Real-time Monitoring of Non-specific Toxicity Using a Saccharomyces cerevisiae Reporter System

Abstract
:
1. Introduction
2. Results and Discussion
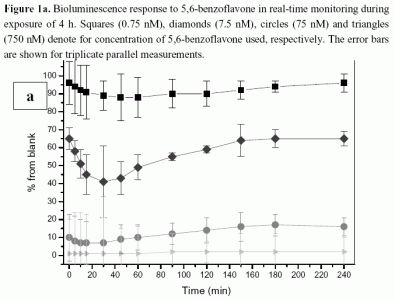
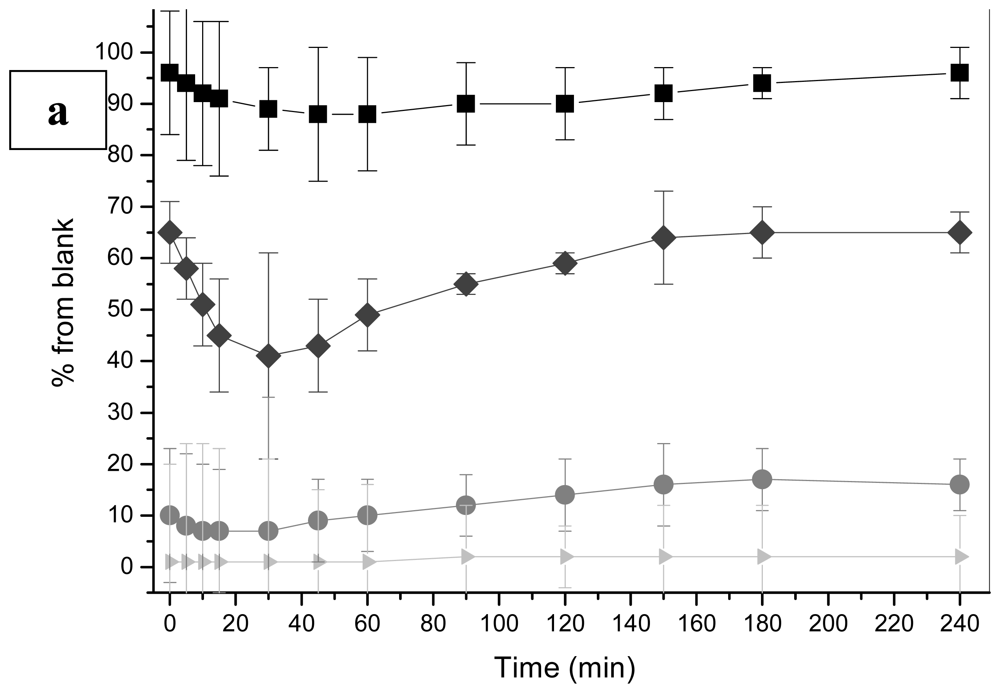
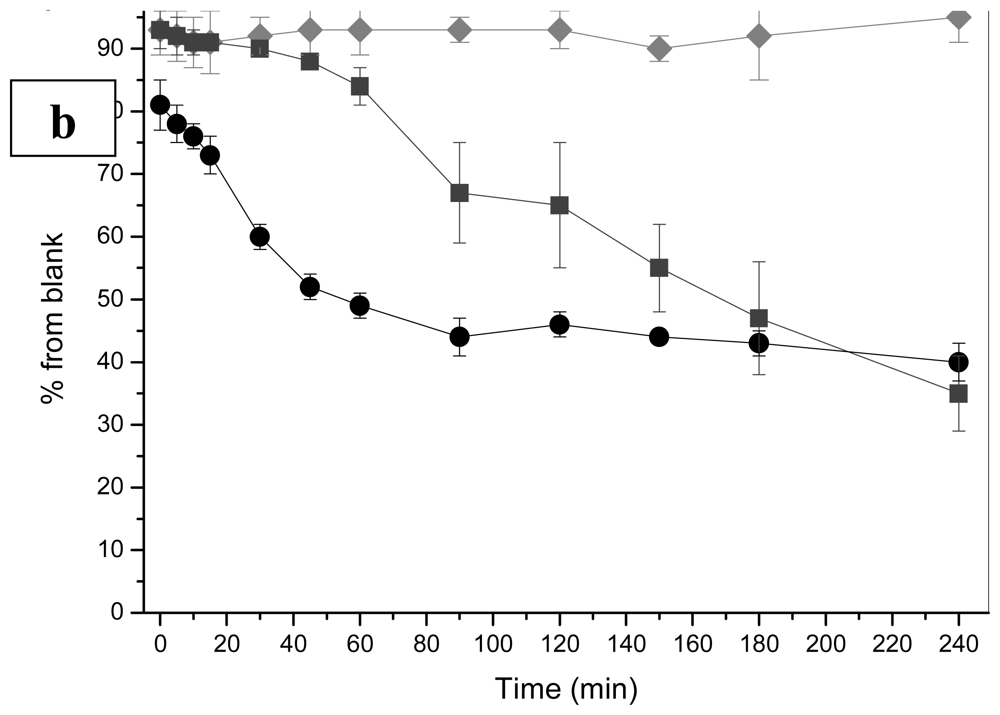
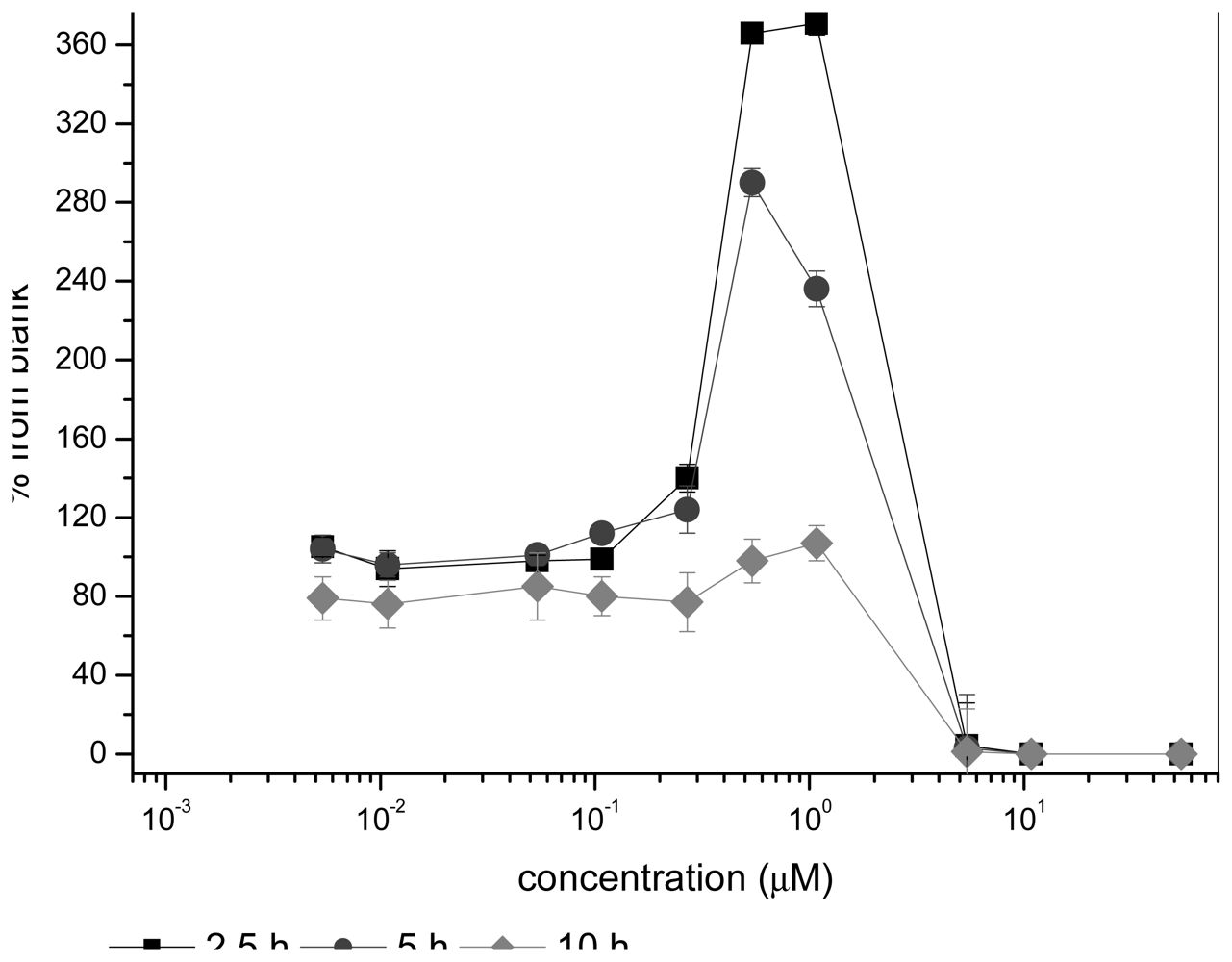
2.1 Bioluminescence assay
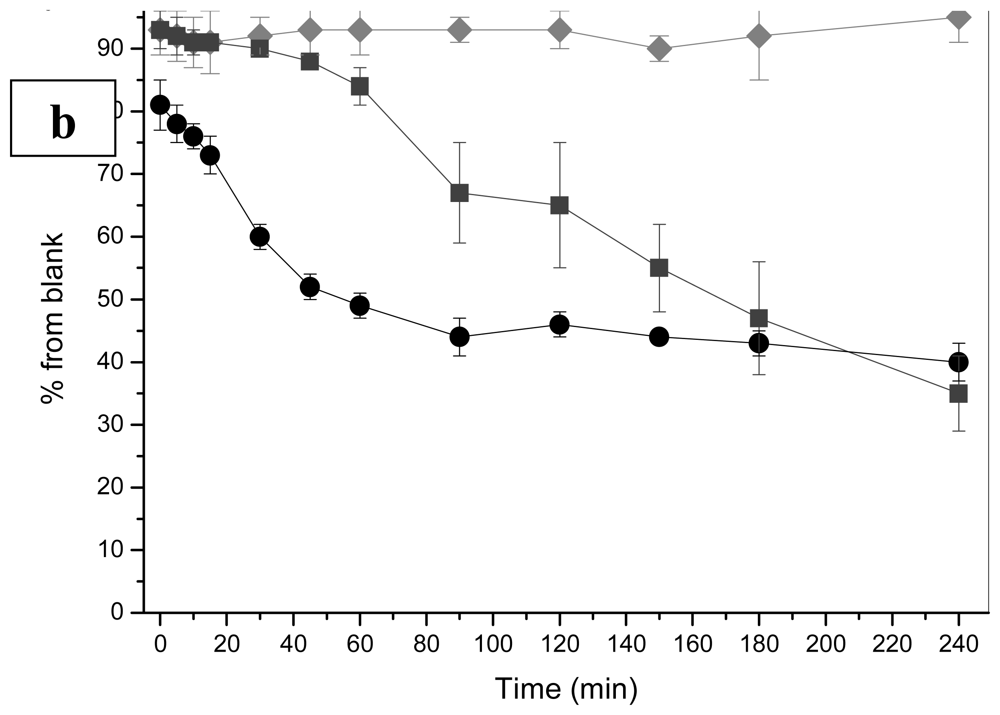
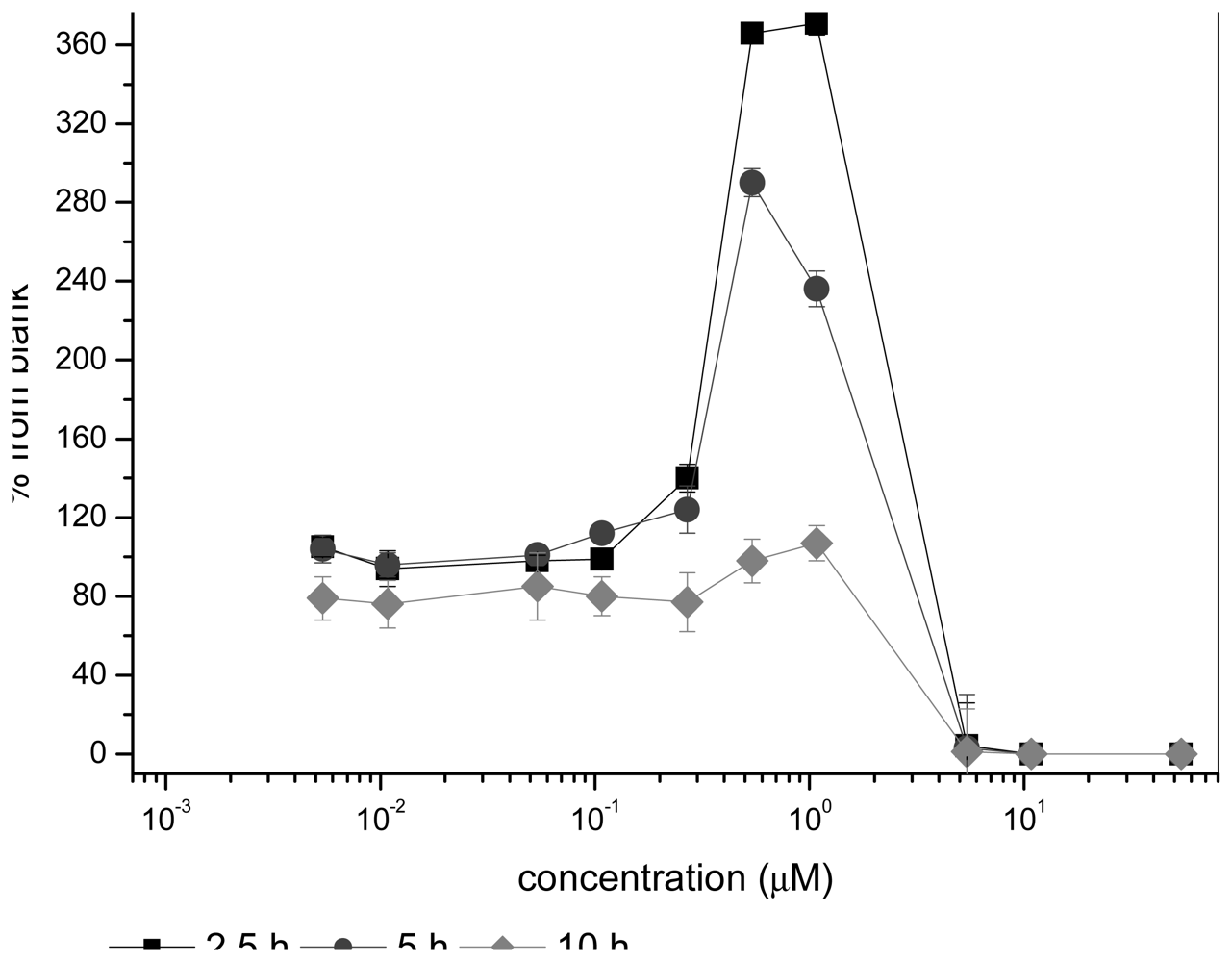
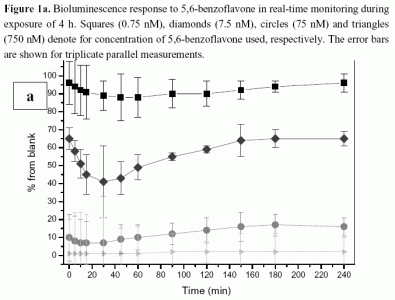
2.1.1 Antimicrobial agents
2.1.2 Heavy metals
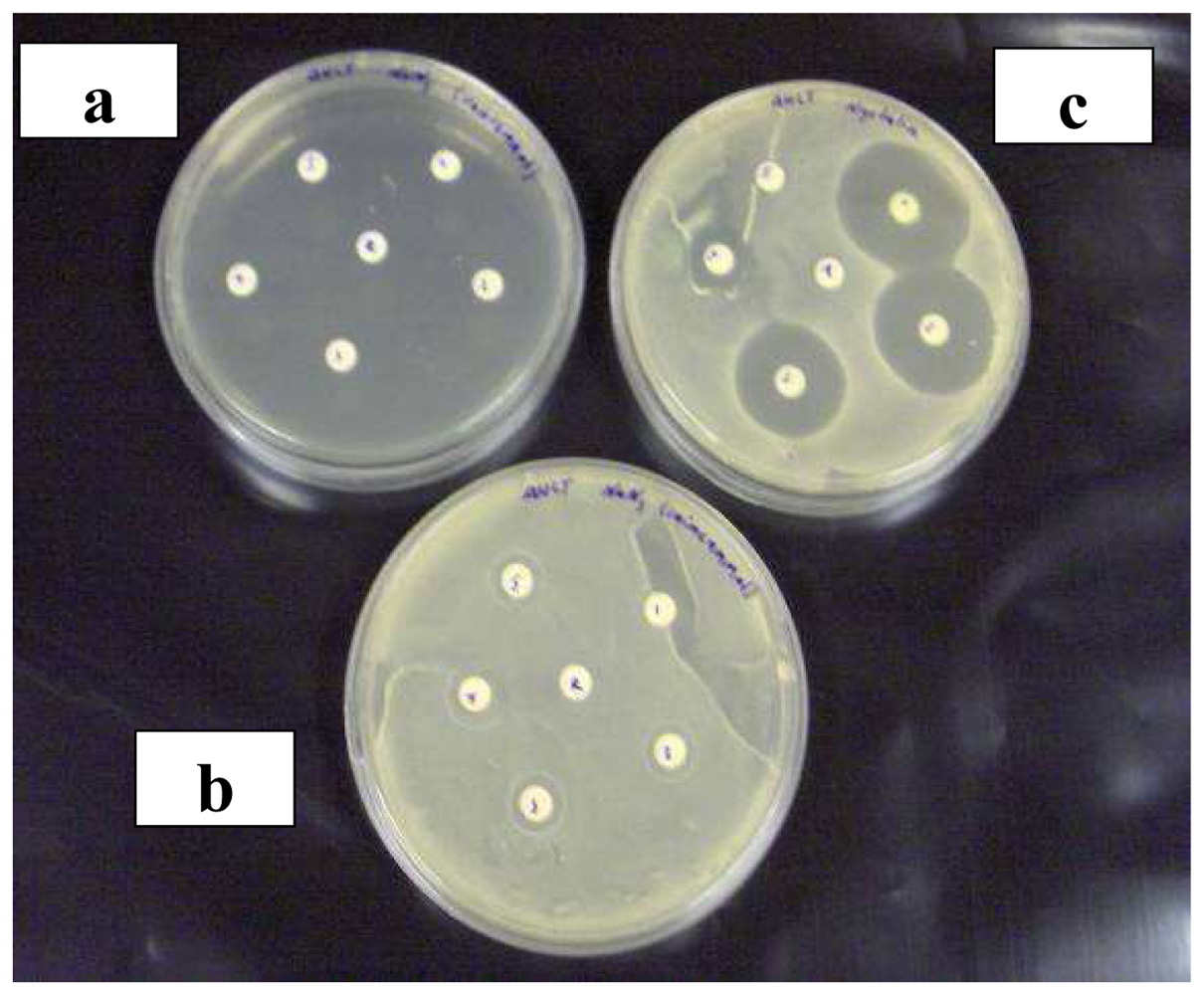
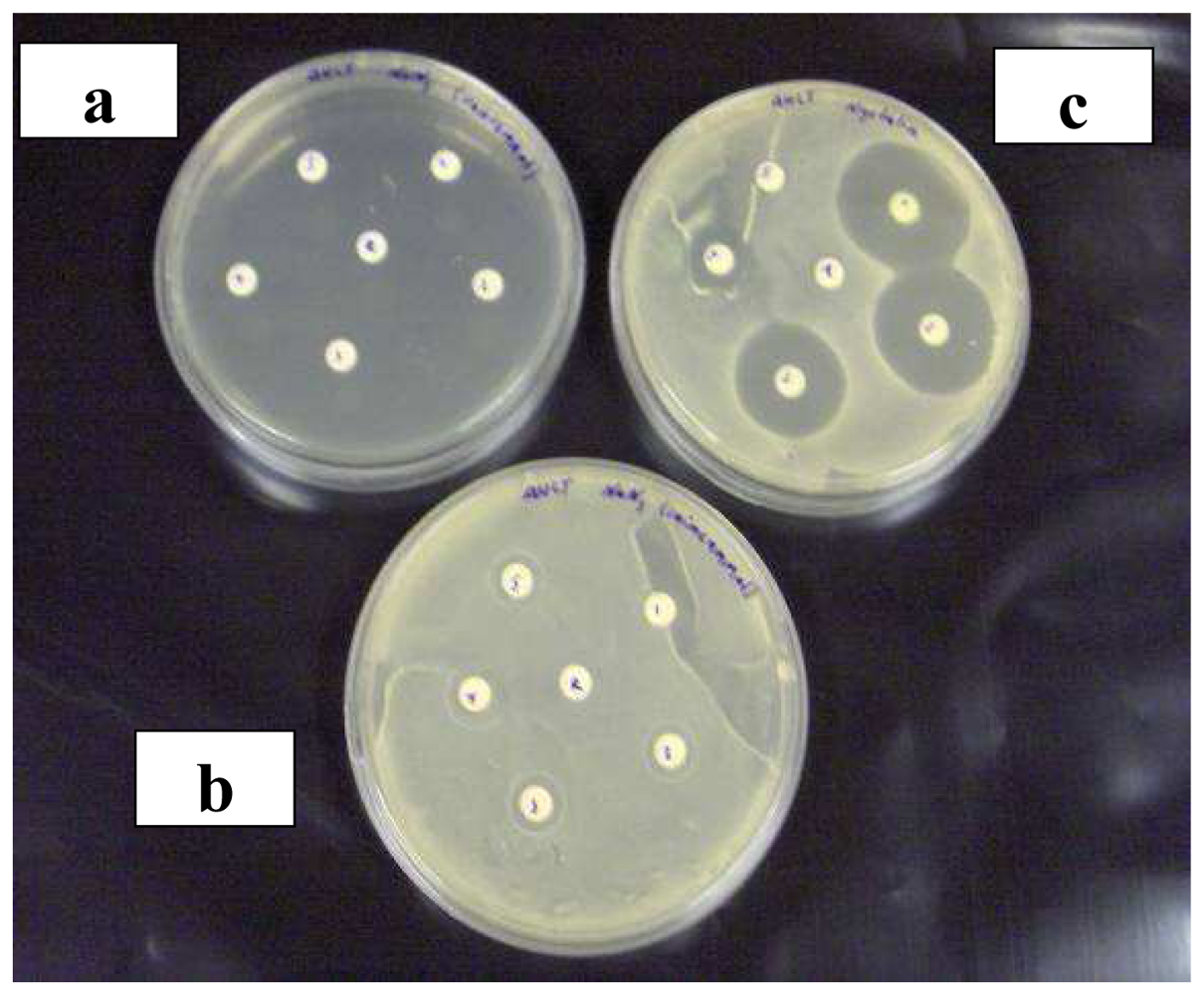
2.2 Agar diffusion assay compared to bioluminescence assay
3. Experimental Section
3.1 Chemicals
3.2 Sample preparation
3.3 Yeast strain cultivation
3.4 Assay procedures
3.4.1 Bioluminescence assay
3.4.2 Agar diffusion assay
4. Conclusions
Acknowledgments
References and Notes
- Bitton, G. Bacterial and biochemical tests for assessing chemical toxicity in the aquatic environment: A review. CRC Crit. Rev. Environ Control 1983, 13, 51–67. [Google Scholar]
- Nascarella, M.A.; Stoffolano, J.G., Jr.; Stanek, E.J., III; Kostecki, P.T.; Calabrese, E.J. Hormesis and stage specific toxicity induced by cadmium in an insect model, the queen blowfly, Phormia regina Meig. Environ. Pollut. 2003, 124, 257–262. [Google Scholar]
- Pichardo, S.; Jos, A.; Zurita, J.L.; Salguero, M.; Cameán, A.M.; Repetto, G. Acute and subacute toxic effects produced by microcystin-YR on the fish cell lines RTG-2 and PLHC-1. Toxicol. in Vitro 2007, 21, 1460–1467. [Google Scholar]
- Zurita, J.L.; Jos, A.; del Peso, A.; Salguero, M.; Cameán, A.M.; López-Artíguez; Repetto, G. Toxicological assessment of indium nitrate on aquatic organisms and investigation of the effects on the PLHC-1 fish cell line. Sci. Total Environ. 2007, 387, 155–165. [Google Scholar]
- Cheung, R.Y.; Rauth, A.M.; Ronaldson, P.T.; Bendayan, R.; Wu, X.Y. In vitro toxicity to breast cancer cells of microsphere-delivered mitomycin C and its combination with doxorubicin. Eur. J. Pharmaceut. Biopharmaceut. 2006, 62, 321–331. [Google Scholar]
- Codina, J.C.; Perez-Garcia, A.; Romero, P.; de Vicente, A.A. Comparison of microbial bioassays for the detection of metal toxicity. Arch. Environ. Contam. Toxicol. 1993, 25, 250–254. [Google Scholar]
- Layton, A.C.; Sanseverino, J.; Gregory, B.W.; Easter, J.P.; Sayler, G.S.; Schultz, T.W. In Vitro Estrogen Reseptor Binding of PCBs: Measured activity and detection of hydroxylated metabolites in a recombinant yeast assay. Toxicol. Appl. Pharmacol. 2002, 180, 157–163. [Google Scholar]
- Henry, S.A.; Patton-Vogt, J.L. Genetic regulation of phospholipid metabolism: yeast as a model eukaryote. Prog. Nucleic Acid Res. Mol. Biol. 1998, 61, 133–79. [Google Scholar]
- Amberg, D.C.; Burke, D.J.; Strathern, J.N. Methods in yeast genetics. A Cold Spring Harbor Laboratory Course Manual; Cold Spring Harbor Laboratory Press: NY, UK, 2005. [Google Scholar]
- Tutulan-Cunita, A.C.; Mikoshki, M.; Mizunuma, M.; Hirata, D.; Miyakawa, T. Mutational analysis of the multidrug resistance ABC transporter Pdr5p with altered drug specificity. Genes Cells 2005, 10, 409–420. [Google Scholar]
- Campanella, L.; Faverso, G.; Mastrofini, D.; Tomassetti, M. Toxicity order of cholanic acids using an immobilised cell biosensor. J. Pharm. Biomed. Anal. 1996, 14, 1007–1013. [Google Scholar]
- Haubenstricker, M.E.; Meier, P.G.; Mancy, K.H.; Brabec, M.J. Rapid toxicity testing based on yeast respiratory activity. Bull. Environ. Contam. Toxicol. 1990, 44, 669–674. [Google Scholar]
- Henderson, G. A comparison of effects of chromate, molybdate and cadmium oxide on respiration in the yeast Saccharomyces cerevisiae. Biol. Metals 1989, 2, 83–88. [Google Scholar]
- Ikebukuro, K.; Miyata, A.; Cho, S.J.; Nomura, Y.; Chang, S.M.; Yamauchi, Y.; Hasebe, Y.; Uchiyama, S.; Karube, I. Microbial cyanide sensor for monitoring river water. J. Biotechnol. 1996, 48, 73–80. [Google Scholar]
- Uribe, S.; Rangel, P.; Espínola, G.; Aguirre, G. Effects of cyclohexane, an industrial solvent, on the yeast Saccharomyces cerevisiae and on isolated yeast mitochondria. Appl. Environ. Mirobiol. 1990, 56, 2114–2119. [Google Scholar]
- Toussaint, M.; Levasseur, G.; Gervais-Bird, J.; Wellinger, R.J.; Elela, S.A.; Conconi, A. A high-throughput method to measure the sensitivity of yeast cells to genotoxic agents in liquid cultures. Mutat. Res. 2006, 606, 92–105. [Google Scholar]
- Schmitt, M.; Gellert, G.; Ludwig, J.; Lichtenberg-Fraté, H. Phenotypic yeast growth analysis for chronic toxicity testing. Ecotoxicol. Environ. Safety 2004, 59, 142–150. [Google Scholar]
- Walmsley, R.M.; Billinton, N.; Heyer, W.-D. Green fluorescent protein as a reporter for the DNA damage-induced gene RAD54 in Saccharomyces cerevisiae. Yeast 1997, 13, 535–545. [Google Scholar]
- Peltola, P.; Ivask, A.; Åström, M.; Virta, M. Lead and Cu in contaminated urban soils: extraction with chemical reagents and bioluminescent bacteria and yeast. Sci. Total Environ. 2005, 350, 194–203. [Google Scholar]
- Hollis, R.P.; Killham, K.; Glover, L.A. Design and application of a biosensor for monitoring toxicity of compounds to eukaryotes. Appl. Environ. Microbiol. 2000, 66, 1676–1679. [Google Scholar]
- Leskinen, P.; Michelini, E.; Picard, D.; Karp, M.; Virta, M. Bioluminescence yeast assays for detecting estrogenic and androgenic activity in different matrices. Chemosphere 2005, 61, 259–266. [Google Scholar]
- Ike, M.; Chen, M.-Y.; Jin, C.-S.; Fujita, M. Acute toxicity, mutagenicity, and estrogenicity of biodegradation products of bisphenol-A. Environ. Toxicol. 2002, 17, 457–461. [Google Scholar]
- Routledge, E.J.; Sumpter, J.P. Estrogenic activity of surfactants and some of their degradation products assessed using a recombinant yeast screen. Environ. Toxicol. Chem. 1996, 15, 241–248. [Google Scholar]
- De Boever, P.; Demaré, W.; Vanderperren, E.; Cooreman, K.; Bossier, P.; Verstraete, W. Optimization of a yeast estrogen screen and its applicability to study the release of estrogenic isoflavones from a soygerm powder. Environ. Health Pers. 2001, 109, 691–697. [Google Scholar]
- Colemana, H.M.; Routledge, E.J.; Sumpter, J.P.; Eggins, B.R.; Byrne, J.A. Rapid loss of estrogenicity of steroid estrogens by UVA photolysis and photocatalysis over an immobilized titanium dioxide catalyst. Water Res. 2004, 38, 3233–3240. [Google Scholar]
- Michelini, E.; Leskinen, P.; Virta, M.; Karp, M.; Mora, A. A new recombinant cell-based bioluminescent assay for sensitive androgen-like compound detection. Biosens. Bioelectron. 2005, 20, 2261–2267. [Google Scholar]
- Rodriguez-Peña, J.M.; Diez-Muñiz, S.; Nombela, C.; Arroyo, J. A yeast strain biosensor to detect cell wall-perturbing agents. J. Biotechnol. 2007, 133, 311–317. [Google Scholar]
- Gould, S.J.; Keller, G.-A.; Subramani, S. Identification of peroxisomal targeting signals located at the carboxy terminus of four peroxisomal proteins. J. Cell Biol. 1988, 897–905. [Google Scholar]
- Leskinen, P.; Virta, M.; Karp, M. One-step measurement of firefly luciferase activity in yeast. Yeast 2003, 20, 1109–1113. [Google Scholar]
- Novi, S.; Pretti, C.; Cognetti, A.M.; Longo, V.; Marchetti, S.; Gervasi, G.G. Biotransformation enzymes and their induction by β-naphtoflavone in adult sea bass (Dicentrarchus labrax). Aquat. Toxicol. 1998, 41, 63–81. [Google Scholar]
- Crespo, J.L.; Hall, M.N. Elucidating TOR signaling and rapamycin action: lessons from Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rew. 2002, 579–591. [Google Scholar]
- Venables, B.; Russell, A.D. Nystatin-induced changes in Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 1975, 7, 121–127. [Google Scholar]
- Bhuiyan, M.S.A.; Ito, Y.; Nakamura, A.; Tanaka, N.; Fujita, K.; Fukui, H.; Takegawa, K. Nystatin effects on vacuolar function in Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 1999, 96, 1075–1082. [Google Scholar]
- Siro, M.-R.; Romar, H.; Lövgren, T. Continous flow method for extraction and bioluminescence assay of ATP on baker's yeast. Eur. J. Appl. Microbiol. Biotechnol. 1982, 15, 258–264. [Google Scholar]
- Davies, J; Spiegelman, G.B; Yim, G. The world of subinhibitory antibiotic concentrations. Curr. Opin. Microbiol. 2006, 9, 445–453. [Google Scholar]
- Yim, G.; de la Cruz, F.; Spiegelman, G.B.; Davies, J. Transcription modulation of Salmonella enterica serovar Typhimurium promoters by sub-MIC levels of rifampin. J. Bacteriol. 2006, 188, 7988–7991. [Google Scholar]
- Galluzzi, L.; Karp, M. Intracellular redox equilibrium and growth phase affect the performance of luciferase-based biosensors. J. Biotechnol. 2007, 127, 188–198. [Google Scholar]
- Sirisattha, S.; Momose, Y.; Kitagawa, E.; Iwahashi, H. Toxicity of anionic detergents determined by Saccharomyces cerevisiae microarray analysis. Water Res. 2004, 38, 61–70. [Google Scholar]
- do Valle Matta, M.A.; Jonniaux, J.-L.; Balzi, E.; Goffeau, A.; van den Hazel, B. Novel target genes of the yeast regulator Pdr1p: a contribution of the TPO1 gene in resistance to quinidine and other drugs. Gene 2001, 272, 111–119. [Google Scholar]
- Mamnum, M.M.; Schüller, C.; Kuchler, K. Expression regulation of the yeast PDR5 ATP-binding cassette (ABC) transporter suggests a role in cellular detoxification during the exponential growth phase. FEBS Lett. 2004, 559, 111–117. [Google Scholar]
- Bauer, B.E.; Wolfger, H.; Kuchler, K. Inventory and function of yeast ABC proteins: about sex, stress, pleiotropic drug and heavy metal resistance. Biochim. Biophys Acta. 1999, 1461, 217–236. [Google Scholar]
- Prévéral, S.; Ansoborlo, E.; Mari, S.; Vavasseur, A.; Forestier, C. Metal(loid)s and radionuclides cytotoxicity in Saccharomyces verevisiae. Role of YCF1, gluthatione and effect of buthionine sulfoximine. Biochimie 2006, 88, 1651–1663. [Google Scholar]
- Lucau-Danila, A.; Lelandais, G.; Kozovska, Z.; Tanty, V.; Delaveau, T.; Devaux, F.; Jacq, C. Early expression of yeast genes affected by chemical stress. Mol. Cell. Biol. 2005, 124, 1860–1868. [Google Scholar]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar]
- Ghosh, M.; Shen, J.; Rosen, B.P. Pathways of As(III) detoxification in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. 1999, 96, 5001–5006. [Google Scholar]
- Beggs, W.H. Protonation of ketoconazole in relation to fungistatic activity. Mycopathology 1991, 116, 3–4. [Google Scholar]
- Minigawa, H.; Kitaura, K.; Nakamizo, N. Effects of pH on the activity of ketoconazole against Candida albicans. Antimicrob. Agents Chemother. 1983, 23, 105–107. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Bioluminescence assay IC50 (μmol/L) | Agar diffusion assay | |
|---|---|---|---|
| Exposure time 2.5 h | Exposure time 5 h | Minimum inhibitory concentration (MIC) (μmol/L) | |
| Antimicrobial agents | |||
| 5,6-benzoflavone | 0.008 | 0.006 | 0.075 |
| Cycloheximide | NTa | 0.7 | 42.7 |
| Ketoconazole | NT | NT | 1.0 |
| Nystatin | 1.6 | 1.4 | 54.0 |
| Rapamycin | 2.8 | 0.03 | 0.2 |
| Sodium azide | 25.8 | 27.4 | - b |
| Sodium dodecyl sulfate | 133.5*103 | 69.5*103 | 1.8*103 |
| Metals | |||
| Arsenic(V)oxide | 2.4*103 | 0.4*103 | 85.5*103 |
| Sodium-m-arsenite | 25.5*103 | 0.5*103 | 14.5*103 |
| Cadmium(II)chloride | 19.3*103 | 2.0*103 | 124.6*103 |
| Copper(I)sulfate | 5.1*103 | ND | 9.8*103 |
| Lead(II)acetate | 71.9*103 | 18.4*103 | - c |
| Magnesium(II)chloride | NT | NAd | 2060.5*103 |
| Zinc(II)chloride | NT | NA | 10265.6*103 |
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Välimaa, A.-L.; Kivistö, A.; Virta, M.; Karp, M. Real-time Monitoring of Non-specific Toxicity Using a Saccharomyces cerevisiae Reporter System. Sensors 2008, 8, 6433-6447. https://doi.org/10.3390/s8106433
Välimaa A-L, Kivistö A, Virta M, Karp M. Real-time Monitoring of Non-specific Toxicity Using a Saccharomyces cerevisiae Reporter System. Sensors. 2008; 8(10):6433-6447. https://doi.org/10.3390/s8106433
Chicago/Turabian StyleVälimaa, Anna-Liisa, Anniina Kivistö, Marko Virta, and Matti Karp. 2008. "Real-time Monitoring of Non-specific Toxicity Using a Saccharomyces cerevisiae Reporter System" Sensors 8, no. 10: 6433-6447. https://doi.org/10.3390/s8106433