Array Biosensor for Toxin Detection: Continued Advances

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
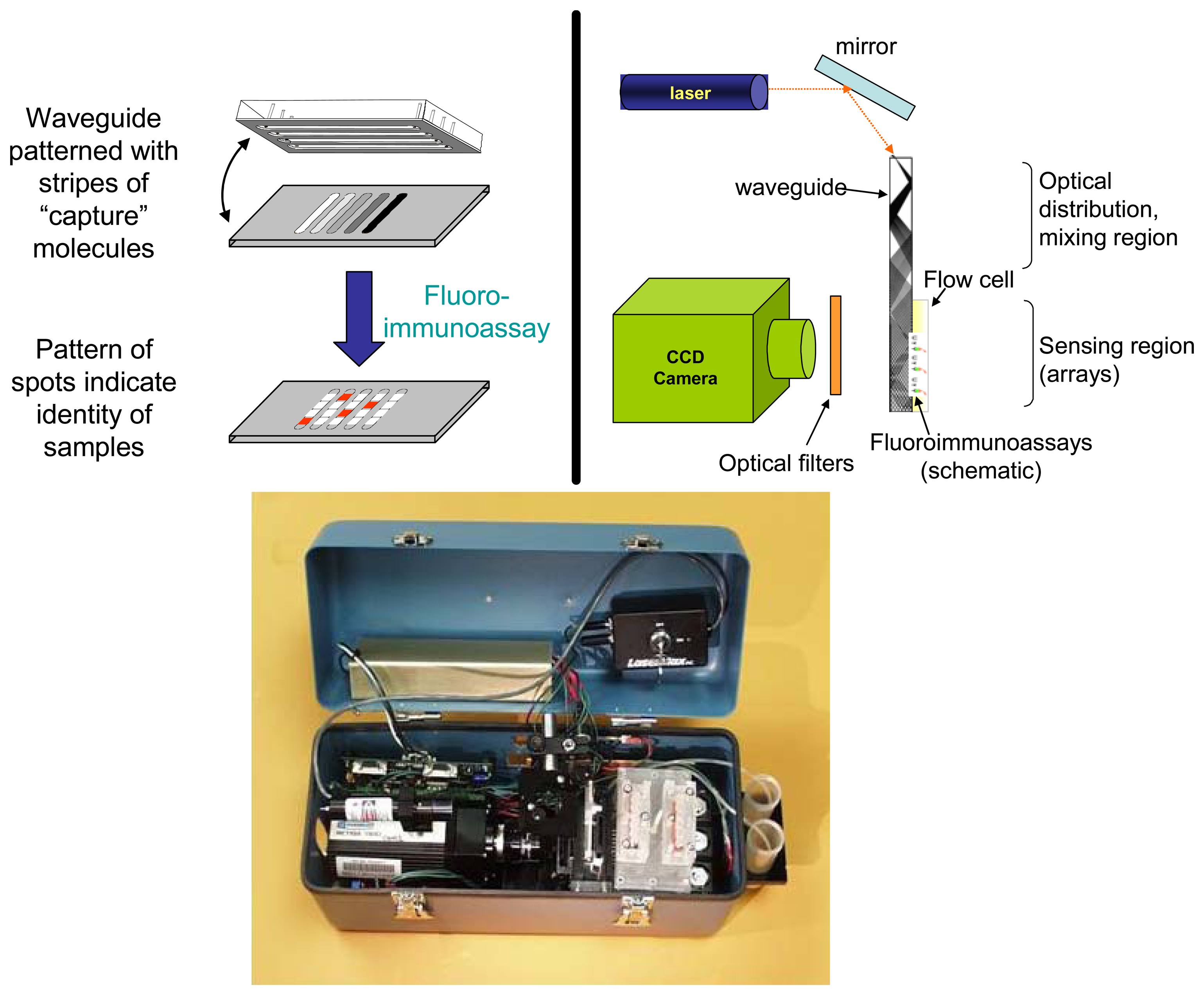
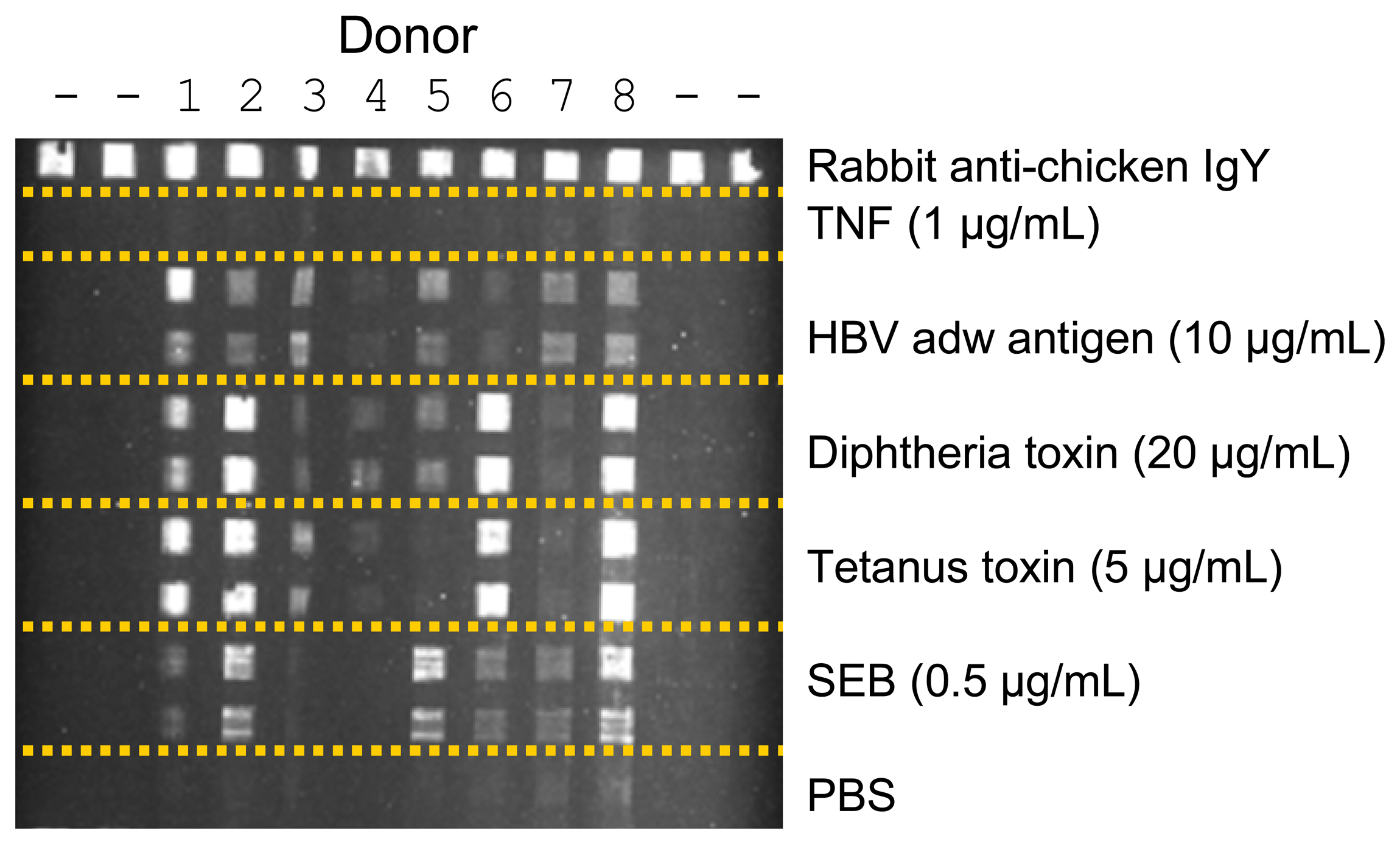
:1. Introduction
2. Applications for food, environmental testing
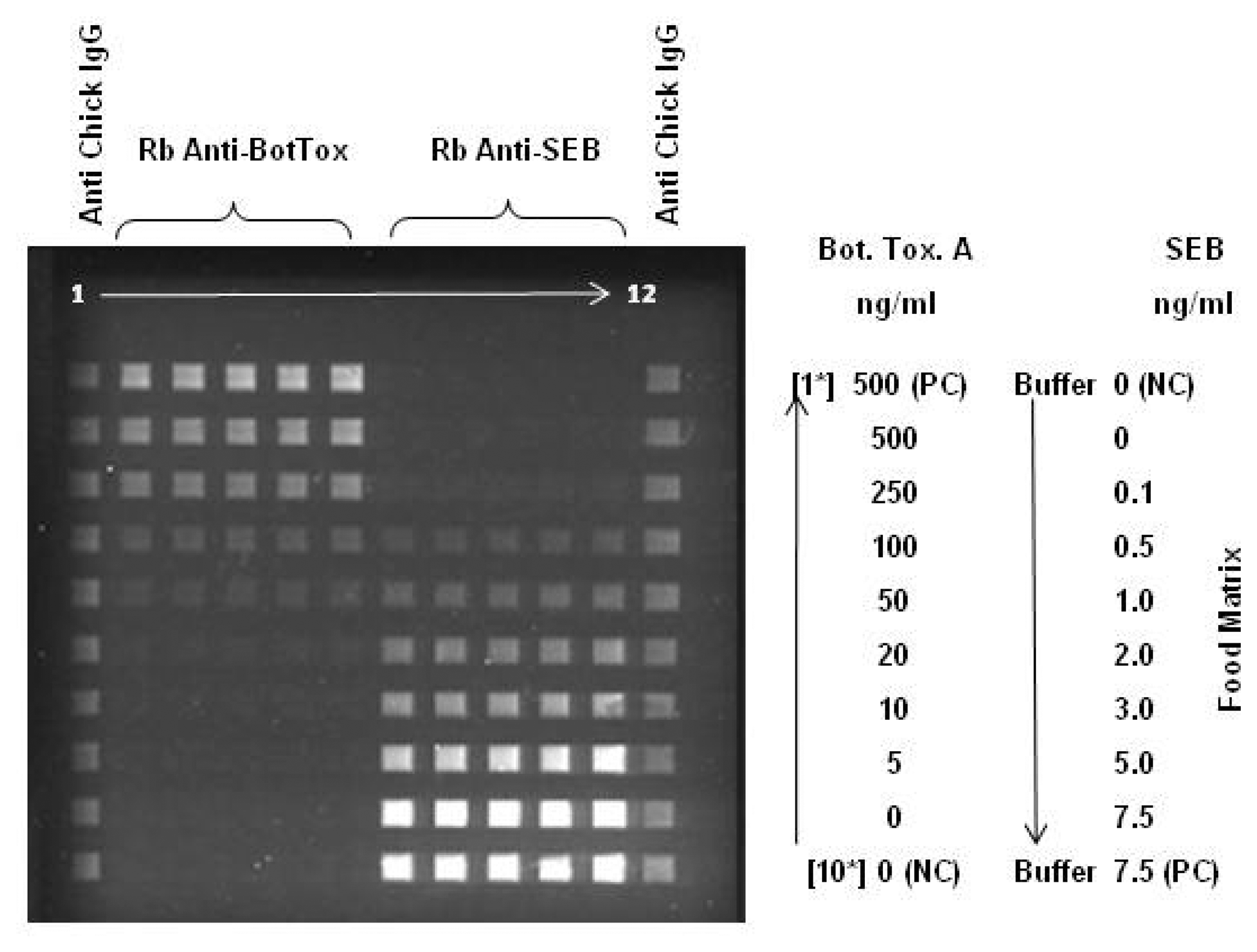
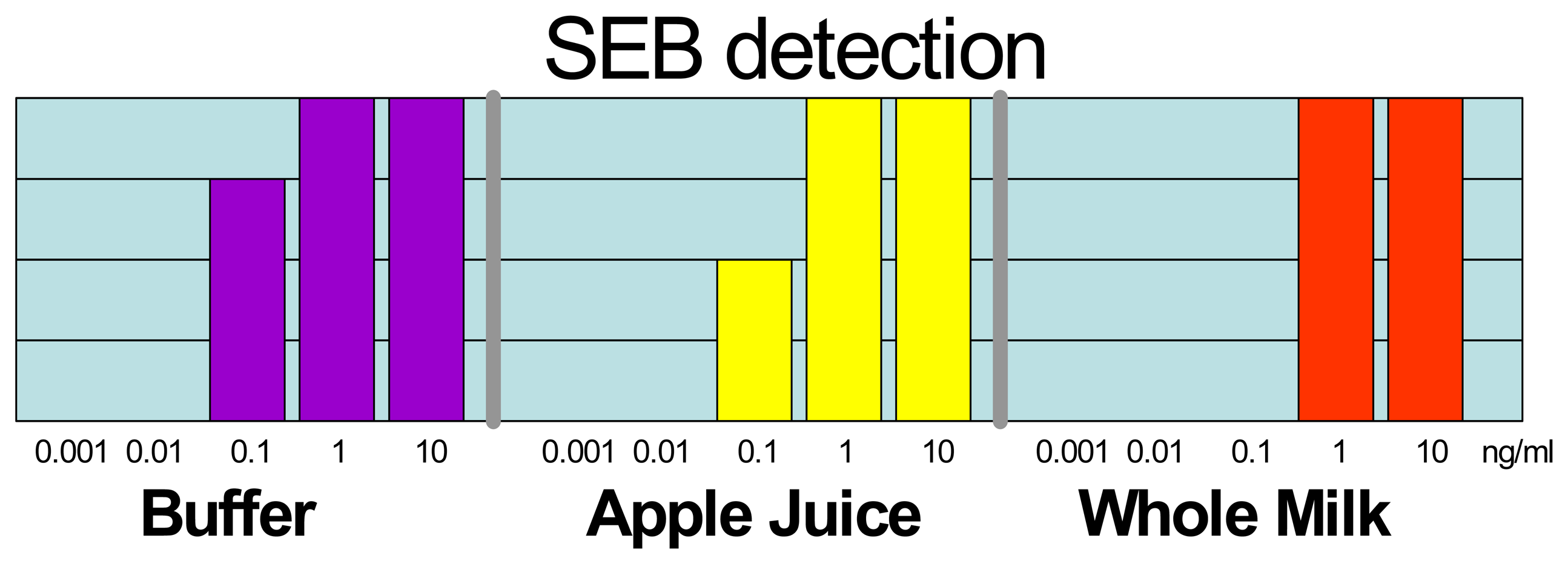
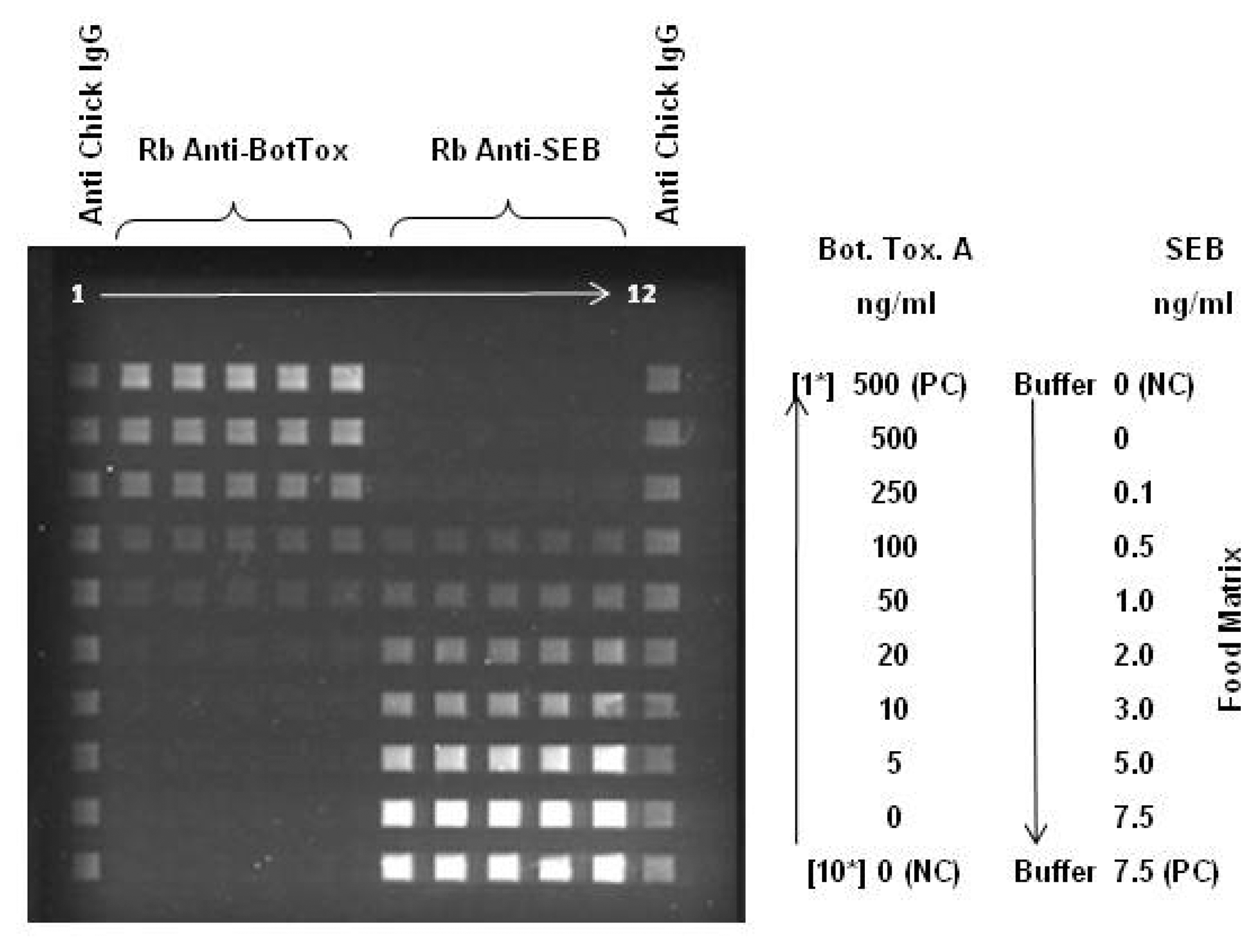
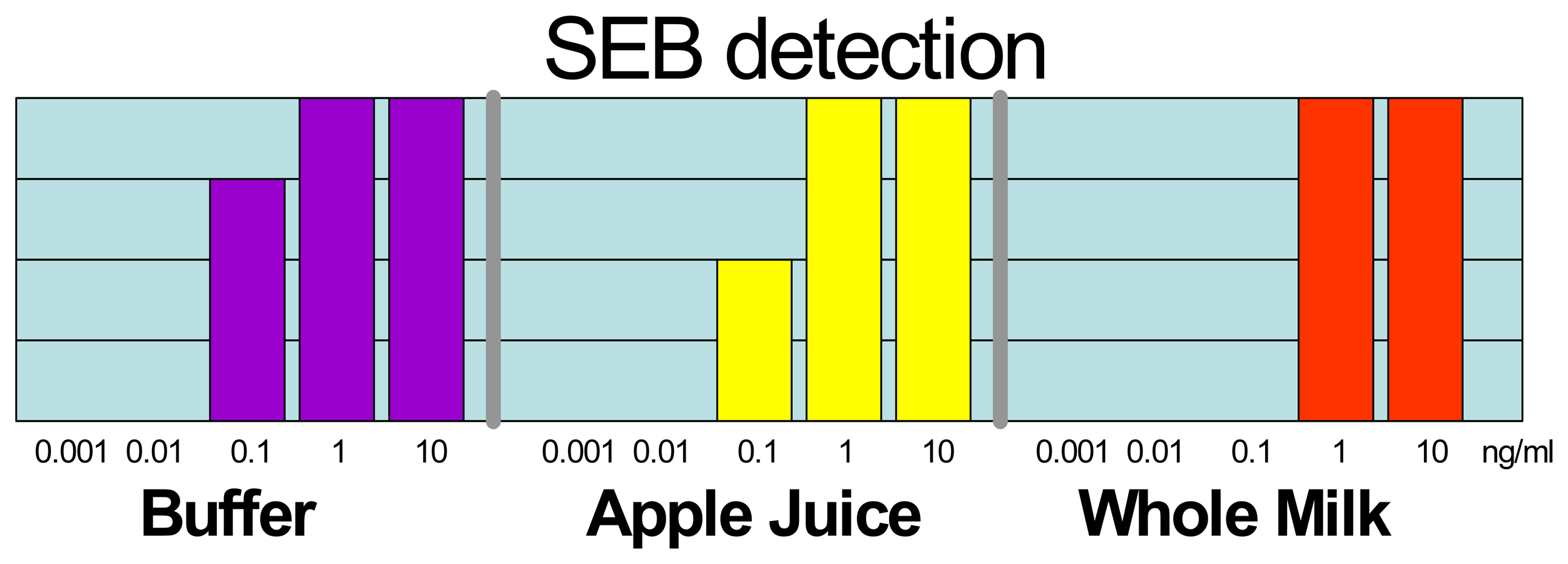
2.1. Large Protein Toxins in Foods
2.2. Mycotoxins in foods and air
2.3 Blind Laboratory Trials
3. Potential clinical applications
4. Expanding the capabilities - Use of alternative recognition species
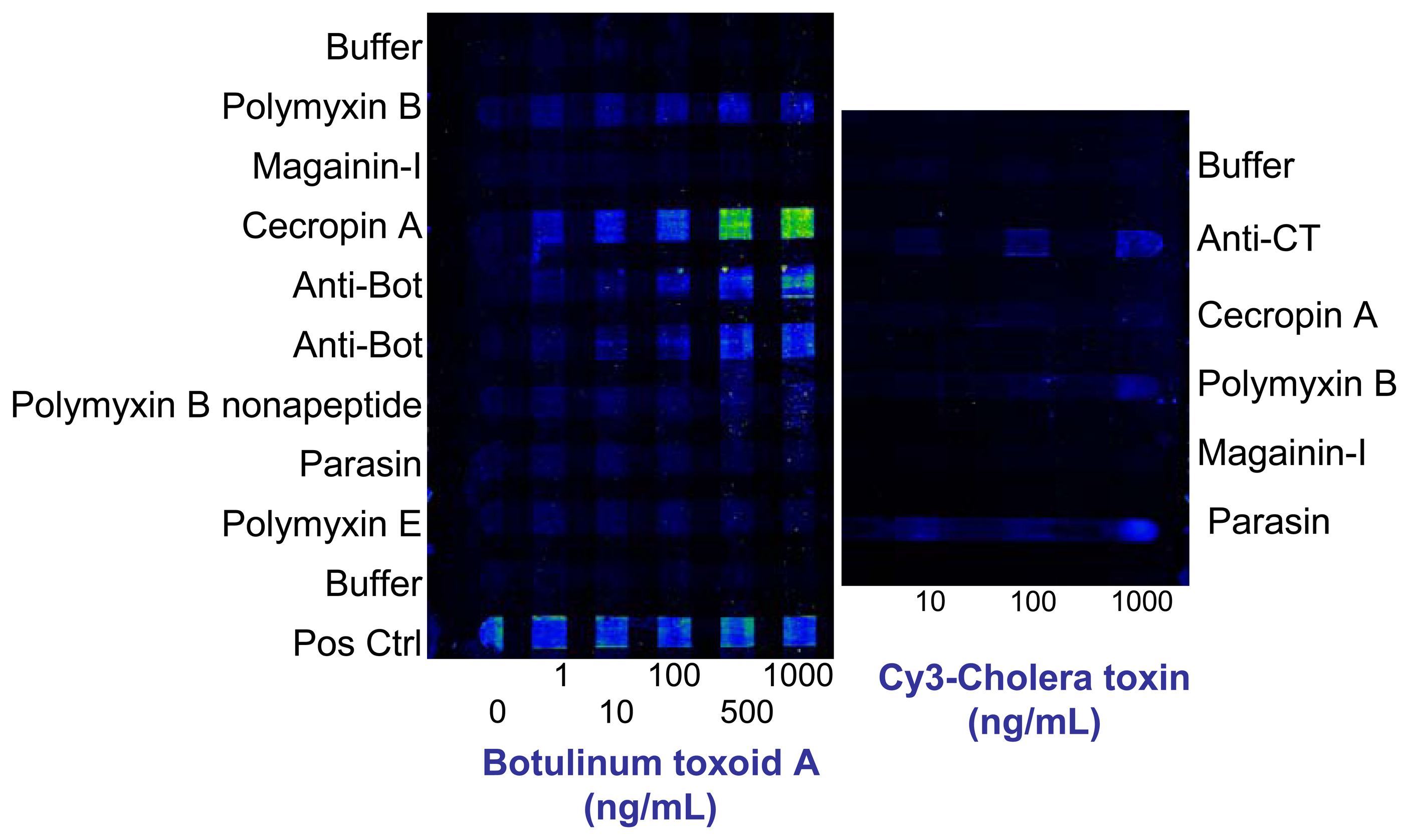
4.1 Use of antimicrobial peptides for toxin detection
4.2. Use of sugars as recognition elements for toxin detection
4.2.1 Immobilization using thiol-terminated linkers
4.2.2 Immobilization using amine-terminated linkers
4.2.3 Immobilization using biotin-terminated linkers and multivalent scaffolds
Conclusion
Acknowledgments
References and Notes
- Ligler, F.S.; Taitt, C.R.; Shriver-Lake, L.C.; Sapsford, K.E.; Shubin, Y.; Golden, J.P. Array biosensor for detection of toxins. Anal. Bioanal. Chem. 2003, 377(3), 469–477. [Google Scholar]
- Feldstein, M.J.; Golden, J.P.; Ligler, F.S.; Rowe, C.A. Reflectively coated optical waveguide and fluidics cell integration. U.S. Pat. 6,192,168, 2001. [Google Scholar]
- Golden, J.P.; Taitt, C.R.; Shriver-Lake, L.C.; Shubin, Y.S.; Ligler, F.S. A portable automated multianalyte biosensor. Talanta 2005, 65(5), 1078–1085. [Google Scholar]
- Ligler, F.S.; Sapsford, K.E.; Golden, J.P.; Shriver-Lake, L.C.; Taitt, C.R.; Dyer, M.A.; Barone, S.; Myatt, C.J. The Array Biosensor: Portable, Automated Systems. Anal. Sci. 2007, 23(1), 5–10. [Google Scholar]
- Sweeney, M. J.; White, S.; Dobson, A. D. W. Mycotoxins in agriculture and food safety. Ir. J. Agr. Food Res. 2000, 39(2), 235–244. [Google Scholar]
- Shriver-Lake, L.C.; Shubin, Y.S.; Ligler, F.S. Detection of Staphylococcal Enterotoxin B in Spiked Food Samples. J. Food Prot. 2003, 66(10), 1851–1856. [Google Scholar]
- Sapsford, K.E.; Taitt, C.R.; Loo, N.; Ligler, F.S. Biosensor detection of botulinum toxoid A and staphylococcal enterotoxin B in food. Appl. Environ. Microbiol. 2005, 71(9), 5590–5592. [Google Scholar]
- Ngundi, M.M.; Shriver-Lake, L.C.; Moore, M.H.; Lassman, M.E.; Ligler, F.S.; Taitt, C.R. Array biosensor for detection of ochratoxin A in cereals and beverages. Anal. Chem. 2005, 77(1), 148–154. [Google Scholar]
- Ngundi, M. M.; Qadri, S.A.; Wallace, E.V.; Moore, M. H.; Lassman, M. E.; Shriver-Lake, L.C.; Ligler, F.S.; Taitt, C.R. Detection of deoxynivalenol in foods and indoor air using an array biosensor. Environ. Sci. Technol. 2006, 40(7), 2352–2356. [Google Scholar]
- Sapsford, K.E.; Taitt, C.R.; Fertig, S.; Moore, M.H.; Lassman, M.E.; Maragos, C.A.; Shriver-Lake, L.C. Indirect competitive immunoassay for detection of aflatoxin B-1 in corn and nut products using the array biosensor. Biosens. Bioelectron. 2006, 21(12), 2298–2305. [Google Scholar]
- Ngundi, M.M.; Shriver-Lake, L.C.; Moore, M.H.; Ligler, F.S.; Taitt, C.R. Multiplexed detection of mycotoxins in foods with a regenerable array. J. Food Prot. 2006, 69(12), 3047–3051. [Google Scholar]
- Ngundi, M.M.; Taitt, C.R. An Array Biosensor for Detection of Bacterial and Toxic Contaminants of Foods. Diagnostic Bacteriology Protocols 2006, 53–68. [Google Scholar]
- Shriver-Lake, L.C.; Erickson, J.S.; Sapsford, K.E.; Ngundi, M.M.; Shaffer, K.M.; Kulagina, N.V.; Hu, J.E.; Gray, S.A.; Golden, J.P.; Ligler, F.S.; Taitt, C.R. Blind laboratory trials for multiple pathogens in spiked food matrices. Anal. Lett. 2007, 40(16-18), 3219–3231. [Google Scholar]
- Golden, J.P.; Shriver-Lake, L.C.; Taitt, C.R.; Fertig, S.; Sapsford, K.E.; Ligler, F.S. A portable array biosensor. In Advanced Environmental, Chemical, and Biological Sensing Technologies II; Philadelphia, PA, USA; SPIE: Philadelphia, PA, USA, 2004; pp. 13–17. [Google Scholar]
- Lin, Y.-Y.; Wang, J.; Liu, G.; Wu, H.; Wai, C.M.; Lin, Y. A nanoparticle label/immunochromatographic electrochemical biosensor for rapid and sensitive detection of prostate-specific antigen. Biosens. Bioelectron. 2008, 23(11), 1659–1665. [Google Scholar]
- Healy, D.A.; Hayes, C.J.; Leonard, P.; McKenna, L.; O'Kennedy, R. Biosensor developments: application to prostate-specific antigen detection. Trends Biotechnol. 2007, 25(3), 125–131. [Google Scholar]
- Wee, K.W.; Kang, G.Y.; Park, J.; Kang, J.Y.; Yoon, D.S.; Park, J.H.; Kim, T.S. Novel electrical detection of label-free disease marker proteins using piezoresistive self-sensing micro-cantilevers. Biosens. Bioelectron. 2005, 20(10), 1932–1938. [Google Scholar]
- Albrecht, C.; Kaeppel, N.; Gauglitz, G. Two immunoassay formats for fully automated CRP detection in human serum. Anal. Bioanal. Chem. 2008, 391(5), 1845–1852. [Google Scholar]
- McBride, J.; Cooper, M. A high sensitivity assay for the inflammatory marker C-Reactive protein employing acoustic biosensing. J. Nanobiotechnology 2008. [Google Scholar] [CrossRef]
- Rowe, C.A.; Bolitho, J.S.; Jane, A.; Bundesen, P.G.; Rylatt, D.B.; Eisenberg, P.R.; Ligler, F.S. Rapid detection of D-dimer using a fiber optic biosensor. Thromb. Haemost. 1998, 79(1), 94–98. [Google Scholar]
- Bergstrand, A.; Ceylan, I.; Quash, G.; Nyden, M.; Holmberg, K. Towards a biosensor immunoassay of protein-bound isopeptides in human plasma. Colloids Surf. B 2008, 66(1), 150–153. [Google Scholar]
- McCauley, T.G.; Hamaguchi, N.; Stanton, M. Aptamer-based biosensor arrays for detection and quantification of biological macromolecules. Anal. Biochem. 2003, 319(2), 244–250. [Google Scholar]
- Rasooly, A.; Jacobson, J. Development of biosensors for cancer clinical testing. Biosens. Bioelectron. 2006, 21(10), 1851–1858. [Google Scholar]
- Joos, T.O.; Fortina, P. Microarrays in Clinical Diagnostics; Human Press: Totowa, New Jersey, 2005. [Google Scholar]
- Lu, H.L.; Goodell, V.; Disis, M.L. Targeting serum antibody for cancer diagnosis: a focus on colorectal cancer. Expert Opin. Ther. Targets 2007, 11(2), 235–244. [Google Scholar]
- Chatterjee, M.; Draghici, S.; Tainsky, M.A. Immunotheranostics: Breaking tolerance in immunotherapy using tumor autoantigens identified on protein microarrays. Curr. Opin. Drug Discovery Dev. 2006, 9(3), 380–385. [Google Scholar]
- Chatterjee, M.; Mohapatra, S.; Ionan, A.; Bawa, G.; Ali-Fehmi, R.; Wang, X.J.; Nowak, J.; Ye, B.; Nahhas, F.A.; Lu, K.; Witkin, S.S.; Fishman, D.; Munkarah, A.; Morris, R.; Levin, N.K.; Shirley, N.N.; Tromp, G.; Abrams, J.; Draghici, S.; Tainsky, M.A. Diagnostic markers of ovarian cancer by high-throughput antigen cloning and detection on arrays. Cancer Res. 2006, 66(2), 1181–1190. [Google Scholar]
- Thaler, M.; Buhl, A.; Welter, H.; Schreiegg, A.; Kehrel, M.; Alber, B.; Metzger, J.; Luppa, P. Biosensor analyses of serum autoantibodies: application to antiphospholipid syndrome and systemic lupus erythematosus. Anal. Bioanal. Chem. 2008, 1618–2642. [Google Scholar]
- Kurowska, E.; Szymiczek, M.; Gorczyca, W.A.; Kuropatwa, M.; Jakubaszko, J.; Marek, J. Detection of serum antibodies to S-antigen by surface plasmon resonance (SPR). J. Immunoassay Immunochem. 2006, 27(4), 331–340. [Google Scholar]
- Kim, J.Y.; Lee, M.H.; Jung, K.I.; Na, H.Y.; Cha, H.S.; Ko, E.M.; Kim, T.J. Detection of antibodies against glucose 6-phosphate isomerase in synovial fluid of rheumatoid arthritis using surface plasmon resonance (BIAcore). Exp. Mol. Med. 2003, 35(4), 310–316. [Google Scholar]
- Heyries, K.A.; Loughran, M.G.; Hoffmann, D.; Homsy, A.; Blum, L.J.; Marquette, C.A. Microfluidic biochip for chemiluminescent detection of allergen-specific antibodies. Biosens. Bioelectron. 2008, 23(12), 1812–1818. [Google Scholar]
- Wilson, K.M.; Di Camillo, C.; Doughty, L.; Dax, E.M. Humoral immune response to primary rubella virus infection. Clin. Vaccine Immunol. 2006, 13(3), 380–386. [Google Scholar]
- Vaisocherová, H.; Mrkvová, K.; Piliarik, M.; Jinoch, P.; Steinbachová, M.; Homola, J. Surface plasmon resonance biosensor for direct detection of antibody against Epstein-Barr virus. Biosens. Bioelectron. 2007, 22(6), 1020–1026. [Google Scholar]
- Aytur, T.; Foley, J.; Anwar, M.; Boser, B.; Harris, E.; Beatty, P.R. A novel magnetic bead bioassay platform using a microchip-based sensor for infectious disease diagnosis. J. Immunol. Meth. 2006, 314(1-2), 21–29. [Google Scholar]
- Jongerius-Gortemaker, B.G.M.; Goverde, R.L.J.; van Knapen, F.; Bergwerff, A.A. Surface plasmon resonance (BIACORE) detection of serum antibodies against Salmonella enteritidis and Salmonella typhimurium. J. Immunol. Meth. 2002, 266(1-2), 33–44. [Google Scholar]
- Brown, D.R.; Wendland, L.D.; Ortiz, G.J.; Kramer, M.F.; Lim, D.V.; Brown, M.B.; Klein, P.A. Evanescent-wave biosensor for field serodiagnosis of tortoise mycoplasmosis. Vet. Immunol. Immunopathol. 2008, 124(3-4), 322–331. [Google Scholar]
- Nagel, T.; Gajovic-Eicheimann, N.; Tobisch, S.; Schulte-Spechtel, U.; Bier, F.F. Serodiagnosis of Lyme borreliosis infection using surface plasmon resonance. Clin. Chim. Acta 2008, 394(1-2), 110–113. [Google Scholar]
- Thanyani, S.T.; Roberts, V.; Siko, D.G.R.; Vrey, P.; Verschoor, J.A. A novel application of affinity biosensor technology to detect antibodies to mycolic acid in tuberculosis patients. J. Immunol. Methods 2008, 332(1-2), 61–72. [Google Scholar]
- Abad, L.W.; Neumann, M.; Tobias, L.; Obenauer-Kutner, L.; Jacobs, S.; Cullen, C. Development of a biosensor-based method for detection and isotyping of antibody responses to adenoviral-based gene therapy vectors. Anal. Biochem. 2002, 310(1), 107–113. [Google Scholar]
- Moreno-Bondi, M.C.; Taitt, C.R.; Shriver-Lake, L.C.; Ligler, F.S. Multiplexed measurement of serum antibodies using an array biosensor. Biosens. Bioelectron. 2006, 21(10), 1880–1886. [Google Scholar]
- Kobayashi, S.; Takeshima, K.; Park, C.B.; Kim, S.C.; Matsuzaki, K. Interactions of the novel antimicrobial peptide buforin 2 with lipid bilayers: Proline as a translocation promoting factor. Biochemistry 2000, 39(29), 8648–8654. [Google Scholar]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of action of the antimicrobial peptide buforin II: Buforin II kills microorganisms by penetrating the cell membrane and inhibiting cellular functions. Biochem. Biophys. Res. Commun. 1998, 244(1), 253–257. [Google Scholar]
- Xiong, Y.Q.; Yeaman, M.R.; Bayer, A.S. In vitro antibacterial activities of platelet microbicidal protein and neutrophil defensin against Staphylococcus aureus are influenced by antibiotics differing in mechanism of action. Antimicrob. Agents Chemother. 1999, 43(5), 1111–1117. [Google Scholar]
- Kulagina, N.V.; Shaffer, K.M.; Ligler, F.S.; Taitt, C.R. Antimicrobial peptides as new recognition molecules for screening challenging species. Sens. Actuat. B-Chem. 2007, 121(1), 150–157. [Google Scholar]
- Kulagina, N.V.; Shaffer, K.M.; Anderson, G.P.; Ligler, F.S.; Taitt, C.R. Antimicrobial peptide-based array for Escherichia coli and Salmonella screening. Anal. Chim. Acta 2006, 575(1), 9–15. [Google Scholar]
- Kulagina, N.V.; Lassman, M.E.; Ligler, F.S.; Taitt, C.R. Antimicrobial peptides for detection of bacteria in biosensor assays. Anal. Chem. 2005, 77(19), 6504–6508. [Google Scholar]
- Garcia, G.E.; Moorad, D.R.; Gordon, R.K. Buforin I, a natural peptide, inhibits botulinum neurotoxin B activity in vitro. J. Appl. Toxicol. 1999, 19, S19–S22. [Google Scholar]
- Garcia, G.E.; Gordon, R.K.; Moorad, D.R.; Doctor, B.P. Novel peptides capable of inhibiting the enzymatic activity of Botulinum toxin B, and Tetanus neurotoxins useful for preventing or treating toxic poisoning such as Botulinum toxin and tetanus poisoning. WO200069895-A. 2004. [Google Scholar]
- Gordon, R.K.; Moorad, D.R.; Doctor, B.P.; Garcia, G.E. Novel previn compounds useful for inhibiting the protease activity of Botulinum B and tetanus toxins. WO200069891-A. 2007. [Google Scholar]
- Kulagina, N.V.; Anderson, G.P.; Ligler, F.S.; Shaffer, K.M.; Taitt, C.R. Antimicrobial peptides: New recognition molecules for detecting botulinum toxins. Sensors 2007, 7(11), 2808–2824. [Google Scholar]
- Angstrom, J.; Teneberg, S.; Karlsson, K.A. Delineation and Comparison of Ganglioside-Binding epitopes for the Toxins of Vibrio cholerae, Escherichia coli, and Clostridium tetani: Evidence for Overlapping Epitopes. Proc. Natl. Acad. Sci. U.S.A. 1994, 91(25), 11859–11863. [Google Scholar]
- Singh, A.K.; Harrison, S.H.; Schoeniger, J.S. Gangliosides as receptors for biological toxins: Development of sensitive fluoroimmunoassays using ganglioside-bearing liposomes. Anal. Chem. 2000, 72(24), 6019–6024. [Google Scholar]
- Kitamura, M.; Iwamori, M.; Nagai, Y. Interaction Between Clostridium botulinum Neurotoxin and Gangliosides. Biochim. Biophys. Acta 1980, 628(3), 328–335. [Google Scholar]
- Turnbull, W.B.; Precious, B.L.; Homans, S.W. Dissecting the cholera toxin-ganglioside GM1 interaction by isothermal titration calorimetry. J. Am. Chem. Soc. 2004, 126(4), 1047–1054. [Google Scholar]
- Ngundi, M.M.; Kulagina, N.V.; Anderson, G.P.; Taitt, C.R. Nonantibody-based recognition: alternative molecules for detection of pathogens. Expert Rev Proteom. 2006, 3(5), 511–524. [Google Scholar]
- Disney, M.D.; Seeberger, P.H. The use of carbohydrate microarrays to study carbohydrate-cell interactions and to detect pathogens. Chem. Biol. 2004, 11(12), 1701–1707. [Google Scholar]
- Fang, Y.; Frutos, A.G.; Lahiri, J. Ganglioside microarrays for toxin detection. Langmuir 2003, 19(5), 1500–1505. [Google Scholar]
- Ahn, S.; Durst, R.A. Detection of cholera toxin in seafood using a ganglioside-liposome immunoassay. Anal. Bioanal. Chem. 2008, 391(2), 473–478. [Google Scholar]
- Serra, B.; Gamella, M.; Reviejo, A.J.; Pingarron, J.M. Lectin-modified piezoelectric biosensors for bacteria recognition and quantification. Anal. Bioanal. Chem. 2008, 391(5), 1853–1860. [Google Scholar]
- Shen, Z.H.; Huang, M.C.; Xiao, C.D.; Zhang, Y.; Zeng, X.Q.; Wang, P.G. Nonlabeled quartz crystal microbalance biosensor for bacterial detection using carbohydrate and lectin recognitions. Anal. Chem. 2007, 79(6), 2312–2319. [Google Scholar]
- Rowe-Taitt, C.A.; Cras, J.J.; Patterson, C.H.; Golden, J.P.; Ligler, F.S. A ganglioside-based assay for cholera toxin using an array biosensor. Anal. Biochem. 2000, 281(1), 123–133. [Google Scholar]
- Ngundi, M.M.; Taitt, C.R.; McMurry, S.A.; Kahne, D.; Ligler, F.S. Detection of bacterial toxins with monosaccharide arrays. Biosens. Bioelectron. 2006, 21(7), 1195–1201. [Google Scholar]
- Ngundi, M.M.; Taitt, C.R.; Ligler, F.S. Simultaneous determination of kinetic parameters for the binding of cholera toxin to immobilized sialic acid and monoclonal antibody using an array biosensor. Biosens. Bioelectron. 2006, 22(1), 124–130. [Google Scholar]
- Ngundi, M.M.; Taitt, C.R.; Ligler, F.S. Crosslinkers modify affinity of immobilized carbohydrates for cholera toxin. Sens. Lett. 2007, 5(3-4), 621–624. [Google Scholar]
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Taitt, C.R.; Shriver-Lake, L.C.; Ngundi, M.M.; Ligler, F.S. Array Biosensor for Toxin Detection: Continued Advances. Sensors 2008, 8, 8361-8377. https://doi.org/10.3390/s8128361
Taitt CR, Shriver-Lake LC, Ngundi MM, Ligler FS. Array Biosensor for Toxin Detection: Continued Advances. Sensors. 2008; 8(12):8361-8377. https://doi.org/10.3390/s8128361
Chicago/Turabian StyleTaitt, Chris Rowe, Lisa C. Shriver-Lake, Miriam M. Ngundi, and Frances S. Ligler. 2008. "Array Biosensor for Toxin Detection: Continued Advances" Sensors 8, no. 12: 8361-8377. https://doi.org/10.3390/s8128361