Selective D3 Receptor Antagonist SB-277011-A Potentiates the Effect of Cocaine on Extracellular Dopamine in the Nucleus Accumbens: a Dual Core-Shell Voltammetry Study in Anesthetized Rats

Abstract
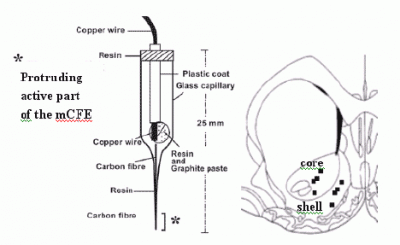
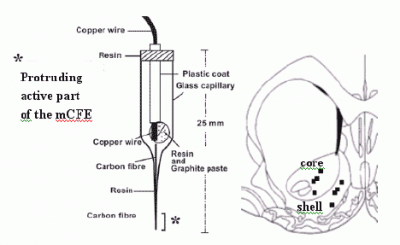
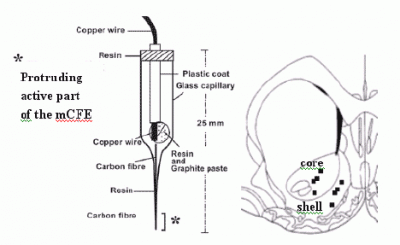
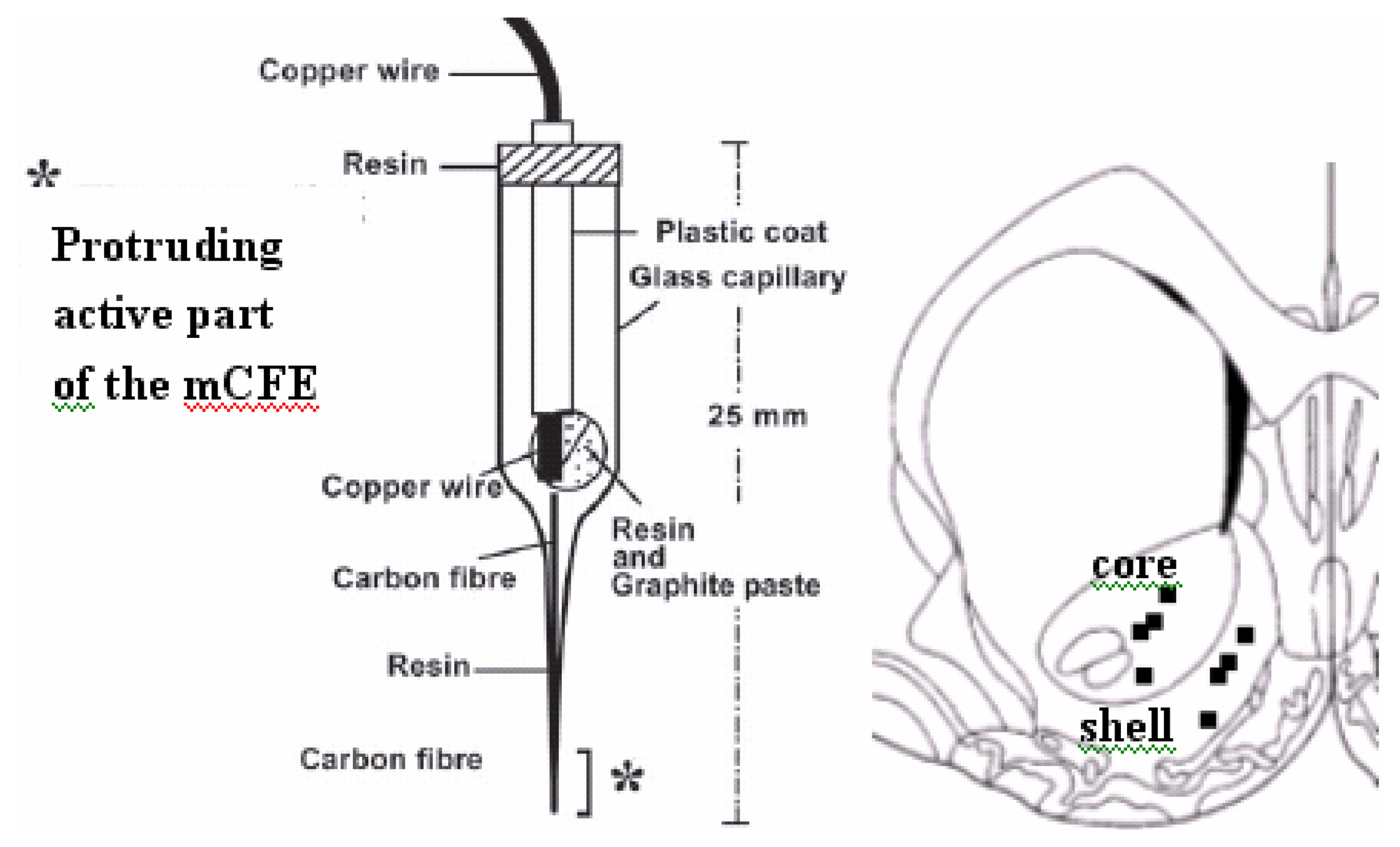
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
1.1. NAc shell and core
1.2. DA receptors in NAc
2. Experimental Section
2.1. Subjects
2.2. Drugs and treatment groups
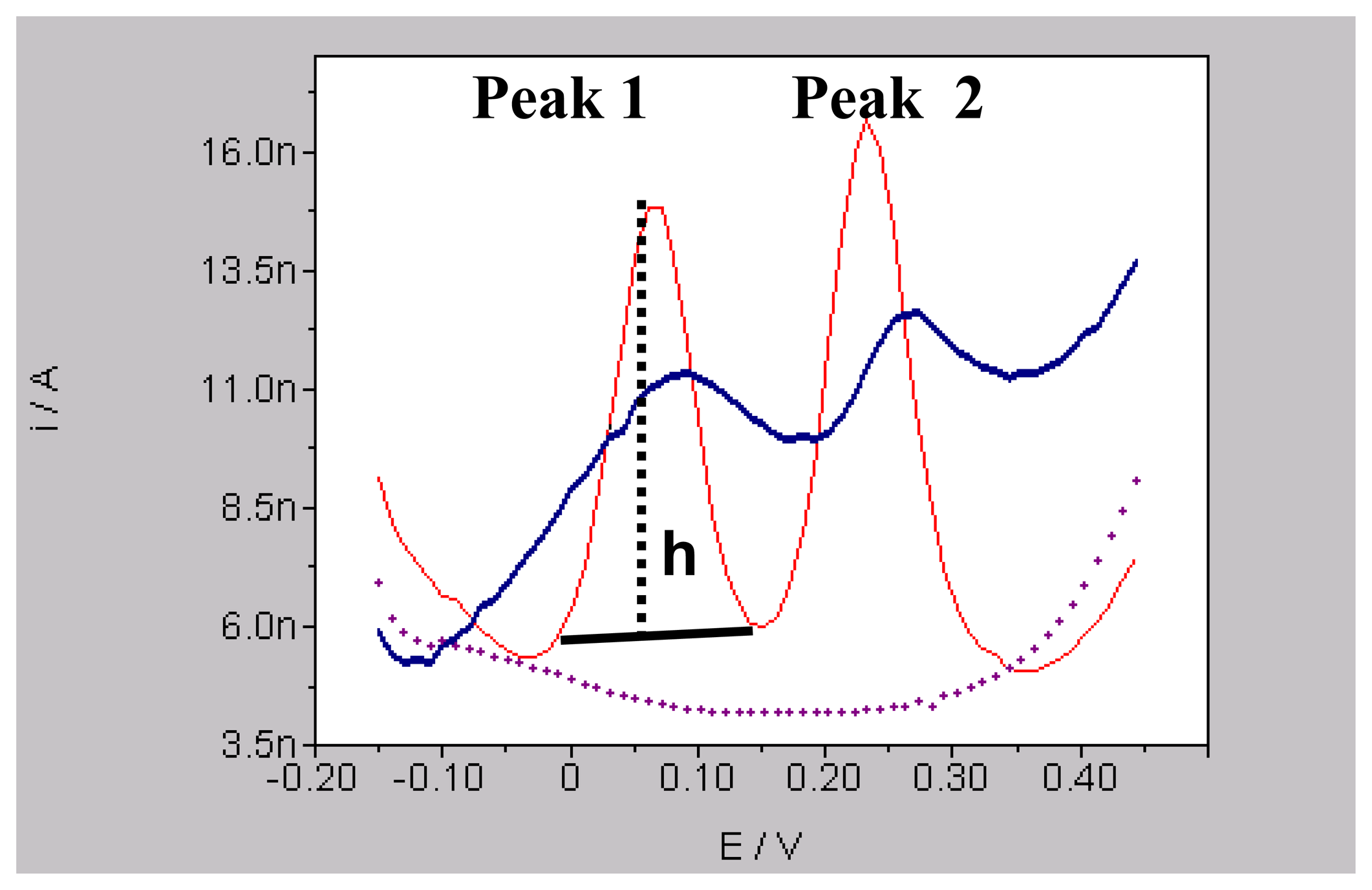
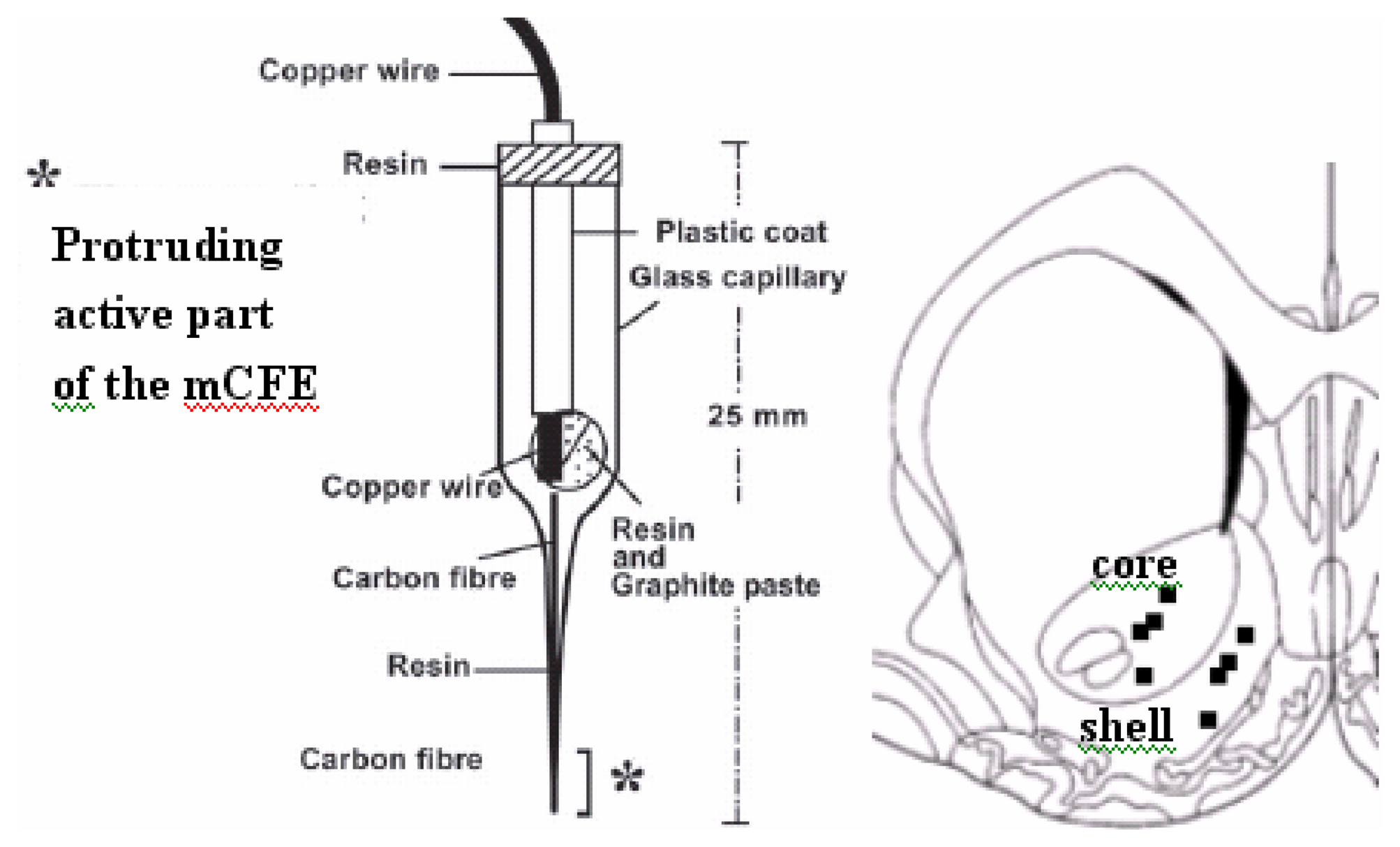
2.3. Voltammetry and carbon fibre microelectrodes (micro-biosensors)
2.4. In vivo voltammetry in dual-probe preparation
2.5 histological analysis of mCFE placement in the NAc
2.6. Data analysis
3. Results and Discussion
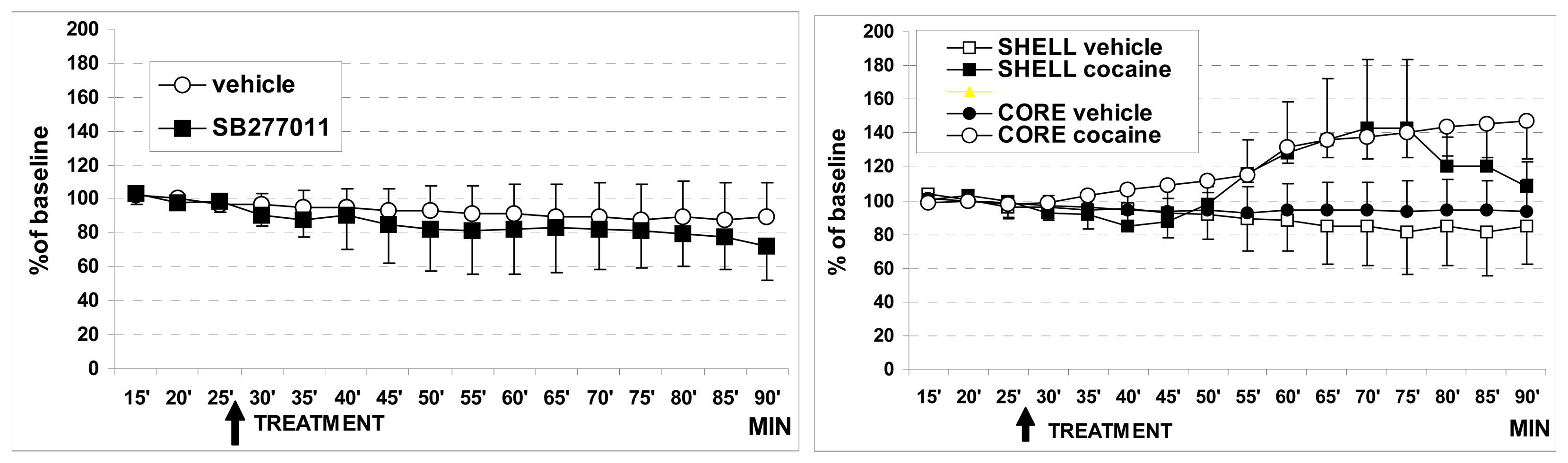
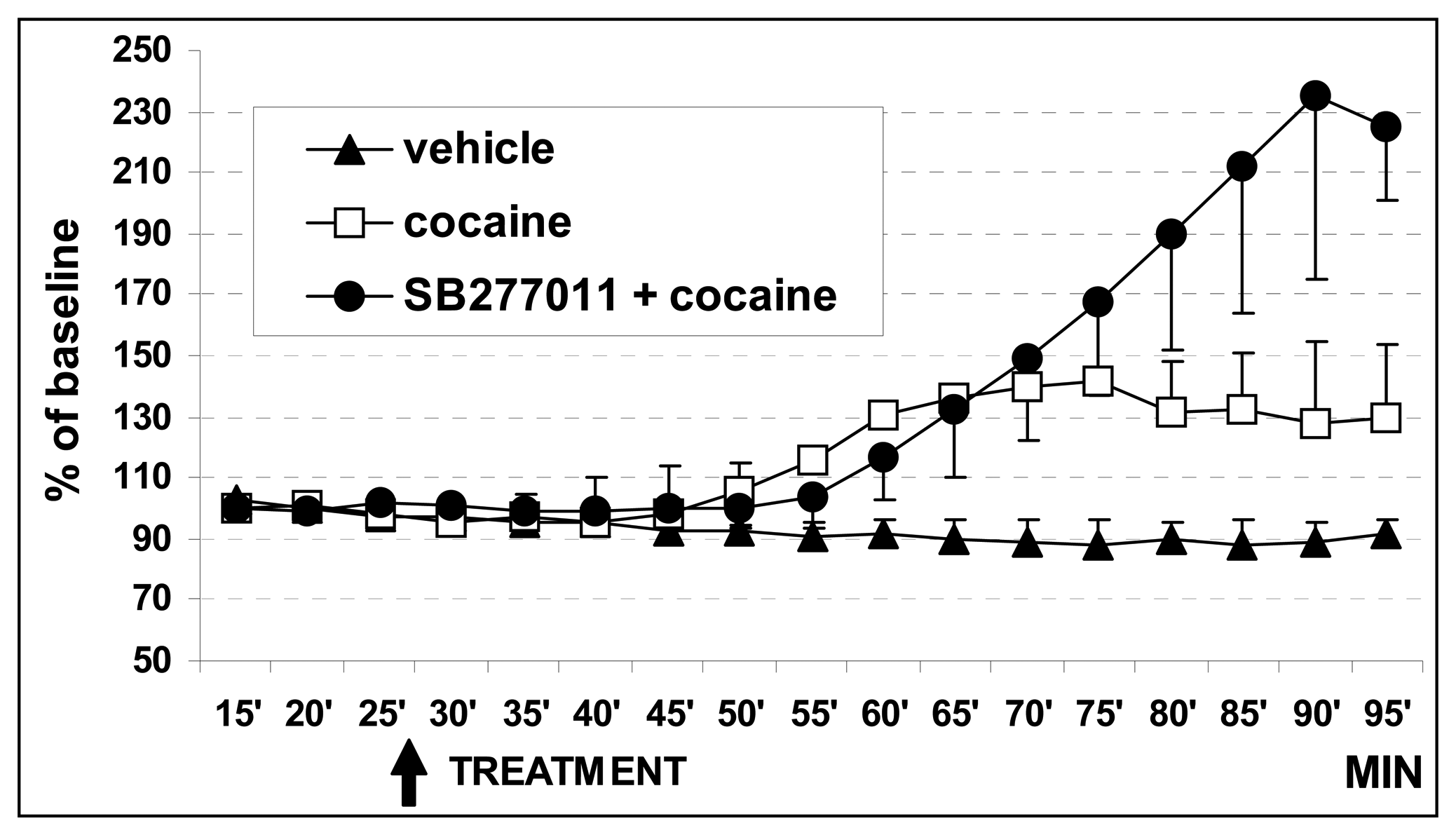
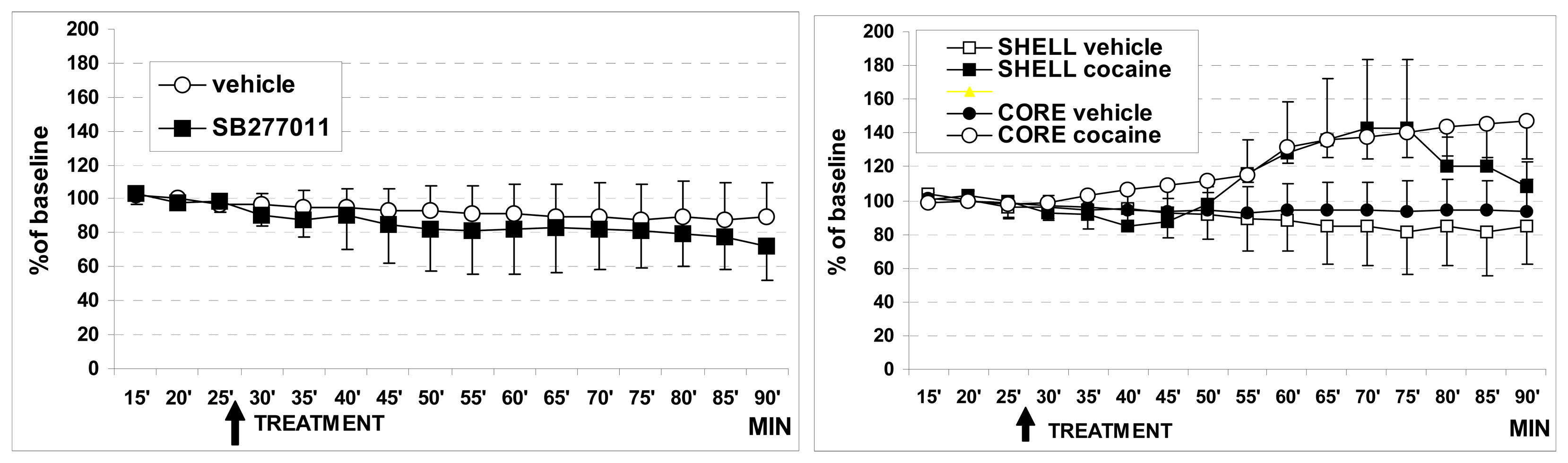
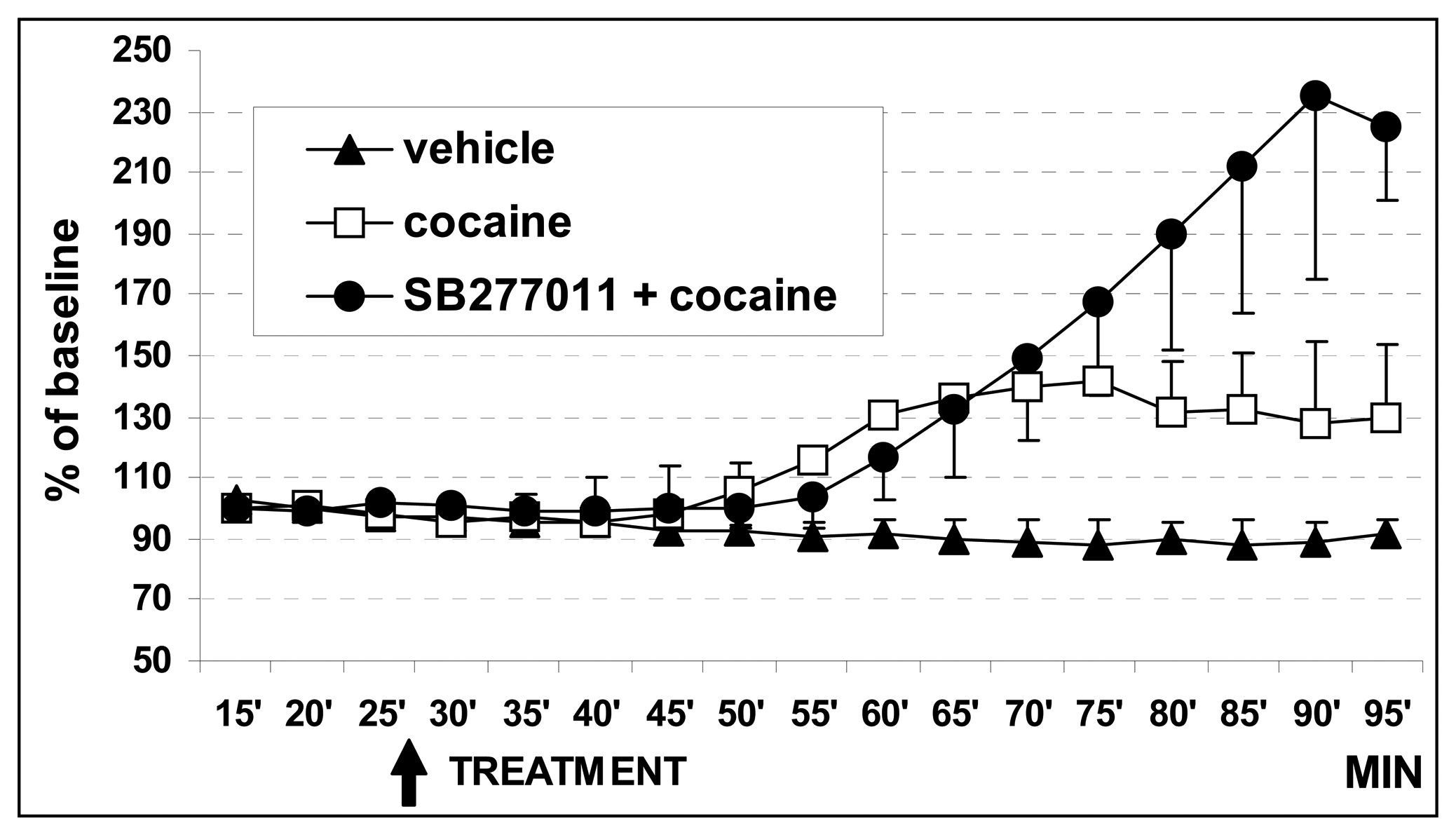
3.1. In vivo dual probe amperometry in core and shell: effect of treatments
3.1.1. Basal DA levels, comparison with the literature
3.1.2. Effect of cocaine on basal DA levels
3.1.3. Effect of SB27701l-A treatment
4. Conclusions
References
- Bozarth, M.A. The mesolimbic dopamine system as a model reward system. In The Mesolimbic Dopamine System: From Motivation to Action; Willner, P., Scheel-Kruger, J., Eds.; Wiley: Chichester, UK, 1991; pp. 301–330. [Google Scholar]
- Wise, R.A. Drug activaction of brain reward pathways. Drug. Alcohol Depend. 1998, 51, 13–22. [Google Scholar]
- Tan, Y.; Brog, J.S.; Williams, E.S.; Zahn, D.S. Morphometric analysis of ventral mesencephalic neurons retrogradely labelled with fluoro-gold following injection in the shell, core and rostral pole of the rat NAccumbens. Brain Res. 1995, 689, 151–156. [Google Scholar]
- Ajheid, G.F.; Heimer, L. New perspectives in basal forebrain organization of special relevance for neuropsychiatric disordera: the striatopallidal, amygdaloid, and corticopetal components of substantia innominata. Neuroscience 1988, 27, 1–39. [Google Scholar]
- Heimer, L.; Zahm, D.S.; Churchill, L.; Kalivas, P.W.; Wohltmann, C. Specificity in the projection patterns of accumbal core and shell in the rat. Neuroscience 1991, 41, 89–125. [Google Scholar]
- Carlezon, W.A., Jr.; Wise, R.A. Rewarding actions of phencyclidine and related drugs in nucleus accumbens shell and frontal cortex. J. Neurosci. 1996, 16, 3112–3122. [Google Scholar]
- Voorn, P.; Gerfen, C.R.; Groenewegen, H.J. Compartmental organization of the ventral striatum of the rat: immunohistochemical distribution of enkephalin, substance P, dopamine, and calciumbinding protein. J. Comp. Neural. 1989, 289, 189–201. [Google Scholar]
- Deutch, A.Y.; Cameron, D.S. Pharmacological characterisation of DA systems in the NAccumbens core and shell. Neuroscience 1992, 46, 49–56. [Google Scholar]
- Deutch, A.Y. Prefrontal cortical dopamine systems and the elaboration of functional corticostriatal clrcults:Implications for schizophrenia and Parkinson's disease. J. Neural. Transm. 1993, 91, 197–221. [Google Scholar]
- Cadoni, C.; Valentini, V.; Di Chiara, G. Behavioral sensitization to Ä9-tetrahydrocannabinol and cross-sensitization with morphine: differential changes in accumbal shell and core dopamine transmission. J. Neurochem. 2008, in press. [Google Scholar]
- Johnson, P.I.; Goodman, J.B.; Condon, R.; Stellar, J.R. Reward shifts and motor responses following microinjections of opiate-specific agonists into either the core or shell of the nucleus accumbens. Psychopharmacology 1995, 120, 195–202. [Google Scholar]
- Essman, W.D.; McGonigle, P.; Lucki, I. Anatomical differentiation within the nucleus accumbens of the locomotor stimulatory actions of selective dopamine agonists and d-amphetamine. Psychopharmacology 1993, 112, 233–241. [Google Scholar]
- Reynolds, S.M.; Berridge, K.C. Emotional environments retune the valence of appetitive versus fearful functions in nucleus accumbens. Nature Neurosci. 2008, 11, 423–425. [Google Scholar]
- Ashby, C.R., Jr.; Paul, M.; Gardner, E.L.; Heidbreder, C.A.; Hagan, J.J. Acute administration of the selective D3 receptor antagonist SB-277011-A blocks the acquisition and expression of the conditioned place preference response to heroin in male rats. Synapse 2003, 48, 154–156. [Google Scholar]
- McKittrick, C.R.; Abercrombie, D. Catecholamine mapping within nucleus accumbens: differences in basal and amphetamine-stimulated efflux of norepinephrine and dopamine in shell and core. J. Neurochem. 2007, 100, 1247–1256. [Google Scholar]
- Rebec, G.V.; Grabner, C.P.; Johnson, M.; Pierce, R.C.; Bardo, M.T. Transient increases in catecholaminergic activity in medial prefrontal cortex and nucleus accumbens shell during novelty. Neuroscience 1997, 76, 707–714. [Google Scholar]
- Jeanblanc, J.; Hoeltzel, A.; Louilot, A. Dissociation in the involvement of dopaminergic neurons innervating the core and shell subregions of the nucleus accumbens in latent inhibition and affective perception. Neuroscience 2002, 111, 315–323. [Google Scholar]
- Heidbreder, C.A.; Gardner, E.L.; Xi, Z.X.; Thanos, P.K.; Mugnaini, M.; Hagan, J.J.; Ashby, C.R., Jr. The Role of Central Dopamine D3 Receptors in Drug Addiction: A Review of Pharmacological Evidence. Brain Res. Brain Res. Rev. 2005, 49, 77–105. [Google Scholar]
- Herroelen, L.; De Backer, J.P.; Wiczak, N.; Flamez, A.; Vauquelin, G.; De Keyser, J. Autoradiographic distribution of D3-type dopamine receptors in human brain using [3H]7-hydroxy-N,N-di-n-propyl-2-aminotretalin. Brain Res. 1994, 648, 222–228. [Google Scholar]
- Suzuki, M.; Hurd, Y.L.; Sokoloff, P.; Schwartz, J.C.; Sedvall, G. D3 dopamine receptor mRNA is widely expressed in the human brain. Brain Res. 1998, 779, 58–74. [Google Scholar]
- Gurevich, E.V.; Joyce, J.N. Distribution of dopamine D3 receptor expressing neurons in the human forebrain: Comparison with D2 receptor expressing neurons. Neuropsychopharmacology 1999, 20, 60–80. [Google Scholar]
- Pilla, M.; Perachon, S.; Garrido, F.; Mann, A.; Wermuth, C.G.; Schwartz, J.C.; Everitt, B.J.; Sokoloff, P. Selective Inhibition of cocaine-seeking behaviour by a partial dopamine D3 receptor agonist. Nature 1999, 400, 1154–1165. [Google Scholar]
- Storch, A.; Schwarz, J. The dopamine transporter: Involvement in selective dopaminergic neurodegeneration. In Neurotoxic factors in Parkinson disease and related disorders; Storch, A., Collins, M.A., Eds.; kluwer academic / plenum publ: NY, USA, 2000; pp. 17–40. [Google Scholar]
- Tamiz, A.P.; Zhang, J.R.; Flippen-Anderson, J.L.; Zhang, M.; Johnson, K.M.; Deschaux, O.; Tella, S.; Kozikowski, A.P. Further SAR studies of piperidine-based analogues of cocaine. 2. Potent dopamine and serotonin reuptake inhibitors. J. Med. Chem. 2000, 43, 1215–1222. [Google Scholar]
- Reavill, C.; Taylor, S.G.; Wood, M.D.; Ashmeade, T.; Austin, N.E.; Jeffrey, P.; Avenell, K.Y.; Boyfield, I.; Hagan, J.J.; Riley, G.J.; Routledge, C.; Wood, M. Pharmacological actions of a novel, high affinity, and selective human dopamine D3 receptor antagonist, Sb277011-a. J Pharmacol. Exp. Ther. 2000, 294, 1154–1165. [Google Scholar]
- Stemp, G.; Ashmeade, T.; Branch, C.L.; Hadley, M.S.; Hunter, A.J.; Johnson, C.N.; Nash, D.J.; Thewlis, K.M.; Vong, A.K.; Austin, N.E.; Jeffrey, P.; Avenell, K.Y.; Boyfield, I.; Hagan, J.J.; Middlemiss, D.N.; Reavill, C.; Riley, G.J.; Routledge, C.; Wood, M. Design and synthesis of trans-N-[4-[2-(6-cyano-1,2,3,4-tetrahydroisoquinolin-2-yl)ethyl]cyclohexyl]-4-quinolinecarbox-amide (SB-277011-A): A potent and selective dopamine D(3) receptor antagonist with high oral bioavailability and CNS penetration in the rat. J. Med. Chem. 2000, 43, 1878–1885. [Google Scholar]
- Cervo, L.; Cocco, A.; Putrella, C.; Heidbreder, C.A. Selective antagonism at dopamine D3 receptors attenuates cocaine-seeking behaviour in the rat. Int. J. Neuropsychopharm. 2007, 10, 167–181. [Google Scholar]
- Xi, Z.X.; Newman, A.H.; Gilbert, J.G.; Pak, A.C.; Peng, X.Q.; Ashby, C.R., Jr.; Gitajn, L.; Gardner, E.L. The novel dopamine D3 receptor antagonist NGB 2904 inhibits cocaine's rewarding effects and cocaine-induced reinstatement of drug-seeking behavior in rats. Neuropsychopharmacology. 2006, 31, 1393–1405. [Google Scholar]
- Vorel, S.R.; Ashby, C.R., Jr.; Paul, M.; Liu, X.; Hayes, R.; Hagan, J.J.; Middlemiss, D.N.; Stemp, G.; Gardner, E.L. Dopamine D3 receptor antagonism inhibits cocaine-seeking and cocaineenhanced brain reward in rats. J. Neurosci. 2002, 22, 9595–9603. [Google Scholar]
- Crespi, F. In vivo voltammetry with micro-biosensors for analysis of neurotransmitter release and metabolism. J.Neurosci. Meth. 1990, 34, 53–65. [Google Scholar]
- Stamford, J.; Crespi, F.; Marsden, C.A. In vivo voltammetric methods for monitoring monoamine release and metabolism. In Practical Approach Series Monitoring Neuronal Activity.; Irl Press at Oxford University Press: Oxford, U.K., 1992; pp. 113–145. [Google Scholar]
- Crespi, F.; Martin, K.F.; Marsden, C.A. Nafion coated carbon fibre electrodes combined with differential pulse voltammetry measure 5HT release in vivo. Neuroscience 1988, 27, 885–896. [Google Scholar]
- Crespi, F.; Mobius, C. In vivo selective monitoring of basal levels of cerebral dopamine using voltammetry with Nafion modified (NA-CRO) carbon fibre micro-electrodes. J.Neurosci. Meth. 1992, 42, 149–161. [Google Scholar]
- Paxinos, G.; Watson, C. The rat brain in stereotaxic coordinates; New York Acad. Press., 1986. [Google Scholar]
- Koob, G.F.; Nestler, E.J. The neurobiology of drug addiction. J. Neuropsychiatry Clin. Neurosci. 1997, 9, 482–497. [Google Scholar]
- Gardner, E.L. What we have learned about addiction from animal models of drug self-administration. Am. J. Addict. 2000, 9, 285–313. [Google Scholar]
- Ashby, C.R., Jr.; Minabe, Y.; Stemp, G.; Hagan, J.J.; Middlemiss, J. Acute and chronic administration of the selective D3 receptor antagonist SB-277011-A alters activity of midbrain dopamine neurons in rats: an in vivo electrophysiological study. Pharmacol. Exp. Ther. 2000, 294, 1166–1174. [Google Scholar]
- Andreoli, M.; Tessari, M.; Pilla, M.; Valerio, E.; Hagan, J.J.; Heidbreder, C.A. Selective antagonism at dopamine D3 receptors prevents nicotine-triggered relapse to nicotine-seeking behaviour. Neuropsychopharmacology 2003, 28, 1272–1280. [Google Scholar]
- Andreoli, M.; Marcon, C.; Hagan, J.J.; Heidbreder, C.A. Effect of selective antagonism at dopamine D3 receptor by SB-277011-A on oral alcohol self-administration in mice. Eur. Neuropsychopharmacology 2003, 13 Suppl. I, S17. [Google Scholar]
- Di Ciano, P.; Underwood, R.J.; Hagan, J.J.; Everitt, B.J. Attenuation of cue-controlled cocaine-seeking by a selective D3 dopamine receptor antagonist SB-277011-A-A. Neuropsychopharmacology 2003, 28, 329–338. [Google Scholar]
- Garris, P.A.; Ciolkowski, E.L.; Pastore, P.; Wightman, R.M. Efflux of dopamine from the synaptic cleft in the nucleus accumbens of the rat brain. J. Neurosci. 1994, 14, 6084–6093. [Google Scholar]
- O'Donnell, P.; Grace, A.A. Dopaminergic modulation of dye coupling between neurons in the core and shell regions of the nucleus accumbens. J. Neurosci. 1993, 13, 3456–3471. [Google Scholar]
- Cools, A.R.; Debeltran, K.K.; Prinssen, E.P.M.; Koshikawa, N. Differential role of core and shell of the nucleus accumbens in jaw movements of rats. Neurosci. Res. Commun. 1993, 13, 55–61. [Google Scholar]
- Prinssen, E.P.M.; Balestra, W.; Bemelmans, F.F.J.; Cools, A.R. Evidence for a role of the shell of the nucleus accumbens in oral behavior of freely moving rats. J. Neurosci. 1994, 14, 1555–1562. [Google Scholar]
- Kalivas, P.W.; Duffy, P. Selective activation of dopamine transmission in the shell of the nucleus accumbens by stress. Brain Res. 1995, 675, 325–328. [Google Scholar]
- Pontieri, F.E.; Tanda, G.; Di Chiara, G. Intravenous cocaine, morphine, and amphetamine preferentially increase extracellular dopamine in the “shell” as compared with the “core” of the rat nucleus accumbens. Proc. Nat. Acad. Sci. USA. 1995, 92, 12304–12308. [Google Scholar]
- Sokolowski, J.D.; Conlan, A.N.; Salamone, J.D. A microdialysis study of nucleus accumbens core and shell dopamine during operant responding in the rat. Neuroscience 1998, 86, 21–28. [Google Scholar]
- Cadoni, C.; Solinas, M.; Di Chiara, G. Psychostimulant sensitization: differential changes in accumbal shell and core dopamine. EUI: J: Pharmacol 2000, 388, 130–140. [Google Scholar]
- Pierce, R.C.; Kalivas, P.W. Amphetamine produces sensitized increases in locomotion and extracellular dopamine preferentiaJly in the nucleus accumbens shell of rats administered repeated cocaine. J. Pharmacol. Exp. Ther. 1995, 275, 1019–1029. [Google Scholar]
- King, D.; Zigmond, M.J.; Finlay, J.M. Effects of dopamine depletion in the medial prefrontal cortex on the stress.induced increase in extracellular dopamine in the nucleus accumbens core and shell. Neuroscience 1997, 77, 141–153. [Google Scholar]
- Barrot, M.; Marinelli, M.; Abrous, D.N.; Rouge-Pont, F.; Le Moal, M.; Piazza, P.V. The dopaminergic hyper-responsiveness of the shell of the nucleus accumbens is hormone-dependent. Eul: J: Neurosci. 2000, 12, 973–979. [Google Scholar]
- Frank, S.; Krum, B.; Spanagel, R. Cocaine-Induced Dopamine Overflow Within the Nucleus Accumbens Measured by In Vivo Microdialysis: A Meta-.Analysis. Synapse 2008, 62, 243–252. [Google Scholar]
- Jones, S.R.; O'Dell, S.J.; Marshall, J.F.; Wightman, R.M. Functional and Anatomical Evidence for Different Dopamine Dynamics in the Core and Shell of the Nucleus Accumbens in Slices of Rat Brain. Synapse 1996, 23, 224–231. [Google Scholar]
- Borland, L.M.; Shi, G.; Yang, H.; Michael, A.C. Voltammetric study of extracellular dopamine near microdialysis probes acutely implanted in the striatum of the anesthetized rat. J. Neurosci. Meth. 2005, 146, 149–158. [Google Scholar]
- Diciano, P.; Coury, A.; Depoortere, R.Y.; Egilmez, Y.; Lane, J.D.; Emmettoglesby, M.W.; Lepiane, F.G.; Phillips, A.G.; Blaha, C.D. Comparison of changes in extracellular dopamine concentrations in the nucleus accumbens during intravenous self-administration of cocaine or d-amphetamine. Behaviour. Pharmacol. 1995, 6, 311–322. [Google Scholar]
- Nomikos, G.G.; Damsma, G.; Wenkstern, D.; Fibiger, H.C. In vivo characterization of locally applied dopamine uptake inhibitors by striatal microdialysis. Synapse. 1990, 6, 106–12. [Google Scholar]
- Allin, R.; Russell, V.A.; Lamm, M.C.; Taljaard, J.J. Regional distribution of monoamines in the nucleus accumbens of the rat. Neurochem. Res. 1988, 13, 937–42. [Google Scholar]
- Dworkin, S.I.; Co, C.; Smith, J. E. Rat brain neurotransmitter turnover rates altered during withdrawal from chronic cocaine administration. Brain Res. 1995, 682, 116–126. [Google Scholar]
- Gurevich, E.V.; Joyce, J.N. Distribution of dopamine D3 receptor expressing neurons in the human orebrain: comparison with D2 receptor expressing neurons. Neuropsychopharmacology 1999, 20, 60–80. [Google Scholar]
- Landwehrmeyer, B.; Mengod, G.; Palacios, J.M. Dopamine D3 receptor mRNA and binding sites in human brain. Brain Res. Mol. Brain Res. 1993, 18, 187–192. [Google Scholar]
- Herroelen, L.; De Backer, J.P.; Wilczak, N.; Flamez, A.; Vauquelin, G.; De Keyser, J. Autoradiographic distribution of D3-type dopamine receptors in human brain using [3H]7-hydroxy-N,N-di-n-propyl-2-aminotetralin. Brain Res. 1994, 648, 222–228. [Google Scholar]
- Parsons, L.H.; Caine, SB; Sokoloff, P.; Schwartz, J.C.; Koob, G.F.; Weiss, F. Neurochemical evidence that postsynaptic nucleus accumbens D3 receptor stimulation enhances cocaine reinforcement. J. Neurochem. 1996, 67, 1078–1089. [Google Scholar]
- Bickel, W.K.; Marsch, L.A.; Carroll, M.E. Deconstructing relative reinforcing efficacy and situating the measures of pharmacological reinforcement with behavioral economics: a theoretical proposal. Psychopharmacology (Berl) 2000, 153, 44–56. [Google Scholar]
- Gilbert, J.; Xi, Z.X.; Campos, A.; Ashby, C.R.; Heidbreder, C.A.; Gardner, E.L. The dopamine D3 receptor antagonist SB27701lA inhibits cocaine reinforcement under fixed-ratio and progressive-ratio schedules. Soc. Neurosci. Abstr. 2003, 422, 10. [Google Scholar]
- Schwarz, A.J.; Gozzi, A.; Reese, T.; Heidbreder, C.A.; Bifone, A. Pharmacological modulation of functional connectivity: the correlation structure underlying the phMRI response to d-amphetamine modified by selective dopamine D3 receptor antagonist SB277011A. Magn. Reson. Imaging 2007, 25, 811–820. [Google Scholar]
- Zocchi, A.; Varnier, G.; Biscaro, B.; Heidbreder, C.A. SB-277011-A, a potent and selective dopamine D3 receptor antagonist, potentiates cocaine-induced increase in dopamine levels in the shell of the nucleus accumbens. 6th Neurochemistry Winter Conference, Solden, Austria; 2004. [Google Scholar]
- Choi, J.; Chen, Y.; Hamel, E.; Jenkins, B.G. Coupling of hemodynamic changes induced by dopamine drugs with dopamine receptor distribution on the cerebral microvasculature. Book of abstracts: 11th Annual Meeting of the International Society of Magnetic Resonance in Medicine' 2003, 356. [Google Scholar]
- Schwarz, A.J.; Gozzi, A.; Reese, T.; Bertani, S.; Crestan, V.; Hagan, J.; Heidbreder, C.A.; Bifone, A. Selective dopamine D3 receptor antagonist SB277011A potentiates phMRI response to acute amphetamine challenge in rat brain. Synapse 2004, 54, 1–10. [Google Scholar]
- Joseph, J.D.; Wang, Y.M.; Miles, P.R.; Budygin, E.A.; Picetti, R.; Gainetdinov, R.R.; Caron, M.G.; Wightman, R.M. Dopamine autoreceptor regulation of release and uptake in mouse brain slices in the absence of D(3) receptors. Neuroscience 2002, 112, 39–49. [Google Scholar]
- Le Foll, B.; Goldberg, S.R.; Sokoloff, P. The dopamine D3 receptor and drug dependence: Effects on reward or beyond? Neuropharmacology 2005, 49, 525–541. [Google Scholar]
- Martinez, D.; Gil, R.; Slifstein, M.; Hwang, D.R.; Huang, Y.; Perez, A. Alcohol dependence is associated with blunted dopamine transmission in the ventral striatum. Biol. Psychiatry 2005, 58, 779–786. [Google Scholar]

© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the CreativeCommons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Congestri, F.; Formenti, F.; Sonntag, V.; Hdou, G.; Crespi, F. Selective D3 Receptor Antagonist SB-277011-A Potentiates the Effect of Cocaine on Extracellular Dopamine in the Nucleus Accumbens: a Dual Core-Shell Voltammetry Study in Anesthetized Rats. Sensors 2008, 8, 6936-6951. https://doi.org/10.3390/s8116936
Congestri F, Formenti F, Sonntag V, Hdou G, Crespi F. Selective D3 Receptor Antagonist SB-277011-A Potentiates the Effect of Cocaine on Extracellular Dopamine in the Nucleus Accumbens: a Dual Core-Shell Voltammetry Study in Anesthetized Rats. Sensors. 2008; 8(11):6936-6951. https://doi.org/10.3390/s8116936
Chicago/Turabian StyleCongestri, Francesco, Francesca Formenti, Viviana Sonntag, Gael Hdou, and Francesco Crespi. 2008. "Selective D3 Receptor Antagonist SB-277011-A Potentiates the Effect of Cocaine on Extracellular Dopamine in the Nucleus Accumbens: a Dual Core-Shell Voltammetry Study in Anesthetized Rats" Sensors 8, no. 11: 6936-6951. https://doi.org/10.3390/s8116936