Construction of a nrdA::luxCDABE Fusion and Its Use in Escherichia coli as a DNA Damage Biosensor

1
College of Life Sciences and Biotechnology, Korea University, Anam-dong, Seongbuk-Gu, Seoul, Republic of Korea
2
Diagnostics Group, Institut Pasteur Korea, Hawolgok-dong, Seongbuk-gu, Seoul, Republic of Korea
*
Author to whom correspondence should be addressed.
Sensors 2008, 8(2), 1297-1307; https://doi.org/10.3390/s8021297
Submission received: 31 January 2008
/
Accepted: 21 February 2008
/
Published: 22 February 2008
(This article belongs to the Special Issue Utilization of Electrochemical Sensors and Biosensors in Biochemistry and Molecular Biology)
Abstract
:The promoter of nrdA gene which is related with DNA synthesis was used to construct a DNA damage sensitive biosensor. A recombinant bioluminescent E. coli strain, BBTNrdA, harboring a plasmid with the nrdA promoter fused to the luxCDABE operon, was successfully constructed. Its response to various chemicals including genotoxic chemicals substantiates it as a DNA damage biosensor. In characterization, three different classes of toxicants were used: DNA damaging chemicals, oxidative stress chemicals, and phenolics. BBTNrdA only responded strongly to DNA damaging chemicals, such as nalidixic acid (NDA), mitomycin C (MMC), 1-methyl-1-nitroso-N-methylguanidine (MNNG), and 4-nitroquinoline N-oxide (4-NQO). In contrast, there were no responses from the oxidative stress chemicals and phenolics, except from hydrogen peroxide (H2O2) which is known to cause DNA damage indirectly. Therefore, the results of the study demonstrate that BBTNrdA can be used as a DNA damage biosensor.
1. Introduction
Due to environmental pollution, specific and sensitive detection methods are in need for environmental contaminants. To investigate the impact of toxic agents on organisms, biological test systems have been developed and applied to in various means. These bioassays are thought as good genotoxicity assessment, as results, rapid and sensitive detection methods for mutagenic condition are in urgent demand for the screening of an overwhelming number of existing substances that are potential DNA-damaging agents. The most widely used indicator, the Ames test is disadvantageous due to the long operation time needed. To overcome that, SOS-dependent bacterial test systems is used for DNA-damaging agents, and their response for those chemicals is known as SOS response. The umu-test is the system induced by that SOS response. That employs a fusion between the umuCD promoter and lacZ gene from Escherichia coli. But the umu-test also has a weak point in that is has a low sensitivity [1].
In response to these problems, other recombinant bacterial sensors were developed. Several of these biosensors have been characterized and widely used, for instance, in specific stress identification and bio-imaging [2-3]. Such sensors contained a variety of reporter genes, such as luxCDABE [4-5], the green fluorescent protein (GFP) [6], luxAB [7] and luc [8]. Among these, the luxCDABE genes can be used to generate bioluminescence in vivo without the need for an extraneous addition of substrate. There have been many reports describing the advantages of luxCDABE, such as its simplicity of analysis and applicability in detecting multiple samples [9-10]. Furthermore, the reaction time needed to generate the bioluminescent responses is very short. Using this procedure, the recA, sulA, umuCD and recN promoters have previously been fused with the luxCDABE genes and the strains carrying these fusions have been used widely in toxicity assays [11-14]. Furthermore, the use of such fusions can be used to study the functionality of a given promoter. Consequently, the nrdA gene was selected for further study as a genotoxic biomarker in part due to its functioning in DNA synthesis but also since it is not regulated by the SOS response in E. coli.
The nrdA gene is well known and encodes for the ribonucleoside diphosphate reductase protein, which is involved in DNA synthesis in Escherichia coli. The ribonucleoside diphosphate reductase is composed of two subunits, referred to as B1 and B2 [15]. Ribonucleoside diphosphate reductase converts ribonucleotides to deoxyribonucleotides and, in this process, oxidizes the thiol group [16]. As well, the expression of the nrdA gene is strongly affected by DNA damage, such as after an exposure to UV light, but is not dependent on LexA [17]. To date, many research groups have studied this gene and its protein and have deduced its function, structure and mechanism, but all of these studies only focused on the molecular aspects of this gene and its protein [18-21].
Therefore, in this study we developed BBTNrdA, a cell-based genotoxicity sensor which is specific in its responses to genotoxins. This E. coli strain harbors a plasmid with the nrdA promoter fused to the luxCDABE operon. Characterization of this strain was performed using exposures to DNA damaging chemicals, oxidative stress-inducing chemicals and phenolics. The results clearly show that BBTNrdA strongly responded to only genotoxic compounds.
2. Results and Discussion
2.1. Response of strain BBTNrdA to DNA damaging chemicals
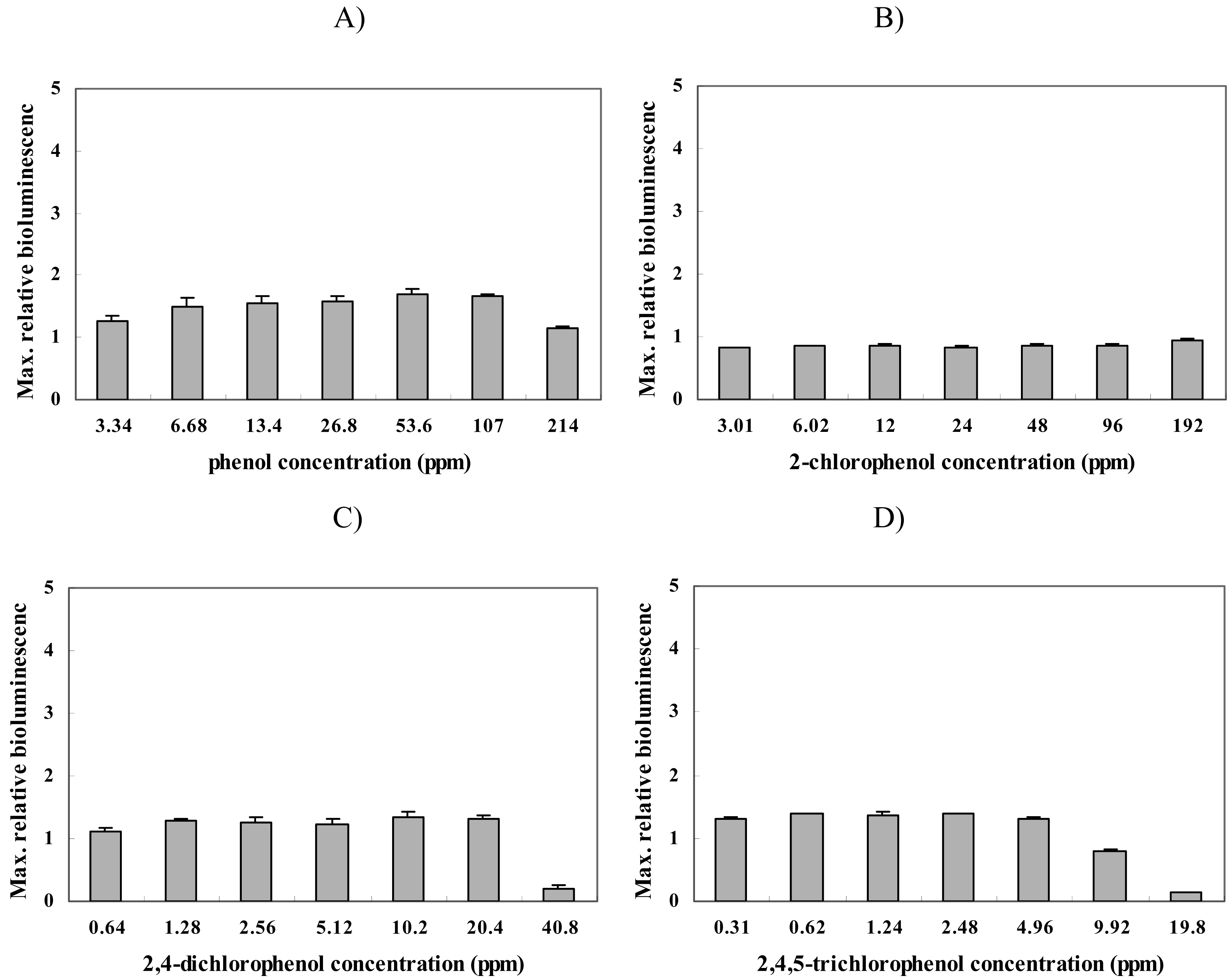
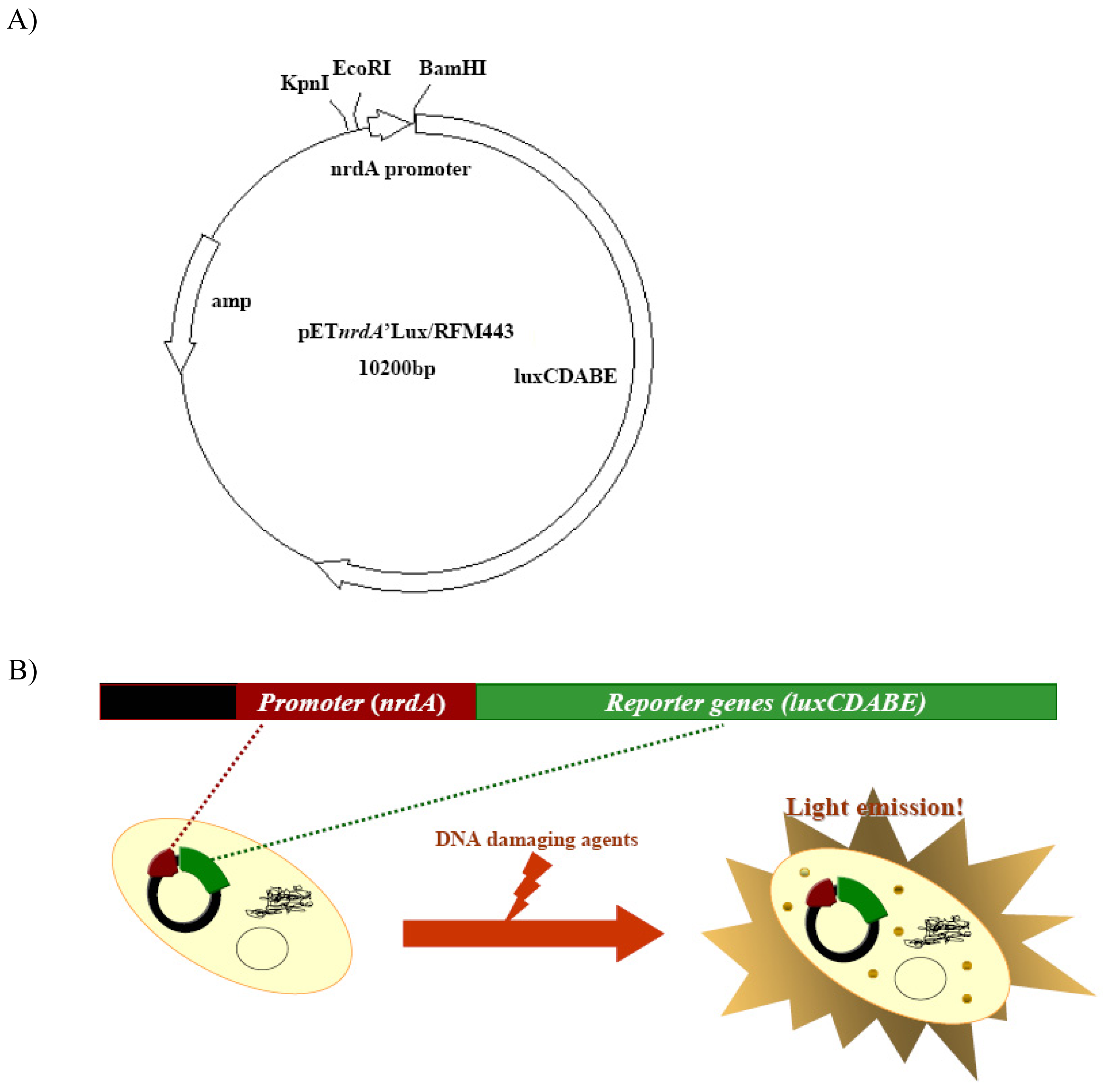
Strain BBTNrdA was constructed by fusing the nrdA promoter with the luxCDABE operon in plasmid pDEW201 and transforming this plasmid into E. coli strain RFM443. This proposed sensor is shown in Figure 1.
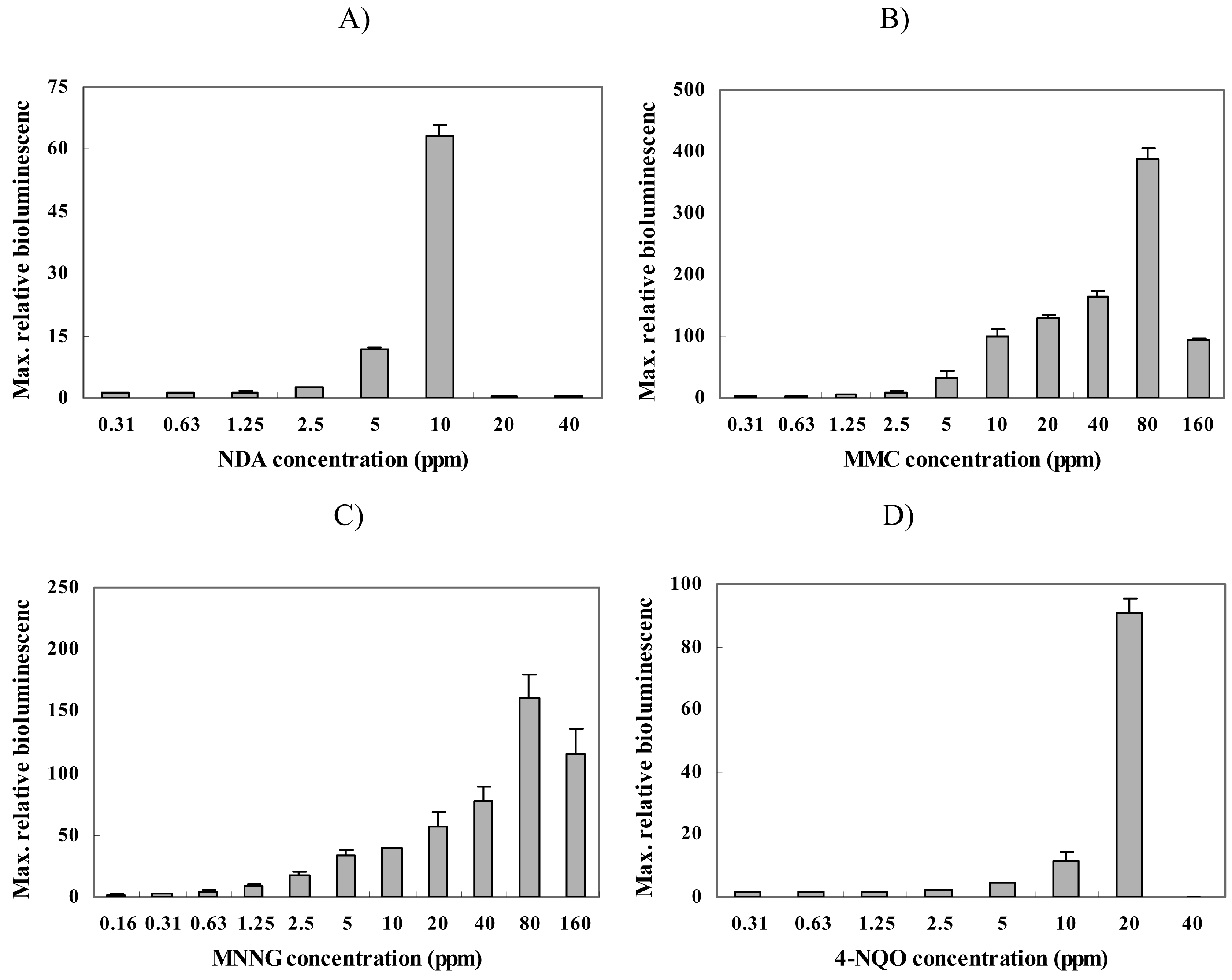
To characterize strain BBTNrdA, we used four different chemicals that are known DNA mutagens [22], i.e., nalidixic acid (NDA), mitomycin C (MMC), 1-methyl-1-nitroso-N-methylguanidine (MNNG), and 4-nitroquinoline N-oxide (4-NQO). Figure 2 shows the responses from this strain for each chemical. Initially, strain BBTNrdA was characterized with NDA and the maximum responses were seen at a concentration of 10 ppm (Figure 2 A), while the minimum detectable concentration (MDC) was 2.5 ppm (Table 1). The strong induction in the RBL (∼65-fold) of this strain clearly shows that this strain was responsive to NDA, which is known to inhibit the synthesis of DNA [23]. Since the nrdA genes encode for a protein specifically involved in DNA synthesis, the responses of strain BBTNrdA to NDA are not surprising.
Likewise, we tested MMC, and found that strain responded very strongly and in a dose-dependent manner (Figure 2 B), with a MRC and MDC of 80 and 0.3125 ppm (Table 1), respectively. In contrast with NDA, MMC crosslinks the two strands of DNA and, as a result, induces apoptosis and arrests the cell cycle in eukaryotic cells [24-26]. From the findings presented here, it is clear that the damage caused by MMC also leads to an induction of the nrdA gene.
Lastly, strain BBTNrdA was exposed to 1-methyl-1-nitroso-N-methylguanidine (MNNG) and 4-nitroquinoline N-oxide (4-NQO). The results for these responses are shown in Figure 2 C and D, respectively. As with MMC and NDA, the bioluminescence of strain BBTNrdA was strongly induced by these compounds and in a dose-dependent manner. MMNG causes alkylation of the cellular proteins and DNA, leading to errors being incorporated in the DNA during replication and repair [27-28]. 4-NQO is also DNA damaging chemical which affects DNA in various ways [29] and its mechanism is similar to the damage caused by exposure to UV [8, 30]. BBTNrdA showed an increase in its bioluminescent emission.
2.2. Response of strain BBTNrdA to other chemicals
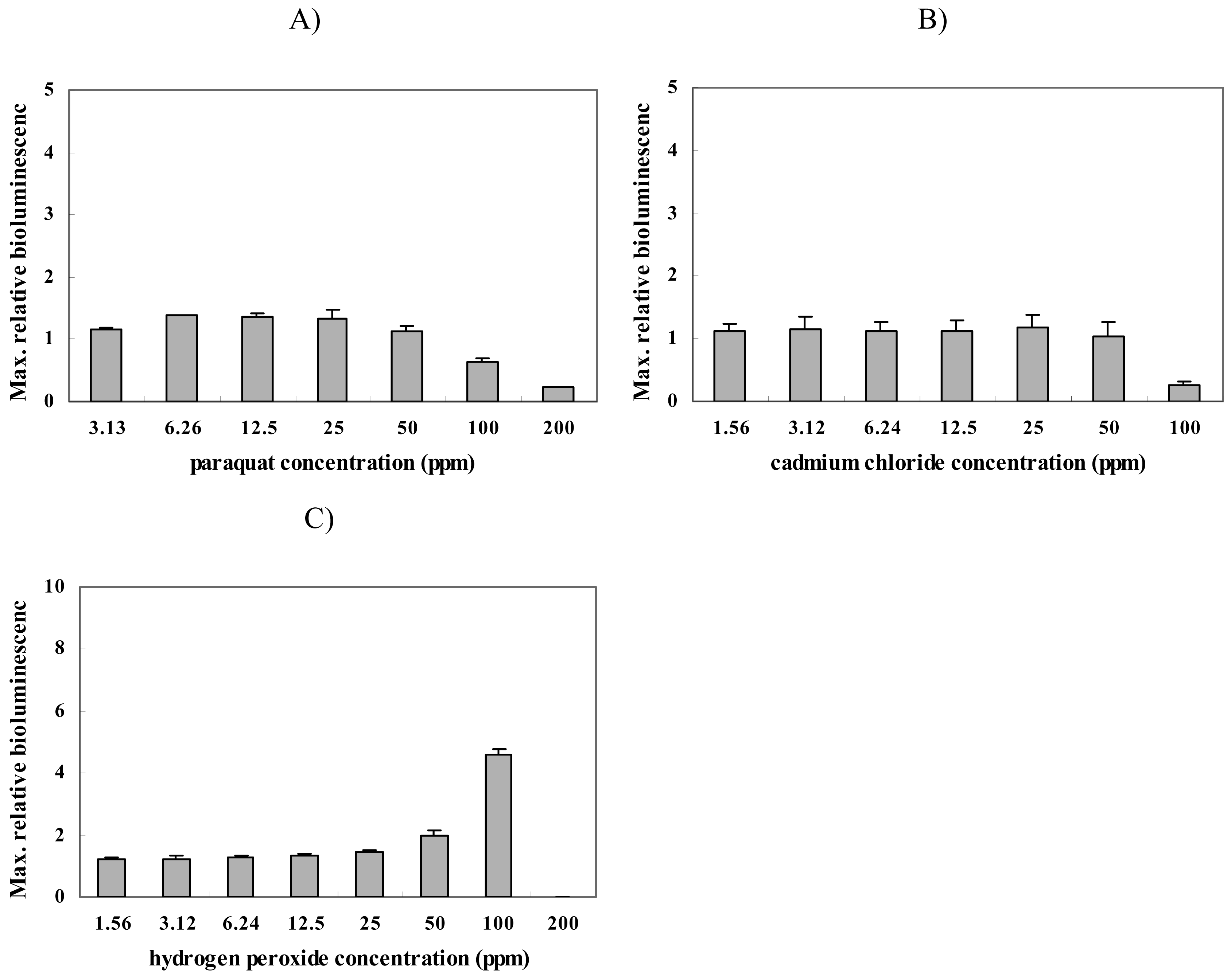
We performed toxicity tests using oxidative stress-inducing chemicals to characterize the nrdA gene expression level when E. coli is exposed to oxidative radicals [31-32]. For this, paraquat, cadmium chloride and hydrogen peroxide (H2O2) were used. Strain BBTNrdA gave no response when exposed to parquat or cadmium chloride (Figure 3 A and B), but a mild response was seen with a hydrogen peroxide (H2O2) exposure (Figure 2 C). It is not surprising that the nrdA::luxCDABE responded to H2O2 since some reports showed that strain DPD2794, another DNA damage-sensitive biosensor, also responded to this compound [25]. Of course, the response mechanisms of each gene for a given chemical are different, but from our results it is clear is that H2O2 can lead to DNA damage.
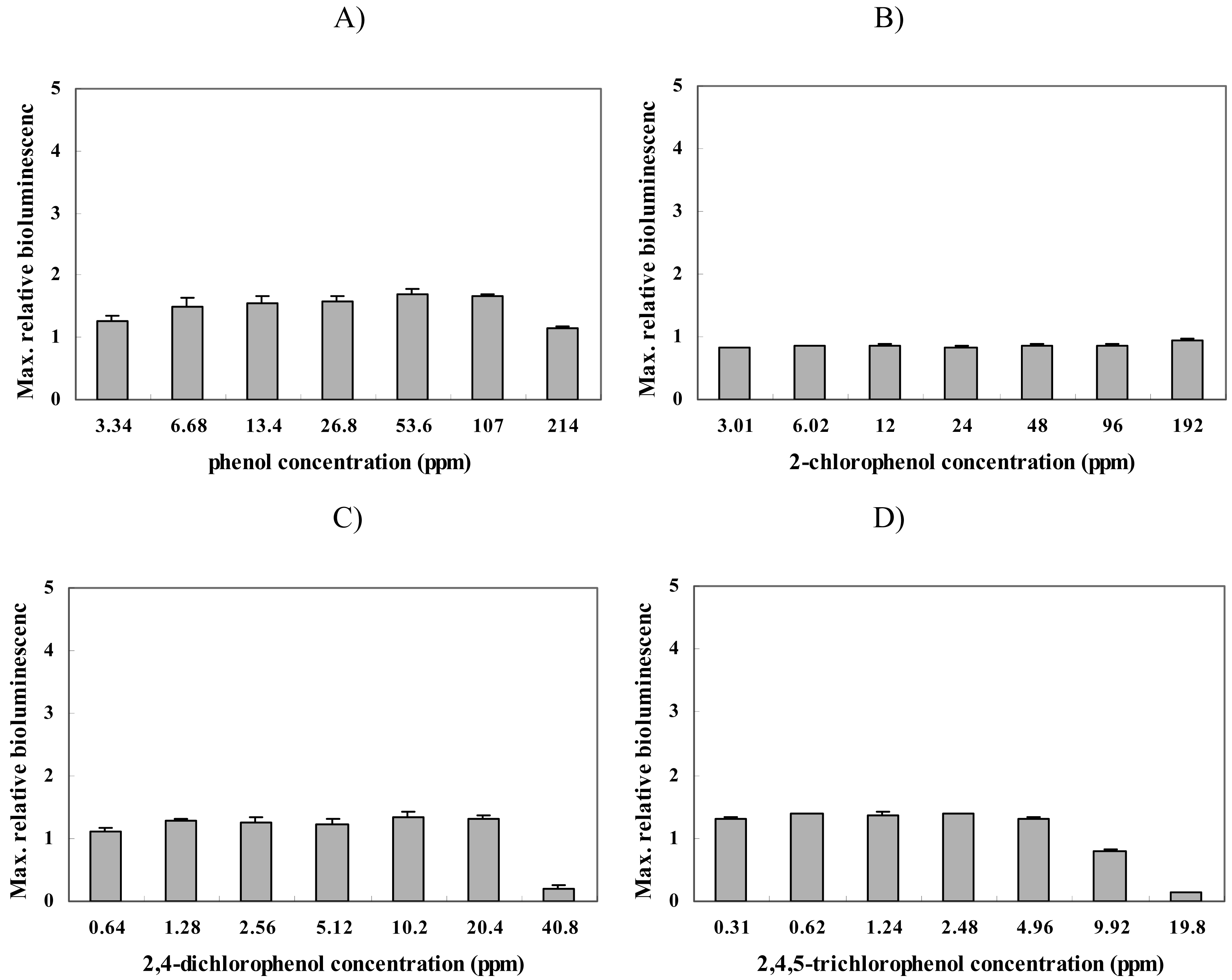
Furthermore, additional experiments were conducted using membrane-damaging chemicals, i.e., phenol, 2-chlorophenol (2-CP), 2,4-dichlorophenol (2,4-DCP), and 2,4,5-trichlorophenol (2,4,5-TCP) [34]. Figure 4 shows that there was no response to these chemicals. This was expected since these compounds should have no effect on the structure or replication of the cellular DNA. Taken together with the results from the oxidative compounds, these results demonstrate that the nrdA gene expression level is not induced by membrane damaging or oxidative toxicants, but only by DNA damaging compounds.
Organisms often encounter abnormal and potentially harmful environments. In response to such conditions, bacterial cells alter their gene expression patterns and, depending on the stress experienced, the production level of specific stress proteins increases to help defend and repair the cells. Consequently, recombinant bioluminescent bacteria are useful tools for examining for toxicants. Depending on the type of promoter region used in the construction of the recombinant bioluminescent bacteria, a variety of sensors can be constructed that respond to a multitude of environmental stimuli. As such, these strains can be utilized to study the mechanisms and classification of specific stress experienced within a given environment. Furthermore, in addition to the analysis of the responses being simple, since it is possible to detect changes in the gene expression levels through the emission of light, these biosensors have other advantages, including their simple usage, quick response time and multiplexing capabilities. As such, they are a powerful tool when applied together in arrays and in the screening of unknown substances.
In this study, we describe a recombinant nrdA-dependent bioluminescent strain that is responsive to DNA damaging chemicals. The results found that BBTNrdA only responds to genotoxic chemicals and hydrogen peroxide, which is known to cause DNA damage indirectly, and there was no significant induction in the bioluminescence of this strain when exposed to the other oxidative stress- and membrane damage-inducing compounds. These results demonstrate that strain BBTNrdA can be used as a sensor for genotoxic conditions.
The nrdA gene encodes for the ribonucleoside diphosphate reductase protein. Consequently, BBTNrdA responds more sensitively to substances related with inhibition of DNA synthesis. Furthermore, it has the potential to be used as a specific biosensor to detect errors during the conversion of ribonucleotides to deoxyribonucleotides in Escherichia coli. Owing to these properties, this strain can be used as a biosensor for detecting genotoxic agents and can be applied in the prescreening of anticancer drugs, which often are used to inhibit DNA synthesis.
3. Experimental Section
3.1. Construction of strain BBTNrdA
Escherichia coli strain RFM443 (strR, laK2, ac△74; was used as a host strain [34]. To construct the recombinant plasmid, pET, the sequence of the nrdA promoter region obtained from the National Center for Biotechnology Information (NCBI) was amplified by PCR using genomic DNA from strain RFM443. The primers used to amplify the nrdA promoter region are 5′-agcagcgaattcaagaaatcgccgaa-3′ and 5′-acttaaggatccgcaatagtttcatg-3′. They include BamH1 and EcoR1 (NEB, USA) restriction sites. After the promoter region was amplified, it was ligated into pDEW201 [35], a promoter-less plasmid with the Photorhabdus luminescens luxCDABE operon, to yield pETnrdA'Lux/RFM443. This plasmid was then transformed into Escherichia coli strain RFM443. and measuring their bioluminescent response. A positive clone that showed good responses when exposed to 2 ppm of mitomycin C (MMC) was selected and named BBTNrdA.
3.2. Culture Condition and Chemical Test Protocol
For the cell culture, one colony of strain BBTNrdA was grown overnight in 3 mL Luria-Bertani (LB) medium (DIFCO, USA) supplemented with 50 μg/ml ampicillin (Sigma, USA) in a 15mL Falcon tube (Corning, MA, USA) in a shaking incubator at 37°C and 200 rpm. In 100 mL of fresh Luria-Bertani (LB) medium (DIFCO, USA) supplemented with 50 μg/ml ampicillin (Sigma, USA), 2 mL of the overnight culture was inoculated and grown until the optical density reached 0.8 at 600 nm, measured using a spectrophotometer (Lambda 12, Perin Elmer, USA). Afterwards, the procedure was repeated one more time, but this time the optical density was allowed to reach only 0.08 at 600 nm. Using this BBTNrdA culture, a 96-well luminometer test was performed. For the experiment the target chemicals were serially diluted and 10 μl were added to each well. Then, 190 μl of the BBTNrdA culture solution was transferred to each of wells in the 96-well plate (Microlite 1, DYNEX Technologies, USA) which already contained known concentrations of the target chemicals. For the control, 10 μl of distilled water was added in place of the target chemicals. This 96-well plate was then placed in the luminometer chamber under controlled conditions (37°C) and constant shaking before measuring the bioluminescence. The measurement interval was at set for every 6 minutes for a total time of 4 hours.
3.3. Chemicals
In this study, all chemicals were purchased from the Sigma-Aldrich Company (USA). They were categorized into three groups depending on the specific stress that they cause. First, the DNA damaging chemicals were nalidixic acid (NDA), mitomycin C (MMC), 1-methyl-1-nitroso-N-methylguanidine (MNNG) and 4-nitroquinoline N-oxide (4-NQO). Likewise, the oxidative stress chemicals included paraquat, cadmium chloride, and hydrogen peroxide (H2O2). The last group was membrane damaging chemicals and included several phenolics - phenol, 2-chlorophenol (2-CP), 2,4-dichlorophenol (2,4-DCP) and 2,4,5-trichlorophenol (2,4,5-TCP).
3.4. Data Analysis
All experiments were conducted three times independently and the results are shown as the average with the standard deviations. The relative bioluminescence (RBL) was recorded after exposure to toxicant and is defined as the bioluminescence value of the test sample divided by that of the control for each time point. To further evaluate the responses for each chemical, the minimum detectable concentration (MDC), which represents the lowest concentration showing an RBL value of at least two, was used. As well, the concentration giving the maximum RBL value is referred to as the maximum response concentration (MRC). Using these two parameters, we characterized the responses of strain BBTNrdA with each of the compounds.
Acknowledgments
This research was supported by the Korea-Israeli Joint Fund Program of the Ministry of Science and Technology (MOST). The authors are grateful for the support.
References and Notes
- Ptitsyn, L. R.; Horneck, G.; Komova, O.; Kozubek, S.; Krasavin, E. A.; Bonev, M.; Rettberg, P. A Biosensor for Environmental Genotoxin Screening Based on an SOS lux Assay in Recombinant Escherichia coli Cells. Appl. Environ. Microbiol. 1997, 63, 4377–4384. [Google Scholar]
- Ahn, J. M.; Kim, B. C.; Gu, M. B. Characterization of gltA::luxCDABE fusion in Escherichia coli as a Toxicity Biosensor. Biotechnol. Bioprocess. Eng. 2006, 11, 16–521. [Google Scholar]
- Kim, B. C.; Youn, C. H.; Ahn, J. M.; Gu, M. B. Screening of target-specific stress-responsive genes for the development of cell-based biosensors using a DNA microarray. Anal. Chem. 2005, 77, 8020–8026. [Google Scholar]
- Billard, P.; DuBow, M. S. Bioluminescence-based assays for detection and characterization of bacteria and chemicals in clinical laboratories. Clin Biochem. 1998, 31, 1–14. [Google Scholar]
- Stewart, G. S; Williams, P. Lux genes and the applications of. bacterial bioluminescence. J. Gen. Microbiol. 1992, 138, 1289–1300. [Google Scholar]
- Morin, J. G.; Hastings, J. W. Energy transfer in a bioluminescent system. J. Cell. Physiol. 1971, 77, 313–318. [Google Scholar]
- Meighen, E. A. Molecular biology of bacterial bioluminescence. Microbiol. Mol. Biol. Rev. 1991, 55, 123–142. [Google Scholar]
- Gu, M. B.; Mitchell, R. J.; Kim, B. C. Whole cell-based biosensors for environmental biomonitoring and application. Adv. Biochem. Eng. Biotechnol. 2004, 87, 269–305. [Google Scholar]
- Belkin, S. Microbial whole-cell sensing systems ofenvironmental pollutants. Curr. Opin. Microbiol. 2003, 6, 206–212. [Google Scholar]
- Daunert, S.; Barrett, G.; Feliciano, J. S.; Shetty, R. S.; Shrestha, S.; Smith-Spencer, W. Genetically engineered whole-cell sensing systems: coupling biological recognition with reporter genes. Chem. Rev. 2000, 100, 2705–2738. [Google Scholar]
- Mitchell, R.J.; Gu, M. B. An Escherichia coli biosensor capable of detecting both genotoxic and oxidative damage. Appl. Microbiol. Biotechnol. 2004, 64, 46–52. [Google Scholar]
- Norman, A.; Hansen, L. H.; Sørensen, S. J. Construction of a ColD cda promoter-based SOS-green fluorescent protein whole-cell biosensor with higher sensitivity toward genotoxic compounds than constructs based on recA, umuDC, or sulA promoters. Appl Environ Microbiol. 2005, 71, 2338–2346. [Google Scholar]
- Baumstark-Khan, C.; Cioara, K.; Rettberg, P.; Horneck, G. Determination of geno- and cytotoxicity of groundwater and sediments using the recombinant SWITCH test. J Environ Sci Health. Part A. 2005, 40, 245–263. [Google Scholar]
- Rabbow, E.; Rettberg, P.; Baumstark-Khan, C.; Horneck, G. SOS-LUX- and LAC-FLUORO-TEST for the quantification of genotoxic and/or cytotoxic effects of heavy metal salts. Anal Chim Act. 2002, 456, 31–39. [Google Scholar]
- Lars, T. Physicochemical Characterization of Ribonucleoside Diphosphate Reductase from Escherichia coli. J. Biol. Che. 1973, 248, 4591–1601. [Google Scholar]
- Stubbe, J. Ribonucleotide Reductases: Amazing and Confusing. J Biol Chem. 1990, 265, 5329–5332. [Google Scholar]
- Courcelle, J.; Khodursky, A.; Peter, B.; Brown, P.O. P.; Hanawalt, C. Comparative gene expression profiles following UV exposure in wild-type and SOS-deficient. Escherichia coli, Genetics. 2001, 158, 41–64. [Google Scholar]
- Erikson, S.; Sjoberg, B.; Karlstrom, O.; Hahne, S. Ribonucleoside Diphosphate Reductase from Escherichia coli. J Biol Chem. 1977, 252, 6132–6138. [Google Scholar]
- Guarino, E.; Jimenez-Sanchez, A.; Guzman, E. C. Defective Ribonucleoside Diphosphate Reductase Impairs Replication Fork Progression in Escherichia coli. J. Bacteriol. 2007, 189, 3496–3501. [Google Scholar]
- Stubbe, J.; Krenitsky, T.; Ator, M. Mechanism of Ribonucleoside Diphosphate Reductase from Echerichia coli. J Biol Chem. 1983, 258, 1625–1630. [Google Scholar]
- Merino, E.; Bolivar, F. The ribonucleoside diphosphate reductase gene (nrdA) of Escherichia coli carries a repetitive extragenicpalindromic (REP) sequence in its 3′ structural terminus. Mol. Microbiol. 1989, 3, 839–841. [Google Scholar]
- Min, J. H.; Gu, M. B. Acclimation and repair of DNA damage in recombinant bioluminescent Escherichia coli. J. Appl. Micorbiol. 2003, 95, 479–483. [Google Scholar]
- Goss, W. A.; Deitz, W. H.; Cook, T. M. Mechanism of Action of Nalidixic Acid on Escherichia coli. J. Bacterio. 1965, 89, 1068–1074. [Google Scholar]
- Tomasz, M. Mitomycin C: small, fast and deadly (but very selective). Chemistry & Biology 1995, 2, 575–579. [Google Scholar]
- Min, J. H.; Kim, E. J.; LaRossa, R. A.; Gu, M. B. Distinct responses of a recA::luxCDABE Escherichia coli strain to direct and indirect DNA damaging agents. Mutat. Res. 1999, 442, 61–68. [Google Scholar]
- McKenna, D. J.; Gallus, M. S.; McKeown, R. C.; Downes, S.; McKelvey-Martin, V. J. Modification of the alkaline Comet assay to allow simultaneous evaluation of mitomycin C-induced DNA cross-link damage and repair of specific DNA sequences in RT4 cells. DNA Repair (Amst.) 2003, 2, 879–890. [Google Scholar]
- Lee, C.; Michael, D. T. Enhanced Resistance to Nitrosoguanidine Killing and Mutagenesis in a DNA Gyrase Mutant of Escherichia coli. J. Bacteriol. 1982, 151, 764–770. [Google Scholar]
- Roy, M. K.; Kuwabara, Y.; Hara, K.; Watanabe, Y.; Tamai, Y. Antimutagenic effect of amino acids on the mutagenicity of N-methyl-N'-nitro-N-nitrosoguanidine (MNNG). Biosci. Biotechnol. Biochem. 2002, 66, 1400–1402. [Google Scholar]
- Dai, H.; Ichikawa-Ryo, H.; Kondo, S. Dose-rate dependence of mutagenesis by ultraviolet radiation and 4-nitroquinoline 1-oxide in Escherichia coli. Jpn. J. Genet. 1983, 58, 283–295. [Google Scholar]
- Poot, M.; Gollahon, K. A.; Emond, M. J.; Silber, J. R.; Rabinovitch, P. S. Werner syndrome diploid fibroblasts are sensitive to 4-nitroquinoline-N-oxide and 8-methoxypsoralen: implications for the disease phenotype1. The FASEB Journal. 2002, 16, 757–758. [Google Scholar]
- Mitchell., R. J.; Gu, M. B. Construction and characterization of novel dual stress-responsive bacterial biosensors. Biosens. Bioelectron. 2004, 19, 977–985. [Google Scholar]
- Min, J. H.; Gu, M. B. Adaptive Responses of Escherichia coli for Oxidative and Protein Damages Using Bioluminescence Reporter. J. Microbiol. Biotechn. 2004, 14, 466–469. [Google Scholar]
- Choi, S. H.; Gu, M. B. Phenolic toxicitydetection and classification through the use of a recombinant bioluminescent Escherichia coli. Environ. Toxicol. Chem. 2001, 20, 248–255. [Google Scholar]
- Drolet, M.; Phoenix, P.; Menzel, R.; Masse, E.; Liu, L. F.; Crouch, R. J. Overexpression of RNase H partially complements the growth defect of an Escherichia coli Δ topA mutant: R-loop formation is a major problem in the absence of DNA topoisomerase I. Proc. Natl. Acad. Sci. 1995, 92, 3526–3530. [Google Scholar]
- Vandyk, T. K.; Rosson, R. A. Photorhabdus luminescens luxCDABE promoter probe vectors. Methods Mol. Biol. 1995, 102, 85–95. [Google Scholar]
Figure 1.
Schematic of the proposed biosensor in this study A) plasmid map of BBTNrdA and B) principle of its responses to DNA damaging agents.
Figure 1.
Schematic of the proposed biosensor in this study A) plasmid map of BBTNrdA and B) principle of its responses to DNA damaging agents.
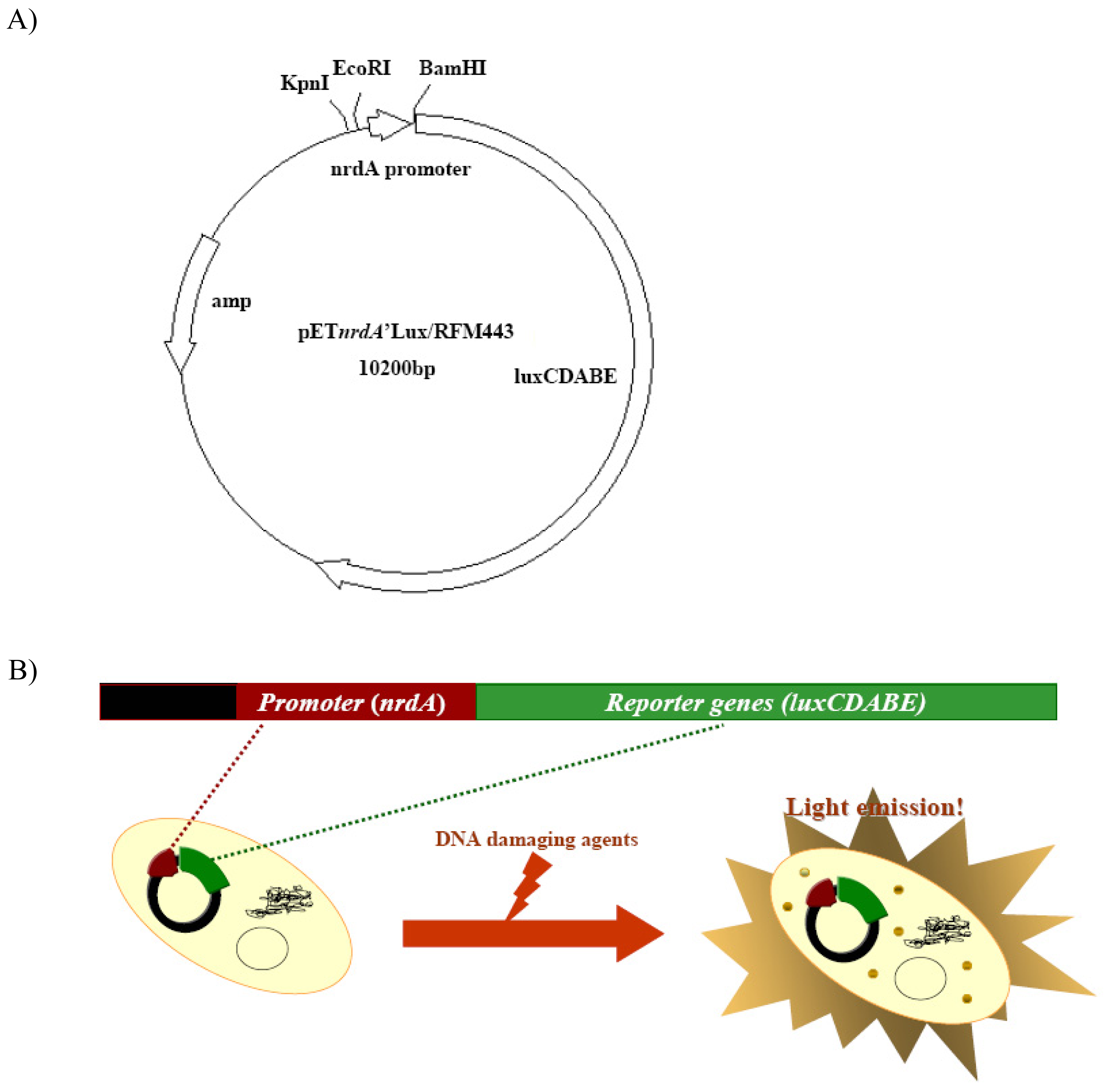
Figure 2.
Maximum relative luminescence values seen from strain BBTNrdA after being exposed to different concentration of (A) nalidixic acid (NDA), (B) mitomycin C (MMC), (C) 1-methyl-1-nitroso-N-methylguanidine (MNNG), and (D) 4-nitroquinoline N-oxide (4-NQO).
Figure 2.
Maximum relative luminescence values seen from strain BBTNrdA after being exposed to different concentration of (A) nalidixic acid (NDA), (B) mitomycin C (MMC), (C) 1-methyl-1-nitroso-N-methylguanidine (MNNG), and (D) 4-nitroquinoline N-oxide (4-NQO).

Figure 3.
Maximum relative luminescence values seen from strain BBTNrdA after being exposed to different concentrations of (A) paraquat, (B) cadmium chloride and (C) hydrogen peroxide.
Figure 3.
Maximum relative luminescence values seen from strain BBTNrdA after being exposed to different concentrations of (A) paraquat, (B) cadmium chloride and (C) hydrogen peroxide.
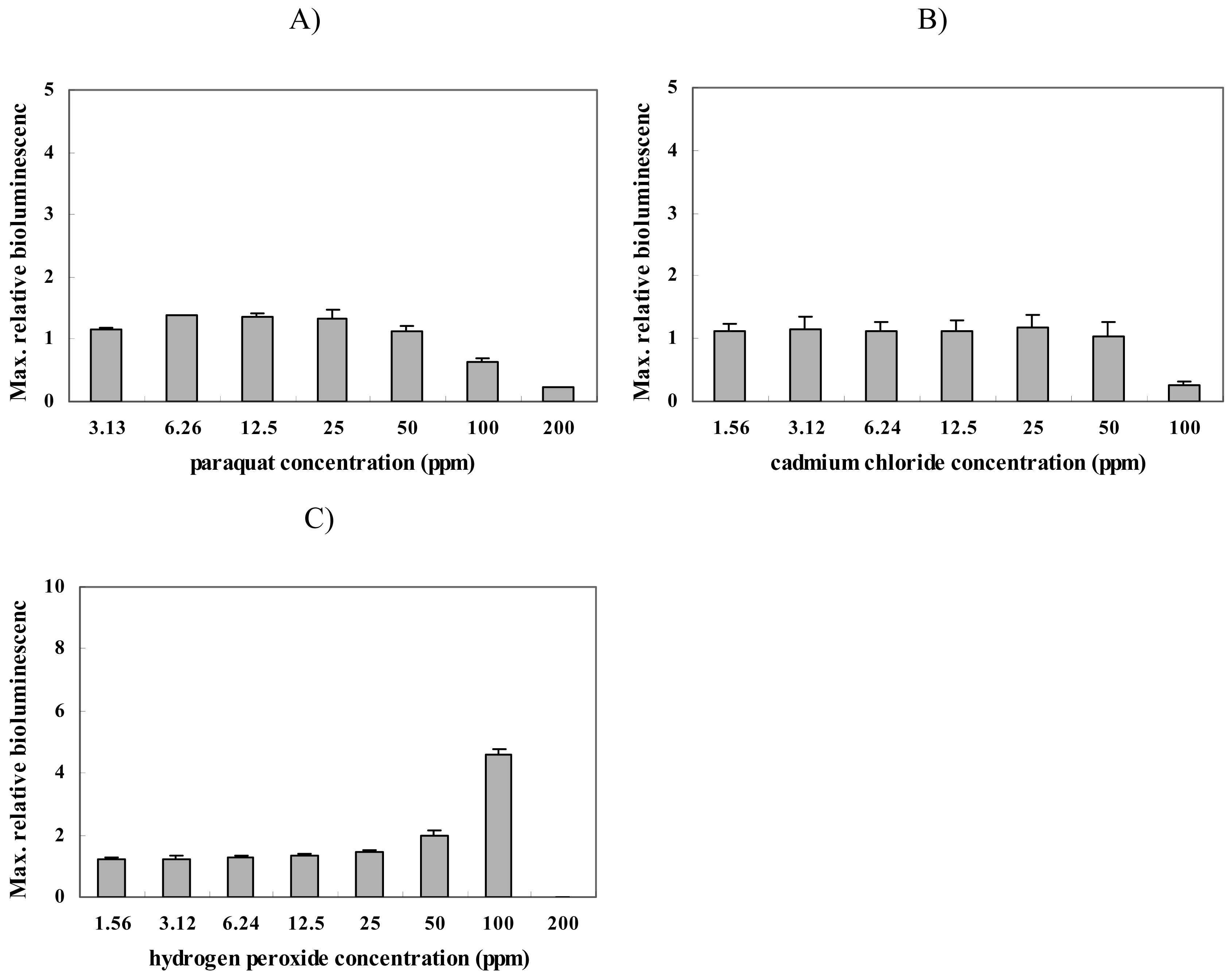
Figure 4.
Maximum relative luminescence values seen from strain BBTNrdA after exposure to different concentration of (A) phenol, (B) 2-chlorophenol (2-CP), (C) 2,4-dichlorophenol (2,4-DCP) and (D) 2,4,5-trichlorophenol (2,4,5-TCP).
Figure 4.
Maximum relative luminescence values seen from strain BBTNrdA after exposure to different concentration of (A) phenol, (B) 2-chlorophenol (2-CP), (C) 2,4-dichlorophenol (2,4-DCP) and (D) 2,4,5-trichlorophenol (2,4,5-TCP).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories | Chemicals | MDC (ppm) | MRC (ppm) |
|---|---|---|---|
| DNA damage | nalidixic acid | 2.5 | 10 |
| mitomycin C | 0.3125 | 80 | |
| 1-methyl-1-nitroso-N-methylguanidine | 0.1563 | 80 | |
| 4-nitroquinoline N-oxide | 2.5 | 20 | |
| Oxidative damage | paraquat | ND | ND |
| cadmium chloride | ND | ND | |
| hydrogen peroxide | 50 | 100 | |
| Phenol compounds | phenol | ND | ND |
| 2-chlorophenol | ND | ND | |
| 2,4-dichlorophenol | ND | ND | |
| 2,4,5-trichlorophenol | ND | ND |
aMDC : Minimum Detectable Concentration (ND: Not Detected)
bMRC : Maximum Response Concentration (ND: Not Detected)
© 2008 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
MDPI and ACS Style
Hwang, E.T.; Ahn, J.-M.; Kim, B.C.; Gu, M.B. Construction of a nrdA::luxCDABE Fusion and Its Use in Escherichia coli as a DNA Damage Biosensor. Sensors 2008, 8, 1297-1307. https://doi.org/10.3390/s8021297
AMA Style
Hwang ET, Ahn J-M, Kim BC, Gu MB. Construction of a nrdA::luxCDABE Fusion and Its Use in Escherichia coli as a DNA Damage Biosensor. Sensors. 2008; 8(2):1297-1307. https://doi.org/10.3390/s8021297
Chicago/Turabian StyleHwang, Ee Taek, Joo- Myung Ahn, Byoung Chan Kim, and Man Bock Gu. 2008. "Construction of a nrdA::luxCDABE Fusion and Its Use in Escherichia coli as a DNA Damage Biosensor" Sensors 8, no. 2: 1297-1307. https://doi.org/10.3390/s8021297