' nontranslated RNA signals required for replication of hepatitis C virus RNA. J. Virol. 2003, 77, 3557–3568. [Google Scholar] [CrossRef]
- Yi, M.; Lemon, S.M. Structure-function analysis of the 3' stem-loop of hepatitis C virus genomic RNA and its role in viral RNA replication. RNA 2003, 9, 331–345. [Google Scholar] [CrossRef]
- Lee, H.; Shin, H.; Wimmer, E.; Paul, A.V. cis-Acting RNA signals in the NS5B C-terminal coding sequence of the hepatitis C virus genome. J. Virol. 2004, 78, 10865–10877. [Google Scholar] [CrossRef]
- You, S.; Stump, D.D.; Branch, A.D.; Rice, C.M. A cis-acting replication element in the sequence encoding the NS5B RNA-dependent RNA polymerase is required for hepatitis C virus RNA replication. J. Virol. 2004, 78, 1352–1366. [Google Scholar]
- Friebe, P.; Boudet, J.; Simorre, J.P.; Bartenschlager, R. Kissing-loop interaction in the 3' end of the hepatitis C virus genome essential for RNA replication. J. Virol. 2005, 79, 380–392. [Google Scholar]
- You, S.; Rice, C.M. 3' RNA elements in hepatitis C virus replication: Kissing partners and long poly(U). J. Virol. 2008, 82, 184–195. [Google Scholar] [CrossRef]
- Zhang, J.; Yamada, O.; Sakamoto, T.; Yoshida, H.; Araki, H.; Murata, T.; Shimotohno, K. Inhibition of hepatitis C virus replication by pol III-directed overexpression of RNA decoys corresponding to stem-loop structures in the NS5B coding region. Virology 2005, 342, 276–285. [Google Scholar] [CrossRef]
- Diviney, S.; Tuplin, A.; Struthers, M.; Armstrong, V.; Elliott, R.M.; Simmonds, P.; Evans, D.J. A hepatitis C virus cis-acting replication element forms a long-range RNA-RNA interaction with upstream RNA sequences in NS5B. J. Virol. 2008, 82, 9008–9022. [Google Scholar]
- Romero-López, C.; Berzal-Herranz, A. The functional RNA domain 5BSL3.2 within the NS5B coding sequence influences hepatitis C virus IRES-mediated translation. Cell. Mol. Life Sci. 2011. [Google Scholar]
- Romero-López, C.; Berzal-Herranz, A. A long-range RNA-RNA interaction between the 5' and 3' ends of the HCV genome. RNA 2009, 15, 1740–1752. [Google Scholar] [CrossRef]
- Marton, S.; Reyes-Darias, J.A.; Sánchez-Luque, F.J.; Romero-López, C.; Berzal-Herranz, A. In vitro and ex vivo selection procedures for identifying potentially therapeutic DNA and RNA molecules. Molecules 2010, 15, 4610–4638. [Google Scholar]
- Puerta-Fernández, E.; Romero-López, C.; Barroso-delJesus, A.; Berzal-Herranz, A. Ribozymes: Recent advances in the development of RNA tools. FEMS Microbiol. Rev. 2003, 27, 75–97. [Google Scholar] [CrossRef]
- Romero-López, C.; Sánchez-Luque, F.J.; Berzal-Herranz, A. Targets and tools: Recent advances in the development of anti-HCV nucleic acids. Infect. Disord. Drug Targets 2006, 6, 121–145. [Google Scholar] [CrossRef]
- Toulme, J.J.; Darfeuille, F.; Kolb, G.; Chabas, S.; Staedel, C. Modulating viral gene expression by aptamers to RNA structures. Biol. Cell 2003, 95, 229–238. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar]
- Ducongè, F.; Toulme, J.J. In vitro selection identifies key determinants for loop-loop interactions: RNA aptamers selective for the TAR RNA element of HIV-1. RNA 1999, 5, 1605–1614. [Google Scholar] [CrossRef]
- Fukuda, K.; Toyokawa, Y.; Kikuchi, K.; Konno, K.; Ishihara, R.; Fukazawa, C.; Nishikawa, S.; Hasegawa, T. Isolation of RNA aptamers specific for the 3' X tail of HCV. Nucleic Acids Symp. Ser. (Oxf.) 2008, 2, 205–206. [Google Scholar]
- Kikuchi, K.; Umehara, T.; Fukuda, K.; Kuno, A.; Hasegawa, T.; Nishikawa, S. A hepatitis C virus (HCV) internal ribosome entry site (IRES) domain III-IV-targeted aptamer inhibits translation by binding to an apical loop of domain IIId. Nucleic Acids Res. 2005, 33, 683–692. [Google Scholar]
- Kolb, G.; Reigadas, S.; Castanotto, D.; Faure, A.; Ventura, M.; Rossi, J.J.; Toulme, J.J. Endogenous expression of an anti-TAR aptamer reduces HIV-1 replication. RNA Biol. 2006, 3, 150–156. [Google Scholar] [CrossRef]
- Romero-López, C.; Díaz-González, R.; Berzal-Herranz, A. Inhibition of hepatitis C virus internal ribosome entry site-mediated translation by an RNA targeting the conserved IIIf domain. Cell. Mol. Life Sci. 2007, 64, 2994–3006. [Google Scholar] [CrossRef]
- Romero-López, C.; Díaz-González, R.; Barroso-delJesus, A.; Berzal-Herranz, A. Inhibition of hepatitis C virus replication and internal ribosome entry site-dependent translation by an RNA molecule. J. Gen. Virol. 2009, 90, 1659–1669. [Google Scholar] [CrossRef]
- Tallet-Lopez, B.; Aldaz-Carroll, L.; Chabas, S.; Dausse, E.; Staedel, C.; Toulme, J.J. Antisense oligonucleotides targeted to the domain IIId of the hepatitis C virus IRES compete with 40S ribosomal subunit binding and prevent in vitro translation. Nucleic Acids Res. 2003, 31, 734–742. [Google Scholar] [CrossRef]
- Lohmann, V.; Korner, F.; Koch, J.; Herian, U.; Theilmann, L.; Bartenschlager, R. Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science 1999, 285, 110–113. [Google Scholar] [CrossRef]
- Theissen, G.; Richter, A.; Lukacs, N. Degree of biotinylation in nucleic acids estimated by a gel retardation assay. Anal. Biochem. 1989, 179, 98–105. [Google Scholar]
- Larrea, E.; Aldabe, R.; Molano, E.; Fernandez-Rodriguez, C.M.; Ametzazurra, A.; Civeira, M.P.; Prieto, J. Altered expression and activation of signal transducers and activators of transcription (STATs) in hepatitis C virus infection: In vivo and in vitro studies. Gut 2006, 55, 1188–1196. [Google Scholar]
- Martell, M.; Gómez, J.; Esteban, J.I.; Sauleda, S.; Quer, J.; Cabot, B.; Esteban, R.; Guardia, J. High-throughput real-time reverse transcription-PCR quantitation of hepatitis C virus RNA. J. Clin. Microbiol. 1999, 37, 327–332. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Marton, S.; Berzal-Herranz, B.; Garmendia, E.; Cueto, F.J.; Berzal-Herranz, A. Anti-HCV RNA Aptamers Targeting the Genomic cis-Acting Replication Element. Pharmaceuticals 2012, 5, 49-60. https://doi.org/10.3390/ph5010049
Marton S, Berzal-Herranz B, Garmendia E, Cueto FJ, Berzal-Herranz A. Anti-HCV RNA Aptamers Targeting the Genomic cis-Acting Replication Element. Pharmaceuticals. 2012; 5(1):49-60. https://doi.org/10.3390/ph5010049
Chicago/Turabian StyleMarton, Soledad, Beatriz Berzal-Herranz, Eva Garmendia, Francisco J. Cueto, and Alfredo Berzal-Herranz. 2012. "Anti-HCV RNA Aptamers Targeting the Genomic cis-Acting Replication Element" Pharmaceuticals 5, no. 1: 49-60. https://doi.org/10.3390/ph5010049